Comprehensive Genomic Analysis and Expression Profiling of Diacylglycerol Kinase (DGK) Gene Family in Soybean (Glycine max) under Abiotic Stresses

Abstract
:1. Introduction
2. Results
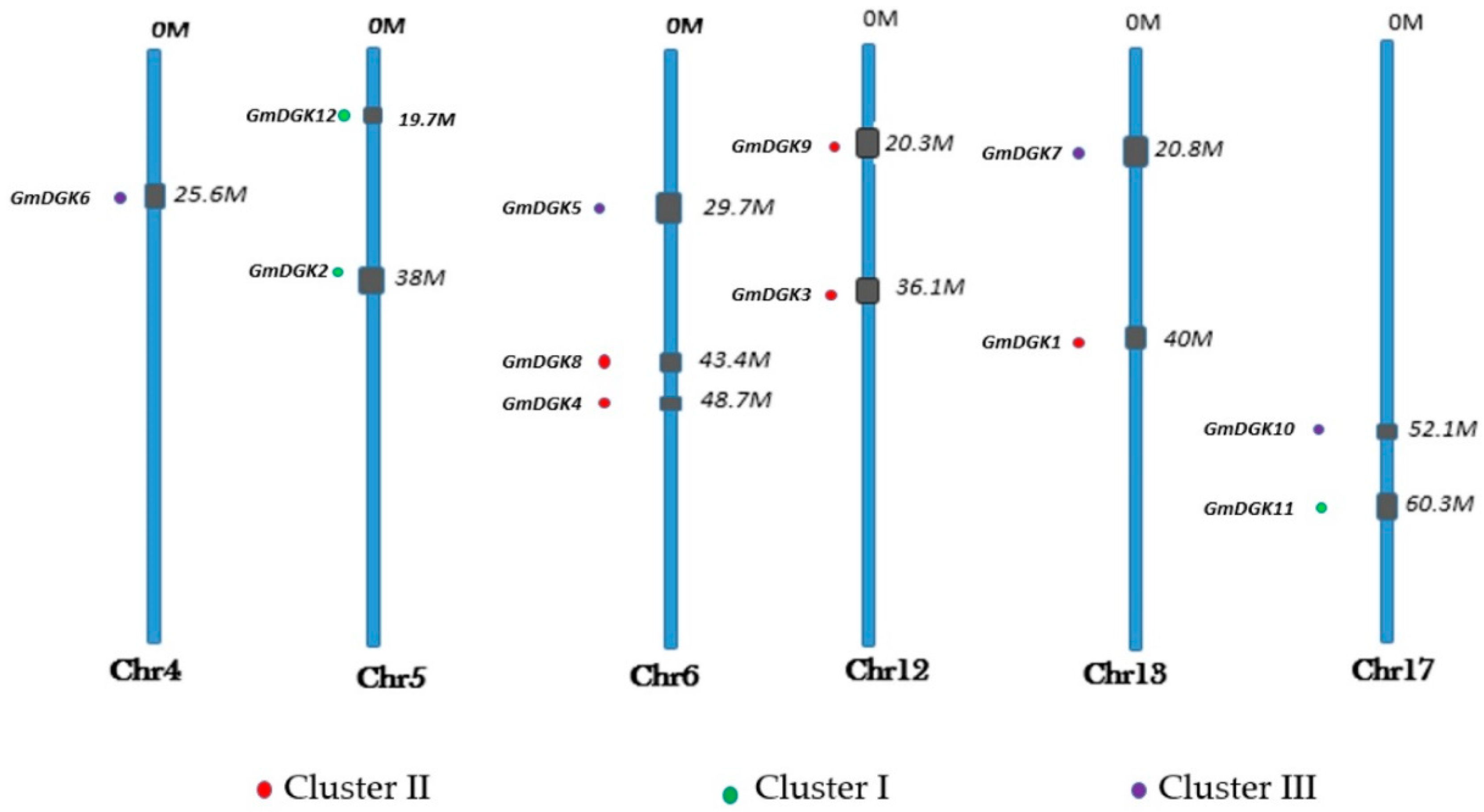
2.1. Genome-Wide Identification and Chromosomal Distribution of DGK Gene Family Members in Soybean
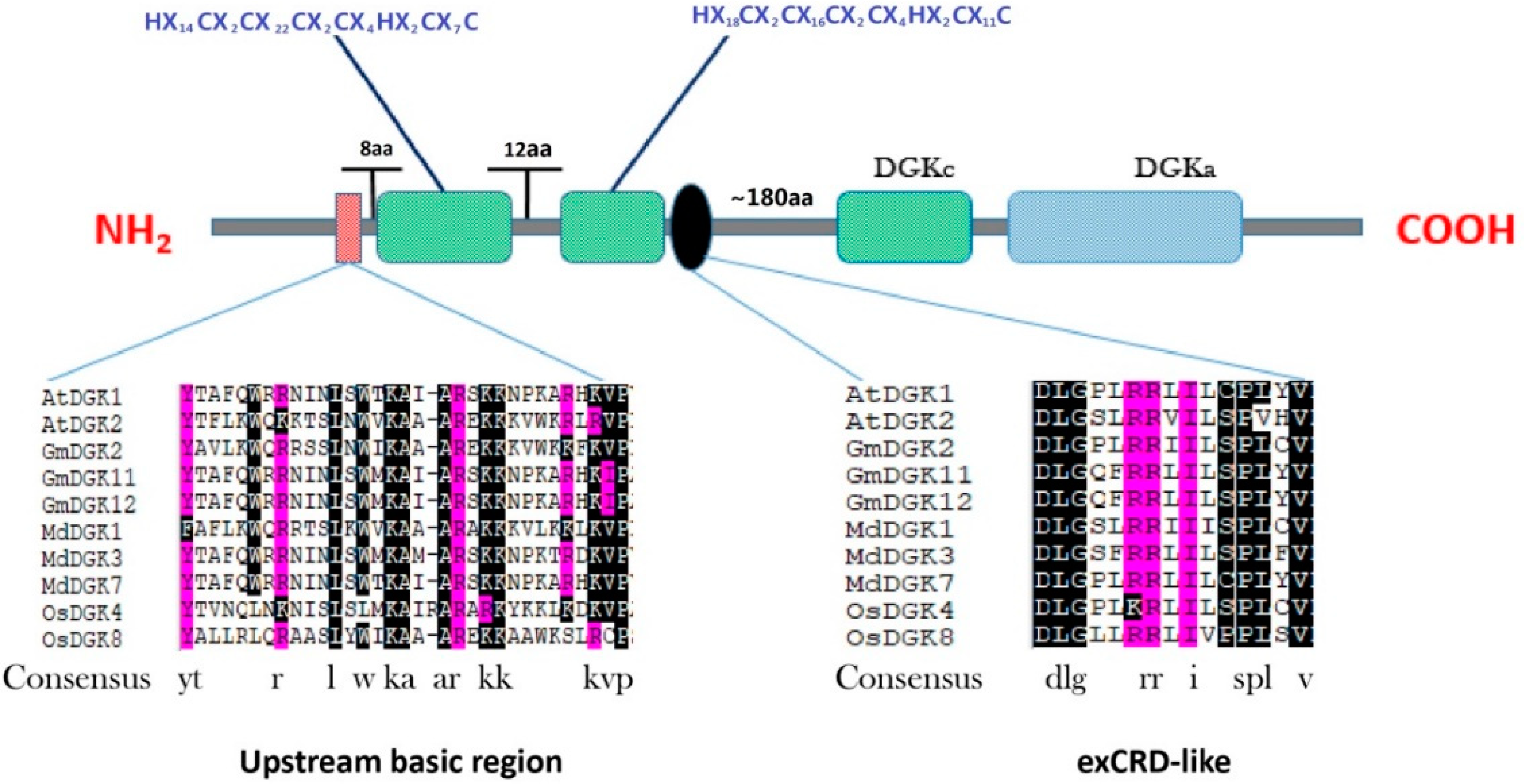
2.2. Multiple Sequence Alignments and Sequence Characterization of the Soybean DGK Genes
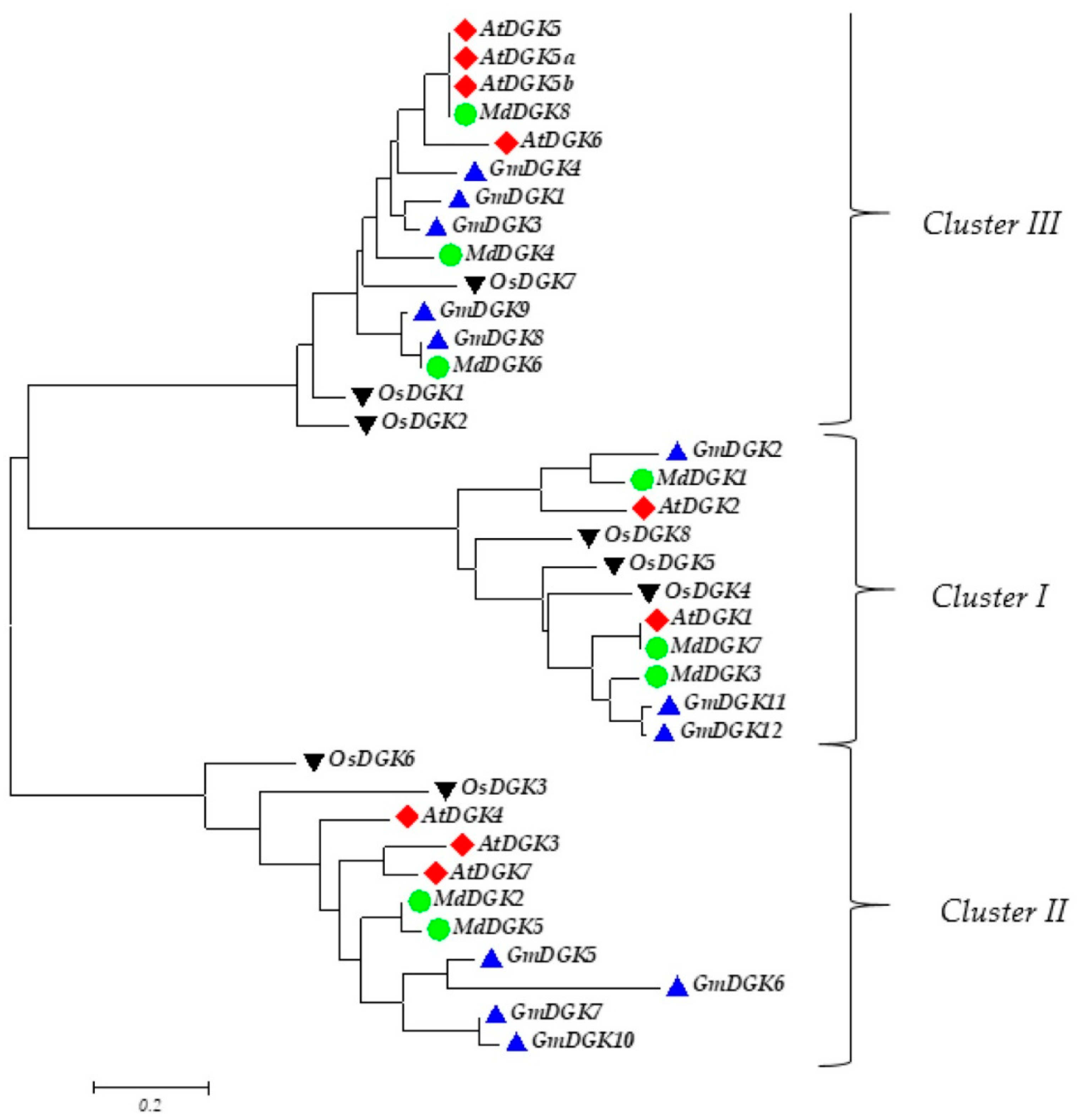
2.3. Phylogenetic Analysis of Soybean DGK Gene Family
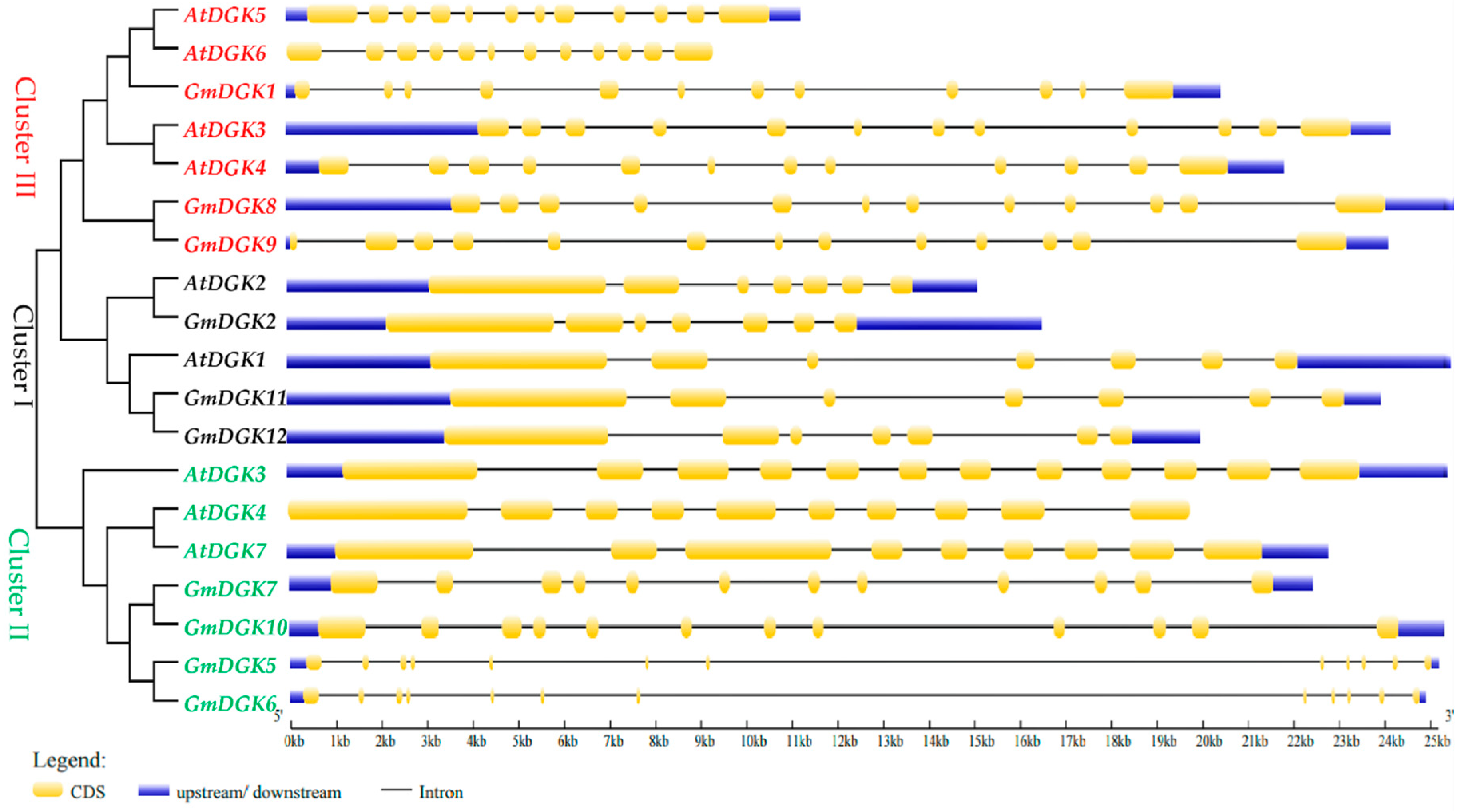
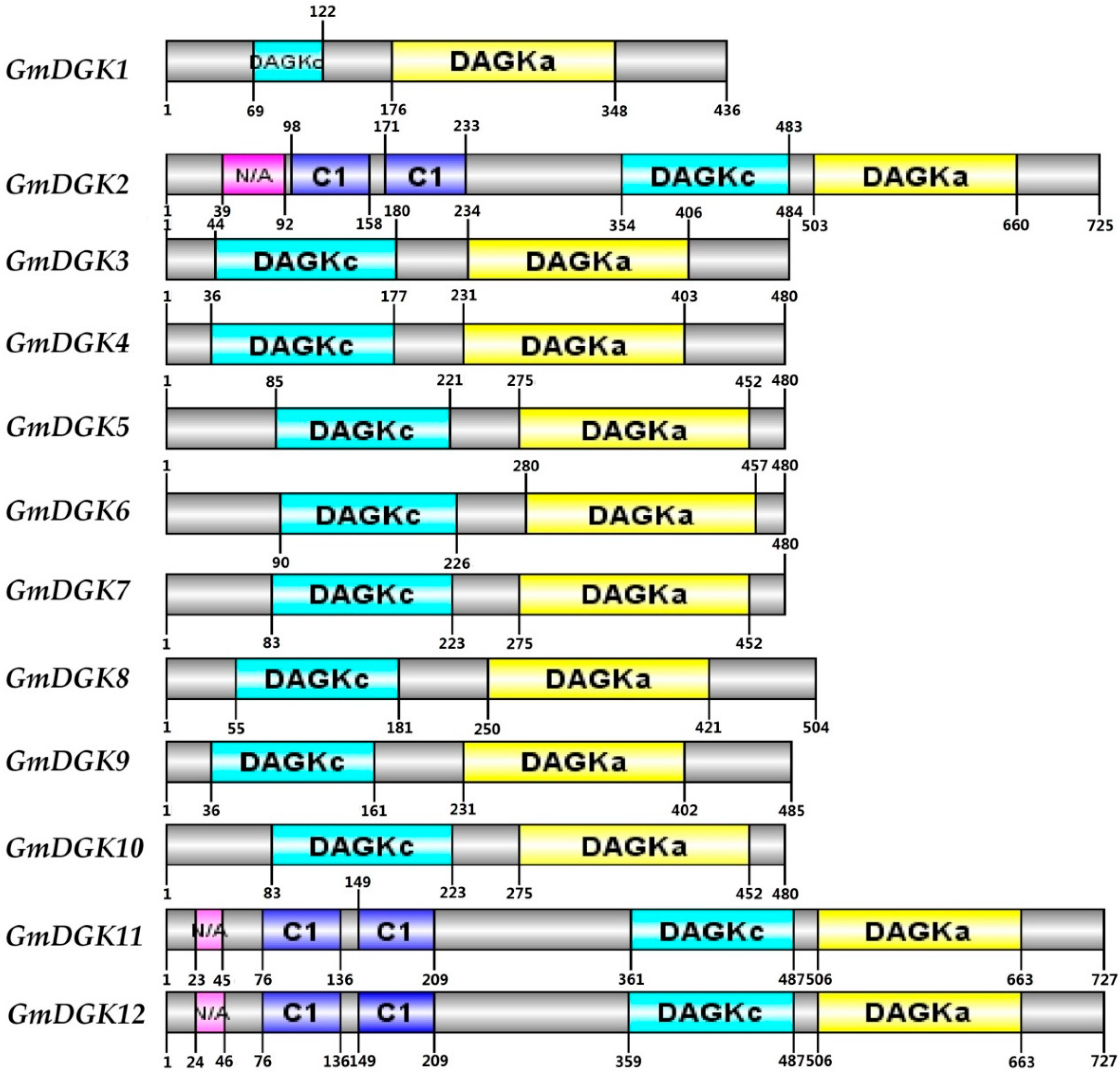
2.4. Structures and Protein Motifs of Soybean DGK Genes
2.5. Protein Domains in Soybean DGKs
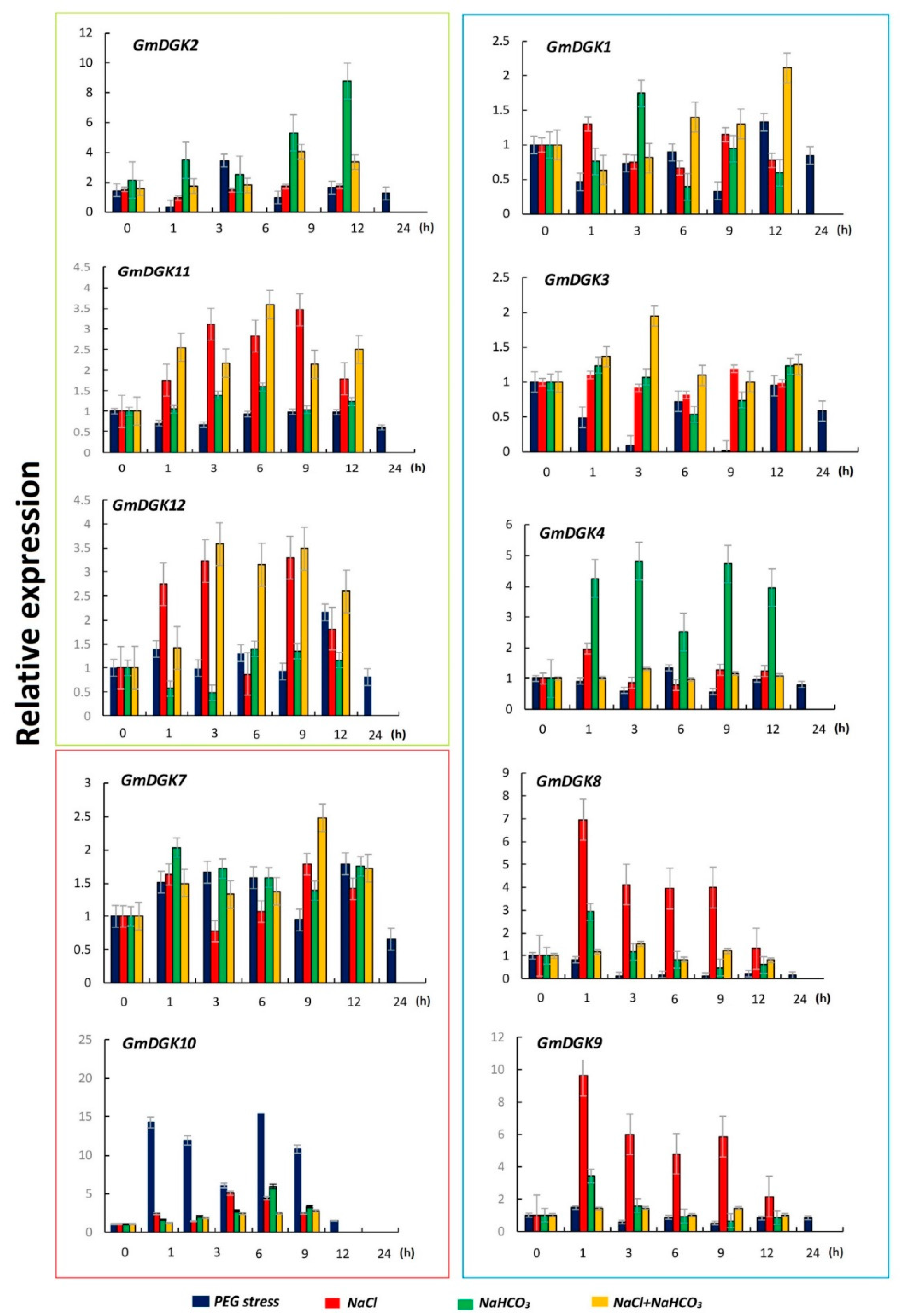
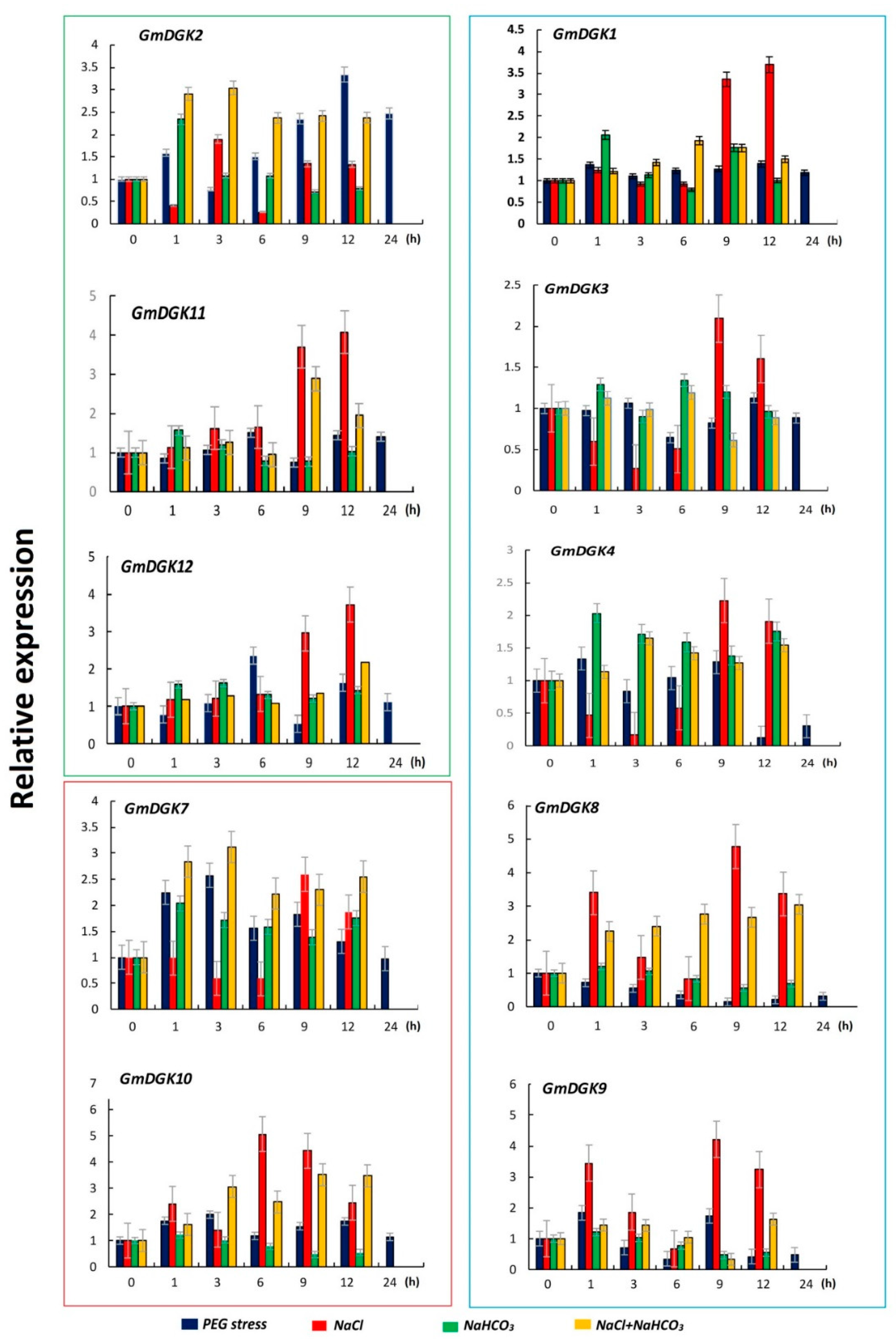
2.6. Expression Analysis of GmDGKs under Abiotic Stress
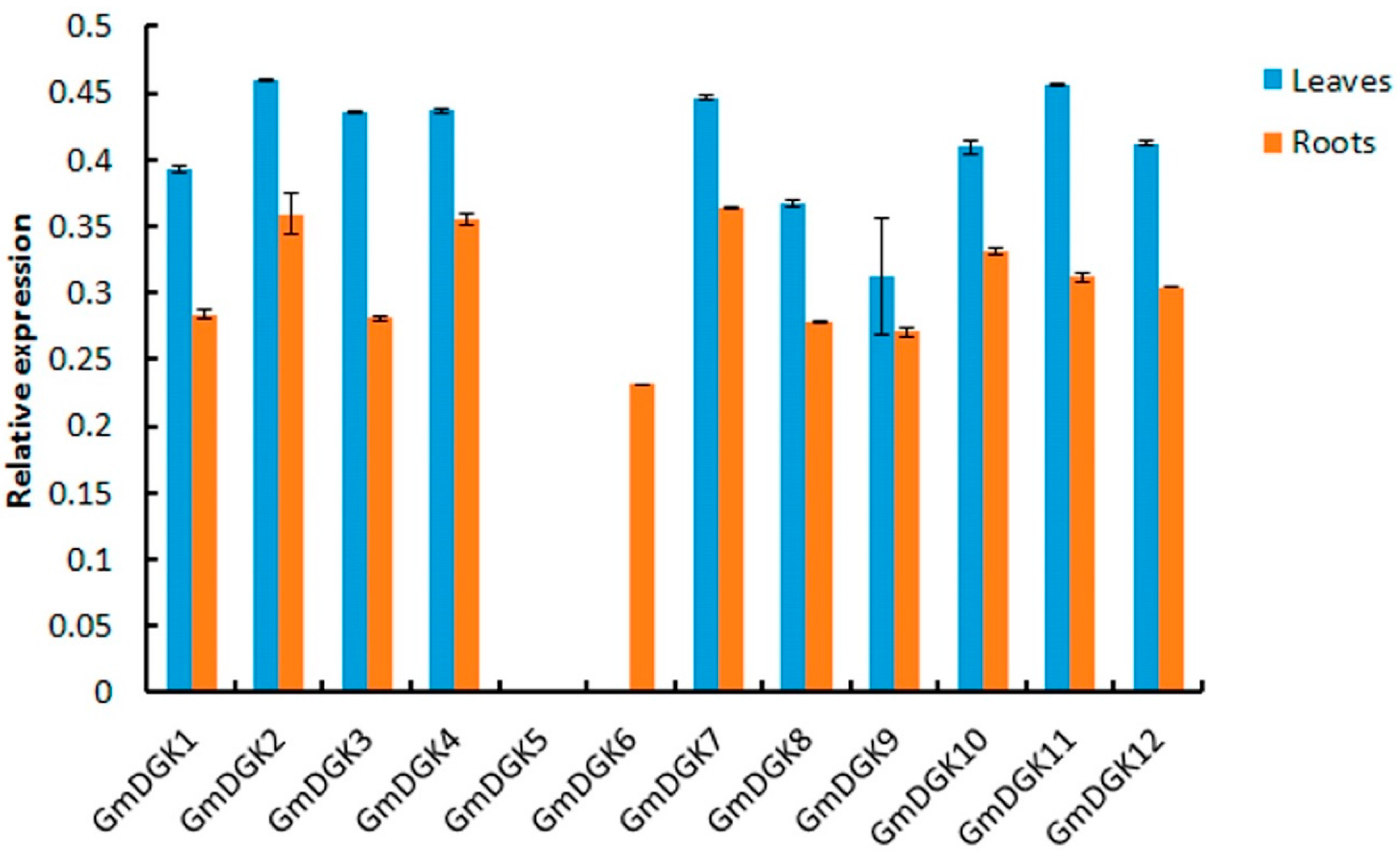
2.7. Expression Profiles of Soybean DGK Genes in Different Tissues
2.8. Prediction of Subcellular Locations for DGK Proteins in Soybean
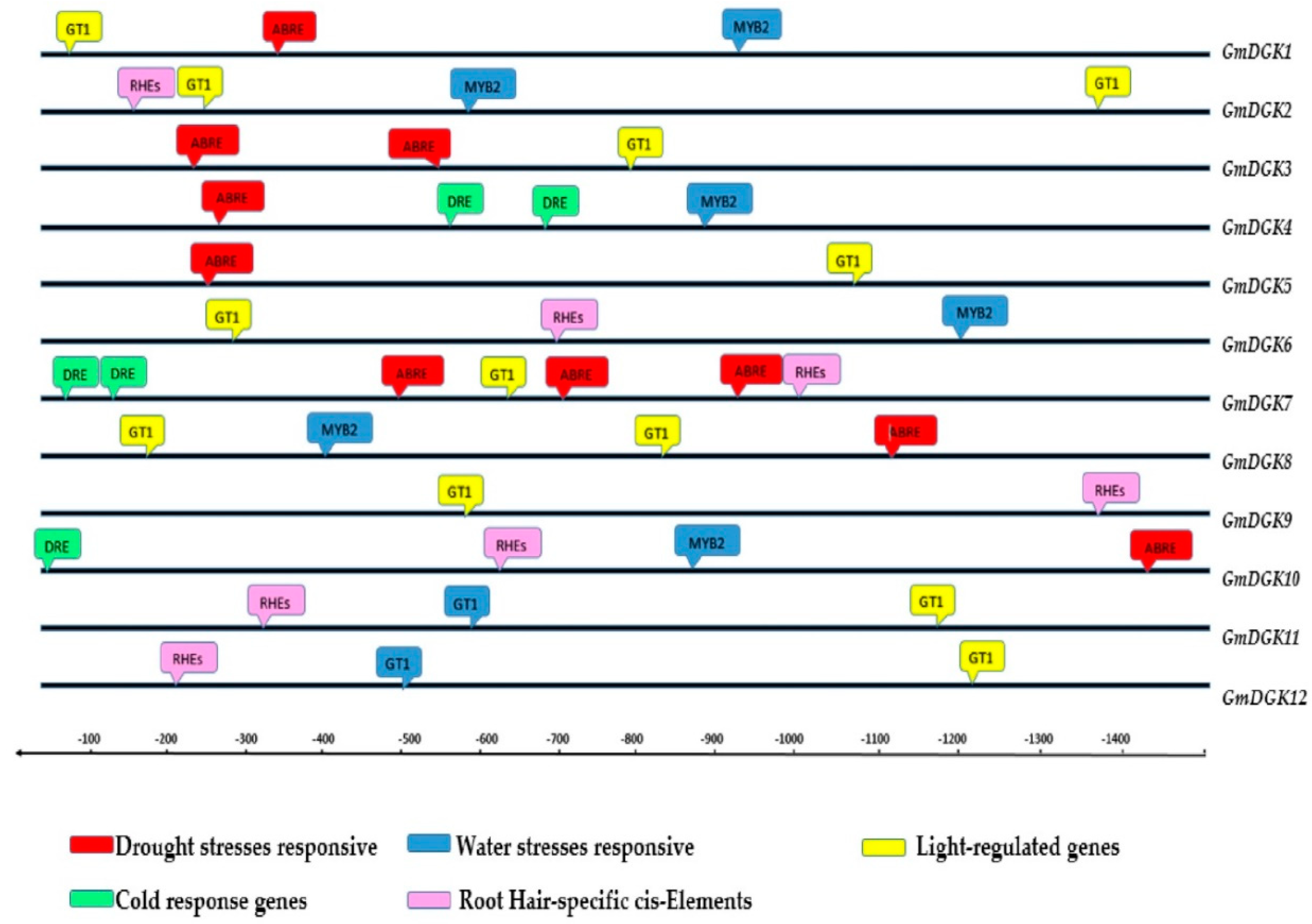
2.9. Cis-Acting Elements in the Promoter of the Soybean DGK Genes
3. Discussion
4. Materials and Methods
4.1. Localization and Identification of the Soybean DGK Genes on Chromosomes
4.2. Multiple Sequence Alignments and Sequence Characterization of the Soybean DGK Genes
4.3. Exon/Intron Organization and Motifs
4.4. Proteins Domain Analysis and the Phylogenetic Relationship of GmDGK Proteins
4.5. Predictions of Subcellular Localization and Promoter Element Analysis for Soybean DGK Genes
4.6. Plant Materials and Stress Treatments
4.7. RNA Extraction, Complementary DNA Synthesis, and Expression Analysis of Selected Soybean DGK Genes
4.8. Quantitative Real-Time PCR Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ABA | abscisic acid |
| qRT-PCR | quantitative real-time PCR |
| SA | salicylic acid |
| GA | gibberellins |
| MeJA | methyl jasmonate |
| DGK | diacylglycerol kinase |
| PI | isoelectric point |
| aa | amino acid |
References
- Fathi, A.; Tari, D.B. Effect of Drought Stress and its Mechanism in Plants. Int. J. Life Sci. 2016, 10, 1. [Google Scholar] [CrossRef]
- Mahajan, S.; Tuteja, N. Cold, salinity and drought stress: An overview. Arch. Biochem. Biophys. 2005, 444, 139–158. [Google Scholar] [CrossRef]
- Pagano, M.C.; Miransari, M. The importance of soybean production worldwide. Abiotic Biotic Stresses Soybean Prod. 2016, 1–26. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, T.; Ding, S. Effect of drought and rewatering on photosynthetic physio-ecological characteristics of soybean. Acta Ecol. Sin. 2006, 26, 2073–2078. [Google Scholar] [CrossRef]
- Souza, G.M.; Catuchi, T.A.; Bertolli, S.C.; Soratto, R.P. Soybean Under Water Deficit: Physiological and Yield Responses. In A Comprehensive Survey of International Soybean Research—Genetics, Physiology, Agronomy and Nitrogen Relationships; InTech: London, UK, 2013. [Google Scholar] [CrossRef] [Green Version]
- Yang, P.H.; Li, G.Q.; Guo, L.; Wu, S.J. Effect of drought stress on plasma membrane permeability of soybean varieties during the flowering-podding stage. Agric. Res. Arid Areas 2003, 21, 127–130. [Google Scholar]
- Munnik, T.; Vermeer, J.E.M. Osmotic stress-induced phosphoinositide and inositol phosphate signaling in plants. Plant Cell Environ. 2010, 33, 655–669. [Google Scholar] [CrossRef] [PubMed]
- Hou, Q.; Ufer, G.; Bartels, D. Lipid signaling in plant responses to abiotic stress. Plant Cell Environ. 2016, 39, 1029–1048. [Google Scholar] [CrossRef]
- Arisz, S.A.; Testerink, C.; Munnik, T. Plant PA signaling via diacylglycerol kinase. Biochim. Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2009, 1791, 869–875. [Google Scholar] [CrossRef]
- Wissing, J.B.; Wagner, K.G. Diacylglycerol kinase from suspension cultured plant cells: Characterization and subcellular localization. Plant Physiol. 1992, 98, 1148–1153. [Google Scholar] [CrossRef]
- Snedden, W.A.; Blumwald, E. Alternative splicing of a novel diacylglycerol kinase in tomato leads to a calmodulin-binding isoform. Plant J. 2000, 24, 317–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez-Merino, F.C.; Brearley, C.A.; Ornatowska, M.; Abdel-Haliem, M.E.F.; Zanor, M.-I.; Mueller-Roeber, B. AtDGK2, a Novel Diacylglycerol Kinase from Arabidopsis thaliana, Phosphorylates 1-Stearoyl-2-arachidonoyl-sn-glycerol and 1,2-Dioleoyl-sn-glycerol and Exhibits Cold-inducible Gene Expression. J. Biol. Chem. 2003, 279, 8230–8241. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.H.; Henderson, D.A.; Zhu, J.K. The Arabidopsis cold-responsive transcriptome and its regulation by ICE1. Plant Cell 2005, 17, 3155–3175. [Google Scholar] [CrossRef]
- Li, Y.; Tan, Y.; Shao, Y.; Li, M.; Ma, F. Comprehensive genomic analysis and expression profiling of diacylglycerol kinase gene family in Malus prunifolia (Wild.) Borkh. Gene 2015, 561, 225–234. [Google Scholar] [CrossRef] [PubMed]
- Schmutz, J.; Cannon, S.B.; Schlueter, J.; Ma, J.; Mitros, T.; Nelson, W.; Hyten, D.L.; Song, Q.; Thelen, J.J.; Cheng, J.; et al. Genome sequence of the palaeopolyploid soybean. Nature 2010, 463, 178–183. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Zhao, C.; He, L.; Yan, B.; Dong, J.; Li, Z.; Xu, J. Genome-wide identification and abiotic stress responses of DGK gene family in maize. J. Plant Biochem. Biotechnol. 2018, 27, 156–166. [Google Scholar] [CrossRef]
- Houssa, B.; van Blitterswijk, W.J. Specificity of cysteine-rich domains in diacylglycerol kinases and protein kinases C. Biochem. J. 1998, 331, 677–679. [Google Scholar] [CrossRef] [PubMed]
- Pears, C.J.; Kour, G.; House, C.; Kemp, B.E.; Parker, P.J. Mutagenesis of the pseudosubstrate site of protein kinase C leads to activation. Eur. J. Biochem. 1990, 194, 89–94. [Google Scholar] [CrossRef] [Green Version]
- Pavlopoulos, G.A.; Soldatos, T.G.; Barbosa-Silva, A.; Schneider, R. A reference guide for tree analysis and visualization. BioData Min. 2010, 3. [Google Scholar] [CrossRef]
- Law, Y.-S.; Gudimella, R.; Song, B.-K.; Ratnam, W.; Harikrishna, J.A. Molecular Characterization and Comparative Sequence Analysis of Defense-Related Gene, Oryza rufipogon Receptor-Like Protein Kinase 1. Int. J. Mol. Sci. 2012, 13, 9343–9362. [Google Scholar] [CrossRef]
- Mao, J.; Manik, S.; Shi, S.; Chao, J.; Jin, Y.; Wang, Q.; Liu, H. Mechanisms and Physiological Roles of the CBL-CIPK Networking System in Arabidopsis thaliana. Genes 2016, 7, 62. [Google Scholar] [CrossRef]
- Fan, X.-D.; Wang, J.-Q.; Yang, N.; Dong, Y.-Y.; Liu, L.; Wang, F.-W.; Wang, N.; Chen, H.; Liu, W.; Sun, Y.; et al. Gene expression profiling of soybean leaves and roots under salt, saline–alkali and drought stress by high-throughput Illumina sequencing. Gene 2013, 512, 392–402. [Google Scholar] [CrossRef] [PubMed]
- Hendrix, K.W.; Assefa, H.; Boss, W.F. The polyphosphoinositides, phosphatidylinositol monophosphate and phosphatidylinositol bisphosphate, are present in nuclei isolated from carrot protoplast. Protoplasma 1989, 151, 62–72. [Google Scholar] [CrossRef]
- Lundberg, G.A.; Sommarin, M. Diacylglycerol kinase in plasma membranes from wheat. Biochim. Biophys. Acta (BBA) Lipids Lipid Metab. 1992, 1123, 177–183. [Google Scholar] [CrossRef]
- Tan, Z.; Boss, W.F. Association of Phosphatidylinositol Kinase, Phosphatidylinositol Monophosphate Kinase, and Diacylglycerol Kinase with the Cytoskeleton and F-Actin Fractions of Carrot (Daucus carota L.) Cells Grown in Suspension Culture: Response to Cell Wall-Degrading Enzymes. Plant Physiol. 1992, 100, 2116–2120. [Google Scholar] [CrossRef] [PubMed]
- Testerink, C.; Munnik, T. Phosphatidic acid: A multifunctional stress signaling lipid in plants. Trends Plant Sci. 2005, 10, 368–375. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Su, Y.; Liu, Y.; Kim, S.C.; Fanella, B. Phosphatidic acid as lipid messenger and growth regulators in plants. In Phospholipases in Plant Signalling; Springer: Berlin/Heidelberg, Germany, 2014; pp. 69–92. [Google Scholar]
- Peters, C.; Kim, S.C.; Devaiah, S.; Li, M.; Wang, X. Non-specific phospholipase C5 and diacylglycerol promote lateral root development under mild salt stress in Arabidopsis. Plant Cell Environ. 2014, 37, 2002–2013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Escobar-Sepulveda, H.F.; Trejo-Tellez, L.I.; Perez-Rodriguez, P.; Hidalgo Contreras, J.V.; Gomez-Merino, F.C. Diacylglycerol kinases are widespread in higher plants and display inducible gene expression in response to beneficial elements, metal, and metalloid ions. Front. Plant Sci. 2017, 8, 129. [Google Scholar] [CrossRef] [PubMed]
- Ha, C.V.; Le, D.T.; Nishiyama, R.; Watanabe, Y.; Tran, U.T.; Dong, N.V.; Tran, L.S.P. Characterization of the newly developed soybean cultivar DT2008 in relation to the model variety W82 reveals a new genetic resource for comparative and functional genomics for improved drought tolerance. BioMed Res. Int. 2013, 759657. [Google Scholar] [CrossRef] [PubMed]
- Ha, C.V.; Watanabe, Y.; Tran, U.T.; Le, D.T.; Tanaka, M.; Nguyen, K.H.; Seki, M.; Nguyen, D.V.; Tran, L.-S.P. Comparative analysis of root transcriptomes from two contrasting drought-responsive Williams 82 and DT2008 soybean cultivars under normal and dehydration conditions. Front. Plant Sci. 2015, 6, 551. [Google Scholar] [CrossRef]
- Vaultier, M.-N.; Cantrel, C.; Guerbette, F.; Boutté, Y.; Vergnolle, C.; Çiçek, D.; Bolte, S.; Zachowski, A.; Ruelland, E. The hydrophobic segment of Arabidopsis thaliana cluster I diacylglycerol kinases is sufficient to target the proteins to cell membranes. FEBS Lett. 2008, 582, 1743–1748. [Google Scholar] [CrossRef]
- Ge, H.; Chen, C.; Jing, W.; Zhang, Q.; Wang, H.; Wang, R.; Zhang, W. The Rice Diacylglycerol Kinase Family: Functional Analysis Using Transient RNA Interference. Front. Plant Sci. 2012, 3, 60. [Google Scholar] [CrossRef]
- Lynch, M.; O’Hely, M.; Walsh, B.; Force, A. The probability of preservation of a newly arisen gene duplicate. Genetics 2001, 159, 1789–1804. [Google Scholar] [PubMed]
- Qing, D.-J.; Lu, H.-F.; Li, N.; Dong, H.-T.; Dong, D.-F.; Li, Y.-Z. Comparative Profiles of Gene Expression in Leaves and Roots of Maize Seedlings under Conditions of Salt Stress and the Removal of Salt Stress. Plant Cell Physiol. 2009, 50, 889–903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinclair, T.; Ludlow, M. Influence of Soil Water Supply on the Plant Water Balance of Four Tropical Grain Legumes. Aust. J. Plant Physiol. 1986, 13, 329. [Google Scholar] [CrossRef]
- Zgallai, H.; Steppe, K.; Lemeur, R. Photosynthetic, Physiological and Biochemical Responses of Tomato Plants to Polyethylene Glycol-Induced Water Deficit. J. Integr. Plant Biol. 2005, 47, 1470–1478. [Google Scholar] [CrossRef]
- Sharma, K.K.; Sreelatha, G.; Dayal, S. Pigeonpea (Cajanus cajan L. Millsp.). Agrobact. Protoc. 2006, 359. [Google Scholar] [CrossRef]
- Yuan, S.; Liu, W.-J.; Zhang, N.-H.; Wang, M.-B.; Liang, H.-G.; Hui Lin, H. Effects of water stress on major photosystem II gene expression and protein metabolism in barley leaves. Physiol. Plant. 2005, 125, 464–473. [Google Scholar] [CrossRef]
- Guo, R.; Shi, L.; Yan, C.; Zhong, X.; Gu, F.; Liu, Q.; Xia, X.; Li, H. Ionomic and metabolic responses to neutral salt or alkaline salt stresses in maize (Zea mays L.) seedlings. BMC Plant Biol. 2017, 17, 41. [Google Scholar] [CrossRef]
- Van Baal, J.; de Widt, J.; Divecha, N.; van Blitterswijk, W.J. Translocation of diacylglycerol kinase theta from cytosol to plasma membrane in response to activation of G protein-coupled receptors and protein kinase C. J. Biol. Chem. 2005, 280, 9870–9878. [Google Scholar] [CrossRef]
- Shirai, Y.; Segawa, S.; Kuriyama, M.; Goto, K.; Sakai, N.; Saito, N. Subtype-specific translocation of diacylglycerol kinase alpha and gamma and its correlation with protein kinase C. J. Biol. Chem. 2000, 275, 24760–24766. [Google Scholar] [CrossRef]
- Hobo, T.; Asada, M.; Kowyama, Y.; Hattori, T. ACGT-containing abscisic acid response element (ABRE) and coupling element 3 (CE3) are functionally equivalent. Plant J. 1999, 19, 679–689. [Google Scholar] [CrossRef] [Green Version]
- Xiong, L.; Schumaker, K.S.; Zhu, J.-K. Cell Signaling during Cold, Drought, and Salt Stress. Plant Cell 2002, 14 (Suppl. 1), S165–S183. [Google Scholar] [CrossRef]
- Jiang, C.; Iu, B.; Singh, J. The requirement of a CCGAC cis-acting element for cold induction of the BN115 gene from winter Brassica napus. Plant Mol. Biol. 1996, 30, 679–684. [Google Scholar] [CrossRef]
- Rieping, M.; Schoffl, F. Synergistic effect of upstream sequences, CCAAT box elements, and HSE sequences for enhanced expression of chimeric heat shock genes in transgenic tobacco. Mol. Gen. Genet. 1992, 231, 226–232. [Google Scholar]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular Evolutionary Genetics Analysis Using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony Methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [Green Version]
- Hu, B.; Jin, J.; Guo, A.-Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2014, 31, 1296–1297. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, L.; Thilmony, R.; You, F.M.; Gu, Y.Q.; Coleman-Derr, D. PIECE 2.0: An update for the plant gene structure comparison and evolution database. Nucleic Acids Res. 2016, 45, 1015–1020. [Google Scholar] [CrossRef]
- Liu, W.; Xie, Y.; Ma, J.; Luo, X.; Nie, P.; Zuo, Z.; Lahrmann, U.; Zhao, Q.; Zheng, Y.; Zhao, Y.; et al. IBS: An illustrator for the presentation and visualization of biological sequences. Bioinformatics 2015, 31, 3359–3361. [Google Scholar] [CrossRef]
- Hall, B.G. Building phylogenetic trees from molecular data with MEGA. Mol. Biol. Evol. 2013, 30, 1229–1235. [Google Scholar] [CrossRef]
- Gupta, R.S.; Epand, R.M. Phylogenetic analysis of the diacylglycerol kinase family of proteins and identification of multiple highly-specific conserved inserts and deletions within the catalytic domain that are distinctive characteristics of different classes of DGK homologs. PLoS ONE 2017, 12, e0182758. [Google Scholar] [CrossRef]
- Higo, K.; Ugawa, Y.; Iwamoto, M.; Korenaga, T. Plant cis-acting regulatory DNA elements (PLACE) database: 1999. Nucleic Acids Res. 1999, 27, 297–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, F.; Deng, Y.; Zhou, Y.; Dong, J.; Chen, H.; Dong, Y.; Wang, N.; Li, X.; Li, H. Genome-Wide Analysis and Expression Profiling of the Phospholipase C Gene Family in Soybean (Glycine max). PLoS ONE 2015, 10, e0138467. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Pandey, G.K. Primer design using Primer Express for SYBR Green-based quantitative PCR. Methods Mol. Biol. 2015, 1275, 153–164. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2–ΔΔCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Gene Locus | Gene Identifier | Chr | Locations | Length (bp) | Proteins | |||
|---|---|---|---|---|---|---|---|---|---|
| Protein ID | Length (aa) | pI | (kDa) | ||||||
| GmDGK1 | LOC100813110 | Glyma.13G302200 | 13 | 39928623-39934694 | 1185 | XP 003541921.1 | 436 | 6.77 | 48.62 |
| GmDGK2 | LOC100797649 | Glyma.05G196100 | 5 | 38025457-38031403 | 2178 | XP_006580353.1 | 725 | 7.05 | 78.6 |
| GmDGK3 | LOC100813809 | Glyma.12G200100 | 12 | 36142043-36148329 | 1467 | XP_014632628 | 480 | 5.59 | 53.7 |
| GmDGK4 | LOC100817198 | Glyma.06G299200 | 6 | 48769786-48776248 | 1456 | XP_003526295 | 484 | 5.29 | 54.29 |
| GmDGK5 | LOC100811505 | Glyma.06G223900 | 6 | 29792271-29817479 | 1443 | XP_014632628 | 480 | 6.36 | 53.76 |
| GmDGK6 | LOC100794923 | Glyma.04G143000 | 4 | 25618714-25643610 | 1458 | XP_003552048 | 485 | 6.36 | 54.2 |
| GmDGK7 | LOC100811680 | Glyma.13G093100 | 13 | 20812774-20820984 | 1443 | XP_003542286 | 480 | 7.61 | 53.54 |
| GmDGK8 | LOC100816144 | Glyma.06G254900 | 6 | 43416272-43423748 | 1452 | XP_014632201 | 504 | 7.29 | 56.32 |
| GmDGK9 | LOC100801004 | Glyma.12G146700 | 12 | 20308574-20316372 | 2044 | XP_006592585 | 485 | 6.58 | 54.35 |
| GmDGK10 | LOC100808353 | Glyma.17G067400 | 17 | 5211623-5219147 | 1874 | XP_014625529.3 | 480 | 7.46 | 53.56 |
| GmDGK11 | LOC100794891 | Glyma.17G077100 | 17 | 6032535-6039406 | 2184 | XP_014625346.1 | 727 | 6.04 | 80.97 |
| GmDGK12 | LOC100802426 | Glyma.05G022500 | 5 | 1971267-1978461 | 2184 | XP_006579516 | 727 | 6 | 80.93 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carther, K.F.I.; Ketehouli, T.; Ye, N.; Yang, Y.-H.; Wang, N.; Dong, Y.-Y.; Yao, N.; Liu, X.-M.; Liu, W.-C.; Li, X.-W.; et al. Comprehensive Genomic Analysis and Expression Profiling of Diacylglycerol Kinase (DGK) Gene Family in Soybean (Glycine max) under Abiotic Stresses. Int. J. Mol. Sci. 2019, 20, 1361. https://doi.org/10.3390/ijms20061361
Carther KFI, Ketehouli T, Ye N, Yang Y-H, Wang N, Dong Y-Y, Yao N, Liu X-M, Liu W-C, Li X-W, et al. Comprehensive Genomic Analysis and Expression Profiling of Diacylglycerol Kinase (DGK) Gene Family in Soybean (Glycine max) under Abiotic Stresses. International Journal of Molecular Sciences. 2019; 20(6):1361. https://doi.org/10.3390/ijms20061361
Chicago/Turabian StyleCarther, Kue Foka Idrice, Toi Ketehouli, Nan Ye, Yan-Hai Yang, Nan Wang, Yuan-Yuan Dong, Na Yao, Xiu-Ming Liu, Wei-Can Liu, Xiao-Wei Li, and et al. 2019. "Comprehensive Genomic Analysis and Expression Profiling of Diacylglycerol Kinase (DGK) Gene Family in Soybean (Glycine max) under Abiotic Stresses" International Journal of Molecular Sciences 20, no. 6: 1361. https://doi.org/10.3390/ijms20061361