Intestinal Anti-Inflammatory Activity of the Aqueous Extract from Ipomoea asarifolia in DNBS-Induced Colitis in Rats

 ,
,  , ,
, ,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
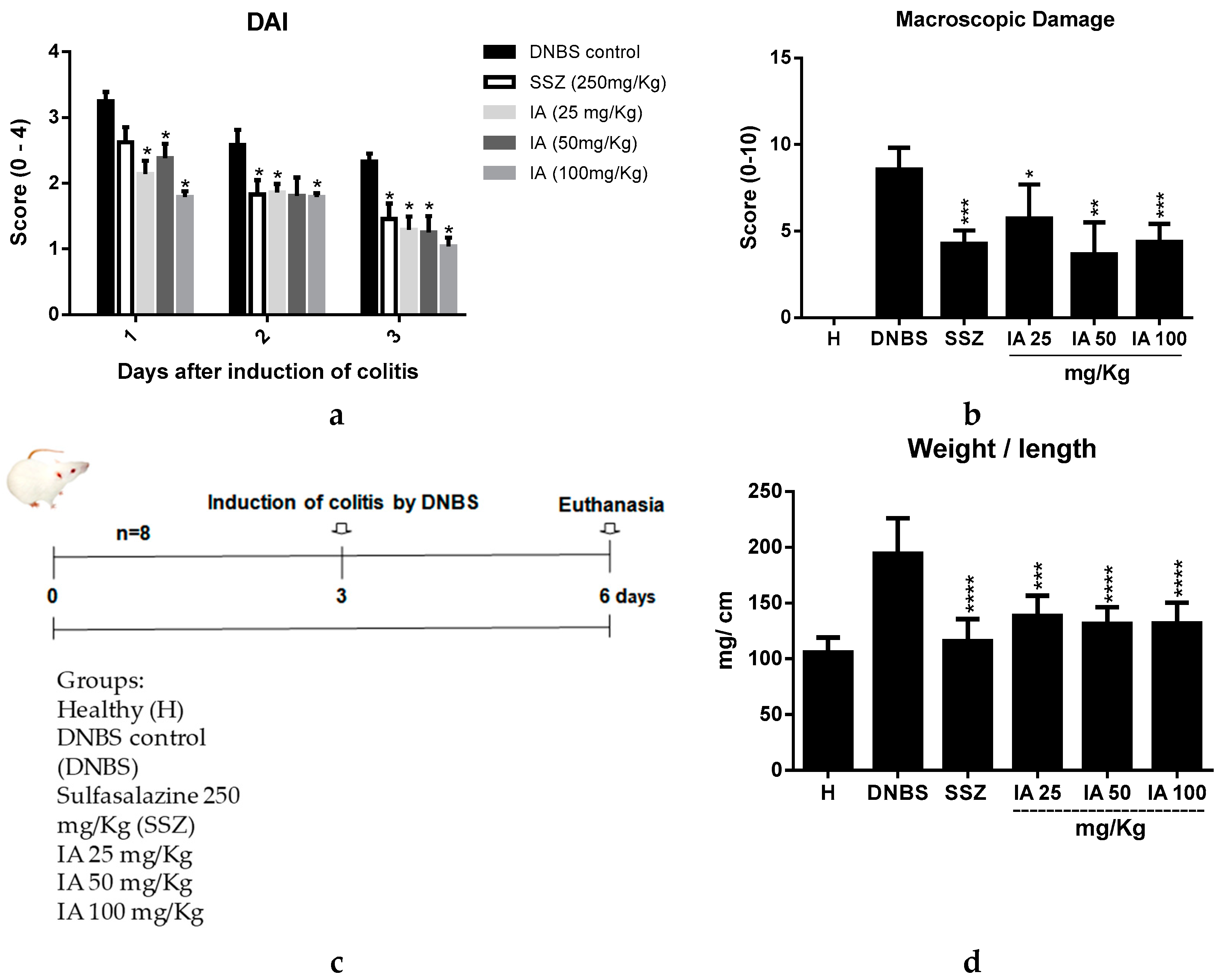
2.1. Effect of Ipomoea asarifolia Aqueous Extract on the Disease Activity Index, Macroscopic Score, and Weight/Colonic Length Relationship
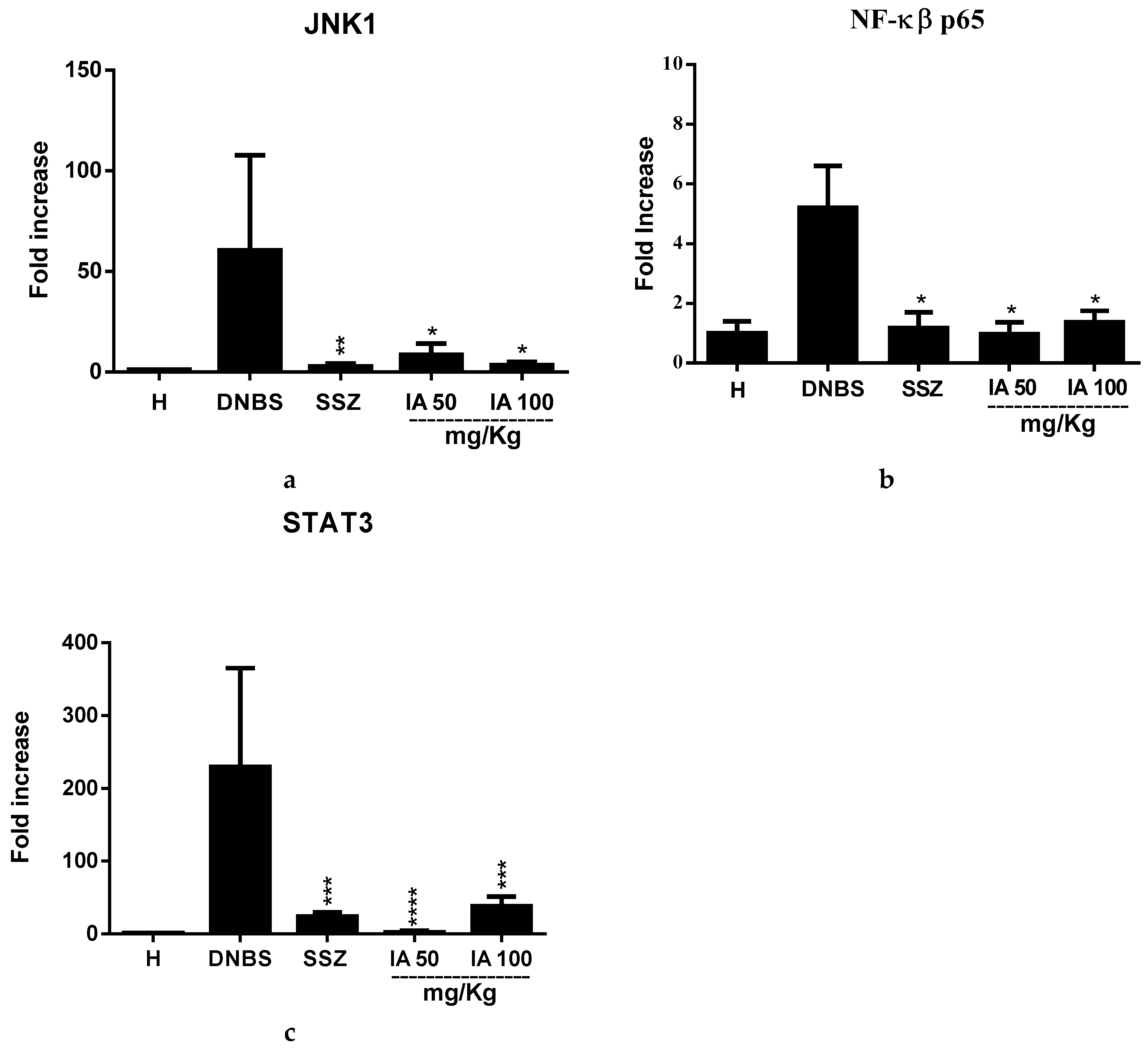
2.2. Effect of the Ipomoea asarifolia Aqueous Extract on the Gene Expresseion of JNK1, NK-κβ-p65, and STAT3
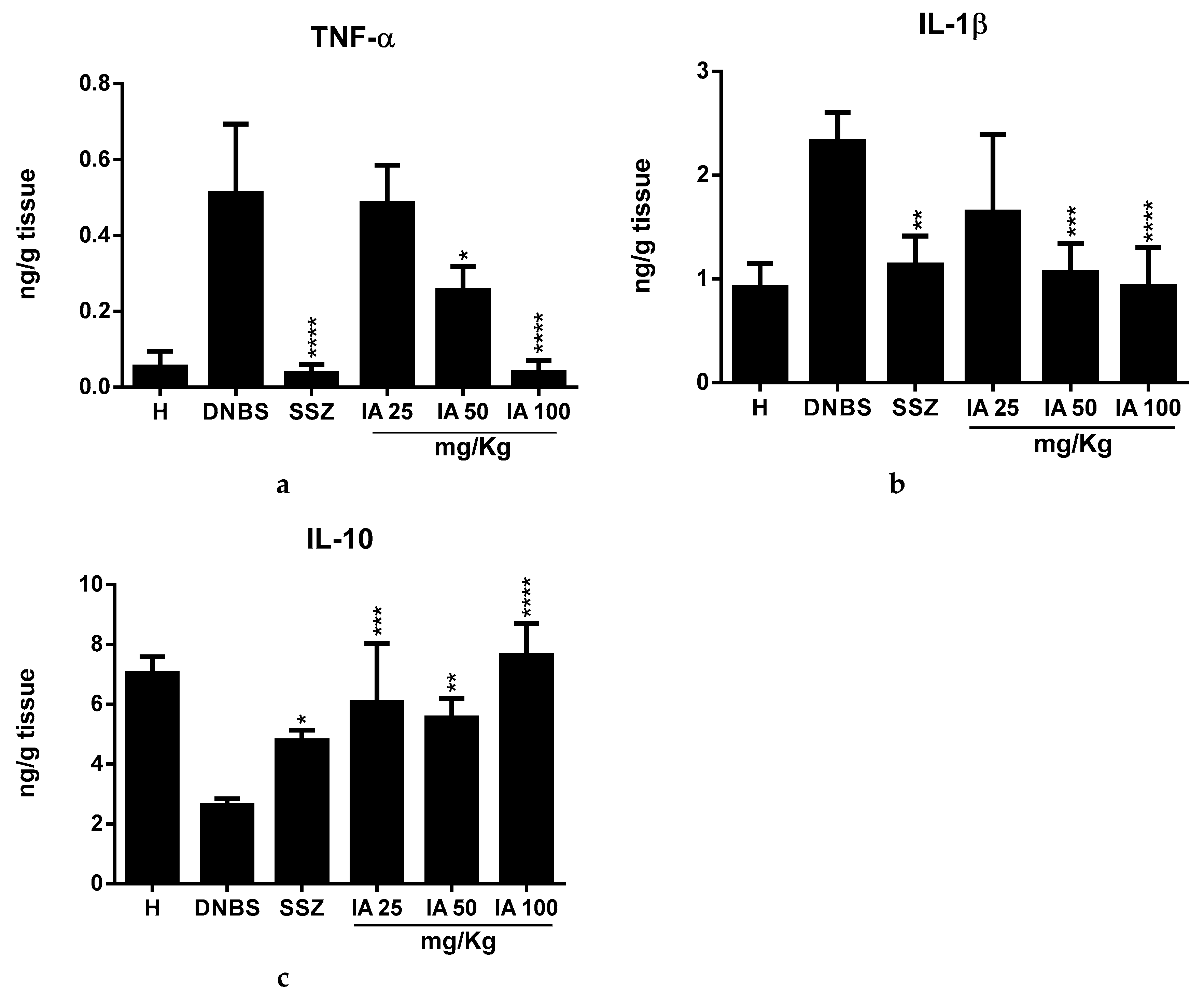
2.3. Effect of the Ipomoea asarifolia Aqueous Extract on the Colonic Levels of TNF-α, IL-1β, and IL-10
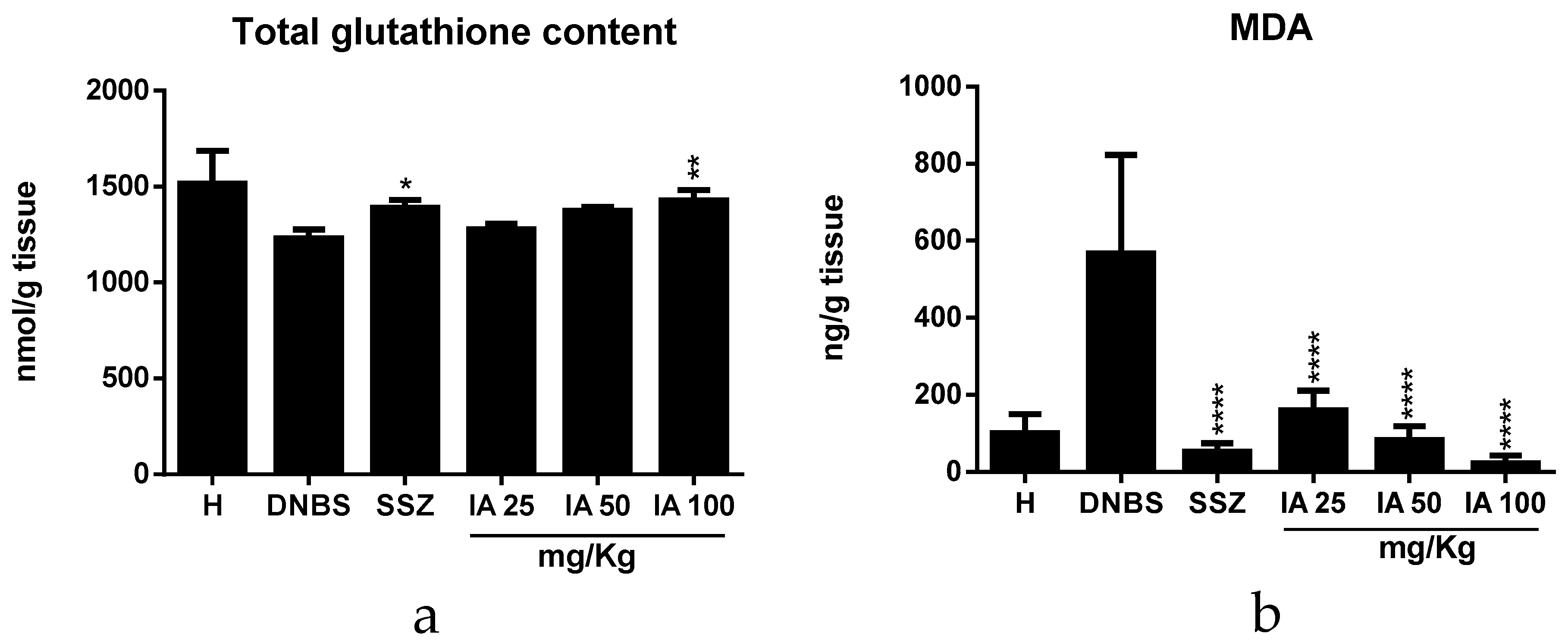
2.4. Effect of Ipomoea asarifolia Aqueous Extract on Colonic Oxidative Stress
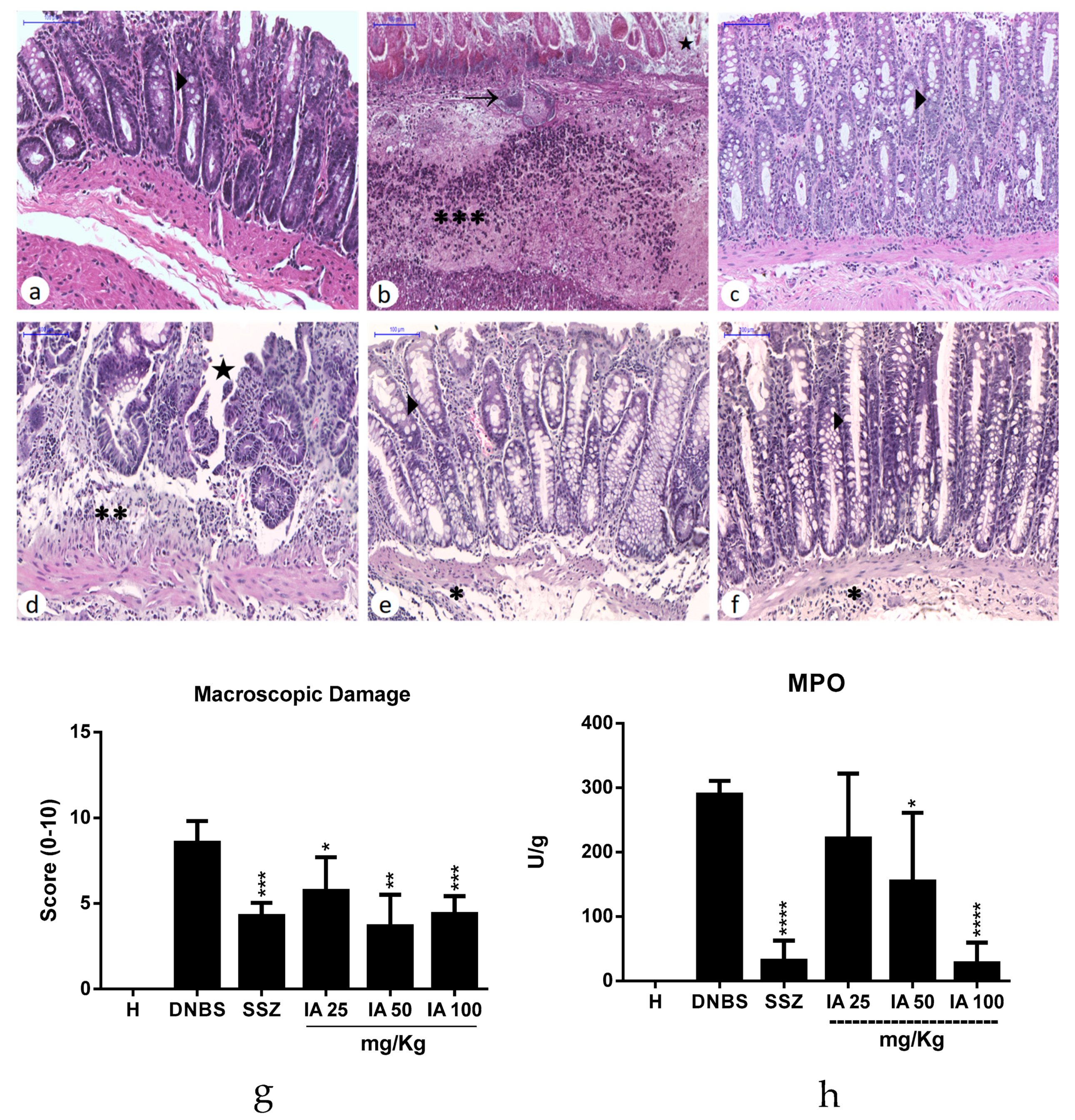
2.5. Histological Analysis and Myeloperoxidase Activity
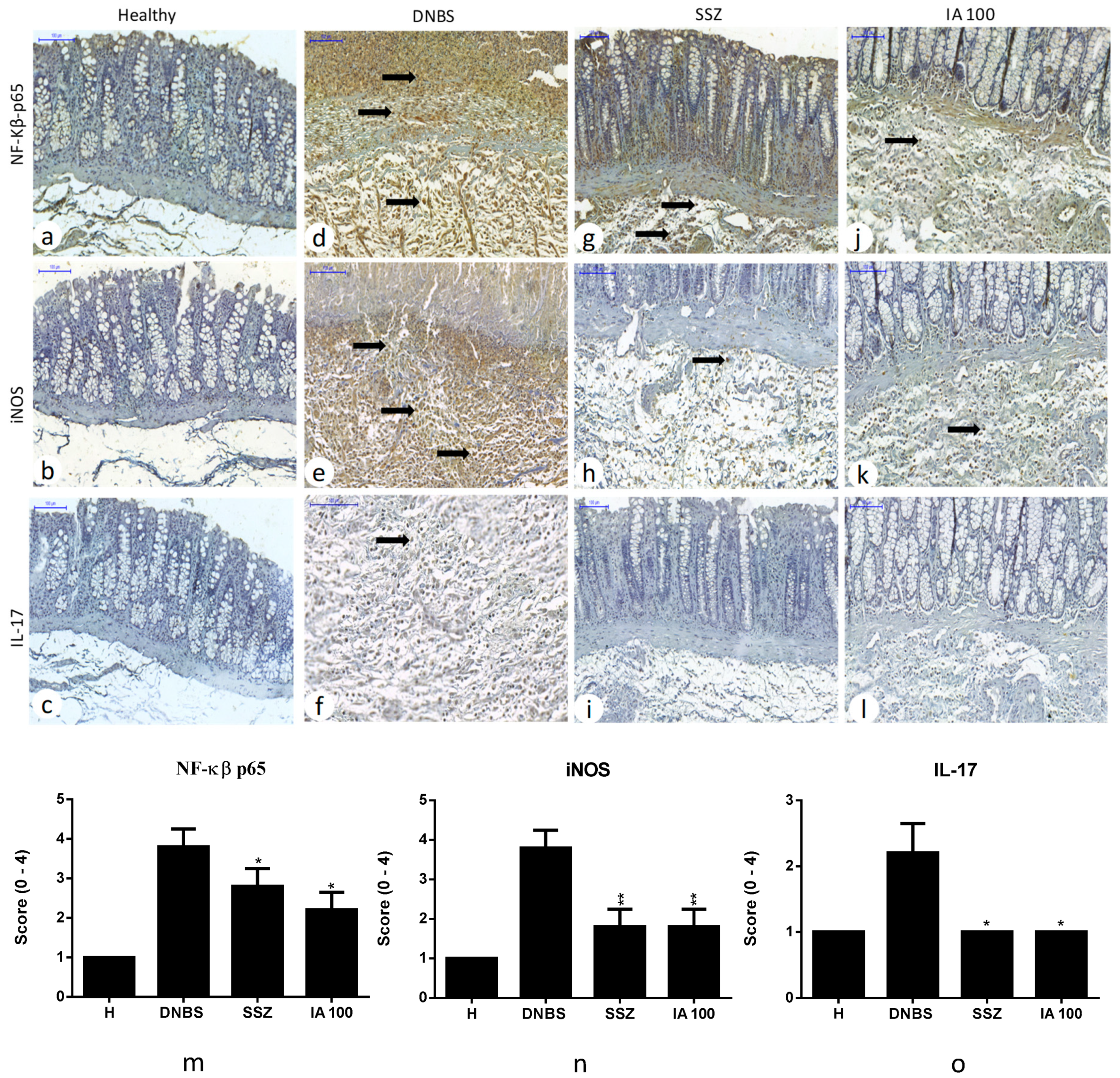
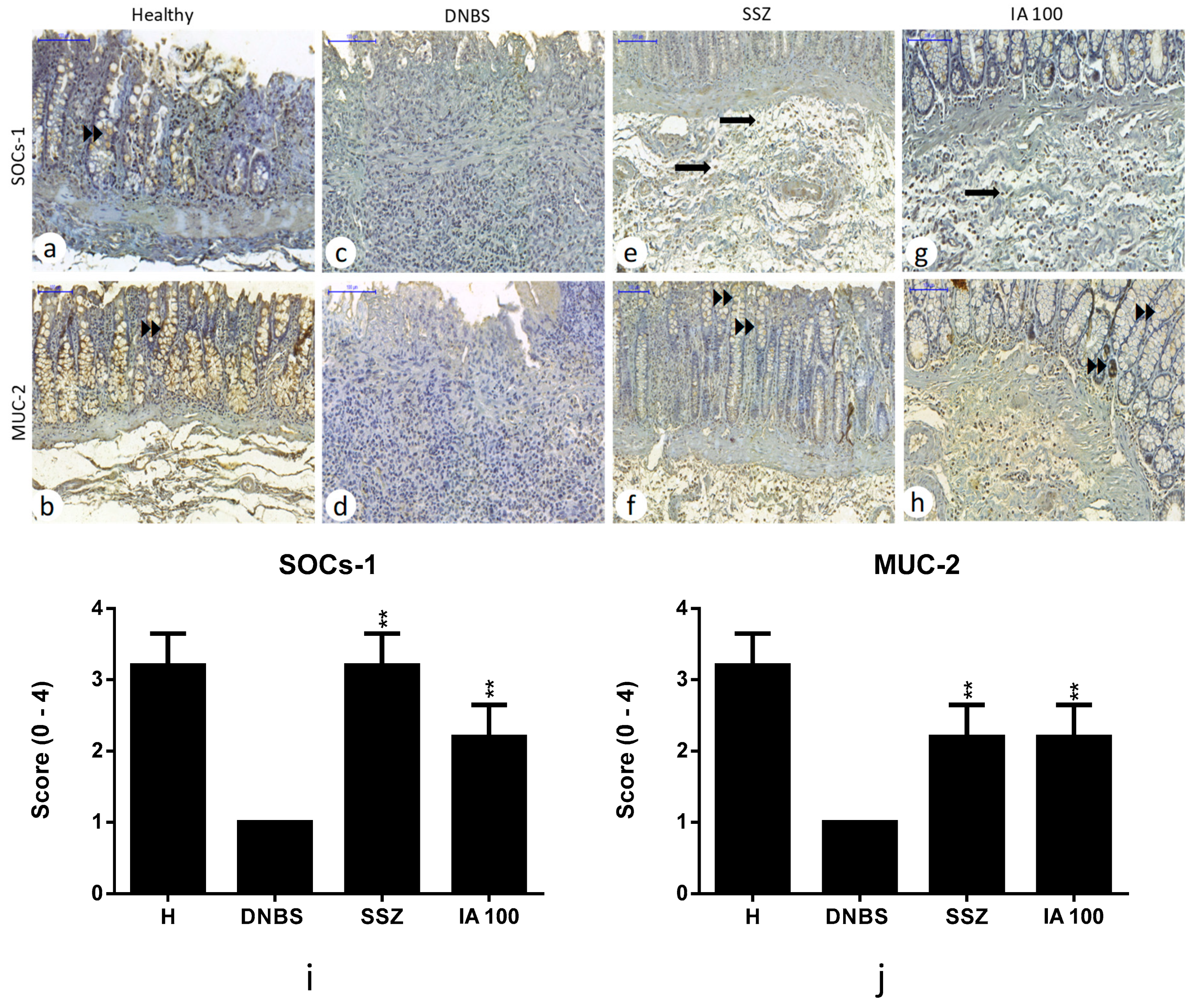
2.6. Immunohistochemical Analysis
3. Discussion
4. Materials and Methods
4.1. Plant Material and Preparation of Extracts
4.2. Animals
4.3. Experimental Design
4.4. DNBS-Induced Colitis
4.5. Evaluation of Intestinal Inflammation
4.6. Gene Expression Analysis
4.7. Histopathological Analysis
4.8. Immunohistochemical Analysis
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| DAI | Disease activity index |
| DNBS | 2,4-Dinitrobenzene sulfonic acid |
| IA | Ipomoea asarifolia |
| IBD | Inflammatory bowel disease |
| IL-10 | Interleukin-10 |
| IL-17 | Interleukin-17 |
| IL-1β | Interleukin-1β |
| iNOS | Inducible nitric oxide synthase |
| JNK1 | c-Jun N-terminal kinases 1 |
| MDA | Malonildialdehyde |
| MDI | Macroscopic damage index |
| MPO | Myeloperoxidase |
| MUC-2 | Mucin-2 |
| NF-κβ | Nuclear factor kappa β |
| SSZ | Sufasalazine |
| SOCs-1 | Cytokine signaling suppressor-1 |
| STAT3 | Activator of transcription 3 |
| TNF-α | Tumor necrosis factor-α |
References
- Wirtz, S.; Neufert, C.; Weigmann, B.; Neurath, M.F. Chemically Induced Mouse Models of Intestinal Inflammation. Nat. Protoc. 2007, 2, 541–546. [Google Scholar] [CrossRef] [PubMed]
- Zhernakova, A.; Festen, E.M.; Franke, L.; Trynka, G.; Van Diemen, C.C.; Monsuur, A.J.; Bevova, M.; Nijmeijer, R.M.; Van Slot, R.; Heijmans, R.; et al. Genetic Analysis of Innate Immunity in Crohn’ S Disease and Ulcerative Colitis Identifies Two Susceptibility Loci Harboring CARD9 and IL18RAP. J. Hum. Genet. 2008, 1202–1210. [Google Scholar] [CrossRef] [PubMed]
- Strober, W.; Fuss, I.; Mannon, P. The Fundamental Basis of Inflammatory Bowel Disease. J. Clin. Investig. 2007, 117, 514–521. [Google Scholar] [CrossRef] [PubMed]
- Wallace, K.L.; Zheng, L.B.; Kanazawa, Y.; Shih, D.Q. Immunopathology of Inflammatory Bowel Disease. World J. Gastroenterol. 2014, 20. [Google Scholar] [CrossRef] [PubMed]
- Neurath, M.F. Cytokines in Inflammatory Bowel Disease. Nat. Rev. Immunol. 2014, 14, 329–342. [Google Scholar] [CrossRef] [PubMed]
- Guan, Q.; Zhang, J. Recent Advances: The Imbalance of Cytokines in the Pathogenesis of Inflammatory Bowel Disease. Mediat. Inflamm. 2017, 2017, 4810258. [Google Scholar] [CrossRef] [PubMed]
- Lista do Brasil-Ipomoea asarifolia (Desr.) Roem. & Schult. Available online: http://floradobrasil.jbrj.gov.br/jabot/FichaPublicaTaxonUC/FichaPublicaTaxonUC.do?id=FB7026 (accessed on 19 September 2018).
- Riet-Correa, F.; Medeiros, R.M.T. Intoxicações por plantas em ruminantes no Brasil e no Uruguai: Importância econômica, controle e riscos para a saúde pública. Pesqui. Vet. Bras. 2001, 21, 38–42. [Google Scholar] [CrossRef]
- Mello, G.W.S.; Oliveira, D.M.; Carvalho, C.J.S.; VianaPires, L.; Costa, F.A.L.; Riet-Correa, F.; Silva, S.M.M. Plantas tóxicas para ruminantes e eqüídeos no norte Piauiense. Pesq. Vet. Bras. 2010, 30, 1–9. [Google Scholar] [CrossRef]
- Akindele, A.J.; Unachukwu, E.G.; Osiagwu, D.D. 90 Days toxicological assessment of hydroethanolic leaf extract of Ipomoea asarifolia (Desr.) Roem. and Schult. (Convolvulaceae) in rats. J. Ethnopharmacol. 2015, 174, 582–594. [Google Scholar] [CrossRef] [PubMed]
- Albuquerque, U.P.; Medeiros, P.M.; Almeida, A.L.S.; Monteiro, J.M.; Neto-Lins, E.M.F.; Melo, J.G.; Santos, J.P. Medicinal plants of the caatinga (semi-arid) vegetation of NE Brazil: A quantitative approach. J. Ethnopharmacol. 2007, 114, 325–354. [Google Scholar] [CrossRef]
- Lawal, U.; Ibrahim, H.; Agunu, A.; Abdulahi, Y. Anti-inflammatory and analgesic activity of water extract from Ipomoea asarifolia Desr (Convolvulaceae). Afr. J. Biotechnol. 2010, 9, 8877–8880. [Google Scholar]
- Jegede, I.A.; Nwinyi, F.C.; Ibrahim, J.; Ugbabe, G.; Dzarma, S.; Kunle, O.F. Investigation of phytochemical, anti- inflammatory and anti-nociceptive properties of Ipomoea asarifolia leaves. J. Med. Plants Res. 2009, 3, 160–165. [Google Scholar]
- Lima, M.C.; Bitencourt, M.A.; Furtado, A.A.; Oliveira Rocha, H.A.; Oliveira, R.M.; da Silva-Júnior, A.A.; Tabosa do Egito, E.S.; Tambourgi, D.V.; Zucolotto, S.M.; Fernandes-Pedrosa Mde, F. Ipomoea Asarifolia Neutralizes Inflammation Induced by Tityus Serrulatus Scorpion Venom. J. Ethnopharmacol. 2014, 153, 890–895. [Google Scholar] [CrossRef] [PubMed]
- Furtado, A.A.; Torres-Rêgo, M.; Lima, M.C.J.S.; Bitencourt, M.A.O.; Estrela, A.B.; Souza da Silva, N.; da Silva Siqueira, E.M.; Tomaz, J.C.; Lopes, N.P.; Silva-Júnior, A.A.; et al. Aqueous Extract from Ipomoea Asarifolia (Convolvulaceae) Leaves and Its Phenolic Compounds Have Anti-Inflammatory Activity in Murine Models of Edema, Peritonitis and Air-Pouch Inflammation. J. Ethnopharmacol. 2016, 192, 225–235. [Google Scholar] [CrossRef] [PubMed]
- Martín, R.; Chain, F.; Miquel, S.; Motta, J.-P.; Vergnolle, N.; Sokol, H.; Langella, P. Using Murine Colitis Models to Analyze Probiotics–host Interactions. FEMS Microbiol. Rev. 2017, 41 (Suppl. 1), S49–S70. [Google Scholar] [CrossRef] [PubMed]
- Cooper, H.S.; Murthy, S.N.; Shah, R.S.; Sedergran, D.J. Clinicopathologic Study of Dextran Sulfate Sodium Experimental Murine Colitis. Lab. Investig. 1993, 69, 238–249. [Google Scholar]
- Bell, C.J.; Gall, D.G.; Wallace, J.L. Disruption of Colonic Electrolyte Transport in Experimental Colitis. Am. J. Physiol. 1995, 268 Pt 1, G622–G630. [Google Scholar] [CrossRef]
- Araújo, D.F.S.; Guerra, G.C.B.; Júnior, R.F.A.; Antunes de Araújo, A.; Antonino de Assis, P.O.; Nunes de Medeiros, A.; Formiga de Sousa, Y.R.; Pintado, M.M.E.; Gálvez, J.; Queiroga, R.C.R.D.E. Goat Whey Ameliorates Intestinal Inflammation on Acetic Acid-Induced Colitis in Rats. J. Dairy Sci. 2016, 99, 1–12. [Google Scholar] [CrossRef]
- Araújo, D.F.S.; Guerra, G.C.B.; Pintado, M.M.E.; Sousa, Y.R.F.; Algieri, F.; Rodriguez-Nogales, A.; Araújo, R.F., Jr.; Gálvez, J.; Queiroga, R.C.R.E.; Rodriguez-Cabezas, M.E. Intestinal Anti-Inflammatory Effects of Goat Whey on DNBS-Induced Colitis in Mice. PLoS ONE 2017, 12, e0185382. [Google Scholar] [CrossRef]
- De Assis, P.O.; Guerra, G.C.; de Souza Araújo, D.F.; de Araújo Júnior, R.F.; Machado, T.A.; de Araújo, A.A.; de Lima, T.A.; Garcia, H.E.; do Egypto, R.D. Intestinal Anti-Inflammatory Activity of Goat Milk and Goat Yoghurt in the Acetic Acid Model of Rat Colitis. Int. Dairy J. 2016, 46, 45–54. [Google Scholar] [CrossRef]
- Lichtenstein, G.R.; Hanauer, S.B.; Sandborn, W.J. Management of Crohn’s Disease in Adults. Am. J. Gastroenterol. 2009, 104, 465–483. [Google Scholar] [CrossRef] [PubMed]
- Farzaei, M.; Rahimi, R.; Abdollahi, M. The Role of Dietary Polyphenols in the Management of Inflammatory Bowel Disease. Curr. Pharm. Biotechnol. 2015, 16, 196–210. [Google Scholar] [CrossRef] [PubMed]
- Zorrilla, P.; Rodriguez-Nogales, A.; Algieri, F.; Garrido-Mesa, N.; Olivares, M.; Rondón, D.; Zarzuelo, A.; Utrilla, M.P.; Galvez, J.; Rodriguez-Cabezas, M.E. Intestinal Anti-Inflammatory Activity of the Polyphenolic-Enriched Extract Amanda® in the Trinitrobenzenesulphonic Acid Model of Rat Colitis. J. Funct. Foods 2014, 2, 449–459. [Google Scholar] [CrossRef]
- Khor, B.; Gardet, A.; Xavier, R.J. Genetics and Pathogenesis of Inflammatory Bowel Disease. Nature 2011, 474, 307–317. [Google Scholar] [CrossRef] [PubMed]
- Borja-Cacho, D.; Matthews, J. NIH Public Access. Nano 2008, 6, 2166–2171. [Google Scholar] [CrossRef]
- Santana-Gálvez, J.; Cisneros-Zevallos, L.; Jacobo-Velázquez, D.A. Chlorogenic Acid: Recent Advances on Its Dual Role as a Food Additive and a Nutraceutical against Metabolic Syndrome. Molecules 2017, 22, 358. [Google Scholar] [CrossRef]
- Nafees, S.; Rashid, S.; Ali, N.; Hasan, S.K.; Sultana, S. Rutin Ameliorates Cyclophosphamide Induced Oxidative Stress and Inflammation in Wistar Rats: Role of NFκB/MAPK Pathway. Chem. Biol. Interact. 2015, 231, 98–107. [Google Scholar] [CrossRef]
- Cobo, E.R.; Kissoon-Singh, V.; Moreau, F.; Chadee, K. Colonic MUC2 Mucin Regulates the Expression and Antimicrobial Activity of β-Defensin 2. Mucosal Immunol. 2015, 8, 1360–1372. [Google Scholar] [CrossRef]
- Lewandowska, H.; Kalinowska, M.; Lewandowski, W.; Stepkowski, T.M.; Brzóska, K. The Role of Natural Polyphenols in Cell Signaling and Cytoprotection against Cancer Development. J. Nutr. Biochem. 2016, 32, 1–19. [Google Scholar] [CrossRef]
- Mascaraque, C.; Aranda, C.; Ocón, B.; Monte, M.J.; Suárez, M.D.; Zarzuelo, A.; Marín, J.J.G.; Martínez-Augustin, O.; De Medina, F.S. Rutin Has Intestinal Antiinflammatory Effects in the CD4+ CD62L+ T Cell Transfer Model of Colitis. Pharmacol. Res. 2014, 90, 48–57. [Google Scholar] [CrossRef]
- Vezza, T.; Rodríguez-Nogales, A.; Algieri, F.; Utrilla, M.P.; Rodriguez-Cabezas, M.E.; Galvez, J. Flavonoids in Inflammatory Bowel Disease: A Review. Nutrients 2016, 8, 211. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Wu, X.; Cao, S.; Cromie, M.; Shen, Y.; Feng, Y.; Yang, H.; Li, L. Chlorogenic Acid Ameliorates Experimental Colitis by Promoting Growth of Akkermansia in Mice. Nutrients 2017, 9, 677. [Google Scholar] [CrossRef] [PubMed]
- Fitzpatrick, L.R.; Wang, J.; Le, T. Caffeic Acid Phenethyl Ester, an Inhibitor of Nuclear Factor-B, Attenuates Bacterial Peptidoglycan Polysaccharide-Induced Colitis in Rats. J. Pharmacol. Exp. Ther. 2001, 299, 915–920. [Google Scholar]
- Alzoghaibi, M.A. Concepts of Oxidative Stress and Antioxidant Defense in Crohn’s Disease. World J. Gastroenterol. 2013, 19, 6540–6547. [Google Scholar] [CrossRef]
- Gaschler, M.M.; Stockwell, B.R. Lipid Peroxidation in Cell Death. Biochem. Biophys. Res. Commun. 2017, 482, 419–425. [Google Scholar] [CrossRef] [PubMed]
- Hussain, S.P.; Hofseth, L.J.; Harris, C.C. Radical Causes of Cancer. Nat. Rev. Cancer 2003, 3, 276–285. [Google Scholar] [CrossRef]
- Langhorst, J.; Elsenbruch, S.; Mueller, T.; Rueffer, A.; Spahn, G.; Michalsen, A.; Dobos, G.J. Comparison of 4 Neutrophil-Derived Proteins in Feces as Indicators of Disease Activity in Ulcerative Colitis. Inflamm. Bowel Dis. 2005, 11, 1085–1091. [Google Scholar] [CrossRef] [PubMed]
- Slater, T.W.; Finkielsztein, A.; Mascarenhas, L.A.; Mehl, L.C.; Butin-Israeli, V.; Sumagin, R. Neutrophil Microparticles Deliver Active Myeloperoxidase to Injured Mucosa To Inhibit Epithelial Wound Healing. J. Immunol. 2017, 198, 2886–2897. [Google Scholar] [CrossRef] [Green Version]
- Duque, G.A.; Descoteaux, A. Macrophage Cytokines: Involvement in Immunity and Infectious Diseases. Front. Immunol. 2014, 5, 1–12. [Google Scholar] [CrossRef]
- Sakthivel, K.M.; Guruvayoorappan, C. Amentoflavone Inhibits iNOS, COX-2 Expression and Modulates Cytokine Profile, NF-κB Signal Transduction Pathways in Rats with Ulcerative Colitis. Int. Immunopharmacol. 2013, 17, 907–916. [Google Scholar] [CrossRef]
- You, B.H.; Chae, H.-S.; Song, J.; Ko, H.W.; Chin, Y.-W.; Choi, Y.H. α-Mangostin Ameliorates Dextran Sulfate Sodium-Induced Colitis through Inhibition of NF-κB and MAPK Pathways. Int. Immunopharmacol. 2017, 49, 212–221. [Google Scholar] [CrossRef] [PubMed]
- Vukelić, I.; Detel, D.; Pučar, L.B.; Potočnjak, I.; Buljević, S.; Domitrović, R. Chlorogenic Acid Ameliorates Experimental Colitis in Mice by Suppressing Signaling Pathways Involved in Inflammatory Response and Apoptosis. Food Chem. Toxicol. 2018, 121, 140–150. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Nagahashi, M.; Kim, E.Y.; Harikumar, K.B.; Yamada, A.; Huang, W.-C.; Hait, N.C.; Allegood, J.C.; Price, M.M.; Avni, D.; et al. Sphingosine-1-Phosphate Links Persistent STAT3 Activation, Chronic Intestinal Inflammation, and Development of Colitis-Associated Cancer. Cancer Cell. 2013, 23, 107–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinon, F.; Burns, K.; Tschopp, J. The Inflammasome: A Molecular Platform Triggering Activation of Inflammatory Caspases and Processing of ProIL-β. Mol. Cell. 2002, 10, 417–426. [Google Scholar] [CrossRef]
- Robertson, A.A.B. Inhibiting In Fl Ammasomes with Small Molecules. In Inflammasomes: Clinical and Therapeutic Implications; Experientia Supplementum; Cordero, M., Alcocer-Gómez, E., Eds.; Springer: Cham, Switzerland, 2018; Volume 108, ISBN 978-3-319-89389-1. [Google Scholar]
- Begue, B.; Verdier, J.; Rieux-Laucat, F.; Goulet, O.; Morali, A.; Canioni, D.; Hugot, J.P.; Daussy, C.; Verkarre, V.; Pigneur, B.; et al. Defective IL10 Signaling Defining a Subgroup of Patients with Inflammatory Bowel Disease. Am. J. Gastroenterol. 2011, 106, 1544–1555. [Google Scholar] [CrossRef]
- Sumagin, R.; Prizant, H.; Lomakina, E.; Waugh, R.E.; Sarelius, I.H. LFA-1 and Mac-1 Define Characteristically Different Intralumenal Crawling and Emigration Patterns for Monocytes and Neutrophils in Situ. J. Immunol. 2010, 185, 7057–7066. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wu, J.; Ye, B.; Wang, Q.; Xie, X.; Shen, H. Protective Effect of Curcumin on TNBS-Induced Intestinal Inflammation Is Mediated through the JAK/STAT Pathway. BMC Complement. Altern. Med. 2016, 16, 299. [Google Scholar] [CrossRef]
- Rubtsov, Y.P.; Rasmussen, J.P.; Chi, E.Y.; Fontenot, J.; Castelli, L.; Ye, X.; Treuting, P.; Siewe, L.; Roers, A.; Henderson, W.R.; et al. Regulatory T Cell-Derived Interleukin-10 Limits Inflammation at Environmental Interfaces. Immunity 2008, 28, 546–558. [Google Scholar] [CrossRef] [Green Version]
- Manzanillo, P.; Eidenschenk, C.; Ouyang, W. Deciphering the Crosstalk among IL-1 and IL-10 Family Cytokines in Intestinal Immunity. Trends Immunol. 2015, 36, 471–478. [Google Scholar] [CrossRef]
- Morris, G.P.; Beck, P.L.; Herridge, M.S.; Depew, W.T.; Szewczuk, M.R.; Wallace, J.L. Hapten-Induced Model of Chronic Inflammation and Ulceration in the Colon Rat. Gastroenterology 1989, 96, 795–803. [Google Scholar] [CrossRef]
- Krawisz, J.E.; Sharon, P.; Stenson, W.F. Quantitative Assay for Acute Intestinal Inflammation Based on Myeloperoxidase Activity. Assessment of Inflammation in Rat and Hamster Models. Gastroenterology 1984, 87, 1344–1350. [Google Scholar] [PubMed]
- Anderson, M.E. Determination of Glutathione and Glutathione Disulfide in Biological Samples. Methods Enzymol. 1985, 113, 548–555. [Google Scholar] [CrossRef] [PubMed]
- Esterbauer, H.; Cheeseman, K.H. Determination of Aldehydic Lipid Peroxidation Products: Malonaldehyde and 4-Hydroxynonenal. Methods Enzymol. 1990, 186, 407–421. [Google Scholar] [CrossRef] [PubMed]
- Wirtz, S.; Popp, V.; Kindermann, M.; Gerlach, K.; Weigmann, B.; Fichtner-Feigl, S.; Neurath, M.F. Chemically Induced Mouse Models of Acute and Chronic Intestinal Inflammation. Nat. Protoc. 2017, 12, 1295–1309. [Google Scholar] [CrossRef] [PubMed]
- Guerra, G.C.B.; de Menezes, M.S.; de Araújo, A.A.; de Araújo Júnior, R.F.; de Medeiros, C.A. Olmesartan Prevented Intra-articular Inflammation Induced by Zymosan in Rats. Biol Pharm. Bull 2016, 39, 1793–1801. [Google Scholar] [CrossRef] [PubMed] [Green Version]







© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Da Silva, V.C.; De Araújo, A.A.; Araújo, D.F.d.S.; Lima, M.C.J.S.; Vasconcelos, R.C.; De Araújo Júnior, R.F.; Langasnner, S.M.Z.; Pedrosa, M.D.F.F.; De Medeiros, C.A.C.X.; Guerra, G.C.B. Intestinal Anti-Inflammatory Activity of the Aqueous Extract from Ipomoea asarifolia in DNBS-Induced Colitis in Rats. Int. J. Mol. Sci. 2018, 19, 4016. https://doi.org/10.3390/ijms19124016
Da Silva VC, De Araújo AA, Araújo DFdS, Lima MCJS, Vasconcelos RC, De Araújo Júnior RF, Langasnner SMZ, Pedrosa MDFF, De Medeiros CACX, Guerra GCB. Intestinal Anti-Inflammatory Activity of the Aqueous Extract from Ipomoea asarifolia in DNBS-Induced Colitis in Rats. International Journal of Molecular Sciences. 2018; 19(12):4016. https://doi.org/10.3390/ijms19124016
Chicago/Turabian StyleDa Silva, Valéria Costa, Aurigena Antunes De Araújo, Daline Fernandes de Souza Araújo, Maíra Conceição Jerônimo Souza Lima, Roseane Carvalho Vasconcelos, Raimundo Fernandes De Araújo Júnior, Silvana Maria Zucolotto Langasnner, Matheus De Freitas Fernandes Pedrosa, Caroline Addison Carvalho Xavier De Medeiros, and Gerlane Coelho Bernardo Guerra. 2018. "Intestinal Anti-Inflammatory Activity of the Aqueous Extract from Ipomoea asarifolia in DNBS-Induced Colitis in Rats" International Journal of Molecular Sciences 19, no. 12: 4016. https://doi.org/10.3390/ijms19124016