Development of SNP, KASP, and SSR Markers by BSR-Seq Technology for Saturation of Genetic Linkage Map and Efficient Detection of Wheat Powdery Mildew Resistance Gene Pm61

 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Evaluation of Resistance to Bgt Isolates in XXSYH
2.2. Genetic Analysis of Powdery Mildew Resistance in XXSYH
2.3. BSR-Seq Analysis of the Bulked RNA Pools with Distinct Phenotypes to Isolate Bgt1
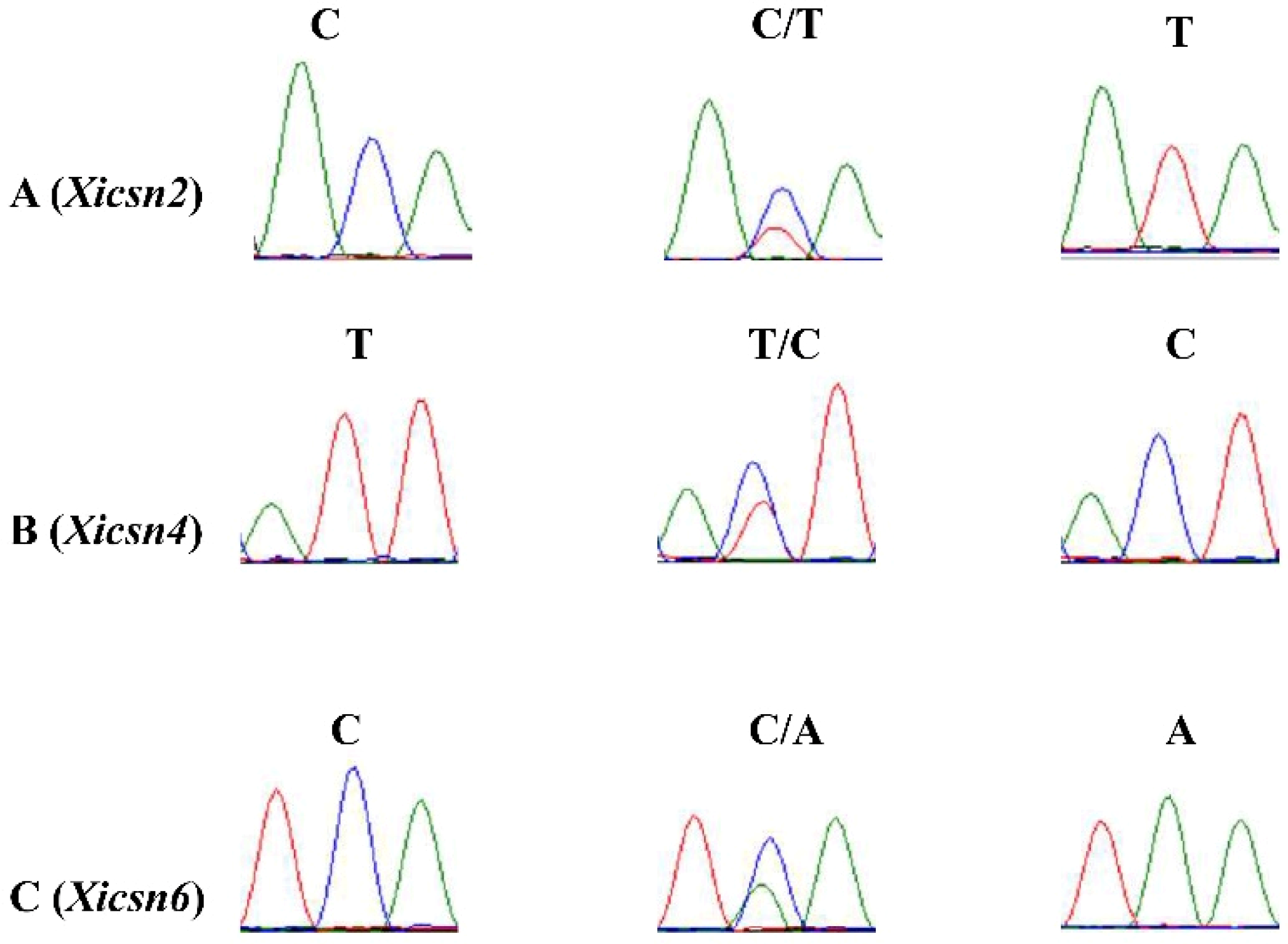
2.4. Validation of the Candidate SNP and Development of SNP Markers
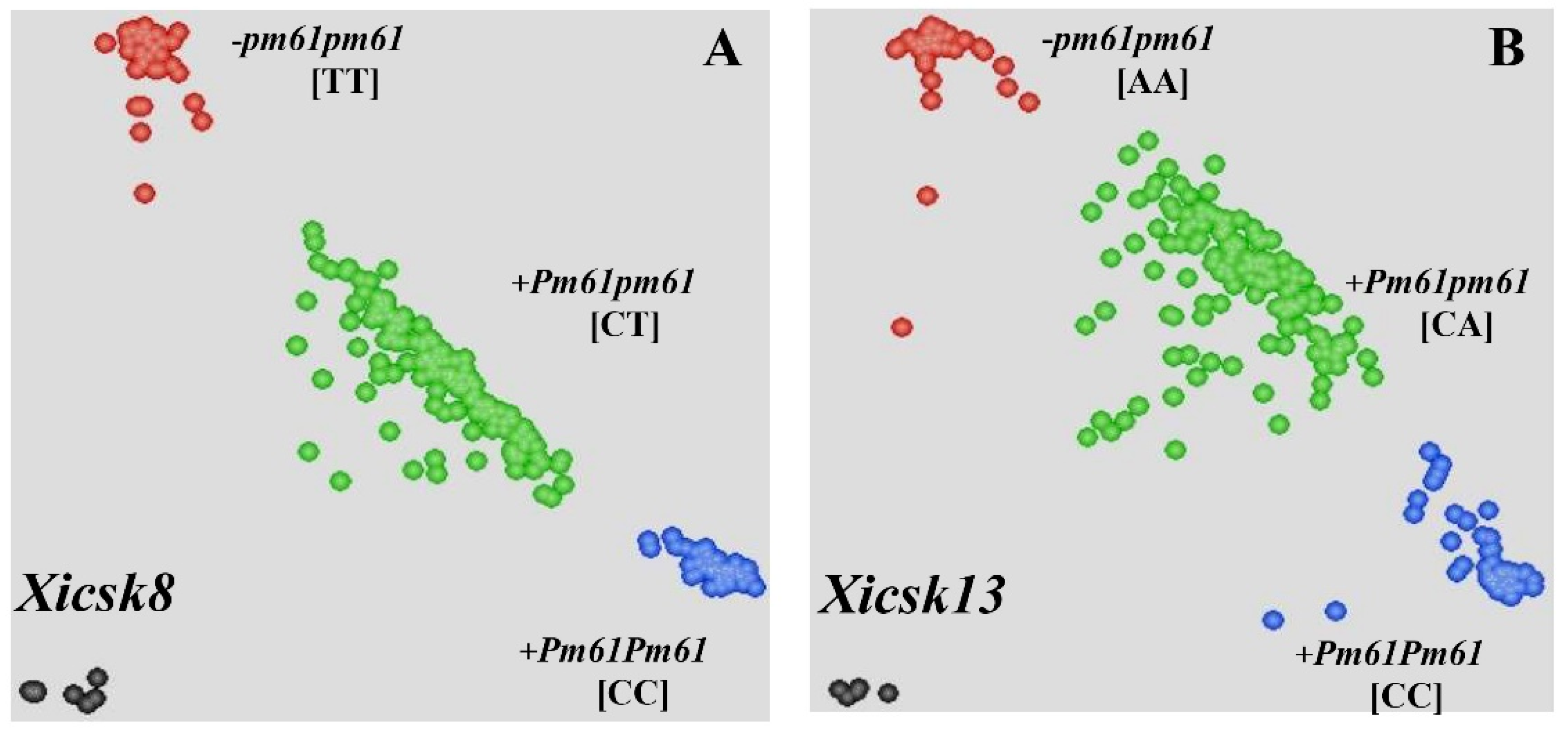
2.5. Development of KASP Markers
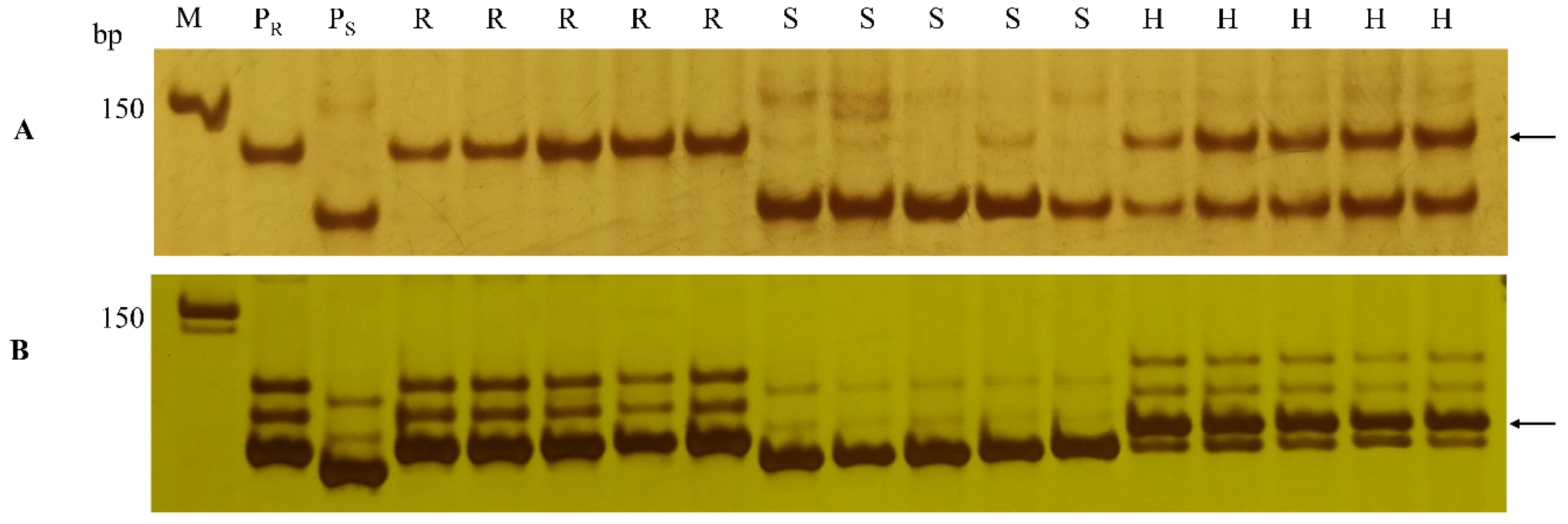
2.6. Development of SSR Markers
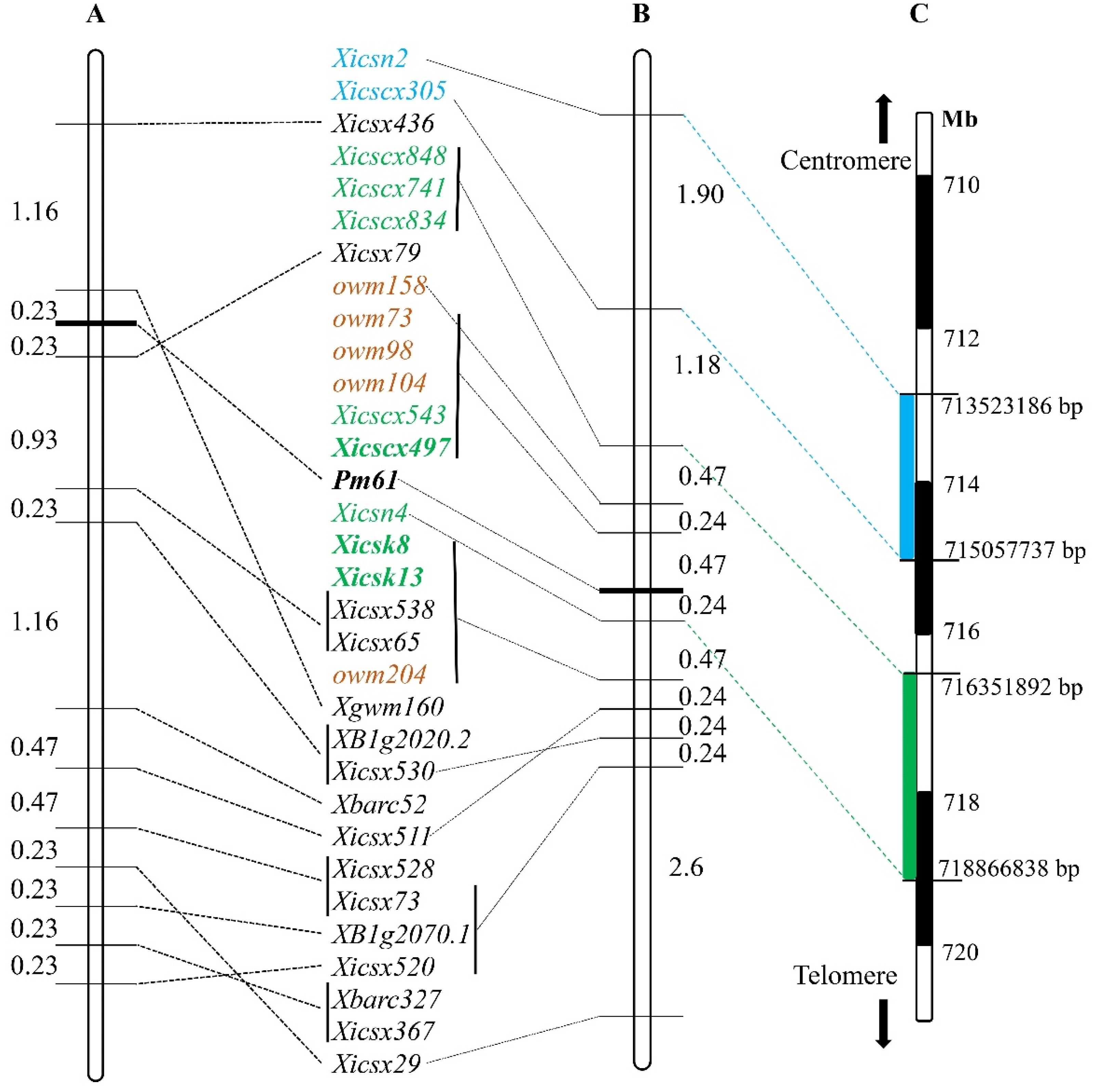
2.7. Construction of the Genetic Linkage Map for Pm61
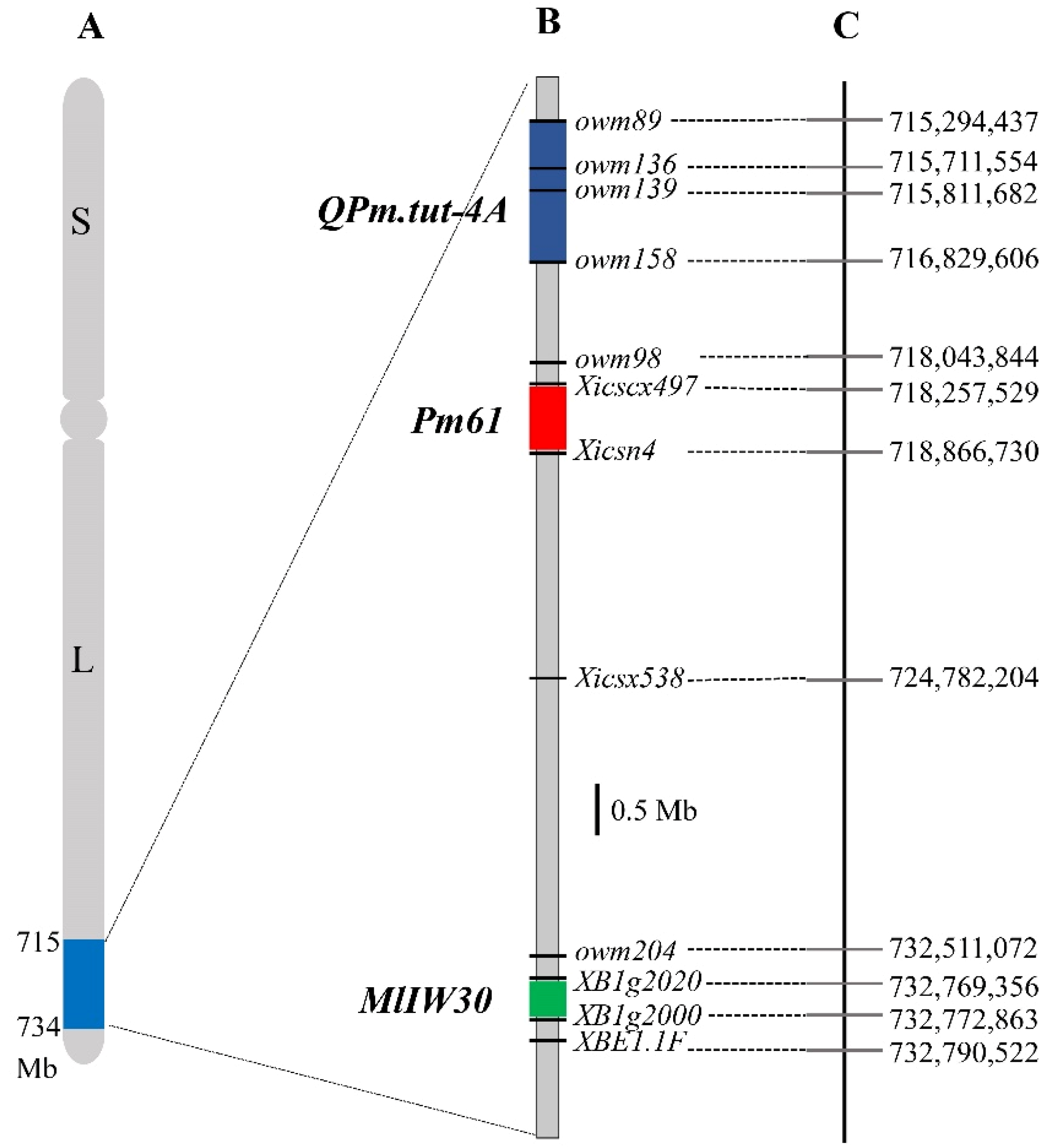
2.8. Physical Locations of Pm61, QPm.tut-4A and MlIW30 on Chromosome Arm 4AL
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Assessments of Resistance to Powdery Mildew
4.3. BSR-Seq Analysis
4.4. DNA Isolation, Amplification and Electrophoresis
4.5. Development and Validation of SNP Markers
4.6. Conversion of SNP Markers to KASP Markers
4.7. Development of SSR Markers
4.8. Construction of High-Density Genetic Linkage Map
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Singh, R.P.; Singh, P.K.; Rutkoski, J.; Hodson, D.P.; He, X.; Jørgenssen, L.N.; Huertaespino, J. Disease impact on wheat yield potential and prospects of genetic control. Annu. Rev. Phytopathol. 2016, 54, 303–322. [Google Scholar] [CrossRef] [PubMed]
- Samobor, V.; Vukobratović, M.; Jošt, M. Effect of powdery mildew attack on quality parameters and experimental bread baking of wheat. Acta Agric. Slov. 2006, 87, 381–391. [Google Scholar]
- Li, J.; Liu, X.H.; Yang, X.W.; Li, Y.C.; Wang, C.Y.; He, D.X. Proteomic analysis of the impacts of powdery mildew on wheat grain. Food Chem. 2018, 261, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Hsam, S.L.K.; Huang, X.Q.; Zeller, F.J. Chromosomal location of genes for resistance to powdery mildew in common wheat (Triticum aestivum L. em. Thell.). 6. Alleles at the Pm5 locus. Theor. Appl. Genet. 2001, 102, 127–133. [Google Scholar] [CrossRef]
- Huang, X.Q.; Wang, L.X.; Xu, M.X.; Röder, M.S. Microsatellite mapping of the powdery mildew resistance gene Pm5e in common wheat (Triticum aestivum L.). Theor. Appl. Genet. 2003, 106, 858–865. [Google Scholar] [CrossRef] [PubMed]
- Zhou, R.H.; Zhu, Z.D.; Kong, X.Y.; Huo, N.X.; Tian, Q.Z.; Li, P.; Jin, C.Y.; Dong, Y.C.; Jia, J.Z. Development of wheat near-isogenic lines for powdery mildew resistance. Theor. Appl. Genet. 2005, 110, 640–648. [Google Scholar] [CrossRef] [PubMed]
- Hu, T.Z.; Li, H.J.; Xie, C.J.; You, M.S.; Yang, Z.M.; Sun, Q.X.; Liu, Z.Y. Molecular mapping and chromosomal location of powdery mildew resistance gene in wheat cultivar Tangmai 4. Acta Agron. Sin. 2008, 34, 1193–1198. [Google Scholar]
- Zhai, W.W.; Duan, X.Y.; Zhou, Y.L.; Ma, H.Q. Inheritance of resistance to powdery mildew in four Chinese landraces. Plant Protect 2008, 34, 37–40. [Google Scholar]
- Xue, F.; Zhai, W.W.; Duan, X.Y.; Zhou, Y.L.; Ji, W.Q. Microsatellite mapping of powdery mildew resistance gene in wheat landrace Xiaobaidong. Acta Agron. Sin. 2009, 35, 1806–1811. [Google Scholar] [CrossRef]
- Fu, B.S.; Zhang, Z.L.; Zhang, Q.F.; Wu, X.Y.; Wu, J.Z.; Cai, S.B. Identification and mapping of a new powdery mildew resistance allele in the Chinese wheat landrace Hongyoumai. Mol. Breed. 2017, 37, 133. [Google Scholar] [CrossRef]
- Xu, X.D.; Jing, F.; Fan, J.R.; Liu, Z.Y.; Li, Q.; Zhou, Y.L.; Ma, Z.H. Identification of the resistance gene to powdery mildew in Chinese wheat landrace Baiyouyantiao. J. Integr. Agric. 2018, 17, 37–45. [Google Scholar] [CrossRef]
- Xu, X.D.; Li, Q.; Ma, Z.H.; Fan, J.R.; Zhou, Y.L. Molecular mapping of powdery mildew resistance gene PmSGD in Chinese wheat landrace Shangeda using RNA-seq with bulk segregant analysis. Mol. Breed. 2018, 38, 23. [Google Scholar] [CrossRef]
- Huang, X.Q.; Hsam, S.L.K.; Zeller, F.J.; Wenzel, G.; Mohler, V. Molecular mapping of the wheat powdery mildew resistance gene Pm24 and marker validation for molecular breeding. Theor. Appl. Genet. 2000, 101, 407–414. [Google Scholar] [CrossRef]
- Xue, F.; Wang, C.Y.; Li, C.; Duan, X.Y.; Zhou, Y.L.; Zhao, N.J.; Wang, Y.J.; Ji, W.Q. Molecular mapping of a powdery mildew resistance gene in common wheat landrace Baihulu and its allelism with Pm24. Theor. Appl. Genet. 2012, 125, 1425–1432. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.X.; Yi, Y.J.; Ma, P.T.; Qie, Y.M.; Fu, X.Y.; Xu, Y.F.; Zhang, X.T.; An, D.G. Molecular tagging of a new broad-spectrum powdery mildew resistance allele Pm2c in Chinese wheat landrace Niaomai. Theor. Appl. Genet. 2015, 128, 2077–2084. [Google Scholar] [CrossRef]
- Ma, H.Q.; Kong, Z.X.; Fu, B.S.; Li, N.; Zhang, L.X.; Jia, H.Y.; Ma, Z.Q. Identification and mapping of a new powdery mildew resistance gene on chromosome 6D of common wheat. Theor. Appl. Genet. 2011, 123, 1099–1106. [Google Scholar] [CrossRef] [PubMed]
- Xiao, M.G.; Song, F.J.; Jiao, J.F.; Wang, X.M.; Xu, H.X.; Li, H.J. Identification of the gene Pm47 on chromosome 7BS conferring resistance to powdery mildew in the Chinese wheat landrace Hongyanglazi. Theor. Appl. Genet. 2013, 126, 1397–1403. [Google Scholar] [CrossRef]
- Fu, B.S.; Chen, Y.; Li, N.; Ma, H.Q.; Kong, Z.X.; Zhang, L.X.; Jia, H.Y.; Ma, Z.Q. pmX: A recessive powdery mildew resistance gene at the Pm4 locus identified in wheat landrace Xiaohongpi. Theor. Appl. Genet. 2013, 126, 913–921. [Google Scholar] [CrossRef]
- Wang, Z.Z.; Li, H.W.; Zhang, D.Y.; Guo, L.; Chen, J.J.; Chen, Y.X.; Wu, Q.H.; Xie, J.Z.; Zhang, Y.; Sun, Q.X.; et al. Genetic and physical mapping of powdery mildew resistance gene MlHLT in Chinese wheat landrace Hulutou. Theor. Appl. Genet. 2015, 128, 365–373. [Google Scholar] [CrossRef]
- Li, G.; Carver, B.F.; Cowger, C.; Bai, G.; Xu, X. Pm223899, a new recessive powdery mildew resistance gene identified in Afghanistan landrace PI 223899. Theor. Appl. Genet. 2018, 131, 2775–2783. [Google Scholar] [CrossRef]
- Tan, C.C.; Li, G.Q.; Cowger, C.; Carver, B.F.; Xu, X.Y. Characterization of Pm63, a powdery mildew resistance gene in Iranian landrace PI 628024. Theor. Appl. Genet. 2018. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.C.; Li, G.Q.; Cowger, C.; Carver, B.F.; Xu, X.Y. Characterization of Pm59, a novel powdery mildew resistance gene in Afghanistan wheat landrace PI 181356. Theor. Appl. Genet. 2018, 131, 1145–1152. [Google Scholar] [CrossRef] [PubMed]
- Newton, A.C.; Akar, T.; Baresel, J.P.; Bebeli, P.J.; Bettencourt, E.; Blandenopoulos, K.V.; Czembor, J.H.; Fasoula, D.A.; Katsiotis, A.; Koutis, K.; et al. Cereal landraces for sustainable agriculture. A review. Agron. Sustain. Dev. 2010, 20, 237–269. [Google Scholar] [CrossRef]
- Kaur, N.; Street, K.; Mackay, M.; Yahiaoui, N.; Keller, B. Molecular approaches for characterization and use of natural disease resistance in wheat. Eur. J. Plant Pathol. 2008, 121, 387–397. [Google Scholar] [CrossRef] [Green Version]
- International Wheat Genome Sequencing Consortium (IWGSC). Shifting the limits in wheat research and breeding using a fully annotated reference genome. Science 2018, 361, eaar7191. [Google Scholar] [CrossRef] [PubMed]
- Rasheed, A.; Wen, W.; Gao, F.; Zhai, S.; Jin, H.; Liu, J.; Guo, Q.; Zhang, Y.; Dreisigacker, S.; Xia, X.; et al. Development and validation of KASP assays for genes underpinning key economic traits in bread wheat. Theor. Appl. Genet. 2016, 129, 1843–1860. [Google Scholar] [CrossRef] [PubMed]
- Qureshi, N.; Kandiah, P.; Gessese, M.K.; Nsabiyera, V.; Wells, V.; Babu, P.; Wong, D.; Hayden, M.J.; Bariana, H.; Bansal, U. Development of co-dominant KASP markers co-segregating with Ug99 effective stem rust resistance gene Sr26 in wheat. Mol. Breed. 2018, 38, 97. [Google Scholar] [CrossRef]
- Qureshi, N.; Bariana, H.S.; Zhang, P.; McIntosh, R.; Bansal, U.K.; Wong, D.; Hayden, M.J.; Dubcovsky, J.; Shankar, M. Genetic relationship of stripe rust resistance genes Yr34 and Yr48 in wheat and identification of linked KASP markers. Plant Dis. 2018, 102, 413–420. [Google Scholar] [CrossRef]
- Trick, M.; Adamski, N.M.; Mugford, S.G.; Jiang, C.; Febrer, M.; Uauy, C. Combining SNP discovery from next-generation sequencing data with bulked segregant analysis (BSA) to fine-map genes in polyploid wheat. BMC Plant Biol. 2012, 12, 14. [Google Scholar] [CrossRef]
- Ramirez-Gonzalez, R.H.; Segovia, V.; Bird, N.; Fenwick, P.; Holdgate, S.; Berry, S.; Jack, P.; Caccamo, M.; Uauy, C. RNA-Seq bulked segregant analysis enables the identification of high-resolution genetic markers for breeding in hexaploid wheat. Plant Biotechnol. J. 2015, 13, 613–624. [Google Scholar] [CrossRef]
- Wang, Y.; Xie, J.Z.; Zhang, H.Z.; Guo, B.M.; Ning, S.Z.; Chen, Y.X.; Lu, P.; Wu, Q.H.; Li, M.M.; Zhang, D.Y.; et al. Mapping stripe rust resistance gene YrZH22 in Chinese wheat cultivar Zhoumai 22 by bulked segregant RNA-Seq (BSR-Seq) and comparative genomics analyses. Theor. Appl. Genet. 2017, 130, 2191–2201. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, H.Z.; Xie, J.Z.; Guo, B.M.; Chen, Y.X.; Zhang, H.Y.; Lu, P.; Wu, Q.H.; Li, M.M.; Zhang, D.Y.; et al. Mapping stripe rust resistance genes by BSR-Seq, YrMM58 and YrHY1 on chromosome 2AS in Chinese wheat lines Mengmai 58 and Huaiyang 1 are Yr17. Crop J. 2018, 6, 91–98. [Google Scholar] [CrossRef]
- Wu, J.H.; Zeng, Q.D.; Wang, Q.L.; Liu, S.J.; Yu, S.Z.; Mu, J.M.; Huang, S.; Sela, H.; Distelfeld, A.; Huang, L.L.; et al. SNP-based pool genotyping and haplotype analysis accelerate fine-mapping of the wheat genomic region containing stripe rust resistance gene Yr26. Theor. Appl. Genet. 2018, 131, 1481–1496. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.P.; Xie, J.Z.; Hu, J.H.; Qiu, D.; Liu, Z.Y.; Li, J.T.; Li, M.M.; Zhang, H.J.; Yang, L.; Liu, H.W.; et al. Development of molecular markers linked to powdery mildew resistance gene Pm4b by combining SNP discovery from transcriptome sequencing data with bulked segregant analysis (BSR-Seq) in wheat. Front. Plant Sci. 2018, 9, 95. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.G.; Hu, J.H.; Song, W.; Qiu, D.; Cui, L.; Wu, P.P.; Zhang, H.J.; Liu, H.W.; Yang, L.; Qu, Y.F.; et al. Pm61: A recessive gene for resistance to powdery mildew in wheat landrace Xuxusanyuehuang identified by comparative genomics analysis. Theor. Appl. Genet. 2018, 131, 2085–2097. [Google Scholar] [CrossRef] [PubMed]
- Geng, M.M.; Zhang, J.; Peng, F.X.; Liu, X.; Lv, X.D.; Mi, Y.Y.; Li, Y.H.; Li, F.; Xie, C.J.; Sun, Q.X. Identification and mapping of MlIW30, a novel powdery mildew resistance gene derived from wild emmer wheat. Mol. Breed. 2016, 36, 130. [Google Scholar] [CrossRef]
- Janáková, E.; Jakobson, I.; Peusha, H.; Abrouk, M.; Škopová, M.; Šimková, H.; Šafář, J.; Jan Vrána, J.; Doležel, J.; Järve, K.; et al. Divergence between bread wheat and Triticum militinae in the powdery mildew resistance QPm.tut-4A locus and its implications for cloning of the resistance gene. Theor. Appl. Genet. 2018. [Google Scholar] [CrossRef]
- Semagn, K.; Babu, R.; Hearne, S.; Olsen, M. Single nucleotide polymorphism genotyping using Kompetitive Allele Specific PCR (KASP): Overview of the technology and its application in crop improvement. Mol. Breed. 2014, 33, 1–14. [Google Scholar] [CrossRef]
- Allen, A.M.; Barker, G.L.; Wilkinson, P.; Burridge, A.; Winfield, M.; Coghill, J.; Uauy, C.; Griffiths, S.; Jack, P.; Berry, S.; et al. Discovery and development of exome-based, co-dominant single nucleotide polymorphism markers in hexaploid wheat (Triticum aestivum L.). Plant Biotechnol. J. 2013, 11, 279–295. [Google Scholar] [CrossRef]
- Liu, Z.; Sun, Q.; Ni, Z.; Yang, T.; McIntosh, R.A. Development of SCAR markers linked to the Pm21 gene conferring resistance to powdery mildew in common wheat. Plant Breed. 1999, 118, 215–219. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef] [PubMed]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.; Daly, M.; et al. The Genome Analysis Toolkit: A MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef] [PubMed]
- Saghai-Maroof, M.A.; Soliman, K.M.; Jorgensen, R.A.; Allard, R.W. Ribosomal DNA spacer-length polymorphisms in barley: Mendelian inheritance, chromosomal location, and population dynamics. Proc. Natl. Acad. Sci. USA 1984, 81, 8014–8018. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tiwari, V.K.; Rawat, N.; Gill, B.S.; Huo, N.X.; You, F.M.; Coleman-Derr, D.; Gu, Y.Q. GSP: A web-based platform for designing genome-specific primers in polyploids. Bioinformatics 2016, 32, 2382–2383. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Gonzalez, R.H.; Uauy, C.; Caccamo, M. PolyMarker: A fast polyploid primer design pipeline. Bioinformatics 2015, 31, 2038–2039. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.H.; Meng, J.L. MapDraw: A Microsoft excel macro for drawing genetic linkage maps based on given genetic linkage data. Hereditas 2003, 25, 317–321. [Google Scholar] [PubMed]
- Lincoln, S.E.; Daly, M.J.; Lander, E.S. Constructing Genetic Linkage Maps with MAPMAKER/EXP Version 3.0: A Tutorial and Reference Mannual, 3rd ed.; Whitehead Institute for Medical Research: Cambridge, MA, USA, 1993. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bgt Isolate | XXSYH | Zhongzuo 9504 | Province |
|---|---|---|---|
| 1 | 1 | 4 | Shandong |
| 2 | 1 | 3 | Shandong |
| 3 | 1 | 3 | Shandong |
| 4 | 2 | 3 | Shandong |
| 5 | 1 | 3 | Shandong |
| 6 | 2 | 3 | Shandong |
| 7 | 3 | 3 | Shandong |
| 8 | 3 | 4 | Shanxi |
| 9 | 2 | 3 | Shanxi |
| 10 | 0; | 4 | Shanxi |
| 11 | 3 | 3 | Shanxi |
| 12 | 2 | 3 | Shanxi |
| 13 | 1 | 3 | Beijing |
| 14 | 1 | 3 | Beijing |
| 15 | 1 | 3 | Beijing |
| 16 | 3 | 4 | Beijing |
| 17 | 1 | 3 | Hebei |
| 18 | 3 | 3 | Hebei |
| 19 | 0; | 3 | Hebei |
| 20 | 3 | 4 | Hebei |
| 21 | 3 | 3 | Hebei |
| 22 | 1 | 3 | Hebei |
| 23 | 1 | 3 | Hebei |
| 24 | 3 | 3 | Hebei |
| 25 | 1 | 3 | Hebei |
| 26 | 2 | 4 | Hebei |
| 27 | 3 | 3 | Hebei |
| 28 | 2 | 4 | Hebei |
| 29 | 3 | 3 | Sichuan |
| 30 | 1 | 4 | Sichuan |
| 31 | 2 | 3 | Sichuan |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, J.; Li, J.; Wu, P.; Li, Y.; Qiu, D.; Qu, Y.; Xie, J.; Zhang, H.; Yang, L.; Fu, T.; et al. Development of SNP, KASP, and SSR Markers by BSR-Seq Technology for Saturation of Genetic Linkage Map and Efficient Detection of Wheat Powdery Mildew Resistance Gene Pm61. Int. J. Mol. Sci. 2019, 20, 750. https://doi.org/10.3390/ijms20030750
Hu J, Li J, Wu P, Li Y, Qiu D, Qu Y, Xie J, Zhang H, Yang L, Fu T, et al. Development of SNP, KASP, and SSR Markers by BSR-Seq Technology for Saturation of Genetic Linkage Map and Efficient Detection of Wheat Powdery Mildew Resistance Gene Pm61. International Journal of Molecular Sciences. 2019; 20(3):750. https://doi.org/10.3390/ijms20030750
Chicago/Turabian StyleHu, Jinghuang, Jingting Li, Peipei Wu, Yahui Li, Dan Qiu, Yunfeng Qu, Jingzhong Xie, Hongjun Zhang, Li Yang, Tiantian Fu, and et al. 2019. "Development of SNP, KASP, and SSR Markers by BSR-Seq Technology for Saturation of Genetic Linkage Map and Efficient Detection of Wheat Powdery Mildew Resistance Gene Pm61" International Journal of Molecular Sciences 20, no. 3: 750. https://doi.org/10.3390/ijms20030750