Flavonones from Penthorum chinense Ameliorate Hepatic Steatosis by Activating the SIRT1/AMPK Pathway in HepG2 Cells


Abstract
:1. Introdution
2. Results
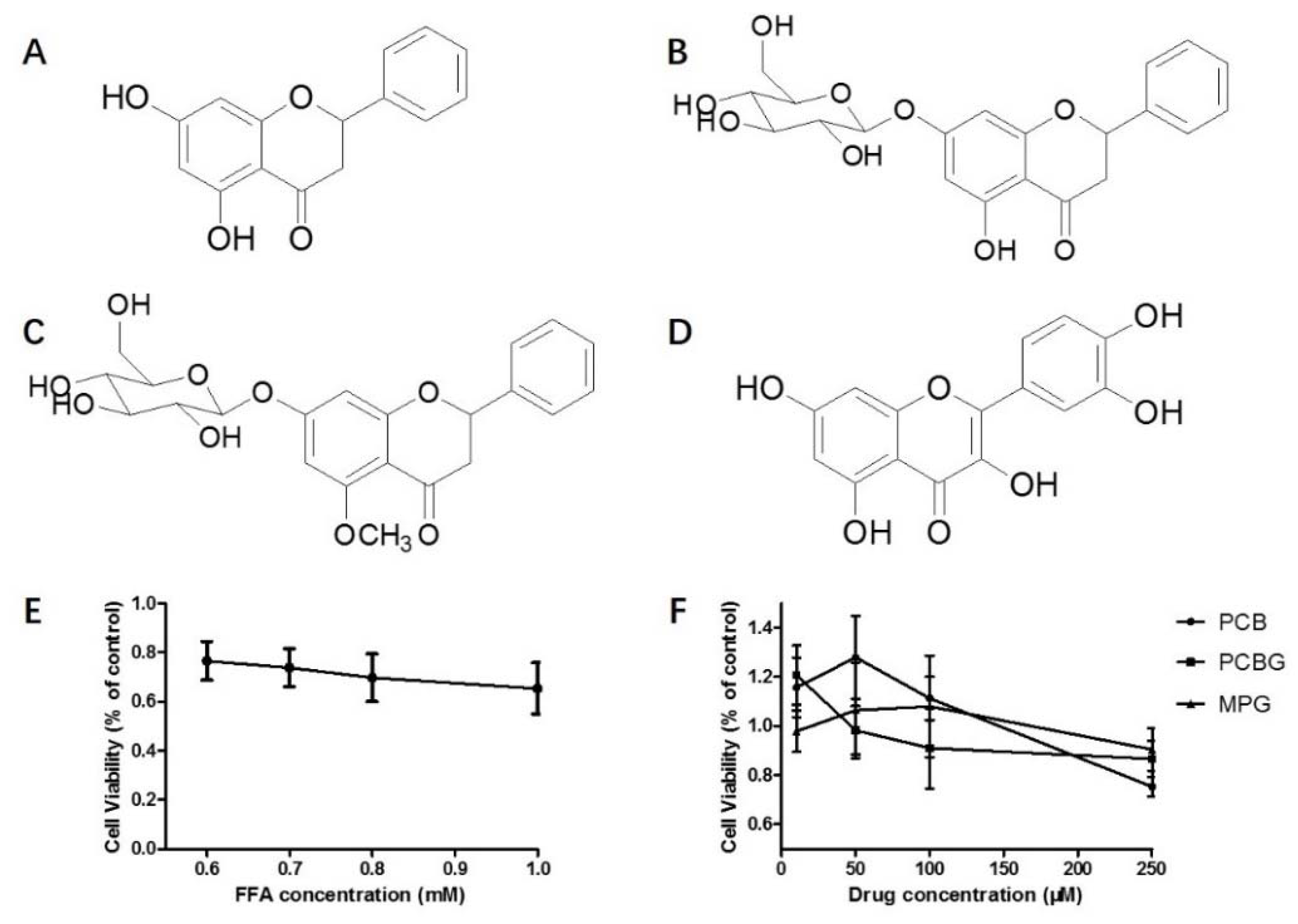
2.1. FFA-Induced Cytotoxicity and Concentration Screening of PCB, PCBG, and MPG
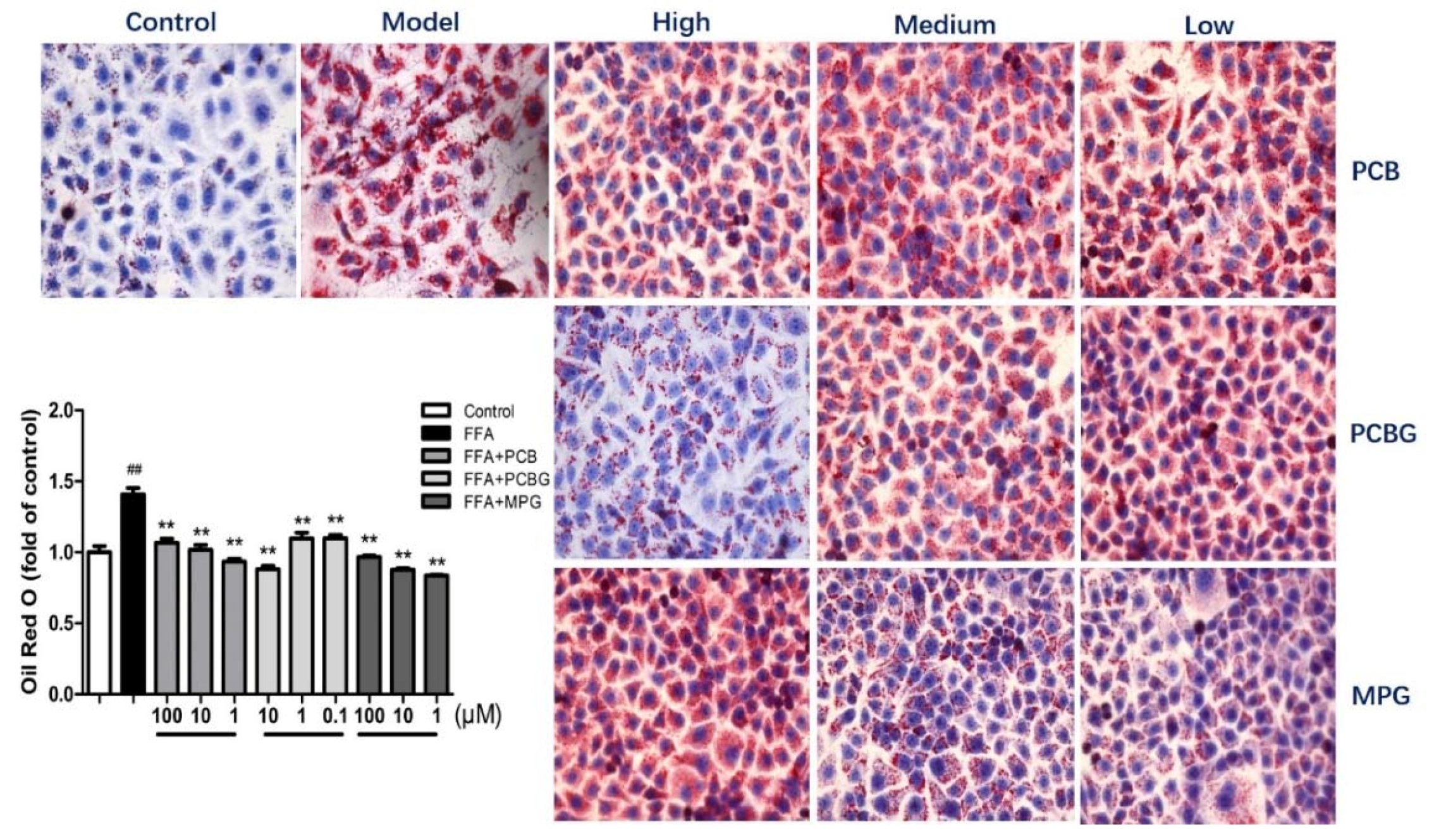
2.2. PCB, PCBG, and MPG Inhibited Intracellular Lipid Accumulation in HepG2 Cells
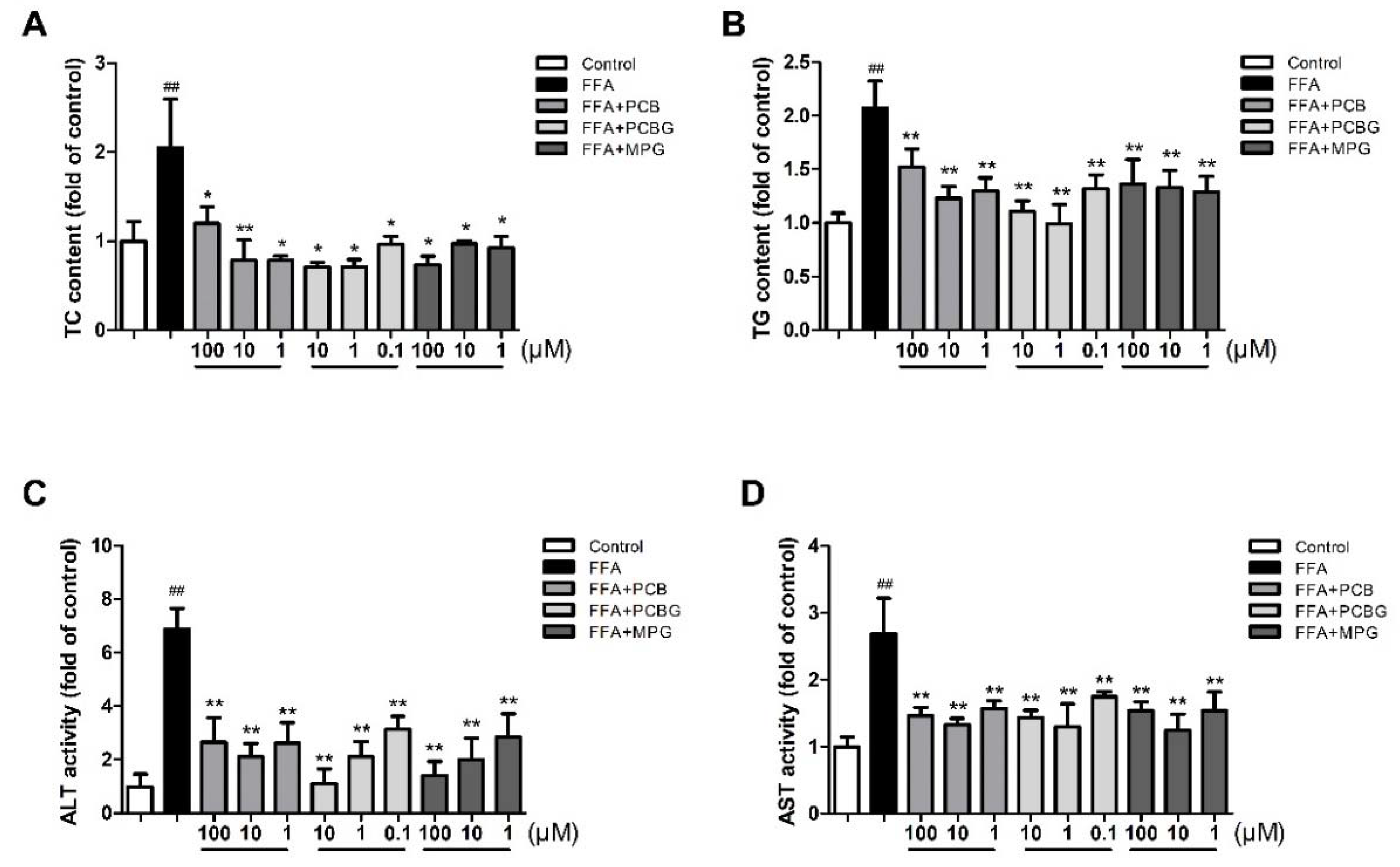
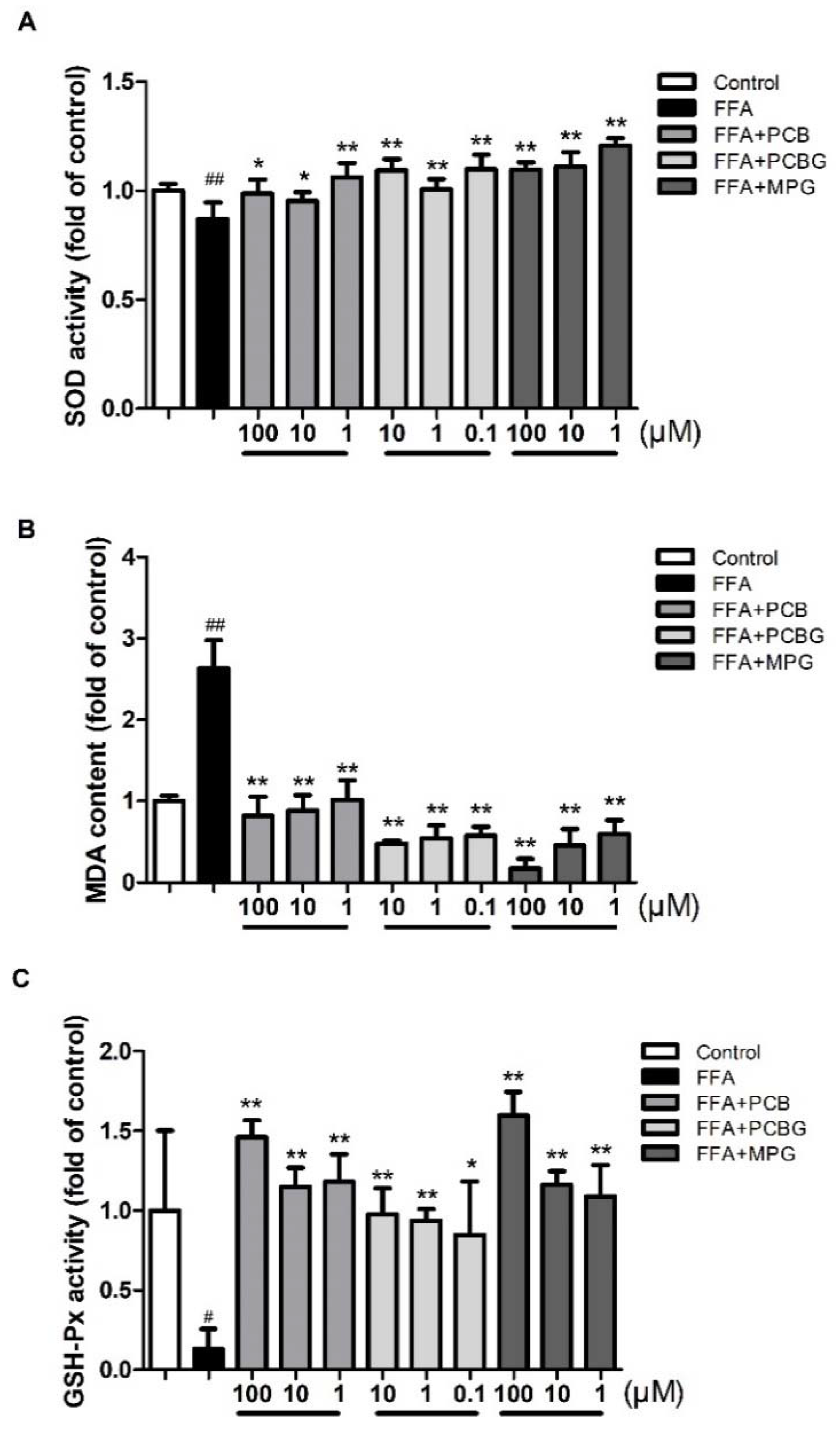
2.3. PCB, PCBG, and MPG Weakened Lipid Levels and Up-Regulated Antioxidant Enzymes
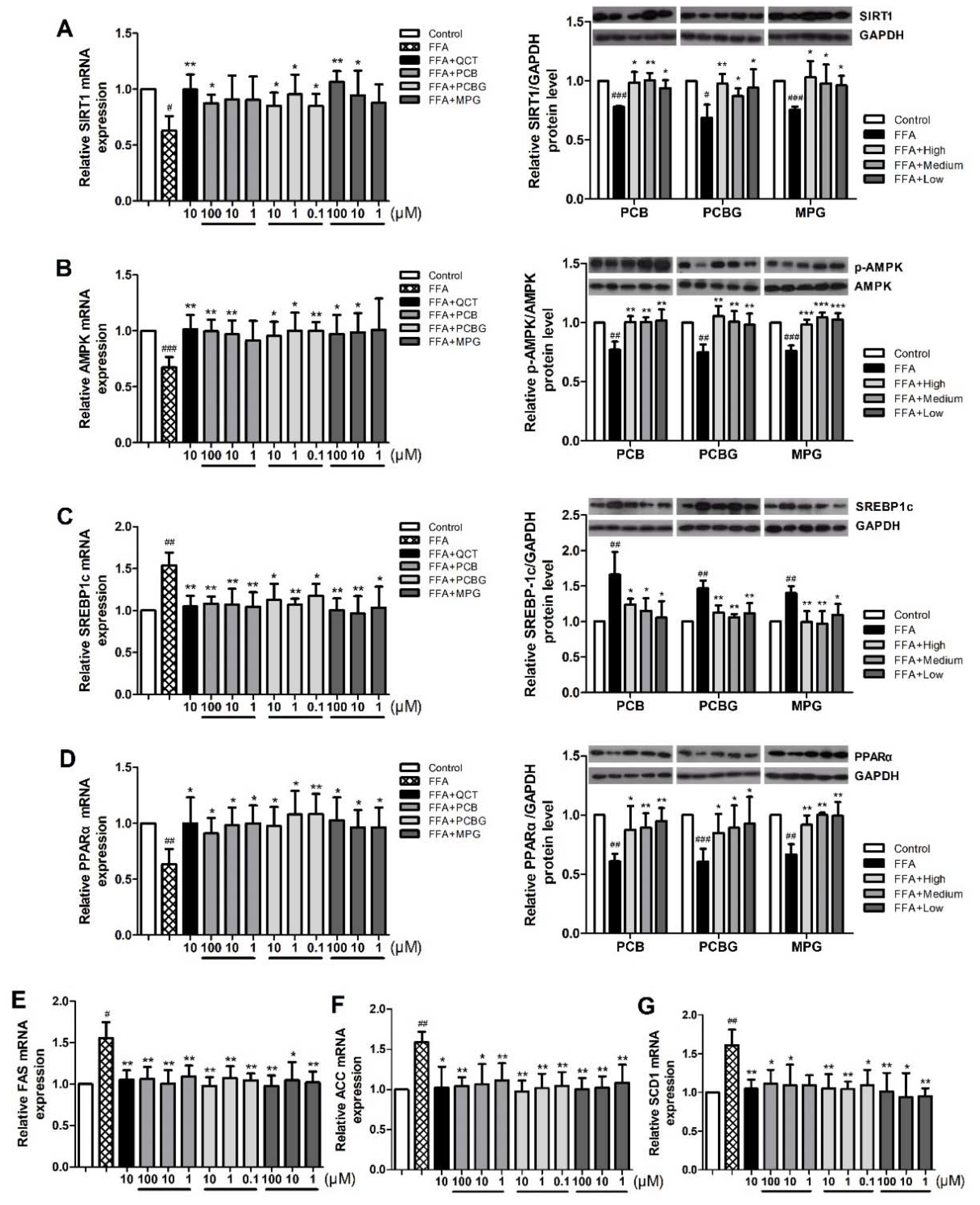
2.4. Effects of PCB, PCBG, and MPG on the Expression of Factors Associated with Hepatic Lipid Accumulation
2.5. Effects of PCB, PCBG, and MPG on AMPK and SIRT1 Activities in HepG2 Cells
2.6. Docking Studies
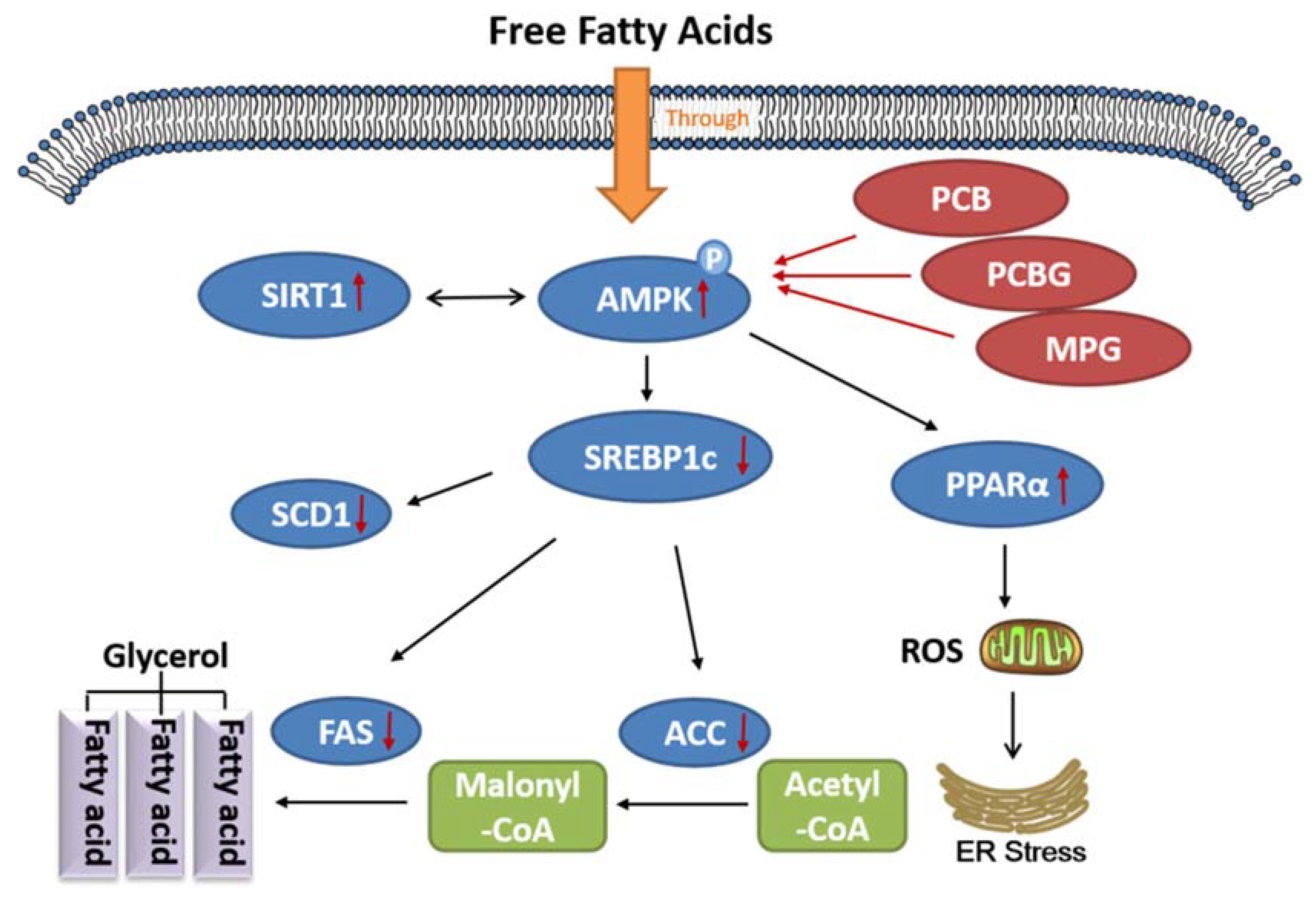
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Measurement of Cell Viability
4.3. Oil Red O Staining
4.4. Biochemical Assay
4.5. Western Blots Analysis
4.6. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
4.7. Molecular Docking
4.8. Statistics
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Younossi, Z.M.; Koenig, A.B.; Abdelatif, D.; Fazel, Y.; Henry, L.; Wymer, M. Global epidemiology of nonalcoholic fatty liver disease-Meta-analytic assessment of prevalence, incidence, and outcomes. Hepatology 2016, 64, 73–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, J.G.; Kim, S.U.; Wong, V.W. New trends on obesity and nafld in asia. J. Hepatol. 2017, 67, 862–873. [Google Scholar] [CrossRef] [PubMed]
- Reccia, I.; Kumar, J.; Akladios, C.; Virdis, F.; Pai, M.; Habib, N.; Spalding, D. Non-alcoholic fatty liver disease: A sign of systemic disease. Metabolism 2017, 72, 94–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abenavoli, L. Non-alcoholic fatty liver disease: Today and tomorrow. Rev. Recent Clin. Trials 2014, 9, 125. [Google Scholar] [CrossRef] [PubMed]
- Chalasani, N.; Younossi, Z.; Lavine, J.E.; Diehl, A.M.; Brunt, E.M.; Cusi, K.; Charlton, M.; Sanyal, A.J. The diagnosis and management of non-alcoholic fatty liver disease: Practice guideline by the American Gastroenterological Association, American Association for the Study of Liver Diseases, and American College of Gastroenterology. Gastroenterology 2012, 142, 1592–1609. [Google Scholar] [CrossRef] [PubMed]
- Dyson, J.; Jaques, B.; Chattopadyhay, D.; Lochan, R.; Graham, J.; Das, D.; Aslam, T.; Patanwala, I.; Gaggar, S.; Cole, M.; et al. Hepatocellular cancer: The impact of obesity, type 2 diabetes and a multidisciplinary team. J. Hepatol. 2014, 60, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Musso, G.; Gambino, R.; Cassader, M.; Pagano, G. Meta-analysis: Natural history of non-alcoholic fatty liver disease (NAFLD) and diagnostic accuracy of non-invasive tests for liver disease severity. Ann. Med. 2011, 43, 617–649. [Google Scholar] [CrossRef] [PubMed]
- Nascimbeni, F.; Pais, R.; Bellentani, S.; Day, C.P.; Ratziu, V.; Loria, P.; Lonardo, A. From NAFLD in clinical practice to answers from guidelines. J. Hepatol. 2013, 59, 859–871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Than, N.N.; Newsome, P.N. A concise review of non-alcoholic fatty liver disease. Atherosclerosis 2015, 239, 192–202. [Google Scholar] [CrossRef] [PubMed]
- Stanković, M.N.; Mladenović, D.R.; Đuričić, I.; Šobajić, S.S.; Timić, J.; Jorgačević, B.; Aleksić, V.; Vučević, D.B.; Ješić-Vukićević, R.; Radosavljević, T.S. Time-dependent changes and association between liver free fatty acids, serum lipid profile and histological features in mice model of nonalcoholic fatty liver disease. Arch. Med. Res. 2014, 45, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Cantó, C.; Gerharthines, Z.; Feige, J.N.; Lagouge, M.; Noriega, L.; Milne, J.C.; Elliott, P.J.; Puigserver, P.; Auwerx, J. AMPK regulates energy expenditure by modulating NAD+ metabolism and SIRT1 activity. Nature 2009, 458, 1056–1060. [Google Scholar] [CrossRef] [PubMed]
- Lan, F.; Cacicedo, J.M.; Ruderman, N.; Ido, Y. SIRT1 modulation of the acetylation status, cytosolic localization, and activity of LKB1. Possible role in AMP-activated protein kinase activation. J. Biol. Chem. 2008, 283, 27628–27635. [Google Scholar] [CrossRef] [PubMed]
- Lim, C.T.; Kola, B.; Korbonits, M. AMPK as a mediator of hormonal signalling. J. Mol. Endocrinol. 2010, 44, 87–97. [Google Scholar] [CrossRef] [PubMed]
- You, M.; Matsumoto, M.; Pacold, C.M.; Cho, W.K.; Crabb, D.W. The role of AMP-activated protein kinase in the action of ethanol in the liver. Gastroenterology 2004, 127, 1798–1808. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.; Myers, R.; Li, Y.; Chen, Y.; Shen, X.; Fenykmelody, J.; Wu, M.; Ventre, J.; Doebber, T.; Fujii, N.; et al. Role of AMP-activated protein kinase in mechanism of metformin action. J. Clin. Investg. 2001, 108, 1167–1174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferre, P.; Foufelle, F. Hepatic steatosis: A role for de novo lipogenesis and the transcription factor SREBP-1c. Diabetes Obes. Metab. 2010, 12, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Fortin, É.; Blouin, R.; Lapointe, J.; Petit, H.V.; Palin, M.F. Linoleic acid, α-linolenic acid and enterolactone affect lipid oxidation and expression of lipid metabolism and antioxidant-related genes in hepatic tissue of dairy cows. Br. J. Nutr. 2017, 117, 1199–1211. [Google Scholar] [CrossRef] [PubMed]
- Kohjima, M.; Higuchi, N.; Kato, M.; Kotoh, K.; Yoshimoto, T.; Fujino, T.; Yada, M.; Yada, R.; Harada, N.; Enjoji, M.; et al. SREBP-1c, regulated by the insulin and AMPK signaling pathways, plays a role in nonalcoholic fatty liver disease. Int. J. Mol. Med. 2008, 21, 507–511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chirala, S.S.; Jayakumar, A.; Gu, Z.W.; Wakil, S.J. Human fatty acid synthase: Role of interdomain in the formation of catalytically active synthase dimer. Proc. Natl. Acad. Sci. USA 2001, 98, 3104–3108. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.H.; Shiu, S.M.; Wu, M.T.; Chen, W.L.; Wang, S.G.; Lee, H.M. Monacolin K affects lipid metabolism through SIRT1/AMPK pathway in HepG2 cells. Arch. Pharm. Res. 2013, 36, 1541–1551. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Wu, X.; Jiang, Y.; Zhang, D.Y. The research progress of Penthorum chinense Pursh. Food Drug 2013, 15, 202–205. [Google Scholar]
- Zeng, Q.H.; Zhang, X.W.; Xu, X.L.; Jiang, M.H.; Xu, K.P.; Piao, J.H.; Zhu, L.; Chen, J.; Jiang, J.G. Antioxidant and anticomplement functions of flavonoids extracted from Penthorum chinense Pursh. Food Funct. 2013, 4, 1811–1818. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.P.; Song, Y.Y.; Zhou, Y.X.; Liu, J.L.; He, S.; Zhang, D.Y. Experiment research about resistant effects of Penthorum chinense on nonalcoholic fatty liver. Chin. J. Exp. Tradit. Med. Form. 2014, 20, 125–129. [Google Scholar]
- Chen, X.R.; Yao, H.; Jiang, Y.; Wu, S.M.; Zhu, X.F.; Zhou, X.Q.; Cai, Y.M.; Zhou, Y.H.; Chen, J.J.; Wang, L.T. Evaluate of the therapeutic efficiency of the Chinese traditional medicine Gansu granule on chronic hepatitis B. Chin. J. Hepatol. 2004, 12, 50. [Google Scholar]
- Xu, C.X.; Ni, X.Y. Clinic observation of treatment on NAFLD using Gansu granule with polyenophosphorylcholine. J. Pract. Tradit. Chin. Med. 2014, 30, 857. [Google Scholar]
- Kapoor, S. Comment on isolation and identification of compounds from Penthorum chinense Pursh with antioxidant and antihepatocarcinoma properties: Pinocembrin and its rapidly emerging neuroprotective effects. J. Agric. Food. Chem. 2013, 61, 1416. [Google Scholar] [CrossRef] [PubMed]
- Rasul, A.; Millimouno, F.M.; Ali, E.W.; Ali, M.; Li, J.; Li, X. Pinocembrin: A novel natural compound with versatile pharmacological and biological activities. Biomed. Res. Int. 2013, 2013, 379850. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Jiang, Y.; Liu, H.L.; Chen, X.Q.; Wu, X.; Zhang, D.Y. A new flavanone from the aerial parts of Penthorum chinense. Nat. Prod. Res. 2014, 28, 70–73. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.W.; Qiu, F.; Chen, X.Q.; Ba, Y.Y.; Wang, X.; Wu, X. In-vivo absorption of pinocembrin-7-O-β-d-glucoside in rats and its in-vitro biotransformation. Sci. Rep. 2016, 6, 29340. [Google Scholar] [CrossRef] [PubMed]
- Kang, O.H.; Kim, S.B.; Seo, Y.S.; Joung, D.K.; Mun, S.H.; Choi, J.G.; Lee, Y.M.; Kang, D.G.; Lee, H.S. Kwon DY Curcumin decreases oleic acid-induced lipid accumulation via AMPK phosphorylation in hepatocarcinoma cells. Eur. Rev. Med. Pharmacol. Sci. 2013, 17, 2578–2586. [Google Scholar] [PubMed]
- Li, Y.; Xu, S.; Mihaylova, M.; Zheng, B.; Hou, X.; Jiang, B.; Park, O.; Luo, Z.; Lefai, E.; Shyy, J.Y.J.; et al. AMPK Phosphorylates and Inhibits SREBP Activity to Attenuate Hepatic Steatosis and Atherosclerosis in Diet-Induced Insulin-Resistant Mice. Cell Metab. 2007, 13, 376–388. [Google Scholar] [CrossRef] [PubMed]
- Sinclair, D.A.; Guarente, L. Unlocking the secrets of longevity genes. Sci. Am. 2006, 294, 48–51, 54–57. [Google Scholar] [CrossRef] [PubMed]
- Pisonero-Vaquero, S.; González-Gallego, J.; Sánchez-Campos, S.; García-Mediavilla, M.V. Flavonoids and related compounds in non-alcoholic fatty liver disease therapy. Curr. Med. Chem. 2015, 22, 2991–3012. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Jiang, Y.; Chen, X.; Yu, P.; Wang, M.; Wu, X.; Zhang, D. Identification and quantitation of major phenolic compounds from Penthorum chinense Pursh. by HPLC with tandem mass spectrometry and HPLC with diode array detection. J. Sep. Sci. 2015, 38, 2789–2796. [Google Scholar] [CrossRef] [PubMed]
- Kwan, H.Y.; Fong, W.F.; Yang, Z.; Yu, Z.L.; Hsiao, W.L. Inhibition of DNA-dependent protein kinase reduced palmitate and oleate-induced lipid accumulation in HepG2 cells. Eur. J. Nutr. 2013, 52, 1621–1630. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Chan, P.K.; Pan, S.Y.; Kwon, K.H.; Ye, Y.; Chu, J.H.; Fong, W.F.; Tsui, W.M.; Yu, Z.L. ERp57 is up-regulated in free fatty acids-induced steatotic L-02 cells and human nonalcoholic fatty livers. J. Cell. Biochem. 2010, 110, 1447–1456. [Google Scholar] [CrossRef] [PubMed]
- Kawano, Y.; Cohen, D.E. Mechanisms of hepatic triglyceride accumulation in non-alcoholic fatty liver disease. J. Gastroenterol. 2013, 48, 434–441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasetti, R.B.; Rajasekhar, M.D.; Kondeti, V.K.; Fatima, S.S.; Swapna, S.; Ramesh, B.; Rao, C.A. Antihyperglycemic and antihyperlipidemic activities of methanol:water (4:1) fraction isolated from aqueous extract of Syzygium alternifolium seeds in streptozotocin induced diabetic rats. Food Chem. Toxicol. 2010, 48, 1078–1084. [Google Scholar] [CrossRef] [PubMed]
- Ruskovska, T.; Bernlohr, D.A. Oxidative stress and protein carbonylation in adipose tissue-implications for insulin resistance and diabetes mellitus. J. Proteomics 2013, 92, 323–334. [Google Scholar] [CrossRef] [PubMed]
- Keshari, A.K.; Kumar, G.; Kushwaha, P.S.; Bhardwaj, M.; Kumar, P.; Rawat, A.; Kumar, D.; Prakash, A.; Ghosh, B.; Saha, S. Isolated flavonoids from Ficus racemosa stem bark possess antidiabetic, hypolipidemic and protective effects in albino Wistar rats. J. Ethnopharmacol. 2016, 181, 252–262. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.L.; Kang, C.H.; Wang, S.G.; Lee, H.M. Alpha-Lipoic acid regulates lipid metabolism through induction of sirtuin 1 (SIRT1) and activation of AMP-activated protein kinase. Diabetologia 2012, 55, 1824–1835. [Google Scholar] [CrossRef] [PubMed]
- Park, E.J.; Kim, Y.M.; Kim, H.J.; Jang, S.Y.; Oh, M.H.; Lee, D.H.; Chang, K.C. (S)YS-51, a novel isoquinoline alkaloid, attenuates obesity-associated non-alcoholic fatty liver disease in mice by suppressing lipogenesis, inflammation and coagulation. Eur. J. Pharmacol. 2016, 788, 200–209. [Google Scholar] [CrossRef] [PubMed]
- Chau, M.D.; Gao, J.; Yang, Q.; Wu, Z.; Gromada, J. Fibroblast growth factor 21 regulates energy metabolism by activating the AMPK-SIRT1-PGC-1α pathway. Proc. Natl. Acad. Sci. USA 2010, 107, 12553–12558. [Google Scholar] [CrossRef] [PubMed]
- Price, N.L.; Gomes, A.P.; Ling, A.J.; Duarte, F.V.; Martinmontalvo, A.; North, B.J.; Agarwal, B.; Ye, L.; Ramadori, G.; Teodoro, J.S.; et al. Sirt1 is required for ampk activation and the beneficial effects of resveratrol on mitochondrial function. Cell Metab. 2012, 15, 675–690. [Google Scholar] [CrossRef] [PubMed]
- Lv, Q.; Zhen, Q.; Liu, L.; Gao, R.; Yang, S.; Zhou, H.; Goswami, R.; Li, Q. AMP-kinase pathway is involved in tumor necrosis factor alpha-induced lipid accumulation in human hepatoma cells. Life Sci. 2015, 131, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Scott, J.W.; Hawley, S.A.; Green, K.A.; Anis, M.; Stewart, G.; Scullion, G.A.; Norman, D.G.; Hardie, D.G. CBS domains form energy-sensing modules whose binding of adenosine ligands is disrupted by disease mutations. J. Clin. Investig. 2004, 113, 274–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quan, H.Y.; Kim, D.Y.; Kim, S.J.; Jo, H.K.; Kim, G.W.; Chung, S.H. Betulinic acid alleviates non-alcoholic fatty liver by inhibiting SREBP1 activity via the AMPK-mTOR-SREBP signaling pathway. Biochem. Pharmacol. 2013, 85, 1330–1340. [Google Scholar] [CrossRef] [PubMed]
- Berger, J.P.; Akiyama, T.E.; Meinke, P.T. PPARs: Therapeutic targets for metabolic disease. Trends Pharmacol. Sci. 2005, 26, 244–251. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.; Zhang, X.; Zhang, L.; Bian, H.X.; Xu, N.; Bao, B.; Liu, J. Quercetin reduces obesity-associated ATM infiltration and inflammation in mice: A mechanism including AMPKalpha1/SIRT1. J. Lipid Res. 2014, 55, 363–374. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Luo, Y.; Deng, H.; Qin, S.; Tang, W.; Zeng, L.; Zhou, B. Hugan Qingzhi medication ameliorates hepatic steatosis by activating AMPK and PPARalpha pathways in L02 cells and HepG2 cells. J. Ethnopharmacol. 2014, 154, 229–239. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.T.; Xu, F.Y.; Yu, X.Z.; Shang, W.B. Research progress on therapeutic targets of active components in Chinese herbs for treatment of nonalcoholic fatty liver disease. China J. Chin. Mater. Med. 2017, 42, 1109–1112. [Google Scholar]
- Li, Z.; Feng, S.; Zhou, L.; Liu, S.; Cheng, J. NS5ATP6 modulates intracellular triglyceride content through FGF21 and independently of SIRT1 and SREBP1. Biochem. Biophys. Res. Commun. 2016, 475, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Zeng, L.; Tang, W.; Yin, J.; Feng, L.; Li, Y.; Yao, X.; Zhou, B. Alisol A 24-Acetate Prevents Hepatic Steatosis and Metabolic Disorders in HepG2 Cells. Cell. Physiol. Biochem. 2016, 40, 453–464. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.H.; Yang, J.H.; Shin, B.Y.; Seo, K.; Shin, S.M.; Cho, I.J.; Ki, S.H. Resveratrol inhibits LXRalpha-dependent hepatic lipogenesis through novel antioxidant Sestrin2 gene induction. Toxicol. Appl. Pharmacol. 2013, 271, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.S.; Wu, Q.; Wang, C.Q.; Wang, X.N.; Huang, J.; Zhao, J.J.; Mao, S.S.; Zhang, G.H.; Xu, X.C.; Zhang, N. miR-340 inhibition of breast cancer cell migration and invasion through targeting of oncoprotein c-Met. Cancer 2011, 117, 2842–2852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spasic, A.; Serafini, J.; Mathews, D.H. The Amber ff99 Force Field Predicts Relative Free Energy Changes for RNA Helix Formation. J. Chem. Theory. Comput. 2012, 8, 2497–2505. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Targets | Ligand | Docking Score | H-Bonds | Residue of Hydrogen Bond | Targets | Ligand | Docking Score | H-Bonds | Residue of Hydrogen Bond |
|---|---|---|---|---|---|---|---|---|---|
| SIRT1 (4ZZJ) | QCT | 7.4850 | 6 | GLY263, ASN465, GLU467, ARG276, GLU656 | AMPK (4ZHX) | QCT | 5.1547 | 6 | GLU94, MET93, VAL96, LEU22, GLU100 |
| PCB | 5.0529 | 3 | ALA262, SER442, GLN294 | PCB | 3.7372 | 3 | SER97, TYR95, ASP103 | ||
| PCBG | 7.2696 | 9 | ARG466, GLU467, ASN465, GLY263, ALA262, GLN345, GLN294 | PCBG | 5.6765 | 7 | VAL96, TYR95, SER97, ASP103, GLU100 | ||
| MPG | 7.0478 | 4 | SER442, ASP272, ARG274 | MPG | 6.3571 | 2 | GLU100, ASN144 | ||
| PPARα (3KDU) | QCT | 5.7407 | 1 | THR279 | FAS (5C37) | QCT | 5.3856 | 3 | ARG1917, ASN1945 |
| PCB | 3.1022 | 3 | ALA333, THR279, CYS275 | PCB | 4.4753 | 1 | ILE1946 | ||
| PCBG | 4.8008 | 4 | TYR464, PHE273, PHE351, MET355 | PCBG | 7.2480 | 4 | ARG1917, VAL1973, GLY1895, PHE1896 | ||
| MPG | 6.5830 | 5 | TYR464, MET355, CYS276, GLU269, PHE273 | MPG | 8.1074 | 5 | ARG1917, VAL1973, PHE1896, GLY1895, GLY1897 | ||
| ACC1 (3TVU) | QCT | 4.6247 | 8 | SER1999, ARG1954, ARG1996, PHE1956, GLY1958, MET1963 | SCD1 (4ZYO) | QCT | 5.5432 | 3 | LYS189, ASP156 |
| PCB | 3.3302 | 4 | ARG1996, GLY1998, SER1999, PHE1956 | PCB | 5.5185 | 2 | LYS189, ARG155 | ||
| PCBG | 4.8381 | 6 | PHE1956, SER1999, ARG1996, ARG1954, GLU2026 | PCBG | 5.8544 | 5 | LYS194, ASP156, LYS189, ASN75, ASN148 | ||
| MPG | 4.5763 | 2 | PHE1956, ARG1954 | MPG | 4.9069 | 4 | LYS194, LYS189, GLU152 |
| Gene Name | Forward Primer (5′–3′) | Reverse Primer (5′–3′) | Reference |
|---|---|---|---|
| SIRT1 | GCCAGAGTCCAAGTTTAGAAGA | CCATCAGTCCCAAATCCAG | [52] |
| AMPK | CAGGCATATGGTGGTCCATAGAG | TCATGGGATCCACCTGCAGC | [18] |
| SREBP1c | ATACCACCAGCGTCTACC | CACCAACAGCCCATTGAG | [48] |
| PPARα | AGCAAGGAAGGGTTGTGGCAAA | ATGGACTCGGAAGCAGGAAGGT | [53] |
| FAS | CGGCTCGCCCACCT | CGGGCCGCAAAGC | [48] |
| ACC | GCTGCTCGGATCACTAGTGAA | TTCTGCTATCAGTCTGTCCAG | [54] |
| SCD1 | CCTCTACTTGGAAGACGACATTCGC | GCAGCCGAGCTTTGTAAGAGCGGT | [54] |
| GAPDH | TGCACCACCAACTGCTTAGC | GGCATGGACTGTGGTCATGAG | [55] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, W.-W.; Wang, X.; Chen, X.-Q.; Ba, Y.-Y.; Zhang, N.; Xu, R.-R.; Zhao, W.-W.; Wu, X. Flavonones from Penthorum chinense Ameliorate Hepatic Steatosis by Activating the SIRT1/AMPK Pathway in HepG2 Cells. Int. J. Mol. Sci. 2018, 19, 2555. https://doi.org/10.3390/ijms19092555
Guo W-W, Wang X, Chen X-Q, Ba Y-Y, Zhang N, Xu R-R, Zhao W-W, Wu X. Flavonones from Penthorum chinense Ameliorate Hepatic Steatosis by Activating the SIRT1/AMPK Pathway in HepG2 Cells. International Journal of Molecular Sciences. 2018; 19(9):2555. https://doi.org/10.3390/ijms19092555
Chicago/Turabian StyleGuo, Wei-Wei, Xing Wang, Xiao-Qing Chen, Yin-Ying Ba, Nan Zhang, Rong-Rong Xu, Wen-Wen Zhao, and Xia Wu. 2018. "Flavonones from Penthorum chinense Ameliorate Hepatic Steatosis by Activating the SIRT1/AMPK Pathway in HepG2 Cells" International Journal of Molecular Sciences 19, no. 9: 2555. https://doi.org/10.3390/ijms19092555