Protection of Spleen Tissue of γ-ray Irradiated Mice against Immunosuppressive and Oxidative Effects of Radiation by Adenosine 5′-Monophosphate

Abstract
:1. Introduction
2. Results
2.1. Effect of 5′-AMP on Immune Organ Index
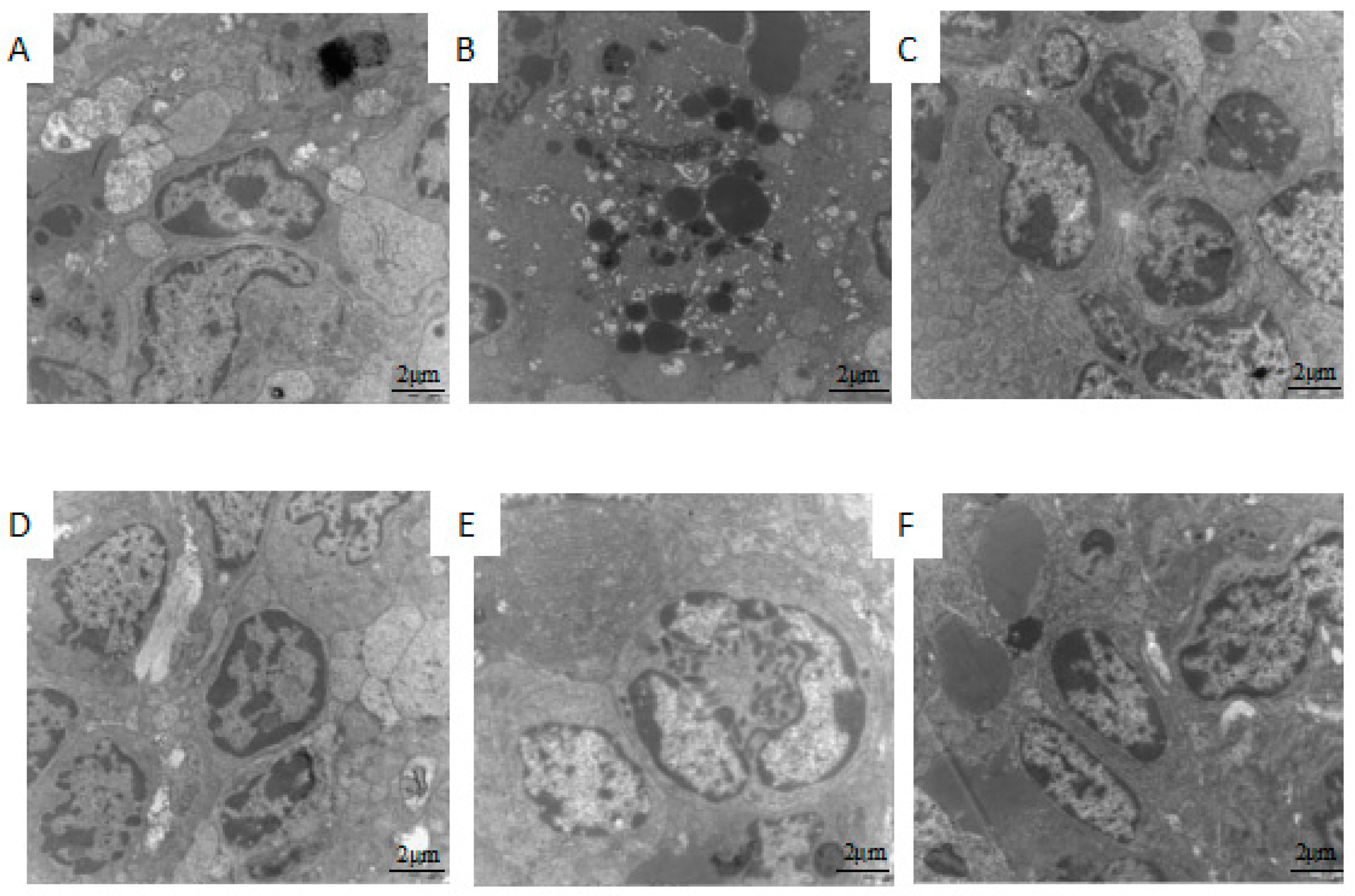
2.2. Effect of 5′-AMP on Spleen Ultra-Structure after Whole Body Irradiation of Mice
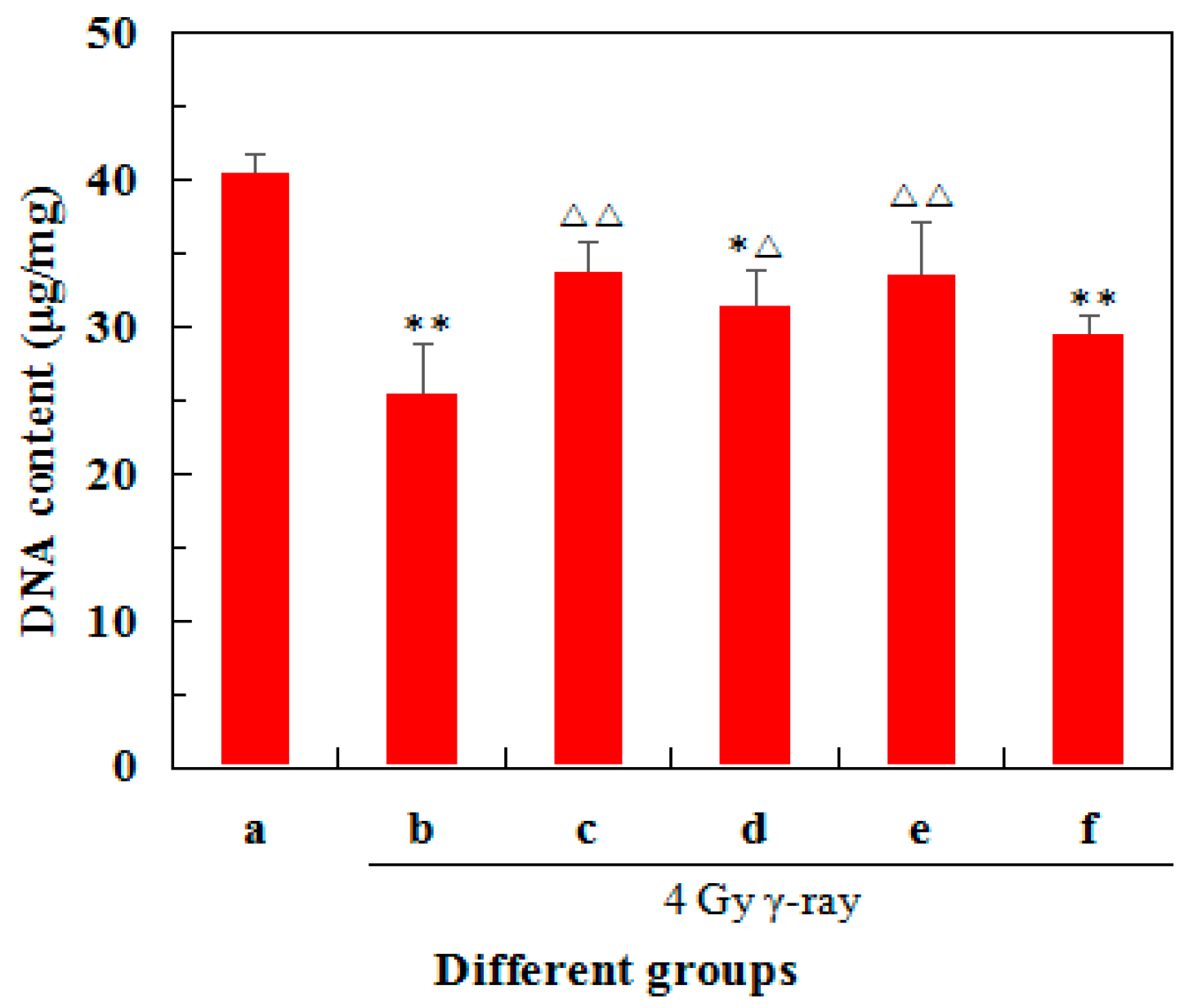
2.3. Effects of 5′-AMP on DNA Concentrations in Radiated Mice
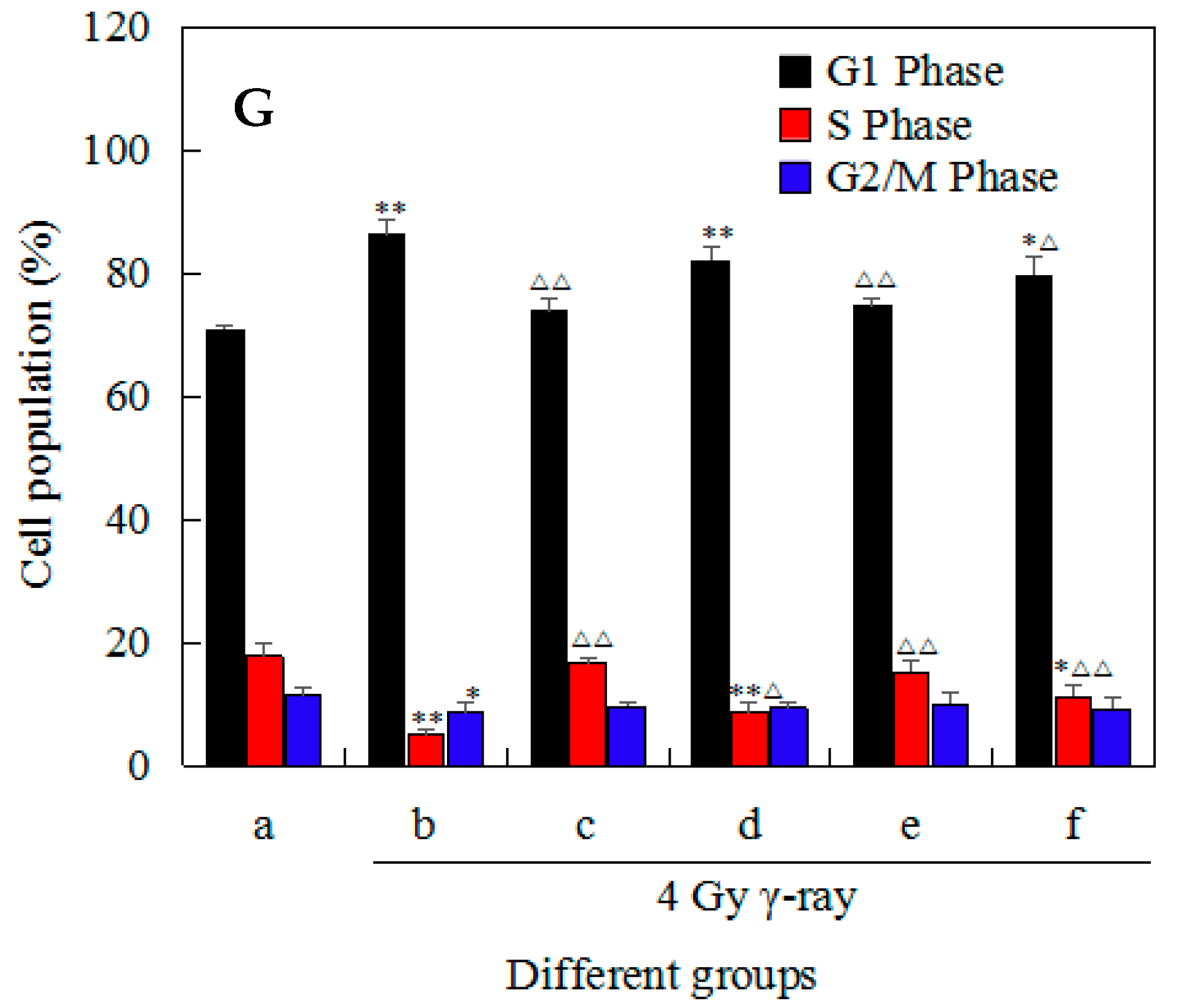
2.4. Effects of 5′-AMP on Radiation-Induced Cell-Cycle Arrest in Mouse Splenocytes
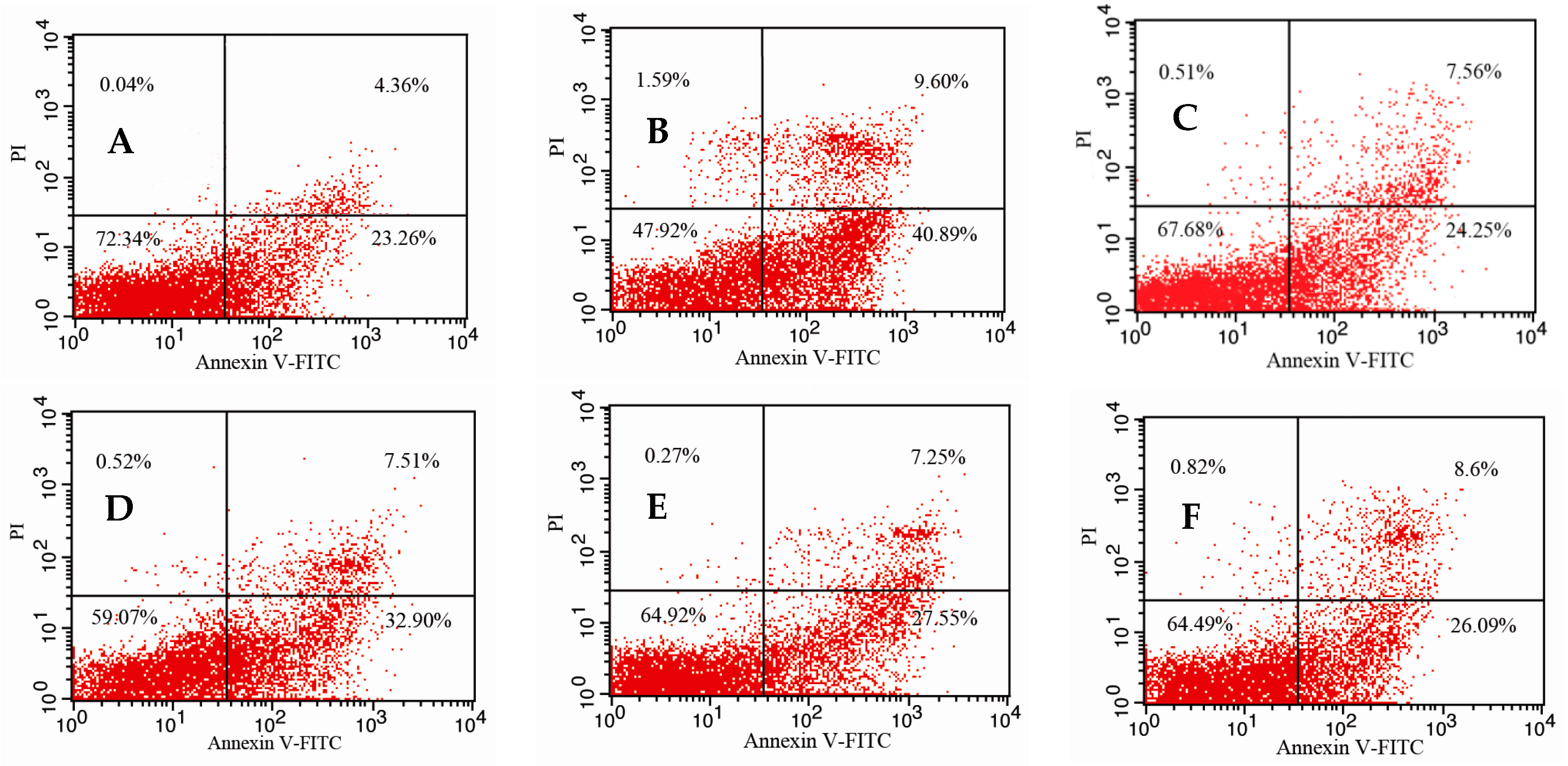
2.5. Effects of 5′-AMP on Splenocytes Apoptosis Induced by Irradiation
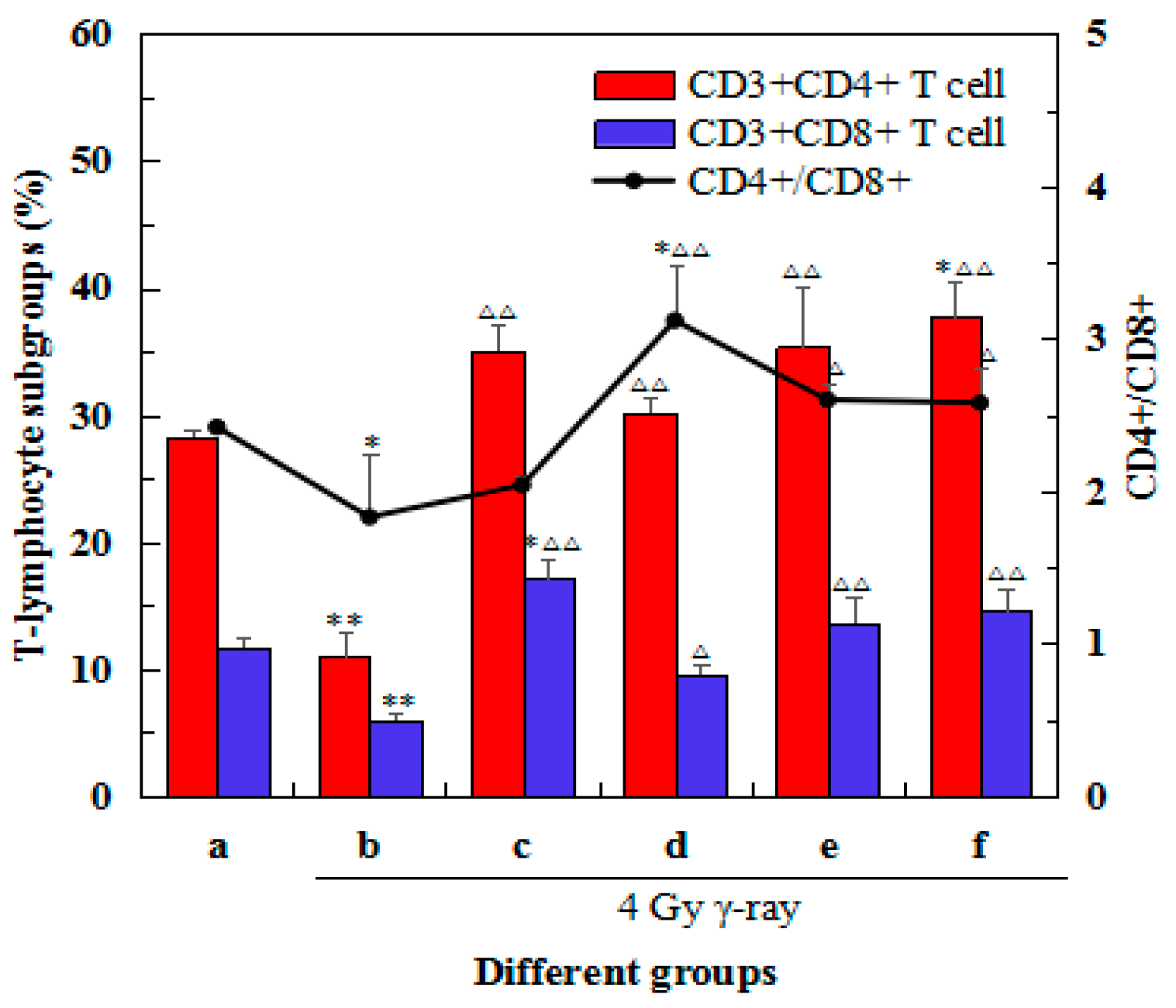
2.6. Effects of 5′-AMP on the Changes in Spleen Lymphocyte Subsets after Irradiation
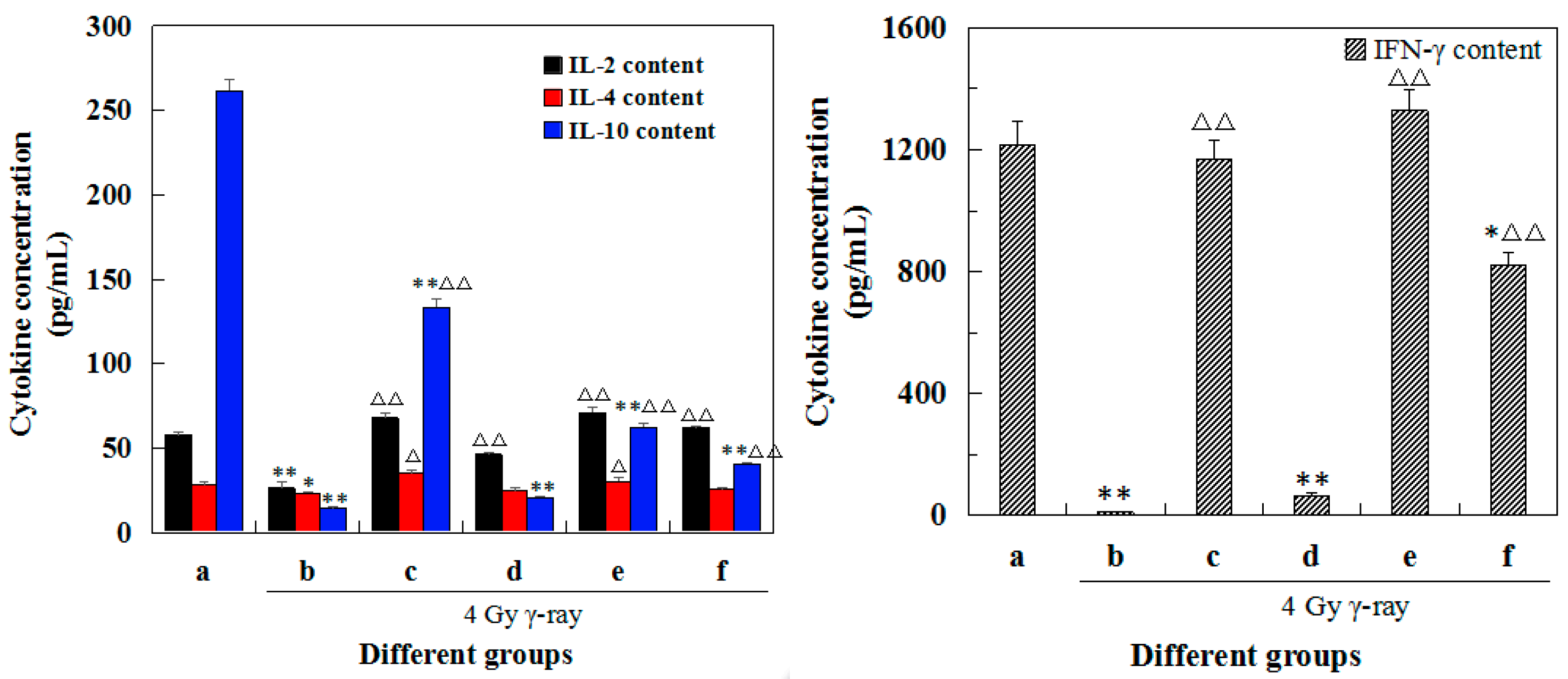
2.7. Effects of 5′-AMP on the Cytokine Concentrations in Lymphocyte Supernatants after Irradiation
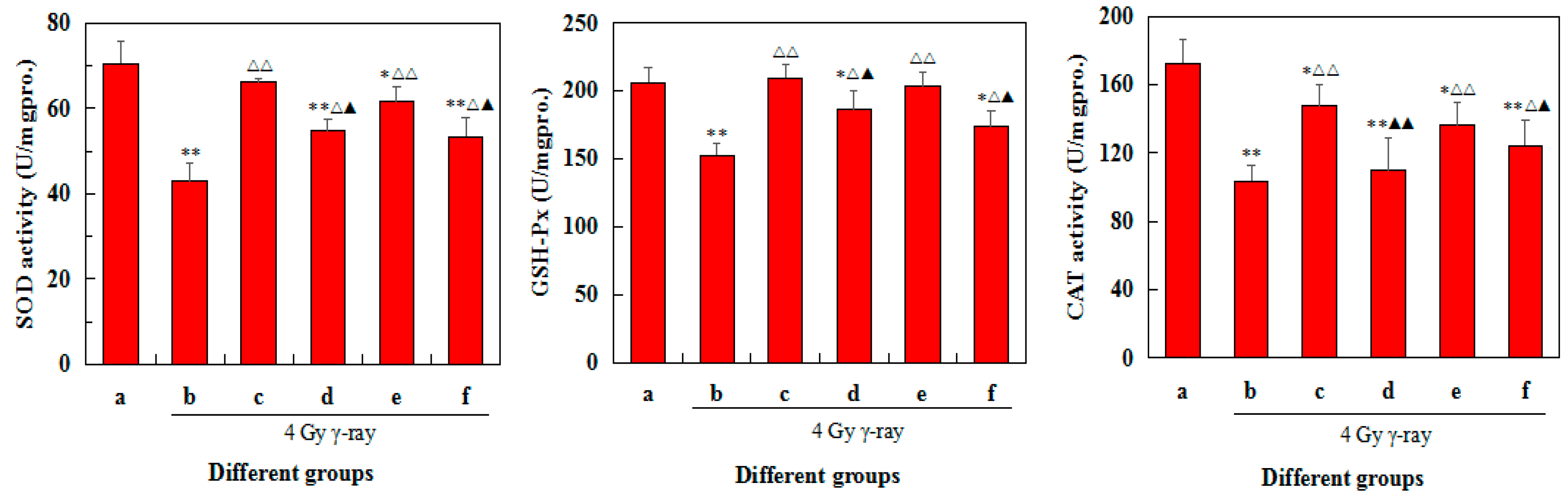
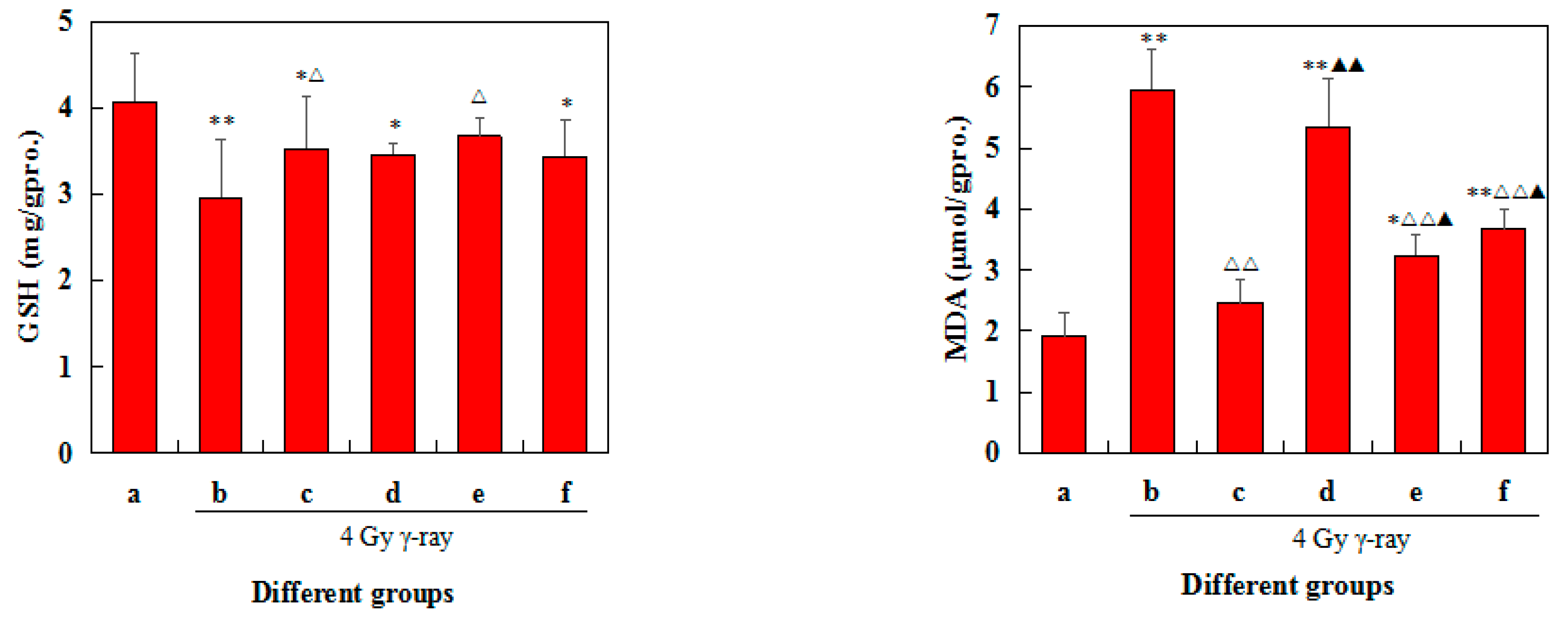
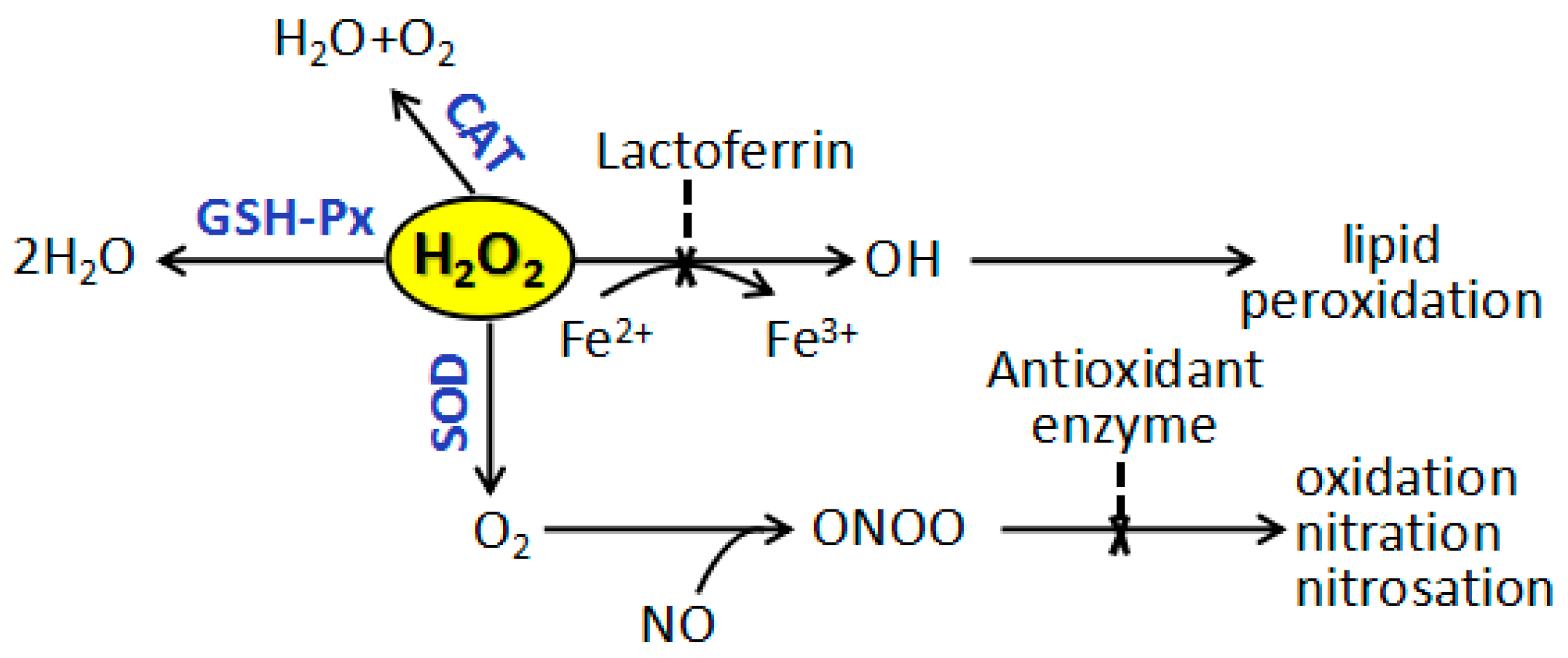
2.8. Effects of 5′-AMP on Antioxidant Defence of Spleens in Irradiated Mice
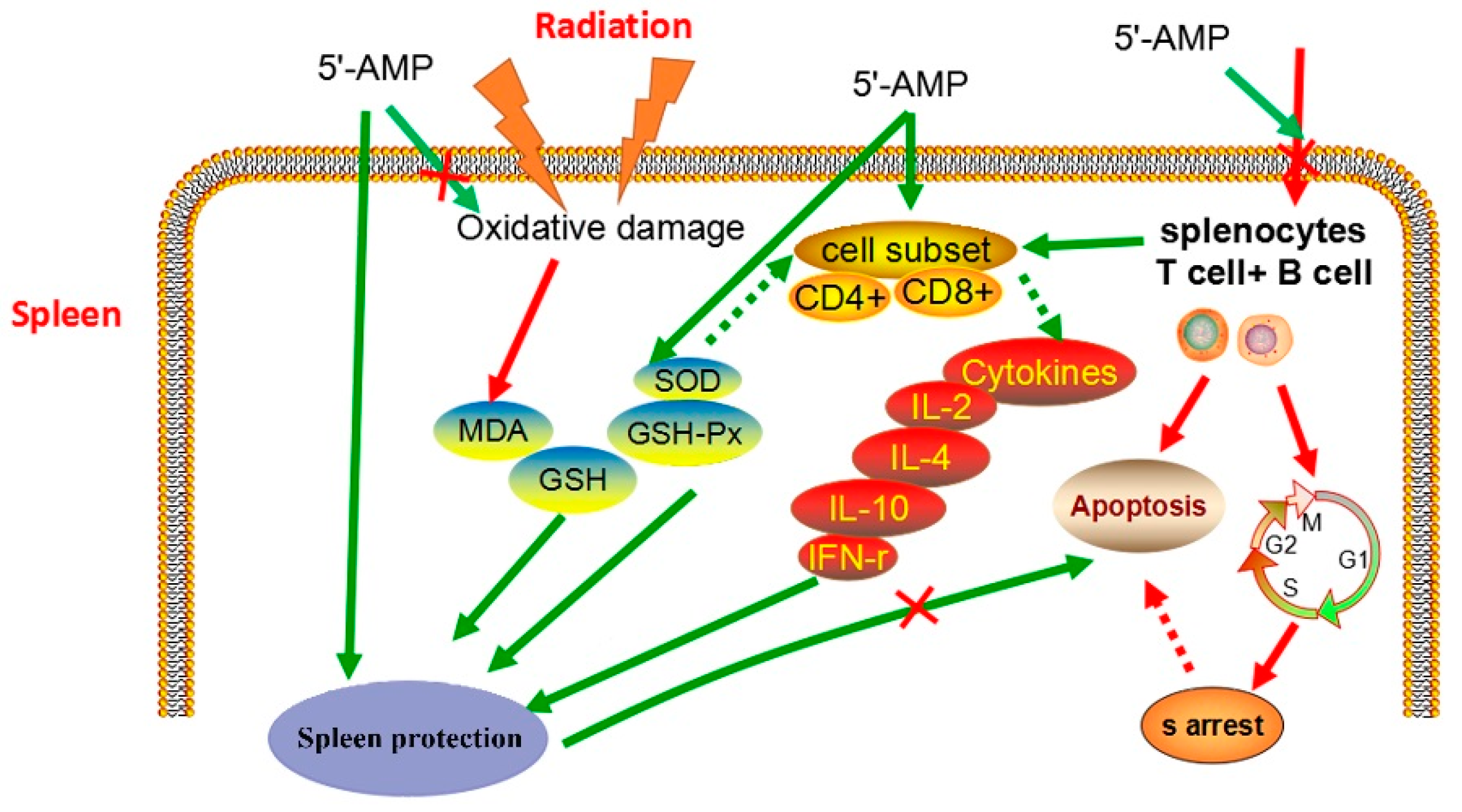
3. Discussion
4. Materials and Methods
4.1. Experimental Agents
4.2. Experimental Animals
4.3. Experimental Design
- Group I: Normal mice (Normal group)
- Group II: Radiation (Model group)
- Group III: Radiation + berberine hydrochloride (Positive control group)
- Group IV: Radiation + 5′-AMP (0.08 g/kgBW/day)
- Group V: Radiation + 5′-AMP (0.16 g/kgBW/day)
- Group VI: Radiation + 5′-AMP (0.64 g/kgBW/day)
4.4. Electron Microscopy Examination of Spleens in Mice
4.5. Measurement of Total DNA Contents in Mouse Spleen
4.6. Isolation of Lymphocytes from the Spleens of Mice
4.7. Analysis of Splenocyte Cell Cycle
4.8. Analysis of Splenocyte Apoptosis
4.9. Measurement of Cytokine Concentrations in the Supernatants of Lymphocytes
4.10. Analysis of Splenocyte Subsets
4.11. Determination of the T-SOD, GSH-Px, GSH, and MDA
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Carver, J.D.; Walker, W.A. The role of nucleotides in human nutrition. J. Nutr. Biochem. 1995, 6, 58–72. [Google Scholar] [CrossRef]
- Cosgrove, M. Nucleotides. Nutrition 1998, 14, 748–751. [Google Scholar] [CrossRef]
- Frankič, T.; Pajk, T.; Rezar, V.; Levart, A.; Salobir, J. The role of dietary nucleotides in reduction of DNA damage induced by T-2 toxin and deoxynivalenol in chicken leukocytes. Food Chem. Toxicol. 2006, 44, 1838–1844. [Google Scholar] [CrossRef] [PubMed]
- Jyonouchi, H. Nucleotide actions on humoral immune responses. J. Nutr. 1994, 124 (Suppl. S1), 138S–143S. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Muñoz, R.; de Sánchez, V.C. In vivo correlation between liver and blood energy status as evidenced by chronic treatment of carbon tetrachloride and adenosine to rats. Can. J. Physiol. Pharmacol. 1994, 72, 1252–1256. [Google Scholar] [CrossRef] [PubMed]
- Pizzini, R.P.; Kumar, S.; Kulkarni, A.D.; Rudolph, F.B.; Van Buren, C.T. Dietary nucleotides reverse malnutrition and starvation-induced immunosuppression. Arch. Surg. 1990, 125, 86–90. [Google Scholar] [CrossRef] [PubMed]
- Hu, G.; Yang, F.Q. Biological activities of nucleosides and their analogues in dietary foods. Chem. Rapid Commun. 2014, 2, 22–28. [Google Scholar]
- Guo, J.P.; Guo, B.Y.; Zhang, H.L.; Xu, W.; Zhang, W.B.; Mai, K.S. Effects of nucleotides on growth performance, immune response, disease resistance and intestinal morphology in shrimp Litopenaeus vannamei fed with a low fish meal diet. Aquac. Int. 2016, 24, 1007–1023. [Google Scholar] [CrossRef]
- Barros, M.M.; Guimaraes, I.G.; Pezzato, L.E.; de Oliveira Orsi, R.; Fernandes Junior, A.C.; Teixeira, C.P.; Fleuri, L.F.; Padovani, C.R. The effects of dietary nucleotide mixture on growth performance, haematological and immunological parameters of Nile tilapia. Aquac. Res. 2015, 46, 987–993. [Google Scholar] [CrossRef]
- Hossain, M.S.; Koshio, S.; Ishikawa, M.; Yokoyama, S.; Sony, N.M. Dietary nucleotide administration influences growth, immune responses and oxidative stress resistance of juvenile red sea bream (Pagrus major). Aquaculture 2016, 455, 41–49. [Google Scholar] [CrossRef]
- Shiaua, S.Y.; Gabaudan, J.; Lin, Y.H. Dietary nucleotide supplementation enhances immune responses and survival to Streptococcus iniae in hybrid tilapia fed diet containing low fish meal. Aquac. Rep. 2015, 2, 77–81. [Google Scholar] [CrossRef]
- Baidya, S.; Shivananda, M.H.; Jagadeesh, T.D.; Sonowal, S. Effect of nucleotide on growth, immune responses and resistance of labeo rohita to aeromonas hydrophila infection. J. Aquac. Mar. Biol. 2015, 2, 00037. [Google Scholar]
- Gil, A. Modulation of the immune response mediated by dietary nucleotides. Eur. J. Clin. Nutr. 2002, 56 (Suppl. S3), S1–S4. [Google Scholar] [CrossRef] [PubMed]
- Che, L.Q.; Hu, L.; Liu, Y.; Yan, C.; Peng, X.; Xu, Q.; Wang, R.; Cheng, Y.F.; Chen, H.; Fang, Z.F.; et al. Dietary nucleotides supplementation improves the intestinal development and immune function of neonates with intra-uterine growth restriction in a pig model. PLoS ONE 2015, 11, e0157314. [Google Scholar] [CrossRef] [PubMed]
- Holen, E.; Bjorge, O.A.; Jonsson, R. Dietary nucleotides and human immune cells. II. Modulation of PBMC growth and cytokine secretion. Nutrition 2005, 21, 1003–1009. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Ran, C.; He, S.X.; Zhang, J.L.; Hu, J.; Yang, Y.L.; Du, Z.Y.; Yang, Y.N.; Zhou, Z.G. Effects of dietary yeast nucleotides on growth, non-specific immunity, intestine growth and intestinal microbiota of juvenile hybrid tilapia Oreochromis niloticus ♀ Oreochromis aureus ♂. Anim. Nutr. 2015, 1, 244–251. [Google Scholar] [CrossRef]
- Hossain, M.S.; Koshio, S.; Ishikawa, M.; Yokoyama, S.; Sony, N.M. Dietary effects of adenosine monophosphate to enhance growth, digestibility, innate immune responses and stress resistance of juvenile red sea bream, Pagrus major. Fish Shellfish Immunol. 2016, 56, 523–533. [Google Scholar] [CrossRef] [PubMed]
- Hu, T.J.; Wei, X.J.; Zhang, X.; Cheng, F.S.; Shuai, X.H.; Zhang, L.; Kang, L. Protective effect of Potentilla anserine polysaccharide (PAP) on hydrogen peroxide induced apoptosis in murine splenic lymphocytes. Carbohydr. Polym. 2010, 79, 356–361. [Google Scholar] [CrossRef]
- Cheng, C.L.; Zhao, H.T.; Wang, Z.Y.; Lu, W.H.; Wang, L.; Wang, R.C.; Yao, L. The effect of 5′-adenylic acid on hepatic proteome of mice radiated by 60Co γ-ray. Int. J. Mol. Sci. 2014, 15, 186–202. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.L.; Wang, L.; Zhao, H.T.; Yu, W.H.; Yao, L.; Wang, Z.Y.; Ma, L.M. The rotection 5′-adenylic acid against radiation-induced oxidative stress. Heilongjiang Xumu Shouyi 2012, 5, 151–153. (In Chinese) [Google Scholar]
- Wang, L.F.; Gong, X.; Le, G.W.; Shi, Y.H. Dietary nucleotides protect thymocyte DNA from damage induced by cyclophosphamide in mice. J. Anim. Physiol. Anim. Nutr. 2008, 92, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Rudolph, F.B.; Kulkarni, A.D.; Fanslow, W.C.; Pizzini, R.P.; Kumar, S.; Van Buren, C.T. Role of RNA as a dietary source of pyrimidines and purines in immune function. Nutrition 1990, 6, 45–52. [Google Scholar] [PubMed]
- Ishikawa, T.; Kokura, S.; Sakamoto, N.; Okayama, T.; Endo, M.; Tscuchiya, R.; Okajima, M.; Matsuyama, T.; Adachi, S.; Kamada, K.; et al. Whole blood interferon-γ levels predict the therapeutic effects of adoptive T-cell therapy in patients with advanced pancreatic cancer. Int. J. Cancer 2013, 133, 1119–1125. [Google Scholar] [CrossRef] [PubMed]
- Akdis, C.A.; Joss, A.; Akdis, M.; Blaser, K. Mechanism of IL-10-induced T cell inactivation in allergic inflammation and normal response to allergens. Int. Arch. Allergy Immunol. 2012, 124, 180–182. [Google Scholar] [CrossRef] [PubMed]
- Nagafuchi, S.; Hachimura, S.; Totsuka, M.; Takahashi, T.; Goto, M.; Kuwata, T.T.; Habu, S.; Kaminogawa, S. Dietary nucleotides can up-regulate antigen-specific Th1 immune responses and suppress antigen-specific IgE responses in mice. Int. Arch. Allergy Immunol. 2000, 122, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, A.D.; Rudolph, F.B.; Van Buren, C.T. The role of dietary sources of nucleotides in immune function: A review. J. Nutr. 1994, 124, 1442S–1446S. [Google Scholar] [CrossRef] [PubMed]
- Carver, J. Dietary nucleotides: Effects on the immune and gastrointestinal systems. Acta Paediatr. 1999, 88, 83–88. [Google Scholar] [CrossRef]
- Joseph, J.; Panicker, S.N.; Janardhanan, K.K. Protective effect of polysaccharide-protein complex from a polypore mushroom, Phellinus rimosus against radiation-induced oxidative stress. Redox Rep. 2012, 17, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Wang, Z.; Ma, F.; Yang, X.; Cheng, C.; Yao, L. Protective effect of anthocyanin from Lonicera caerulea var. Edulis on radiation-induced damage in mice. Int. J. Mol. Sci. 2012, 13, 11773–11782. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.; An, H.J.; Jun, S.H.; Kim, T.J.; Lim, S.A.; Park, G.; Na, H.B.; Park, Y.; Hyeon, T.; Yee, C.; et al. Single step isolation and activation of primary CD3+ T lymphocytes using alcohol-dispersed electrospun magnetic nanofibers. Nano Lett. 2012, 12, 4018–4024. [Google Scholar] [CrossRef] [PubMed]
- Yao, L.; Wang, Z.; Zhao, H.; Cheng, C.L.; Fu, X.Y.; Liu, J.R.; Yang, X. Protective effects of polysaccharides from soybean meal against X-ray radiation induced damage in mouse spleen lymphocytes. Int. J. Mol. Sci. 2011, 12, 8096–8104. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Spleen Index |
|---|---|
| Group I: Normal group | 0.4869 ± 0.1659 |
| Group II: Model group | 0.1431 ± 0.0368 ** |
| Group III: Positive control group | 0.2214 ± 0.0556 **,Δ |
| Group IV: 5′-AMP (0.08 g/kgBW/day) | 0.1754 ± 0.0332 ** |
| Group V: 5′-AMP (0.16 g/kgBW/day) | 0.2244 ± 0.0933 **,Δ |
| Group VI: 5′-AMP (0.64 g/kgBW/day) | 0.1906 ± 0.0251 ** |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, C.; Yi, J.; Wang, R.; Cheng, L.; Wang, Z.; Lu, W. Protection of Spleen Tissue of γ-ray Irradiated Mice against Immunosuppressive and Oxidative Effects of Radiation by Adenosine 5′-Monophosphate. Int. J. Mol. Sci. 2018, 19, 1273. https://doi.org/10.3390/ijms19051273
Cheng C, Yi J, Wang R, Cheng L, Wang Z, Lu W. Protection of Spleen Tissue of γ-ray Irradiated Mice against Immunosuppressive and Oxidative Effects of Radiation by Adenosine 5′-Monophosphate. International Journal of Molecular Sciences. 2018; 19(5):1273. https://doi.org/10.3390/ijms19051273
Chicago/Turabian StyleCheng, Cuilin, Juanjuan Yi, Rongchun Wang, Li Cheng, Zhenyu Wang, and Weihong Lu. 2018. "Protection of Spleen Tissue of γ-ray Irradiated Mice against Immunosuppressive and Oxidative Effects of Radiation by Adenosine 5′-Monophosphate" International Journal of Molecular Sciences 19, no. 5: 1273. https://doi.org/10.3390/ijms19051273