Differentiation of Motor Neuron-Like Cells from Tonsil-Derived Mesenchymal Stem Cells and Their Possible Application to Neuromuscular Junction Formation



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
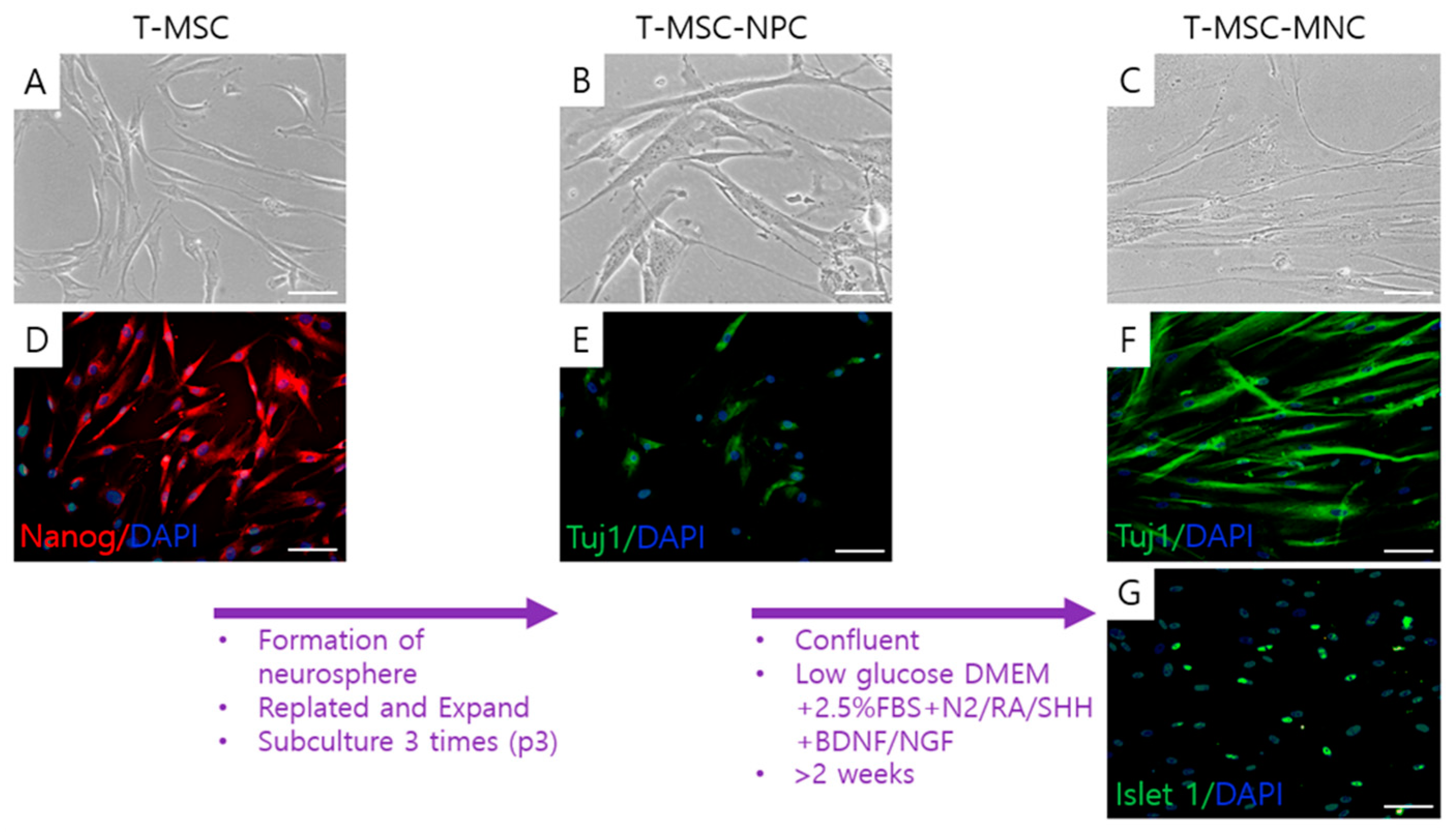
2.1. Differentiation of MN-Like Cells from T-MSCs
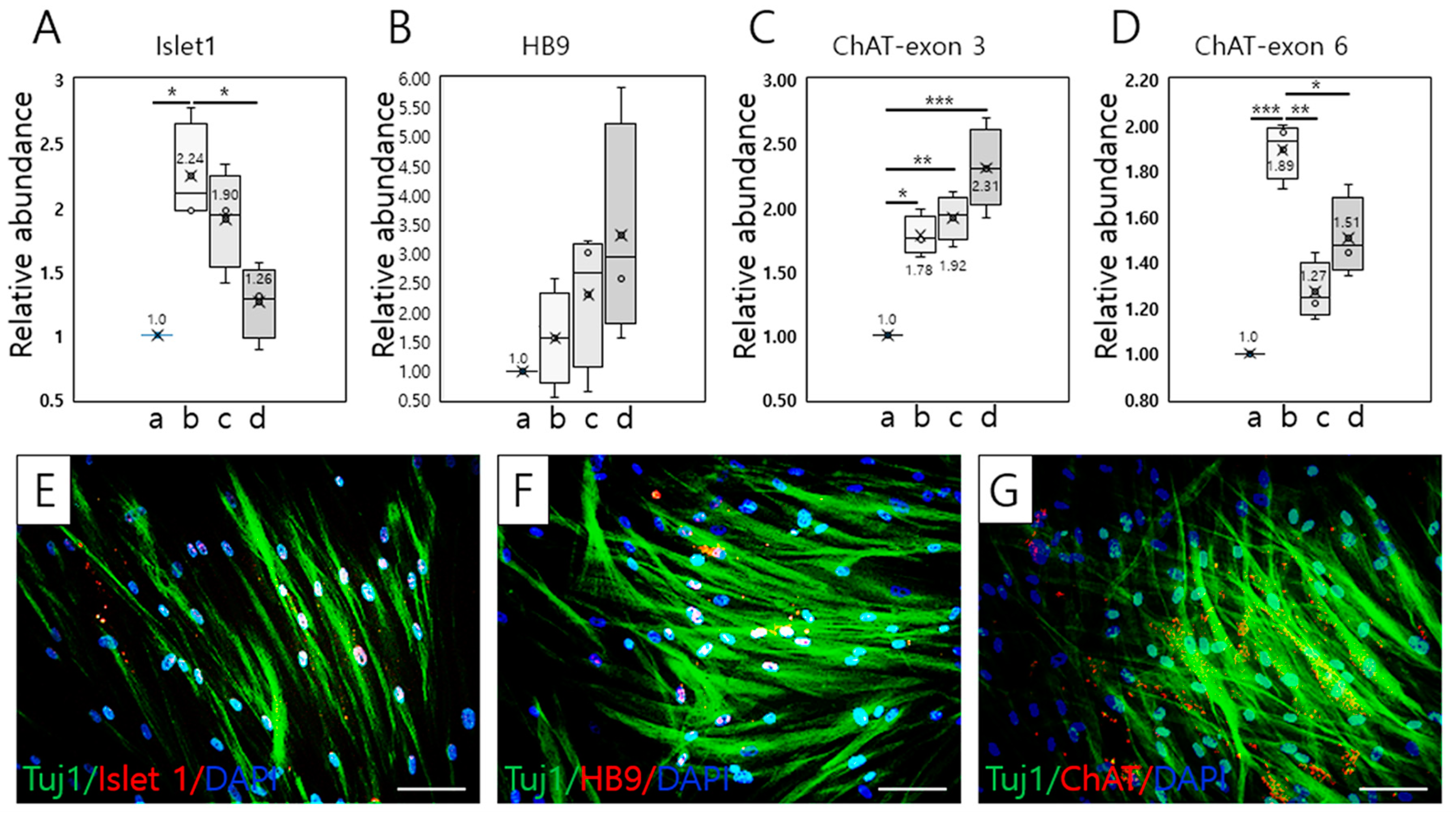
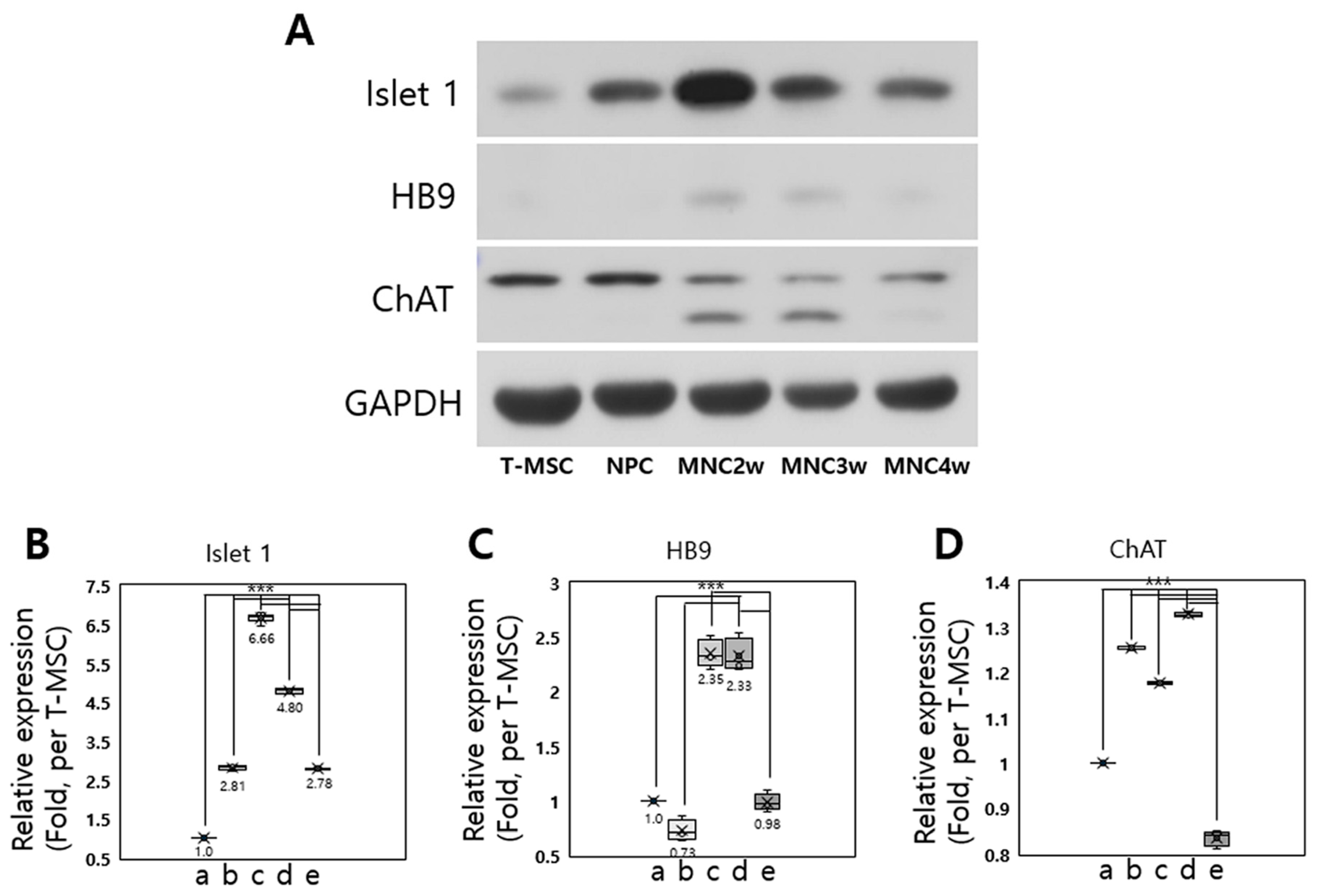
2.2. Detection of Motor Neuronal Markers in T-MSC-MNCs
2.2.1. RT–qPCR
2.2.2. Immunocytochemistry
2.2.3. Western Blot Analysis
2.3. Effective MN Differentiation from T-MSCs
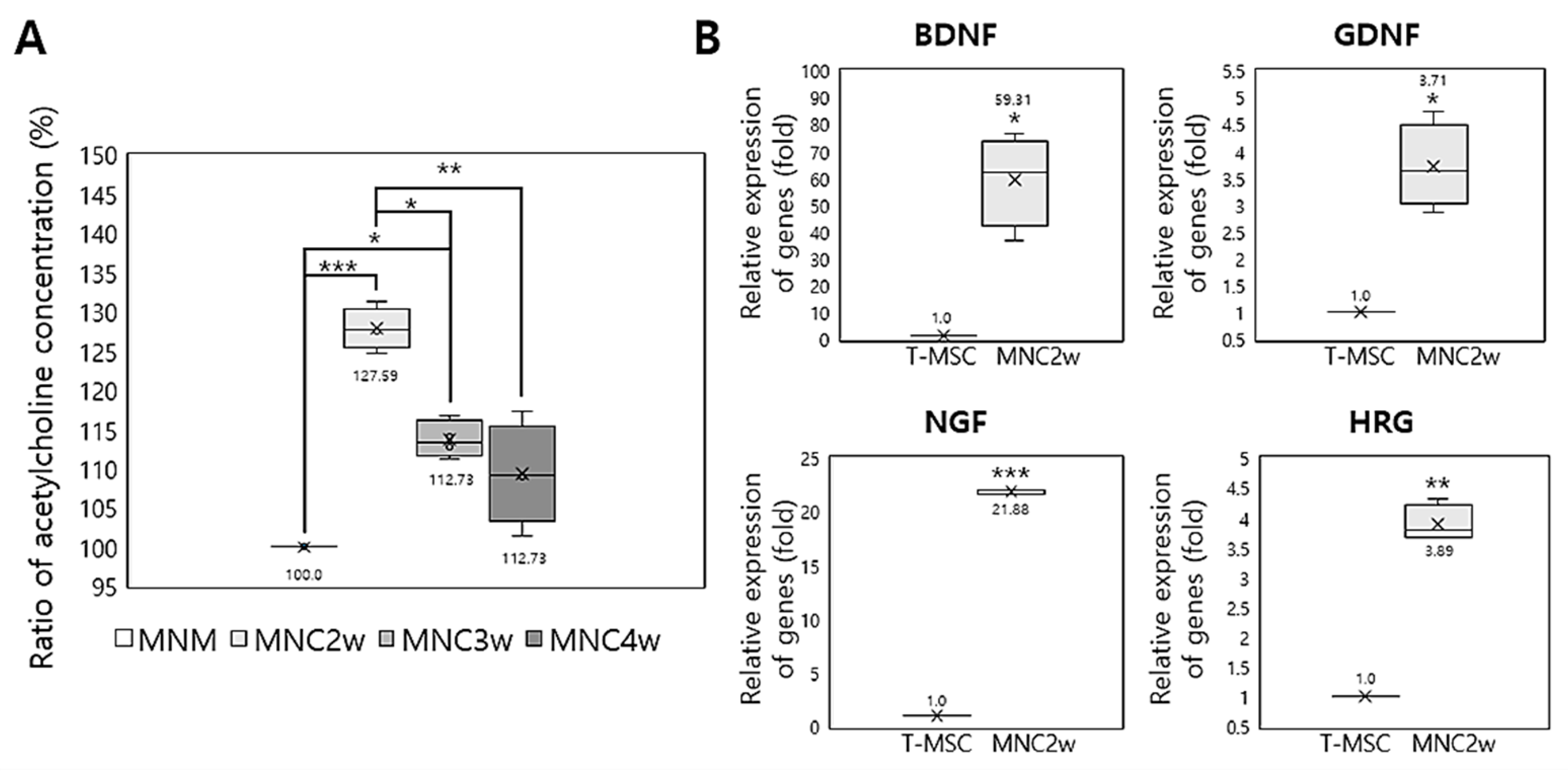
2.4. Expression of Neurotrophic Factors by T-MSC-MNCs
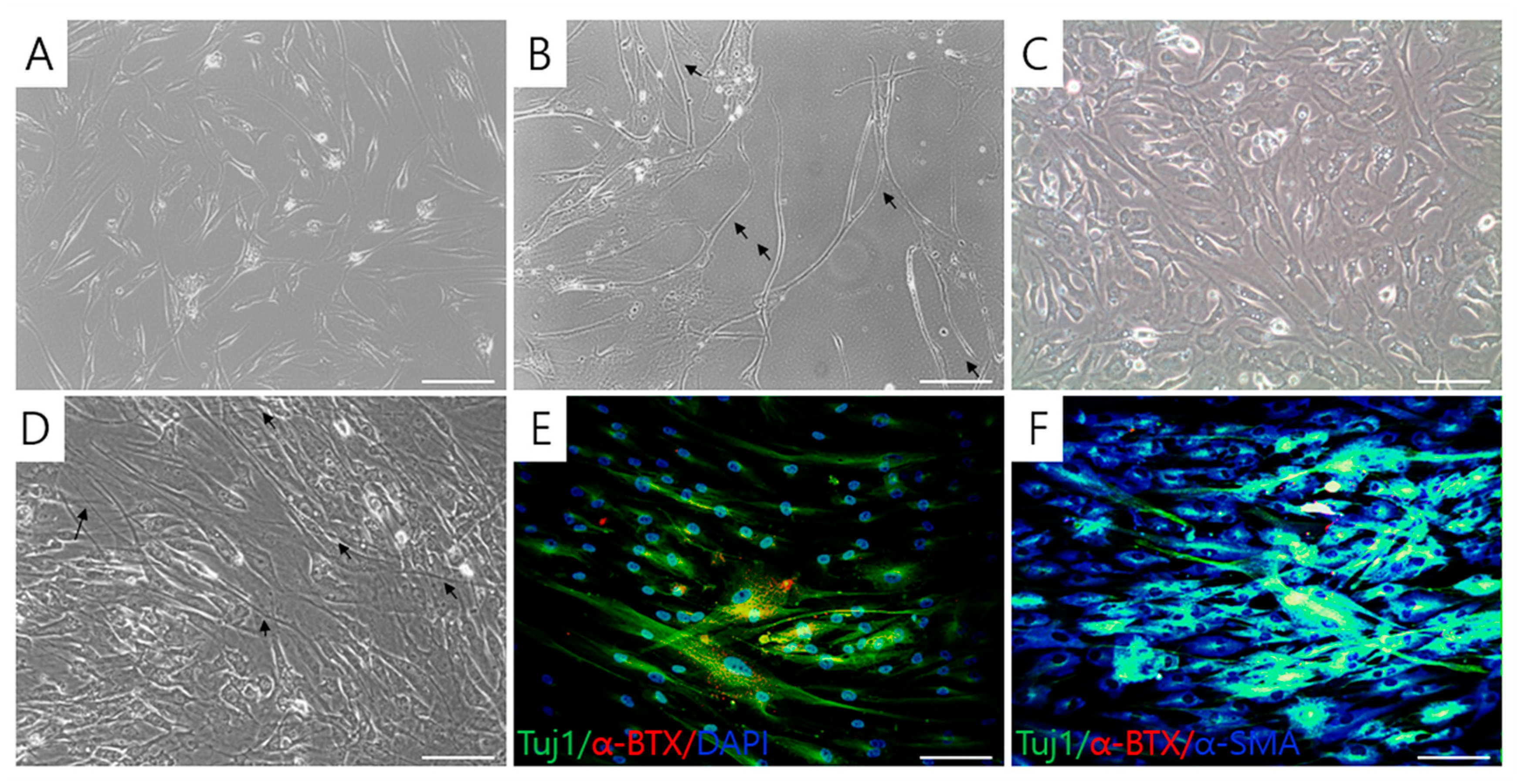
2.5. Formation of Acetylcholine Receptor Clusters in Cocultures of T-MSC-MNCs with Human Skeletal Muscle Cells
3. Discussion
4. Material and Methods
4.1. Ethics Statement
4.2. Preparation of T-MSCs and their Differentiation into MN-like Cells
4.3. RT–qPCR
4.4. Western Blotting
4.5. Immunocytochemistry
4.6. Enzyme-Driven Reaction Assay for Acetylcholine
4.7. Coculture of Human Skeletal Muscle Cells with T-MSC-MNCs
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| T-MSC | tonsil-derived mesenchymal stem cell |
| MN | motor neuron |
| MND | motor neuron diseases |
| ALS | amyotrophic lateral sclerosis |
| ESC | embryonic stem cell |
| iPSC | induced pluripotent stem cell |
| NPC | neural progenitor cell |
| PEI | polyethyleneimine |
| PNS | peripheral nervous system |
| CNS | central nervous system |
| HB9 | HB9/HLXB9 |
| ChAT | choline acetyltransferase |
| pChAT | Peripheral type ChAT |
| MNM | motor neuronal induction medium |
| BDNF | brain-derived neurotrophic factor |
| GDNF | glial cell-derived neurotrophic factor |
| NGF | nerve growth factor |
| HRG | heregulin |
| hSKMC | human skeletal muscle cells |
| AchR | acetylcholine receptors |
| α-BTX | α-bungarotoxin |
| NMJs | neuromuscular junctions |
| RA | retinoic acid |
| Shh | sonic hedgehog |
| cChAT | common type of ChAT |
| GFRα | GDNF family receptor α |
| GAPDH | glyceraldehyde 3-phosphate dehydrogenase |
| FBS | fetal bovine serum |
| PBS | phosphate-buffered solution |
| CM | conditioned media |
| α-SMA | α-smooth muscle actin |
| SEM | standard error of the mean |
| ANOVA | analysis of variance |
References
- Djouad, F.; Jackson, W.M.; Bobick, B.E.; Janjanin, S.; Song, Y.; Huang, G.T.; Tuan, R.S. Activin A expression regulates multipotency of mesenchymal progenitor cells. Stem Cell Res. Ther. 2010, 1, 11. [Google Scholar] [CrossRef] [PubMed]
- Ryu, K.H.; Cho, K.A.; Park, H.S.; Kim, J.Y.; Woo, S.Y.; Jo, I.; Choi, Y.H.; Park, Y.M.; Jung, S.C.; Chung, S.M.; et al. Tonsil-derived mesenchymal stromal cells: Evaluation of biologic, immunologic and genetic factors for successful banking. Cytotherapy 2012, 14, 1193–1202. [Google Scholar] [CrossRef] [PubMed]
- Park, M.; Kim, Y.H.; Ryu, J.H.; Woo, S.Y.; Ryu, K.H. Immune Suppressive Effects of Tonsil-Derived Mesenchymal Stem Cells on Mouse Bone-Marrow-Derived Dendritic Cells. Stem Cells Int. 2015, 2015, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.; Park, Y.S.; Kim, H.S.; Kim, H.Y.; Jin, Y.M.; Jung, S.C.; Ryu, K.H.; Jo, I. Characterization of long-term in vitro culture-related alterations of human tonsil-derived mesenchymal stem cells: Role for CCN1 in replicative senescence-associated increase in osteogenic differentiation. J. Anat. 2014, 225, 510–518. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.S.; Lee, B.J.; Park, H.Y.; Song, J.S.; Shin, S.C.; Lee, J.C.; Wang, S.G.; Jung, J.S. Effects of Donor Age, Long-Term Passage Culture, and Cryopreservation on Tonsil-Derived Mesenchymal Stem Cells. Cell. Physiol. Biochem. 2015, 36, 85–99. [Google Scholar] [CrossRef] [PubMed]
- Moon, H.J.; Patel, M.; Chung, H.; Jeong, B. Nanocomposite versus Mesocomposite for Osteogenic Differentiation of Tonsil-Derived Mesenchymal Stem Cells. Adv. Healthc. Mater. 2016, 5, 353–363. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Choi, Y.; Jung, N.; Yu, Y.; Ryu, K.H.; Kim, H.S.; Jo, I.; Choi, B.O.; Jung, S.C. Myogenic differentiation potential of human tonsil-derived mesenchymal stem cells and their potential for use to promote skeletal muscle regeneration. Int. J. Mol. Med. 2016, 37, 1209–1220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.J.; Park, M.H.; Moon, H.J.; Park, J.H.; Ko, D.Y.; Jeong, B. Polypeptide Thermogels as a Three Dimensional Culture Scaffold for Hepatogenic Differentiation of Human Tonsil-Derived Mesenchymal Stem Cells. ACS Appl. Mater. Interfaces 2014, 6, 17034–17043. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.Y.; Kim, Y.R.; Park, W.J.; Kim, H.S.; Jung, S.C.; Woo, S.Y.; Jo, I.; Ryu, K.H.; Park, J.W. Characterisation of insulin-producing cells differentiated from tonsil derived mesenchymal stem cells. Differentiation 2015, 90, 27–39. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.S.; Hwang, J.Y.; Jun, Y.; Jin, Y.M.; Kim, G.; Kim, H.Y.; Kim, H.S.; Lee, S.H.; Jo, I. Scaffold-free parathyroid tissue engineering using tonsil-derived mesenchymal stem cells. Acta Biomater. 2016, 35, 215–227. [Google Scholar] [CrossRef]
- Jung, N.; Park, S.; Choi, Y.; Park, J.W.; Hong, Y.; Park, H.; Yu, Y.; Kwak, G.; Kim, H.; Ryu, K.H.; et al. Tonsil-Derived Mesenchymal Stem Cells Differentiate into a Schwann Cell Phenotype and Promote Peripheral Nerve Regeneration. Int. J. Mol. Sci. 2016, 17, 1867. [Google Scholar] [CrossRef]
- Patel, M.; Moon, H.J.; Jung, B.K.; Jeong, B. Microsphere-Incorporated Hybrid Thermogel for Neuronal Differentiation of Tonsil Derived Mesenchymal Stem Cells. Adv. Healthc. Mater. 2015, 4, 1565–1574. [Google Scholar] [CrossRef] [PubMed]
- Tortora, G.; Derrickson, B. Principles of Anatomy and Physiology, 14th ed.; John Wiley Sons Inc.: Hoboken, NJ, USA, 2014; ISBN 978-1-118-34500-9. [Google Scholar]
- Mitne-Neto, M.; Machado-Costa, M.; Marchetto, M.C.N.; Bengtson, M.H.; Joazeiro, C.A.; Tsuda, H.; Bellen, H.J.; Silva, H.C.A.; Oliveira, A.S.B.; Lazar, M.; et al. Downregulation of VAPB expression in motor neurons derived from induced pluripotent stem cells of ALS8 patients. Hum. Mol. Genet. 2011, 20, 3642–3652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Statland, J.M.; Barohn, R.J.; McVey, A.L.; Katz, J.S.; Dimachkie, M.M. Patterns of Weakness, Classification of Motor Neuron Disease, and Clinical Diagnosis of Sporadic Amyotrophic Lateral Sclerosis. Neurol. Clin. 2015, 33, 735–748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qu, Q.; Li, D.; Louis, K.R.; Li, X.; Yang, H.; Sun, Q.; Crandall, S.R.; Tsang, S.; Zhou, J.; Cox, C.L.; et al. High-efficiency motor neuron differentiation from human pluripotent stem cells and the function of Islet-1. Nat. Commun. 2014, 5, 3449. [Google Scholar] [CrossRef] [PubMed]
- Shimojo, D.; Onodera, K.; Doi-Torii, Y.; Ishihara, Y.; Hattori, C.; Miwa, Y.; Tanaka, S.; Okada, R.; Ohyama, M.; Shoji, M.; et al. Rapid, efficient, and simple motor neuron differentiation from human pluripotent stem cells. Mol. Brain 2015, 8, 79. [Google Scholar] [CrossRef]
- Sances, S.; Bruijn, L.I.; Chandran, S.; Eggan, K.; Ho, R.; Klim, J.R.; Livesey, M.R.; Lowry, E.; Macklis, J.D.; Rushton, D.; et al. Modeling ALS with motor neurons derived from human induced pluripotent stem cells. Nat. Neurosci. 2016, 19, 542–553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bossolasco, P.; Sassone, F.; Gumina, V.; Peverelli, S.; Garzo, M.; Silani, V. Motor neuron differentiation of iPSCs obtained from peripheral blood of a mutant TARDBP ALS patient. Stem Cell Res. 2018, 30, 61–68. [Google Scholar] [CrossRef]
- Liu, X.; Li, D.; Jiang, D.; Fang, Y. Acetylcholine secretion by motor neuron-like cells from umbilical cord mesenchymal stem cells. Neural Regen. Res. 2013, 8, 2086. [Google Scholar]
- Abdullah, R.H.; Yaseen, N.Y.; Salih, S.M.; Al-Juboory, A.A.; Hassan, A.; Al-Shammari, A.M. Induction of mice adult bone marrow mesenchymal stem cells into functional motor neuron-like cells. J. Chem. Neuroanat. 2016, 77, 129–142. [Google Scholar] [CrossRef]
- Bahrami, N.; Bayat, M.; Mohamadnia, A.; Khakbiz, M.; Yazdankhah, M.; Ai, J.; Ebrahimi-Barough, S. Purmorphamine as a Shh Signaling Activator Small Molecule Promotes Motor Neuron Differentiation of Mesenchymal Stem Cells Cultured on Nanofibrous PCL Scaffold. Mol. Neurobiol. 2017, 54, 5668–5675. [Google Scholar] [CrossRef] [PubMed]
- Jang, S.; Kang, Y.H.; Ullah, I.; Shivakumar, S.; Rho, G.J.; Cho, Y.C.; Sung, I.Y.; Park, B.W. Cholinergic Nerve Differentiation of Mesenchymal Stem Cells Derived from Long-Term Cryopreserved Human Dental Pulp In Vitro and Analysis of Their Motor Nerve Regeneration Potential In Vivo. Int. J. Mol. Sci. 2018, 19, 2434. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Koh, S.E.; Maeng, S.; Lee, W.D.; Lim, J.; Lee, Y.J. Neural progenitors generated from the mesenchymal stem cells of first-trimester human placenta matured in the hypoxic-ischemic rat brain and mediated restoration of locomotor activity. Placenta 2011, 32, 269–276. [Google Scholar] [CrossRef] [PubMed]
- Huang, E.J.; Reichardt, L.F. Neurotrophins: Roles in Neuronal Development and Function. Annu. Rev. Neurosci. 2001, 24, 677–736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bianchi, F.; Malboubi, M.; Li, Y.; George, J.H.; Jerusalem, A.; Szele, F.; Thompson, M.S.; Ye, H. Rapid and efficient differentiation of functional motor neurons from human iPSC for neural injury modelling. Stem Cell Res. 2018, 32, 126–134. [Google Scholar] [CrossRef] [PubMed]
- Faravelli, I.; Bucchia, M.; Rinchetti, P.; Nizzardo, M.; Simone, C.; Frattini, E.; Corti, S. Motor neuron derivation from human embryonic and induced pluripotent stem cells: Experimental approaches and clinical perspectives. Stem Cell Res. Ther. 2014, 5, 87. [Google Scholar] [CrossRef] [PubMed]
- Strauss, L.; Kemper, R. Human Choline Acetyltransferase Gene Maps to Region 1 Oql I-q22.2 by in Situ Hybridization. Genomics 1991, 2, 396–398. [Google Scholar] [CrossRef]
- Tooyama, I.; Kimura, H. A protein encoded by an alternative splice variant of choline acetyltransferase mRNA is localized preferentially in peripheral nerve cells and fibers. J. Chem. Neuroanat. 2000, 17, 217–226. [Google Scholar] [CrossRef]
- Chiocchetti, R.; Poole, D.; Kimura, H.; Aimi, Y.; Robbins, H.; Castelucci, P.; Furness, J. Evidence that two forms of choline acetyltransferase are differentially expressed in subclasses of enteric neurons. Cell Tissue Res. 2003, 311, 11–22. [Google Scholar] [CrossRef]
- Levitan, I.B.; Kaczmarek, L.K. Intercellular Communication. In The Neuron. Cell and Molecular Biology, 4th ed.; Oxford University Press: Oxford, UK, 2015. [Google Scholar]
- Santhanam, N.; Kumanchik, L.; Guo, X.; Sommerhage, F.; Cai, Y.; Jackson, M.; Martin, C.; Saad, G.; McAleer, C.W.; Wang, Y.; et al. Stem cell derived phenotypic human neuromuscular junction model for dose response evaluation of therapeutics. Biomaterials 2018, 166, 64–78. [Google Scholar] [CrossRef]
- Ferraro, E.; Molinari, F.; Berghella, L. Molecular control of neuromuscular junction development. J. Cachexia Sarcopenia Muscle 2012, 3, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Demestre, M.; Orth, M.; Föhr, K.J.; Achberger, K.; Ludolph, A.C.; Liebau, S.; Boeckers, T.M. Formation and characterisation of neuromuscular junctions between hiPSC derived motoneurons and myotubes. Stem Cell Res. 2015, 15, 328–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dharmadasa, T.; Henderson, R.D.; Talman, P.S.; Macdonell, R.A.; Mathers, S.; Schultz, D.W.; Needham, M.; Zoing, M.; Vucic, S.; Kiernan, M.C. Motor neurone disease: Progress and challenges. Med. J. Aust. 2017, 206, 357–362. [Google Scholar] [CrossRef] [PubMed]
- Czarzasta, J.; Habich, A.; Siwek, T.; Czapliński, A.; Maksymowicz, W.; Wojtkiewicz, J. Stem cells for ALS: An overview of possible therapeutic approaches. Int. J. Dev. Neurosci. 2017, 57, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Keefe, K.; Sheikh, I.; Smith, G. Targeting Neurotrophins to Specific Populations of Neurons: NGF, BDNF, and NT-3 and Their Relevance for Treatment of Spinal Cord Injury. Int. J. Mol. Sci. 2017, 18, 548. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, M.; McHugh, J.; Tork, C.; Shelley, B.; Klein, S.M.; Aebischer, P.; Svendsen, C.N. GDNF Secreting Human Neural Progenitor Cells Protect Dying Motor Neurons, but Not Their Projection to Muscle, in a Rat Model of Familial ALS. PLoS ONE 2007, 2, e689. [Google Scholar] [CrossRef]
- Ho, W.H.; Armanini, M.P.; Nuijens, A.; Phillips, H.S.; Osheroff, P.L. Sensory and Motor Neuron-derived Factor. A novel heregulin variant highly expressed in sensory and motor neurons. J. Biol. Chem. 1995, 270, 14523–14532. [Google Scholar] [CrossRef]
- Mei, L.; Xiong, W.C. Neuregulin 1 in neural development, synaptic plasticity and schizophrenia. Nat. Rev. Neurosci. 2008, 9, 437–452. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model forrelative quantification in real-time RT–PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Zhang, L.; Tan, X.; Dong, C.; Zou, L.; Zhao, H.; Zhang, X.; Tian, M.; Jin, G. In vitro differentiation of human umbilical cord mesenchymal stem cells (hUCMSCs), derived from Wharton’s jelly, into choline acetyltransferase (ChAT)-positive cells. Int. J. Dev. Neurosci. 2012, 30, 471–477. [Google Scholar] [CrossRef]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, S.; Kim, J.Y.; Myung, S.; Jung, N.; Choi, Y.; Jung, S.-C. Differentiation of Motor Neuron-Like Cells from Tonsil-Derived Mesenchymal Stem Cells and Their Possible Application to Neuromuscular Junction Formation. Int. J. Mol. Sci. 2019, 20, 2702. https://doi.org/10.3390/ijms20112702
Park S, Kim JY, Myung S, Jung N, Choi Y, Jung S-C. Differentiation of Motor Neuron-Like Cells from Tonsil-Derived Mesenchymal Stem Cells and Their Possible Application to Neuromuscular Junction Formation. International Journal of Molecular Sciences. 2019; 20(11):2702. https://doi.org/10.3390/ijms20112702
Chicago/Turabian StylePark, Saeyoung, Ji Yeon Kim, Seoha Myung, Namhee Jung, Yeonzi Choi, and Sung-Chul Jung. 2019. "Differentiation of Motor Neuron-Like Cells from Tonsil-Derived Mesenchymal Stem Cells and Their Possible Application to Neuromuscular Junction Formation" International Journal of Molecular Sciences 20, no. 11: 2702. https://doi.org/10.3390/ijms20112702