Genome-Wide Characterization, Evolution, and Expression Profiling of VQ Gene Family in Response to Phytohormone Treatments and Abiotic Stress in Eucalyptus grandis

Abstract
:1. Introduction
2. Results
2.1. Identification of the VQ Gene Family in the E. grandis
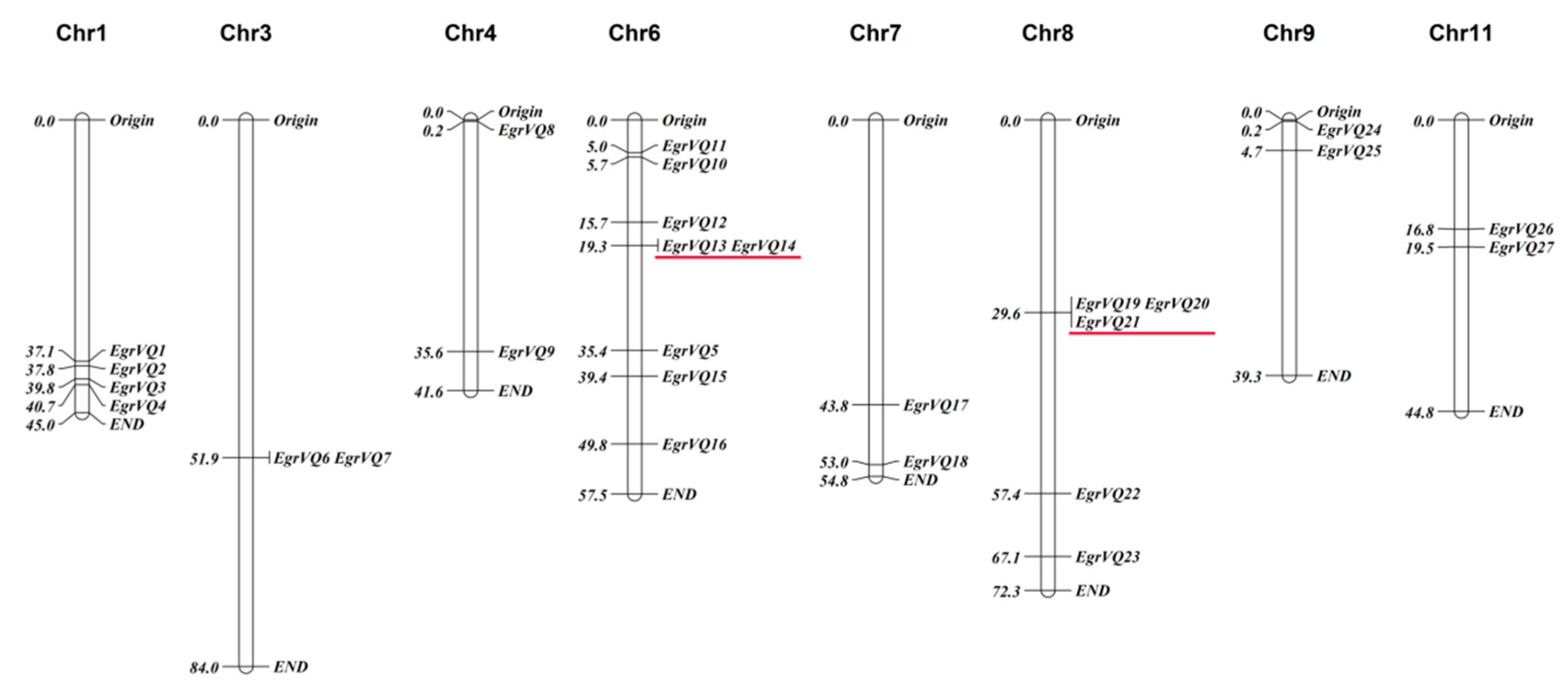
2.2. Mapping EgrVQ Genes on Chromosomes, Gene Duplication, and Analysis of Paralogs and Orthologs
2.3. Identification of Cis-Elements in the Promoter Regions of EgrVQ Genes
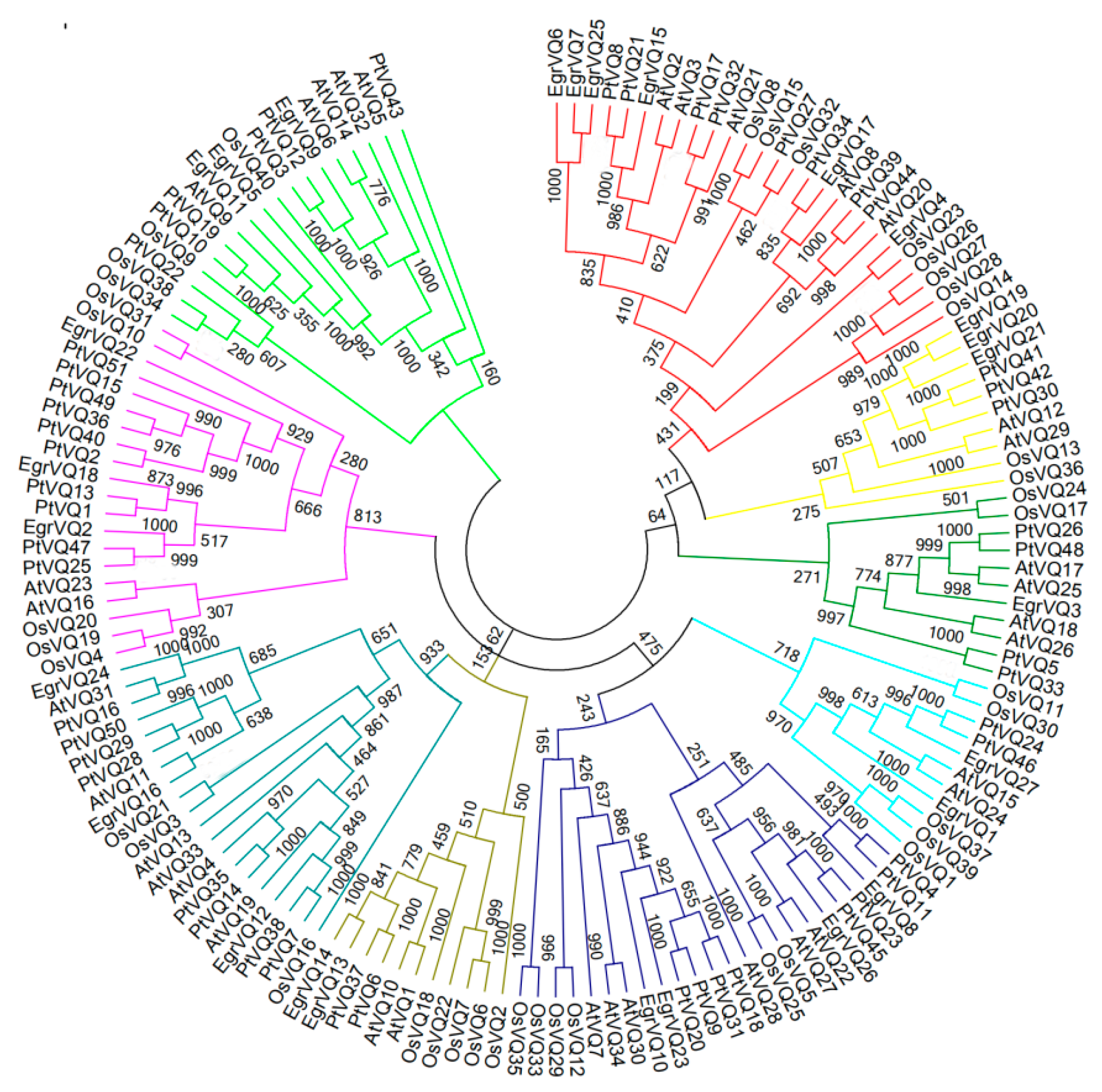
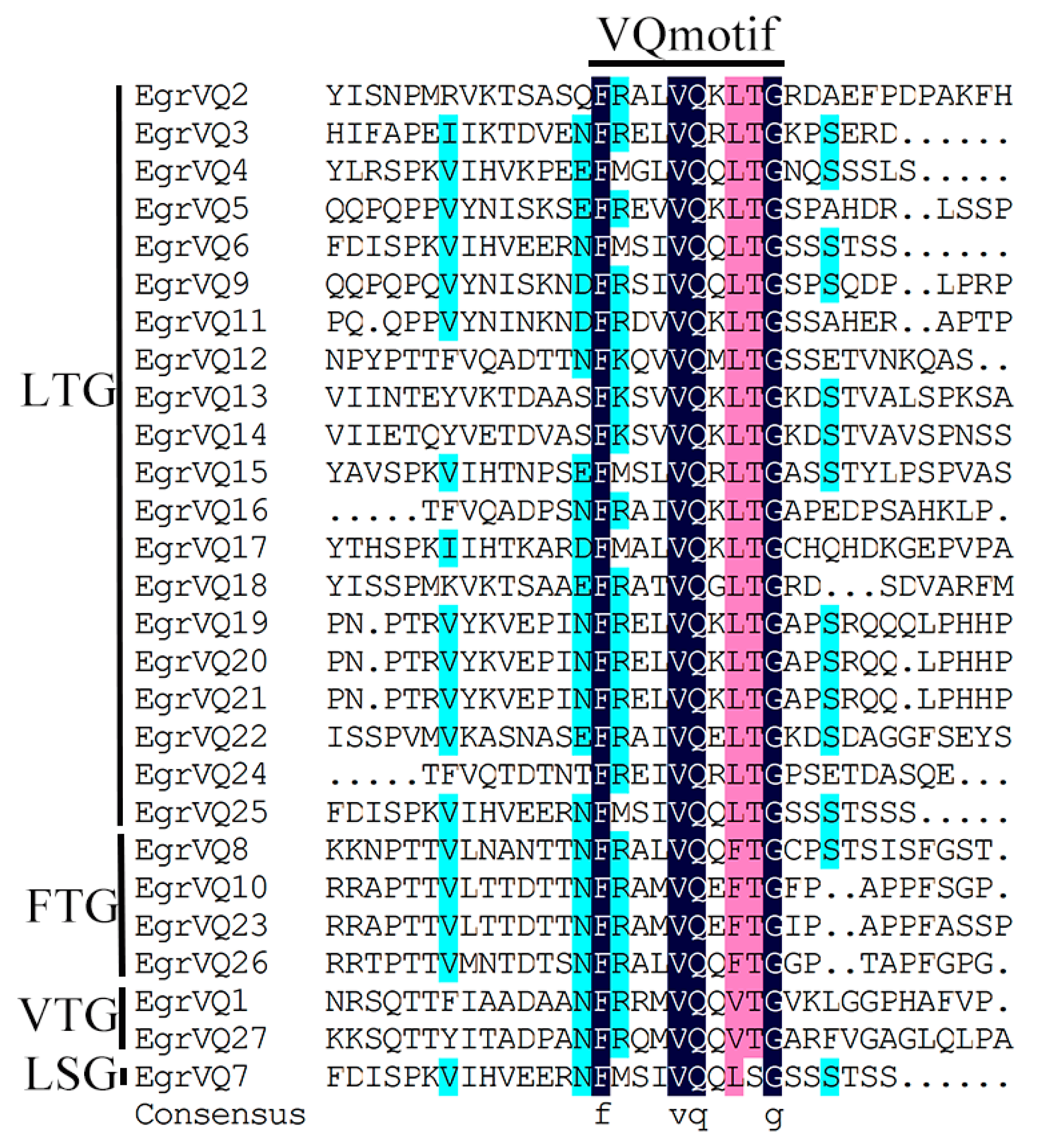
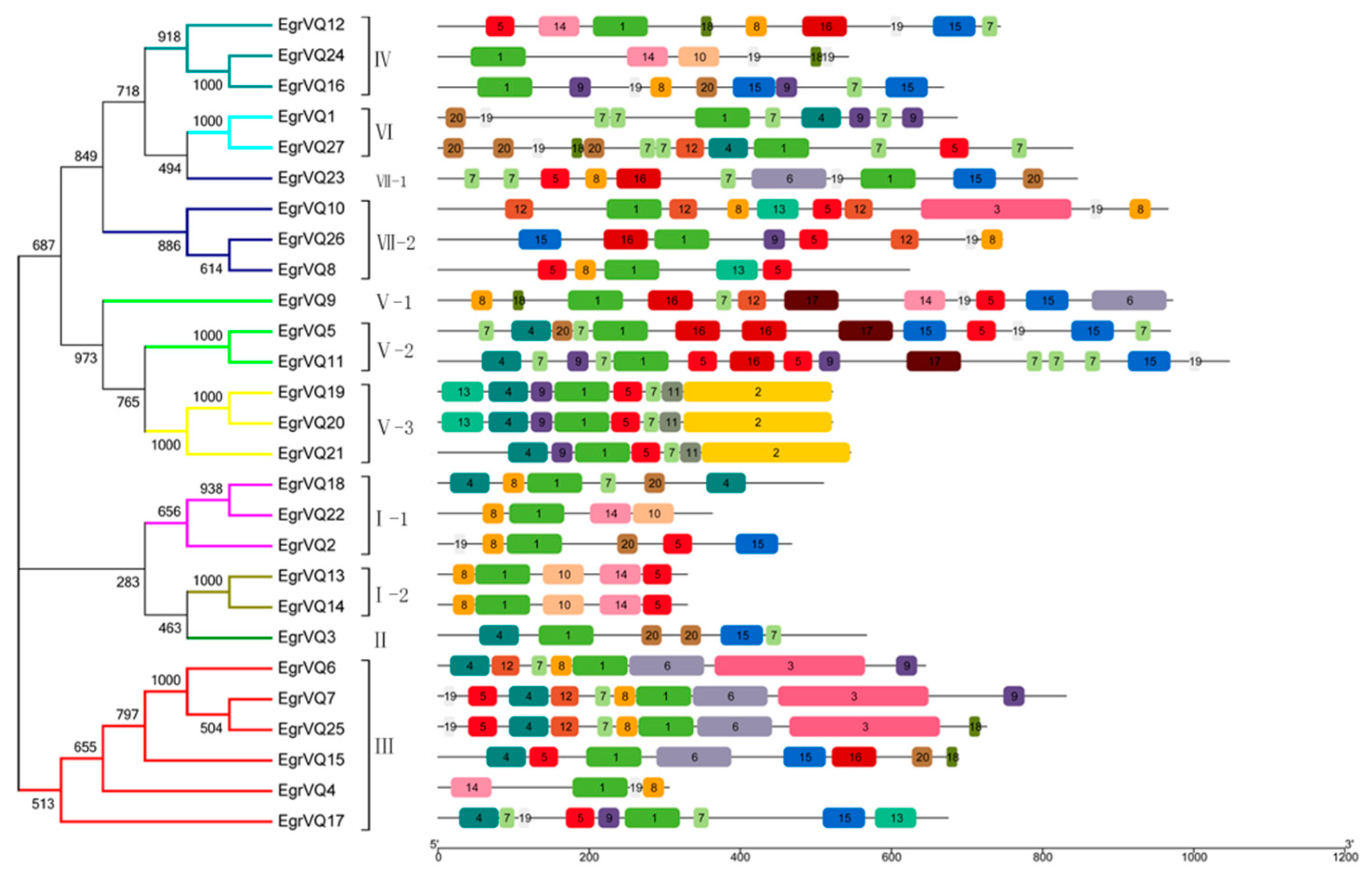
2.4. Multiple Sequence Alignment and Phylogenetic of EgrVQ Proteins
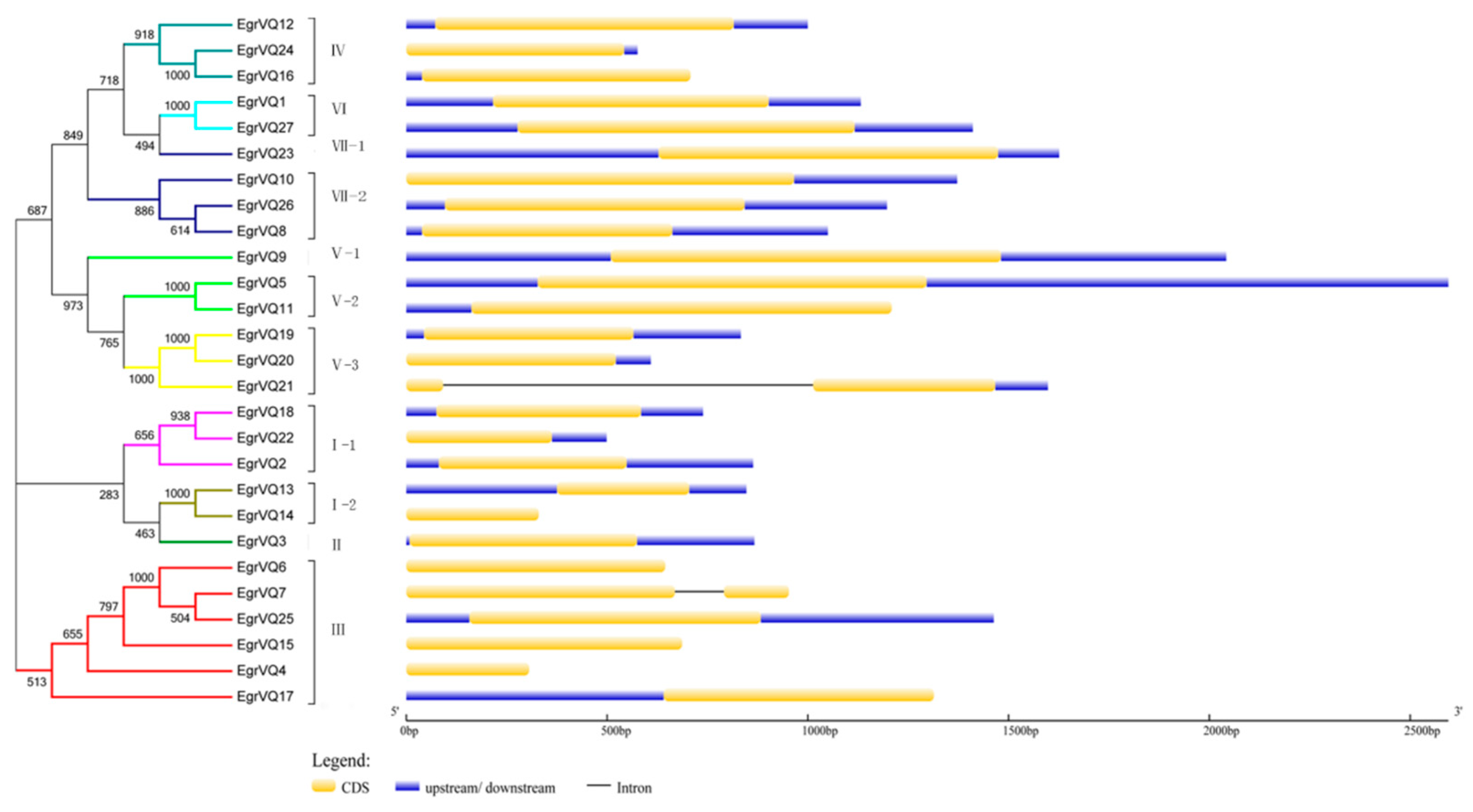
2.5. Analysis of Gene Structural and Conserved motifs of EgrVQ Genes
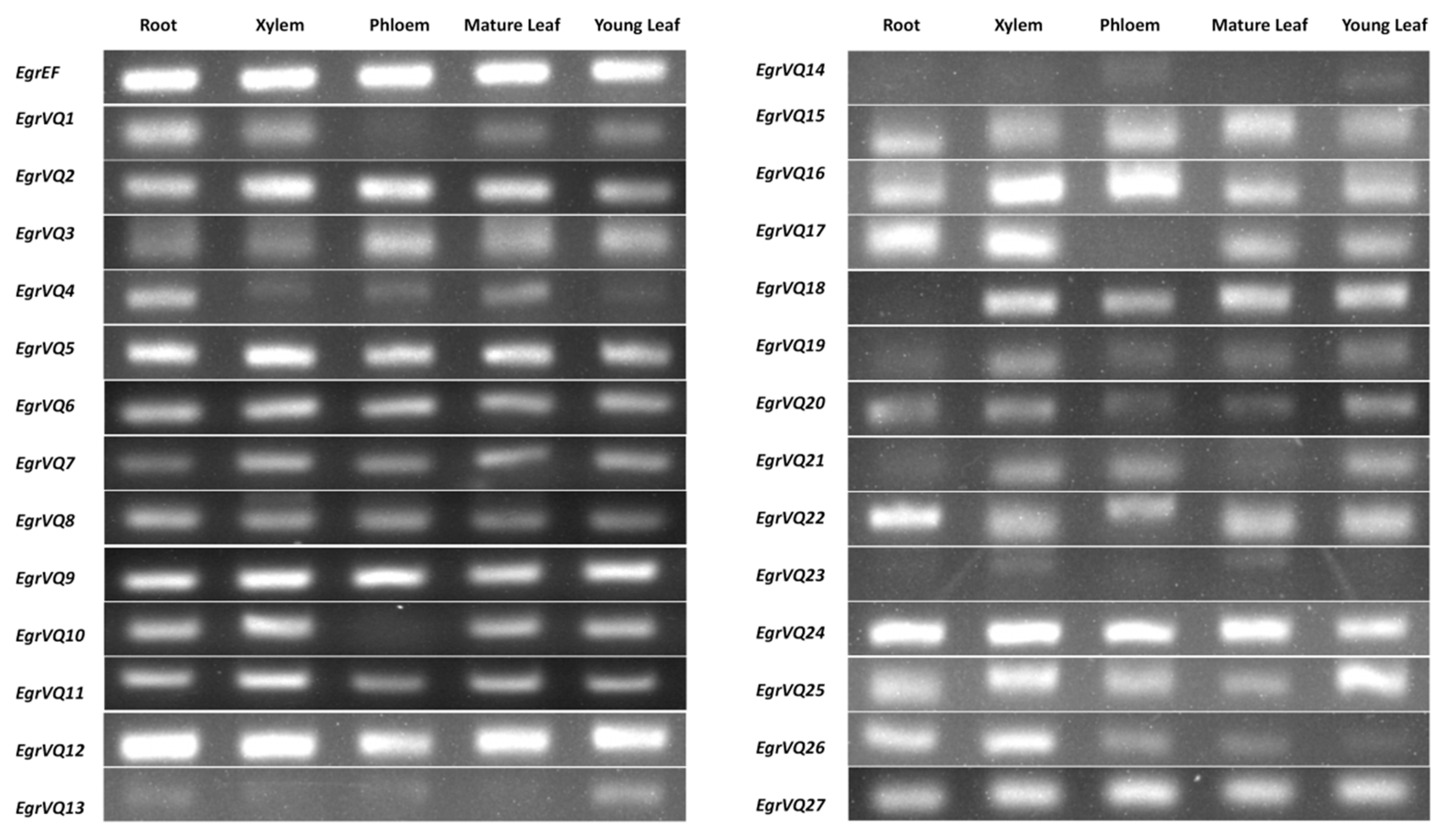
2.6. Expression Patters of EgrVQ Genes in Different Tissues of E. grandis
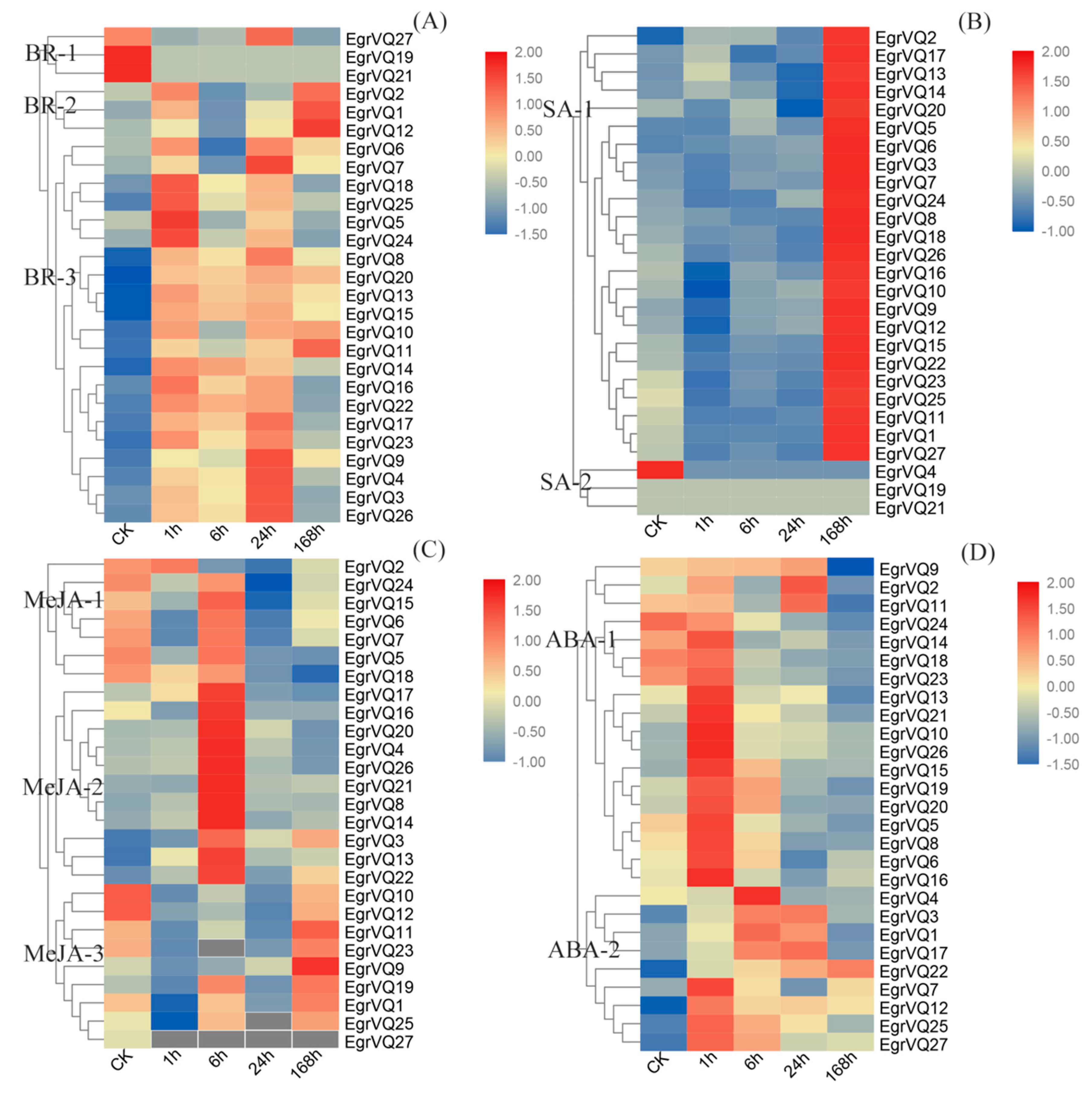
2.7. Expression Profiles of EgrVQ Genes in Response to Hormones
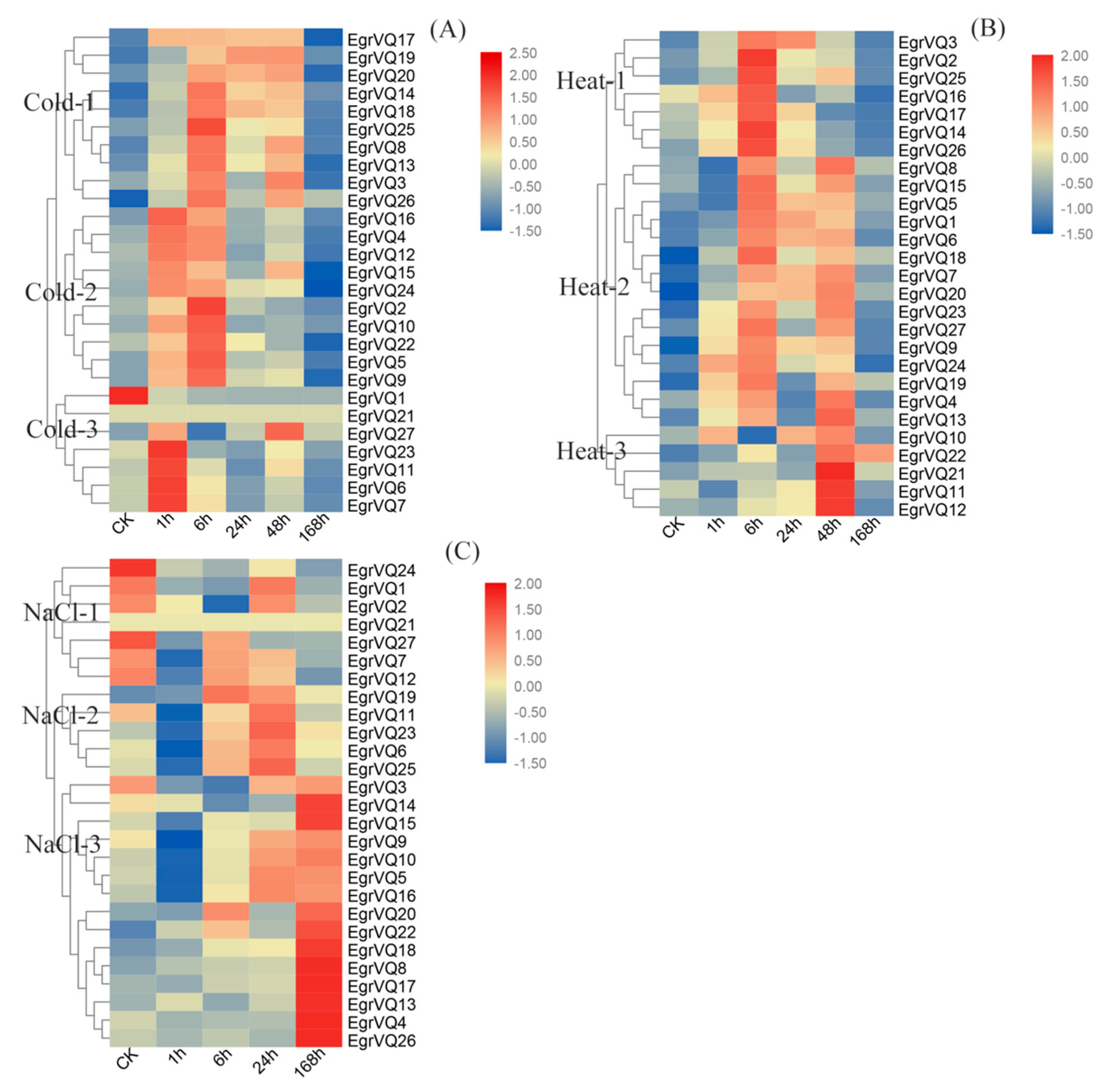
2.8. Expression Profiles of EgrVQ Genes in Response to Stress Treatments
3. Discussion
4. Materials and Methods
4.1. Identification of VQ Gene Family in E. grandis
4.2. Chromosomal Location, Gene Duplication, and Identification of Paralogs and Orthologs of EgrVQ Genes
4.3. Promoter Analysis of EgrVQ Genes
4.4. Phylogenetic Analysis and Multiple Sequence Alignment of EgrVQ Proteins
4.5. Gene Structure and Conserved Motifs Analysis of EgrVQ Genes
4.6. Plant Material and Treatments
4.7. RNA Extraction, Semi-Quantitative Real-Time PCR (RT-PCR), and Real-Time Quantitative PCR (qRT-PCR)
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jing, Y.; Lin, R. The VQ Motif-Containing Protein Family of Plant-Specific Transcriptional Regulators. Plant Physiol. 2015, 169, 371–378. [Google Scholar] [CrossRef] [PubMed]
- Buscaill, P.; Rivas, S. Transcriptional control of plant defence responses. Curr. Opin. Plant Biol. 2014, 20, 35–46. [Google Scholar] [CrossRef]
- Wang, A.; Garcia, D.; Zhang, H.; Feng, K.; Chaudhury, A.; Berger, F.; Peacock, W.J.; Dennis, E.S.; Luo, M. The VQ motif protein IKU1 regulates endosperm growth and seed size in Arabidopsis. Plant J. 2010, 63, 670–679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pecher, P.; Eschen-Lippold, L.; Herklotz, S.; Kuhle, K.; Naumann, K.; Bethke, G.; Uhrig, J.; Weyhe, M.; Scheel, D.; Lee, J. The Arabidopsis thaliana mitogen-activated protein kinases MPK3 and MPK6 target a subclass of ‘VQ-motif’-containing proteins to regulate immune responses. New Phytol. 2014, 203, 592–606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Zhou, Y.; Yang, Y.; Chi, Y.-J.; Zhou, J.; Chen, J.-Y.; Wang, F.; Fan, B.; Shi, K.; Zhou, Y.-H. Structural and functional analysis of VQ motif-containing proteins in Arabidopsis as interacting proteins of WRKY transcription factors. Plant Physiol. 2012, 159, 810–825. [Google Scholar] [CrossRef]
- Li, N.; Li, X.; Xiao, J.; Wang, S. Comprehensive analysis of VQ motif-containing gene expression in rice defense responses to three pathogens. Plant Cell Rep. 2014, 33, 1493–1505. [Google Scholar] [CrossRef]
- Chu, W.; Liu, B.; Wang, Y.; Pan, F.; Chen, Z.; Yan, H.; Yan, X. Genome-wide analysis of poplar VQ gene family and expression profiling under PEG, NaCl, and SA treatments. Tree Genet. Genomes 2016, 12, 124. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, H.; Zhu, D.; Gao, Y.; Yan, H.; Xiang, Y. Genome-wide analysis of VQ motif-containing proteins in Moso bamboo (Phyllostachys edulis). Planta 2017, 246, 165–181. [Google Scholar] [CrossRef]
- Guo, J.; Chen, J.; Yang, J.; Yu, Y.; Yang, Y.; Wang, W. Identification, characterization and expression analysis of the VQ motif-containing gene family in tea plant (Camellia sinensis). BMC Genom. 2018, 19, 710. [Google Scholar] [CrossRef]
- Liu, X.; Wang, X.; Pang, Y.; Liang, J.; Liu, S.; Sun, X.; Tang, K. Molecular cloning and characterization of a novel WRKY gene from Brassica chinensis. Mol. Biol. 2006, 40, 816. [Google Scholar] [CrossRef]
- Chi, Y.; Yang, Y.; Zhou, Y.; Zhou, J.; Fan, B.; Yu, J.Q.; Chen, Z. Protein-Protein Interactions in the Regulation of WRKY Transcription Factors. Mol. Plant 2013, 6, 287–300. [Google Scholar]
- Xie, T.; Chen, C.; Li, C.; Liu, J.; Liu, C.; He, Y. Genome-wide investigation of WRKY gene family in pineapple: Evolution and expression profiles during development and stress. BMC Genom. 2018, 19, 490. [Google Scholar] [CrossRef]
- Lin, Z.; Li, Y.; Wang, F.; Cheng, Y.; Fan, B.; Yu, J.-Q.; Chen, Z. Arabidopsis sigma factor binding proteins are activators of the WRKY33 transcription factor in plant defense. Plant Cell 2011, 23, 3824–3841. [Google Scholar]
- Song, W.; Zhao, H.; Zhang, X.; Lei, L.; Lai, J. Genome-Wide Identification of VQ Motif-Containing Proteins and their Expression Profiles Under Abiotic Stresses in Maize. Front. Plant Sci. 2016, 6, 1177. [Google Scholar] [CrossRef]
- Perruc, E.; Charpenteau, M.; Ramirez, B.C.; Jauneau, A.; Galaud, J.P.; Ranjeva, R.; Ranty, B. A novel calmodulin-binding protein functions as a negative regulator of osmotic stress tolerance in Arabidopsis thaliana seedlings. Plant J. 2004, 38, 410–420. [Google Scholar] [CrossRef]
- Hu, P.; Zhou, W.; Cheng, Z.; Fan, M.; Wang, L.; Xie, D. JAV1 Controls Jasmonate-Regulated Plant Defense. Mol. Cell 2013, 50, 504–515. [Google Scholar]
- Kim, D.Y.; Kwon, S.I.; Choi, C.; Lee, H.; Ahn, I.; Park, S.R.; Bae, S.C.; Lee, S.C.; Hwang, D.J. Expression analysis of rice VQ genes in response to biotic and abiotic stresses. Gene 2013, 529, 208–214. [Google Scholar]
- Wang, X.; Zhang, H.; Sun, G.; Jin, Y.; Qiu, L.J. Identification of active VQ motif-containing genes and the expression patterns under low nitrogen treatment in soybean. Gene 2014, 543, 237–243. [Google Scholar]
- Rushton, P.J.; Somssich, I.E.; Ringler, P.; Shen, Q.J. WRKY transcription factors. Trends Plant Sci. 2010, 15, 247–258. [Google Scholar] [CrossRef]
- Viana, V.E.; Busanello, C.; da Maia, L.C.; Pegoraro, C.; de Oliveira, A.C. Activation of rice WRKY transcription factors: an army of stress fighting soldiers? Curr. Opin. Plant Biol. 2018. [Google Scholar] [CrossRef]
- Birkenbihl, R.P.; Diezel, C.; Somssich, I.E. Arabidopsis WRKY33 is a key transcriptional regulator of hormonal and metabolic responses toward Botrytis cinerea infection. Plant Physiol. 2012, 159, 266–285. [Google Scholar]
- Hu, Y.; Chen, L.; Wang, H.; Zhang, L.; Wang, F.; Yu, D. Arabidopsis transcription factor WRKY8 functions antagonistically with its interacting partner VQ9 to modulate salinity stress tolerance. Plant J. 2013, 74, 730–745. [Google Scholar] [CrossRef] [Green Version]
- Batish, D.R.; Singh, H.P.; Kohli, R.K.; Shalinder, K. Eucalyptus essential oil as a natural pesticide. For. Ecol. Manag. 2008, 256, 2166–2174. [Google Scholar] [CrossRef]
- Barton, A. The Oil Mallee Project: A Multifaceted Industrial Ecology Case Study. J. Ind. Ecol. 2010, 3, 161–176. [Google Scholar]
- Popescu, C.-M.; Popescu, M.-C.; Singurel, G.; Vasile, C.; Argyropoulos, D.S.; Willfor, S. Spectral characterization of eucalyptus wood. Appl. Spectrosc. 2007, 61, 1168–1177. [Google Scholar]
- Myburg, A.A.; Grattapaglia, D.; Tuskan, G.A.; Hellsten, U.; Hayes, R.D.; Grimwood, J.; Jenkins, J.; Lindquist, E.; Tice, H.; Bauer, D. The genome of Eucalyptus grandis. Nature 2014, 510, 356–362. [Google Scholar] [Green Version]
- Peng, Z.; Lu, Y.; Li, L.; Zhao, Q.; Feng, Q.; Gao, Z.; Lu, H.; Hu, T.; Yao, N.; Liu, K. The draft genome of the fast-growing non-timber forest species moso bamboo (Phyllostachys heterocycla). Nat. Genet. 2013, 45, 456–461. [Google Scholar] [Green Version]
- Wang, M.; Vannozzi, A.; Wang, G.; Zhong, Y.; Corso, M.; Cavallini, E.; Cheng, Z.M. A comprehensive survey of the grapevine VQ gene family and its transcriptional correlation with WRKY proteins. Front. Plant Sci. 2015, 6, 417. [Google Scholar] [CrossRef]
- Xie, Y.D.; Li, W.; Guo, D.; Dong, J.; Zhang, Q.; Fu, Y.; Ren, D.; Peng, M.; Xia, Y. The Arabidopsis gene SIGMA FACTOR-BINDING PROTEIN 1 plays a role in the salicylate- and jasmonate-mediated defence responses. Plant Cell Environ. 2010, 33, 828–839. [Google Scholar]
- Li, Y.; Jing, Y.; Li, J.; Xu, G.; Lin, R. Arabidopsis VQ MOTIF-CONTAINING PROTEIN29 Represses Seedling Deetiolation by Interacting with PHYTOCHROME-INTERACTING FACTOR1. Plant Physiol. 2014, 164, 2068–2080. [Google Scholar] [CrossRef]
- Ali, M.R.; Uemura, T.; Ramadan, A.; Adachi, K.; Nemoto, K.; Nozawa, A.; Hoshino, R.; Abe, H.; Sawasaki, T.; Arimura, G.-I. The Ring-type E3 Ubiquitin Ligase JUL1 Targets the VQ-motif Protein JAV1 to Coordinate Jasmonate Signaling. Plant Physiol. 2019, 179, 1273–1284. [Google Scholar] [CrossRef]
- Fariduddin, Q.; Mir, B.A.; Yusuf, M.; Ahmad, A. 24-epibrassinolide and/or putrescine trigger physiological and biochemical responses for the salt stress mitigation in Cucumis sativus L. Photosynthetica 2014, 52, 464–474. [Google Scholar] [CrossRef]
- Clouse, S.D. Brassinosteroid/Abscisic Acid Antagonism in Balancing Growth and Stress. Dev. Cell 2016, 38, 118–120. [Google Scholar]
- Ali, S.S.; Kumar, G.B.S.; Mojibur, K.; Doohan, F.M. Brassinosteroid enhances resistance to fusarium diseases of barley. Phytopathology 2013, 103, 1260–1267. [Google Scholar]
- Nawaz, F.; Naeem, M.; Zulfiqar, B.; Akram, A.; Ashraf, M.Y.; Raheel, M.; Shabbir, R.N.; Hussain, R.A.; Anwar, I.; Aurangzaib, M. Understanding brassinosteroid-regulated mechanisms to improve stress tolerance in plants: A critical review. Environ. Sci. Pollut. Res. Int. 2017, 24, 15959–15975. [Google Scholar]
- Zhang, G.; Wang, F.; Li, J.; Ding, Q.; Zhang, Y.; Li, H.; Zhang, J.; Gao, J. Genome-Wide Identification and Analysis of the VQ Motif-Containing Protein Family in Chinese Cabbage (Brassica rapa L. ssp. Pekinensis). Int. J. Mol. Sci. 2015, 16, 28683–28704. [Google Scholar] [CrossRef]
- Gasteiger, E.; Gattiker, A.; Hoogland, C.; Ivanyi, I.; Appel, R.D.; Bairoch, A. ExPASy: The proteomics server for in-depth protein knowledge and analysis. Nucleic Acids Res. 2003, 31, 3784–3788. [Google Scholar] [CrossRef]
- Horton, P.; Park, K.-J.; Obayashi, T.; Fujita, N.; Harada, H.; Adams-Collier, C.J.; Nakai, K. WoLF PSORT: Protein localization predictor. Nucleic Acids Res. 2007, 35, W585–W587. [Google Scholar] [CrossRef]
- Emanuelsson, O.; Brunak, S.; von Heijne, G.; Nielsen, H. Locating proteins in the cell using TargetP, SignalP and related tools. Nat. Protoc. 2007, 2, 953–971. [Google Scholar] [CrossRef]
- Voorrips, R.E. MapChart: Software for the graphical presentation of linkage maps and QTLs. J. Hered. 2002, 93, 77–78. [Google Scholar] [CrossRef]
- Blanc, G.; Wolfe, K.H. Widespread paleopolyploidy in model plant species inferred from age distributions of duplicate genes. Plant Cell 2004, 16, 1667–1678. [Google Scholar] [CrossRef]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef] [Green Version]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X Windows Interface: Flexible Strategies for Multiple Sequence Alignment Aided by Quality Analysis Tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Hu, B.; Jin, J.; Guo, A.Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2014, 31, 1296–1297. [Google Scholar] [CrossRef]
- Timothy, L.B.; Mikael, B.; Fabian, A.B.; Martin, F.; Charles, E.G.; Luca, C.; Jingyuan, R.; Wilfred, W.L.; William, S.N. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- De Oliveira, L.A.; Breton, M.C.; Bastolla, F.M.; Camargo Sda, S.; Margis, R.; Frazzon, J.; Pasquali, G. Reference Genes for the Normalization of Gene Expression in Eucalyptus Species. Plant Cell Physiol. 2012, 53, 405–422. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Gene ID | Chr | Location Coordinates (5′–3′) | ORF Length (bp) | Exons | Protein Length (aa) | Protein MW (Da) | Protein PI | Subcellular WoLF PSORT | Localization TargetP |
|---|---|---|---|---|---|---|---|---|---|---|
| EgrVQ1 | Eucgr.A02182.1 | 1 | 37113647..37114778 | 687 | 1 | 228 | 22929.82 | 9.50 | Nucleus | Chloroplast |
| EgrVQ2 | Eucgr.A02242.1 | 1 | 37821284..37822147 | 468 | 1 | 155 | 16913.17 | 7.92 | Mitochondrion | Chloroplast |
| EgrVQ3 | Eucgr.A02450.1 | 1 | 39836460..39837326 | 567 | 1 | 188 | 21252.59 | 5.38 | Nucleus | Any other location |
| EgrVQ4 | Eucgr.A02544.1 | 1 | 40671118..40671423 | 306 | 1 | 101 | 11278.94 | 9.36 | Cytoplasm | Chloroplast |
| EgrVQ5 | Eucgr.B00035.1 | 6 | 35375963..35378557 | 969 | 1 | 322 | 34040.85 | 10.02 | Nucleus | Chloroplast |
| EgrVQ6 | Eucgr.C02621.1 | 3 | 51875030..51875674 | 645 | 1 | 215 | 22454.41 | 6.65 | Mitochondrion | Chloroplast |
| EgrVQ7 | Eucgr.C02629.1 | 3 | 51927510..51928462 | 831 | 2 | 276 | 28709.31 | 7.15 | Nucleus | Chloroplast |
| EgrVQ8 | Eucgr.D00026.1 | 4 | 216578..217627 | 62 | 1 | 207 | 22664.50 | 5.91 | Nucleus | Any other location |
| EgrVQ9 | Eucgr.D02200.1 | 4 | 35568137..35570178 | 972 | 1 | 323 | 35209.97 | 10.66 | Nucleus | Any other location |
| EgrVQ10 | Eucgr.F00302.1 | 6 | 5689343..5690714 | 966 | 1 | 321 | 34599.00 | 9.66 | Nucleus | Chloroplast |
| EgrVQ11 | Eucgr.F00371.1 | 6 | 5040555..5041763 | 1047 | 1 | 348 | 37574.61 | 9.99 | Nucleus | Chloroplast |
| EgrVQ12 | Eucgr.F01148.1 | 6 | 15726039..15727038 | 744 | 1 | 247 | 26541.68 | 9.01 | Nucleus | Chloroplast |
| EgrVQ13 | Eucgr.F01407.1 | 6 | 19292825..19293671 | 330 | 1 | 109 | 12303.87 | 7.79 | Chloroplast | Any other location |
| EgrVQ14 | Eucgr.F01408.1 | 6 | 19298931..19299260 | 330 | 1 | 109 | 12161.73 | 6.73 | Cytoplasm | Any other location |
| EgrVQ15 | Eucgr.F02723.1 | 6 | 39354381..39355067 | 687 | 1 | 228 | 24468.88 | 9.47 | Nucleus | Any other location |
| EgrVQ16 | Eucgr.F03907.1 | 6 | 49847862..49848569 | 669 | 1 | 222 | 23853.03 | 9.71 | Nucleus | Any other location |
| EgrVQ17 | Eucgr.G02312.1 | 7 | 43757342..43758655 | 675 | 1 | 224 | 24656.69 | 8.68 | Nucleus | Chloroplast |
| EgrVQ18 | Eucgr.G03218.1 | 7 | 52999148..52999886 | 510 | 1 | 169 | 17744.58 | 6.50 | Nucleus | Any other location |
| EgrVQ19 | Eucgr.H02317.1 | 8 | 29626932..29627764 | 522 | 1 | 173 | 18910.74 | 9.93 | Chloroplast | Any other location |
| EgrVQ20 | Eucgr.H02318.1 | 8 | 29632300..29632908 | 522 | 1 | 173 | 18883.63 | 9.93 | Chloroplast | Any other location |
| EgrVQ21 | Eucgr.H02319.1 | 8 | 29636679..29638276 | 546 | 2 | 181 | 19910.03 | 10.05 | Chloroplast | Secretory pathway |
| EgrVQ22 | Eucgr.H04261.1 | 8 | 57364319..57364817 | 363 | 1 | 120 | 13296.04 | 8.87 | Nucleus | Chloroplast |
| EgrVQ23 | Eucgr.H04807.1 | 8 | 67074212..67075837 | 846 | 1 | 281 | 29271.01 | 8.93 | Nucleus | Any other location |
| EgrVQ24 | Eucgr.I00016.1 | 9 | 223638..224213 | 543 | 1 | 180 | 19892.34 | 9.00 | Nucleus | Any other location |
| EgrVQ25 | Eucgr.I00226.1 | 9 | 4661485..4662947 | 726 | 1 | 241 | 25294.49 | 6.29 | Nucleus | Chloroplast |
| EgrVQ26 | Eucgr.K01342.1 | 11 | 16771278..16772474 | 747 | 1 | 248 | 25715.14 | 10.08 | Nucleus | Any other location |
| EgrVQ27 | Eucgr.K01686.1 | 11 | 19466093..19467503 | 840 | 1 | 279 | 28773.80 | 6.59 | Nucleus | Any other location |
| Egr–Egr | Egr–At | Egr–Os | Egr-Pt |
|---|---|---|---|
| EgrVQ6/EgrVQ7 | EgrVQ1/AtVQ24 | EgrVQ12/OsVQ3 | EgrVQ9/PtVQ3 |
| EgrVQ6/EgrVQ25 | EgrVQ5/AtVQ9 | EgrVQ12/OsVQ21 | EgrVQ9/PtVQ12 |
| EgrVQ19/EgrVQ20 | EgrVQ9/AtVQ6 | EgrVQ15/OsVQ8 | EgrVQ12/PtVQ7 |
| EgrVQ19/EgrVQ21 | EgrVQ11/AtVQ9 | EgrVQ15/OsVQ15 | |
| EgrVQ20/EgrVQ21 | EgrVQ12/AtVQ4 | EgrVQ24/OsVQ3 | |
| EgrVQ12/AtVQ19 | |||
| EgrVQ12/AtVQ33 | |||
| EgrVQ13/AtVQ1 | |||
| EgrVQ13/AtVQ10 | |||
| EgrVQ14/AtVQ1 | |||
| EgrVQ14/AtVQ10 | |||
| EgrVQ15/AtVQ3 | |||
| EgrVQ15/AtVQ21 | |||
| EgrVQ16/AtVQ11 | |||
| EgrVQ24AtVQ31 |
| Gene Name | LTG | FTG | VTG | LTS | LTD | YTG | ITG | LTR | LTV | ATG | LTA | LSG |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Arabidopsis | 24 | 5 | 2 | 1 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| Poplar | 39 | 11 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Chinese Cabbage | 43 | 8 | 3 | 1 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 0 |
| Soybean | 55 | 15 | 2 | 1 | 0 | 1 | 0 | 1 | 0 | 0 | 0 | 0 |
| Grapevine | 14 | 3 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| Maize | 42 | 8 | 6 | 0 | 0 | 0 | 2 | 0 | 0 | 2 | 1 | 0 |
| Rice | 28 | 7 | 4 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 |
| Eucalyptus grandis | 20 | 4 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| Gene Name | ABRE 1 | CGTCA-motif 2 | TGACG-motif 3 | TCA-element 4 | MBS 5 | LTR 6 | DRE 7 |
|---|---|---|---|---|---|---|---|
| EgrVQ1 | 1 | 1 | 1 | 1 | 1 | ||
| EgrVQ2 | 1 | 1 | 1 | 1 | 1 | ||
| EgrVQ3 | 1 | 1 | 1 | 1 | |||
| EgrVQ4 | 1 | 1 | 1 | 1 | 1 | ||
| EgrVQ5 | 1 | 1 | 1 | 1 | |||
| EgrVQ6 | 1 | 1 | 1 | 1 | |||
| EgrVQ7 | 1 | 1 | 1 | 1 | |||
| EgrVQ8 | 1 | 1 | 1 | 1 | |||
| EgrVQ9 | 1 | 1 | 1 | 1 | 1 | ||
| EgrVQ10 | 1 | 1 | 1 | 1 | 1 | 1 | |
| EgrVQ11 | 1 | 1 | 1 | 1 | |||
| EgrVQ12 | 1 | 1 | 1 | ||||
| EgrVQ13 | 1 | 1 | 1 | 1 | |||
| EgrVQ14 | 1 | 1 | 1 | 1 | |||
| EgrVQ15 | 1 | 1 | 1 | 1 | 1 | ||
| EgrVQ16 | 1 | 1 | 1 | 1 | 1 | ||
| EgrVQ17 | 1 | 1 | 1 | 1 | |||
| EgrVQ18 | 1 | 1 | 1 | 1 | 1 | ||
| EgrVQ19 | 2 | 1 | 1 | 1 | 1 | ||
| EgrVQ20 | 1 | 1 | 1 | 1 | |||
| EgrVQ21 | 1 | 1 | 1 | 1 | |||
| EgrVQ22 | 1 | 1 | 1 | 1 | |||
| EgrVQ23 | 1 | 1 | 1 | 1 | 1 | ||
| EgrVQ24 | 1 | 1 | 1 | 1 | |||
| EgrVQ25 | 1 | 1 | 1 | 1 | 1 | ||
| EgrVQ26 | 1 | 1 | 1 | 1 | 1 | ||
| EgrVQ27 | 1 | 1 | 1 | 1 | 1 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, H.; Wang, Y.; Hu, B.; Qiu, Z.; Zeng, B.; Fan, C. Genome-Wide Characterization, Evolution, and Expression Profiling of VQ Gene Family in Response to Phytohormone Treatments and Abiotic Stress in Eucalyptus grandis. Int. J. Mol. Sci. 2019, 20, 1765. https://doi.org/10.3390/ijms20071765
Yan H, Wang Y, Hu B, Qiu Z, Zeng B, Fan C. Genome-Wide Characterization, Evolution, and Expression Profiling of VQ Gene Family in Response to Phytohormone Treatments and Abiotic Stress in Eucalyptus grandis. International Journal of Molecular Sciences. 2019; 20(7):1765. https://doi.org/10.3390/ijms20071765
Chicago/Turabian StyleYan, Huifang, Yujiao Wang, Bing Hu, Zhenfei Qiu, Bingshan Zeng, and Chunjie Fan. 2019. "Genome-Wide Characterization, Evolution, and Expression Profiling of VQ Gene Family in Response to Phytohormone Treatments and Abiotic Stress in Eucalyptus grandis" International Journal of Molecular Sciences 20, no. 7: 1765. https://doi.org/10.3390/ijms20071765