Similarities and Differences between Silver Ions and Silver in Nanoforms as Antibacterial Agents

 ,
,
Abstract
:1. Introduction
2. Properties of Silver Materials vs. Antibacterial Activity
3. Mode of Antibacterial Action of Silver (MoA)
4. Current Limitations and Future Prospects of Silver Materials Usage
5. Summary and Conclusions
Acknowledgments
Conflicts of Interest
Abbreviations
| ABC | ATP-binding cassette |
| ABNC | Active but nonculturable state |
| AgNPs | Silver nanoparticles |
| ATP | Adenosine triphosphate |
| CM | Cytoplasmic membrane |
| DNA | Deoxyribonucleic acid |
| GO | Graphene oxide |
| GSH | Glutathione |
| HME | Heavy-metal efflux |
| IM | Inner membrane |
| MATE | Multidrug and toxic compound extrusion |
| MBC | Minimal bactericidal concentration |
| MDR | Multidrug resistant |
| MFP | Periplasmic membrane fusion protein |
| MFS | Major facilitator superfamily |
| MIC | Minimal inhibitory concentration |
| MoA | Mode of antibacterial action of silver |
| MR-CNS | Methycilin-resistant coagulase-negative S. aureus |
| MRSA | Methicillin-resistant S. aureus |
| n/a | Not applicable |
| OM | Outer membrane |
| OMF | Outer membrane factor |
| OMP | Outer membrane protein |
| O2−• | Superoxide anion |
| OH• | Hydroxyl radical |
| PEG | Polyethylene glycol |
| RND | Resistance-nodulation-division |
| ROS | Reactive oxygen species |
| SMR | Small multidrug resistance |
| TEM | Transmission electron microscopy |
References
- Klasen, H.J. Historical review of the use of silver in the treatment of burns. I. Early uses. Burn. J. Int. Soc. Burn Inj. 2000, 26, 117–130. [Google Scholar] [CrossRef]
- Klasen, H.J. A historical review of the use of silver in the treatment of burns. II. Renewed interest for silver. Burns 2000, 26, 131–138. [Google Scholar] [CrossRef]
- Benli, B.; Yalın, C. The influence of silver and copper ions on the antibacterial activity and local electrical properties of single sepiolite fiber: A conductive atomic force microscopy (C-AFM) study. Appl. Clay Sci. 2017, 146, 449–456. [Google Scholar] [CrossRef]
- Sun, Z.; Fan, C.; Tang, X.; Zhao, J.; Song, Y.; Shao, Z.; Xu, L. Characterization and antibacterial properties of porous fibers containing silver ions. Appl. Surf. Sci. 2016, 387, 828–838. [Google Scholar] [CrossRef]
- Chen, R.; Ni, H.; Zhang, H.; Yue, G.; Zhan, W.; Xiong, P. A preliminary study on antibacterial mechanisms of silver ions implanted stainless steel. Vacuum 2013, 89, 249–253. [Google Scholar] [CrossRef]
- Mohiti-Asli, M.; Pourdeyhimi, B.; Loboa, E.G. Novel, silver-ion-releasing nanofibrous scaffolds exhibit excellent antibacterial efficacy without the use of silver nanoparticles. Acta Biomater. 2014, 10, 2096–2104. [Google Scholar] [CrossRef] [PubMed]
- Dong, H.; Gao, Y.; Sinko, P.J.; Wu, Z.; Xu, J.; Jia, L. The nanotechnology race between China and the United States. Nano Today 2016, 11, 7–12. [Google Scholar] [CrossRef]
- Ying, J.Y. The era of nanotechnology. Nano Today 2008, 3, 1. [Google Scholar] [CrossRef]
- Sheng, Z.; Liu, Y. Potential impacts of silver nanoparticles on bacteria in the aquatic environment. J. Environ. Manag. 2017, 191, 290–296. [Google Scholar] [CrossRef] [PubMed]
- Shang, L.; Dong, S.; Nienhaus, G.U. Ultra-small fluorescent metal nanoclusters: Synthesis and biological applications. Nano Today 2011, 6, 401–418. [Google Scholar] [CrossRef]
- Zheng, K.; Yuan, X.; Goswami, N.; Zhang, Q.; Xie, J. Recent advances in the synthesis, characterization and biomedical applications of ultrasmall thiolated silver nanoclusters. RSC Adv. 2014, 4, 60581–60596. [Google Scholar] [CrossRef]
- Banhart, F.; Kotakoski, J.; Krasheninnikov, A.V. Structural Defects in Graphene. ACS Nano 2011, 5, 26–41. [Google Scholar] [CrossRef] [PubMed]
- Kharissova, O.V.; Dias, H.V.R.; Kharisov, B.I.; Pérez, B.O.; Pérez, V.M.J. The greener synthesis of nanoparticles. Trends Biotechnol. 2013, 31, 240–248. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Kim, Y.-J.; Zhang, D.; Yang, D.-C. Biological Synthesis of Nanoparticles from Plants and Microorganisms. Trends Biotechnol. 2016, 34, 588–599. [Google Scholar] [CrossRef] [PubMed]
- Mittal, A.K.; Chisti, Y.; Banerjee, U.C. Synthesis of metallic nanoparticles using plant extracts. Biotechnol. Adv. 2013, 31, 346–356. [Google Scholar] [CrossRef] [PubMed]
- Sharma, V.K.; Yngard, R.A.; Lin, Y. Silver nanoparticles: Green synthesis and their antimicrobial activities. Adv. Colloid Interface Sci. 2009, 145, 83–96. [Google Scholar] [CrossRef] [PubMed]
- Jyoti, K.; Baunthiyal, M.; Singh, A. Characterization of silver nanoparticles synthesized using Urtica dioica Linn. leaves and their synergistic effects with antibiotics. J. Radiat. Res. Appl. Sci. 2016, 9, 217–227. [Google Scholar] [CrossRef]
- Shrivastava, S.; Bera, T.; Roy, A.; Singh, G.; Ramachandrarao, P.; Dash, D. Characterization of enhanced antibacterial effects of novel silver nanoparticles. Nanotechnology 2007, 18, 225103. [Google Scholar] [CrossRef]
- Naqvi, S.Z.H.; Kiran, U.; Ali, M.I.; Jamal, A.; Hameed, A.; Ahmed, S.; Ali, N. Combined efficacy of biologically synthesized silver nanoparticles and different antibiotics against multidrug-resistant bacteria. Int. J. Nanomed. 2013, 8, 3187–3195. [Google Scholar] [CrossRef] [PubMed]
- Pal, S.; Tak, Y.K.; Song, J.M. Does the antibacterial activity of silver nanoparticles depend on the shape of the nanoparticle? A study of the Gram-negative bacterium Escherichia coli. Appl. Environ. Microbiol. 2007, 73, 1712–1720. [Google Scholar] [CrossRef] [PubMed]
- Lok, C.-N.; Ho, C.-M.; Chen, R.; He, Q.-Y.; Yu, W.-Y.; Sun, H.; Tam, P.K.-H.; Chiu, J.-F.; Che, C.-M. Proteomic analysis of the mode of antibacterial action of silver nanoparticles. J. Proteome Res. 2006, 5, 916–924. [Google Scholar] [CrossRef] [PubMed]
- Sondi, I.; Salopek-Sondi, B. Silver nanoparticles as antimicrobial agent: A case study on E. coli as a model for Gram-negative bacteria. J. Colloid Interface Sci. 2004, 275, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Morones, J.R.; Elechiguerra, J.L.; Camacho, A.; Holt, K.; Kouri, J.B.; Ramírez, J.T.; Yacaman, M.J. The bactericidal effect of silver nanoparticles. Nanotechnology 2005, 16, 2346–2353. [Google Scholar] [CrossRef] [PubMed]
- Mohandas, A.; Krishnan, A.G.; Biswas, R.; Menon, D.; Nair, M.B. Antibacterial and cytocompatible nanotextured Ti surface incorporating silver via single step hydrothermal processing. Mater. Sci. Eng. C Mater. Biol. Appl. 2017, 75, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Jiang, X.; Zhang, J.; Lin, M.; Tang, X.; Zhang, J.; Liu, H. Synthesis and characterization of antimicrobial nanosilver/diatomite nanocomposites and its water treatment application. Appl. Surf. Sci. 2017, 396, 1760–1764. [Google Scholar] [CrossRef]
- Mokhena, T.C.; Luyt, A.S. Electrospun alginate nanofibres impregnated with silver nanoparticles: Preparation, morphology and antibacterial properties. Carbohydr. Polym. 2017, 165, 304–312. [Google Scholar] [CrossRef] [PubMed]
- Raman, G.; Park, S.J.; Sakthivel, N.; Suresh, A.K. Physico-cultural parameters during AgNPs biotransformation with bactericidal activity against human pathogens. Enzym. Microb. Technol. 2017, 100, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Chandraker, K.; Nagwanshi, R.; Jadhav, S.K.; Ghosh, K.K.; Satnami, M.L. Antibacterial properties of amino acid functionalized silver nanoparticles decorated on graphene oxide sheets. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2017, 181, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Moustafa, M.T. Removal of pathogenic bacteria from wastewater using silver nanoparticles synthesized by two fungal species. Water Sci. 2017, 31, 164–176. [Google Scholar] [CrossRef]
- Gao, H.; Yang, H.; Wang, C. Controllable preparation and mechanism of nano-silver mediated by the microemulsion system of the clove oil. Results Phys. 2017, 7, 3130–3136. [Google Scholar] [CrossRef]
- Bindhu, M.R.; Umadevi, M. Antibacterial and catalytic activities of green synthesized silver nanoparticles. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2015, 135, 373–378. [Google Scholar] [CrossRef] [PubMed]
- Fayaz, A.M.; Balaji, K.; Girilal, M.; Yadav, R.; Kalaichelvan, P.T.; Venketesan, R. Biogenic synthesis of silver nanoparticles and their synergistic effect with antibiotics: A study against gram-positive and gram-negative bacteria. Nanomed. Nanotechnol. Biol. Med. 2010, 6, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Bruins, M.R.; Kapil, S.; Oehme, F.W. Microbial resistance to metals in the environment. Ecotoxicol. Environ. Saf. 2000, 45, 198–207. [Google Scholar] [CrossRef] [PubMed]
- Roebben, G.; Rauscher, H.; Amenta, V.; Aschberger, K.; Boix Sanfeliu, A.; Calzolai, L.; Emons, H.; Gaillard, C.; Gibson, N.; Holzwarth, U.; et al. Recommendation of European Union 2011/696/UE; Publications Office of the European Union: Luxembourg, 2014. [Google Scholar]
- Parveen, R.; Shamsi, T.N.; Fatima, S. Nanoparticles-protein interaction: Role in protein aggregation and clinical implications. Int. J. Biol. Macromol. 2017, 94, 386–395. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Zhao, C.; Zhang, Y.; Yao, J.; Yang, W.; Hu, Q.; Wang, C.; Cao, C. Effect of stable antimicrobial nano-silver packaging on inhibiting mildew and in storage of rice. Food Chem. 2017, 215, 477–482. [Google Scholar] [CrossRef] [PubMed]
- Jinu, U.; Gomathi, M.; Saiqa, I.; Geetha, N.; Benelli, G.; Venkatachalam, P. Green engineered biomolecule-capped silver and copper nanohybrids using Prosopis cineraria leaf extract: Enhanced antibacterial activity against microbial pathogens of public health relevance and cytotoxicity on human breast cancer cells (MCF-7). Microb. Pathog. 2017, 105, 86–95. [Google Scholar] [CrossRef] [PubMed]
- Ballottin, D.; Fulaz, S.; Cabrini, F.; Tsukamoto, J.; Durán, N.; Alves, O.L.; Tasic, L. Antimicrobial textiles: Biogenic silver nanoparticles against Candida and Xanthomonas. Mater. Sci. Eng. C 2017, 75, 582–589. [Google Scholar] [CrossRef] [PubMed]
- Li, W.-R.; Sun, T.-L.; Zhou, S.-L.; Ma, Y.-K.; Shi, Q.-S.; Xie, X.-B.; Huang, X.-M. A comparative analysis of antibacterial activity, dynamics and effects of silver ions and silver nanoparticles against four bacterial strains. Int. Biodeterior. Biodegrad. 2017, 123, 304–310. [Google Scholar] [CrossRef]
- Jung, W.K.; Koo, H.C.; Kim, K.W.; Shin, S.; Kim, S.H.; Park, Y.H. Antibacterial Activity and Mechanism of Action of the Silver Ion in Staphylococcus aureus and Escherichia coli. Appl. Environ. Microbiol. 2008, 74, 2171–2178. [Google Scholar] [CrossRef] [PubMed]
- Sütterlin, S.; Tano, E.; Bergsten, A.; Tallberg, A.-B.; Melhus, A. Effects of silver-based wound dressings on the bacterial flora in chronic leg ulcers and its susceptibility in vitro to silver. Acta Derm. Venereol. 2012, 92, 34–39. [Google Scholar] [CrossRef] [PubMed]
- Percival, S.L.; Bowler, P.G.; Russell, D. Bacterial resistance to silver in wound care. J. Hosp. Infect. 2005, 60, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Randall, C.P.; Gupta, A.; Jackson, N.; Busse, D.; O’Neill, A.J. Silver resistance in Gram-negative bacteria: A dissection of endogenous and exogenous mechanisms. J. Antimicrob. Chemother. 2015, 70, 1037–1046. [Google Scholar] [CrossRef] [PubMed]
- Randall, C.P.; Oyama, L.B.; Bostock, J.M.; Chopra, I.; O’Neill, A.J. The silver cation (Ag+): Antistaphylococcal activity, mode of action and resistance studies. J. Antimicrob. Chemother. 2013, 68, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Yamanaka, M.; Hara, K.; Kudo, J. Bactericidal Actions of a Silver Ion Solution on Escherichia coli, Studied by Energy-Filtering Transmission Electron Microscopy and Proteomic Analysis. Appl. Environ. Microbiol. 2005, 71, 7589–7593. [Google Scholar] [CrossRef] [PubMed]
- Koebnik, R.; Locher, K.P.; Van Gelder, P. Structure and function of bacterial outer membrane proteins: Barrels in a nutshell. Mol. Microbiol. 2000, 37, 239–253. [Google Scholar] [CrossRef] [PubMed]
- Schulz, G.E. The structure of bacterial outer membrane proteins. Biochim. Biophys. Acta 2002, 1565, 308–317. [Google Scholar] [CrossRef]
- Radzig, M.A.; Nadtochenko, V.A.; Koksharova, O.A.; Kiwi, J.; Lipasova, V.A.; Khmel, I.A. Antibacterial effects of silver nanoparticles on gram-negative bacteria: Influence on the growth and biofilms formation, mechanisms of action. Colloids Surf. B Biointerfaces 2013, 102, 300–306. [Google Scholar] [CrossRef] [PubMed]
- Yen, M.R.; Peabody, C.R.; Partovi, S.M.; Zhai, Y.; Tseng, Y.H.; Saier, M.H. Protein-translocating outer membrane porins of Gram-negative bacteria. Biochim. Biophys. Acta 2002, 1562, 6–31. [Google Scholar] [CrossRef]
- Li, X.Z.; Nikaido, H.; Williams, K.E. Silver-resistant mutants of Escherichia coli display active efflux of Ag+ and are deficient in porins. J. Bacteriol. 1997, 179, 6127–6132. [Google Scholar] [CrossRef] [PubMed]
- Feng, Q.L.; Wu, J.; Chen, G.Q.; Cui, F.Z.; Kim, T.N.; Kim, J.O. A mechanistic study of the antibacterial effect of silver ions on Escherichia coli and Staphylococcus aureus. J. Biomed. Mater. Res. 2000, 52, 662–668. [Google Scholar] [CrossRef]
- Dakal, T.C.; Kumar, A.; Majumdar, R.S.; Yadav, V. Mechanistic Basis of Antimicrobial Actions of Silver Nanoparticles. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Xiu, Z.-M.; Ma, J.; Alvarez, P.J.J. Differential effect of common ligands and molecular oxygen on antimicrobial activity of silver nanoparticles versus silver ions. Environ. Sci. Technol. 2011, 45, 9003–9008. [Google Scholar] [CrossRef] [PubMed]
- Lok, C.-N.; Ho, C.-M.; Chen, R.; He, Q.-Y.; Yu, W.-Y.; Sun, H.; Tam, P.K.-H.; Chiu, J.-F.; Che, C.-M. Silver nanoparticles: Partial oxidation and antibacterial activities. J. Biol. Inorg. Chem. 2007, 12, 527–534. [Google Scholar] [CrossRef] [PubMed]
- Prabhu, S.; Poulose, E.K. Silver nanoparticles: Mechanism of antimicrobial action, synthesis, medical applications and toxicity effects. Int. Nano Lett. 2012, 2, 32. [Google Scholar] [CrossRef]
- Rai, M.K.; Deshmukh, S.D.; Ingle, A.P.; Gade, A.K. Silver nanoparticles: The powerful nanoweapon against multidrug-resistant bacteria. J. Appl. Microbiol. 2012, 112, 841–852. [Google Scholar] [CrossRef] [PubMed]
- Dubey, P.; Matai, I.; Kumar, S.U.; Sachdev, A.; Bhushan, B.; Gopinath, P. Perturbation of cellular mechanistic system by silver nanoparticle toxicity: Cytotoxic, genotoxic and epigenetic potentials. Adv. Colloid Interface Sci. 2015, 221, 4–21. [Google Scholar] [CrossRef] [PubMed]
- Ramalingam, B.; Parandhaman, T.; Das, S.K. Antibacterial Effects of Biosynthesized Silver Nanoparticles on Surface Ultrastructure and Nanomechanical Properties of Gram-Negative Bacteria viz. Escherichia coli and Pseudomonas aeruginosa. ACS Appl. Mater. Interfaces 2016, 8, 4963–4976. [Google Scholar] [CrossRef] [PubMed]
- Le Pape, H.; Solano-Serena, F.; Contini, P.; Devillers, C.; Maftah, A.; Leprat, P. Involvement of reactive oxygen species in the bactericidal activity of activated carbon fibre supporting silver; Bactericidal activity of ACF(Ag) mediated by ROS. J. Inorg. Biochem. 2004, 98, 1054–1060. [Google Scholar] [CrossRef] [PubMed]
- Joshi, N.; Ngwenya, B.T.; Butler, I.B.; French, C.E. Use of bioreporters and deletion mutants reveals ionic silver and ROS to be equally important in silver nanotoxicity. J. Hazard. Mater. 2015, 287, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Park, H.-J.; Kim, J.Y.; Kim, J.; Lee, J.-H.; Hahn, J.-S.; Gu, M.B.; Yoon, J. Silver-ion-mediated reactive oxygen species generation affecting bactericidal activity. Water Res. 2009, 43, 1027–1032. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Yin, J.-J.; Wamer, W.G.; Lo, Y.M. Mechanistic characterization of titanium dioxide nanoparticle-induced toxicity using electron spin resonance. J. Food Drug Anal. 2014, 22, 76–85. [Google Scholar] [CrossRef] [PubMed]
- Choi, O.; Hu, Z. Size dependent and reactive oxygen species related nanosilver toxicity to nitrifying bacteria. Environ. Sci. Technol. 2008, 42, 4583–4588. [Google Scholar] [CrossRef] [PubMed]
- Mandal, D.; Kumar Dash, S.; Das, B.; Chattopadhyay, S.; Ghosh, T.; Das, D.; Roy, S. Bio-fabricated silver nanoparticles preferentially targets Gram positive depending on cell surface charge. Biomed. Pharmacother. 2016, 83, 548–558. [Google Scholar] [CrossRef] [PubMed]
- Gitipour, A.; Al-Abed, S.R.; Thiel, S.W.; Scheckel, K.G.; Tolaymat, T. Nanosilver as a disinfectant in dental unit waterlines: Assessment of the physicochemical transformations of the AgNPs. Chemosphere 2017, 173, 245–252. [Google Scholar] [CrossRef] [PubMed]
- McGillicuddy, E.; Murray, I.; Kavanagh, S.; Morrison, L.; Fogarty, A.; Cormican, M.; Dockery, P.; Prendergast, M.; Rowan, N.; Morris, D. Silver nanoparticles in the environment: Sources, detection and ecotoxicology. Sci. Total Environ. 2017, 575, 231–246. [Google Scholar] [CrossRef] [PubMed]
- Xiu, Z.; Zhang, Q.; Puppala, H.L.; Colvin, V.L.; Alvarez, P.J.J. Negligible particle-specific antibacterial activity of silver nanoparticles. Nano Lett. 2012, 12, 4271–4275. [Google Scholar] [CrossRef] [PubMed]
- Sotiriou, G.A.; Meyer, A.; Knijnenburg, J.T.N.; Panke, S.; Pratsinis, S.E. Quantifying the Origin of Released Ag+ Ions from Nanosilver. Langmuir 2012, 28, 15929–15936. [Google Scholar] [CrossRef] [PubMed]
- Durán, N.; Marcato, P.D.; Conti, R.D.; Alves, O.L.; Costa, F.T.M.; Brocchi, M. Potential use of silver nanoparticles on pathogenic bacteria, their toxicity and possible mechanisms of action. J. Braz. Chem. Soc. 2010, 21, 949–959. [Google Scholar] [CrossRef]
- Izak-Nau, E.; Huk, A.; Reidy, B.; Uggerud, H.; Vadset, M.; Eiden, S.; Voetz, M.; Himly, M.; Duschl, A.; Dusinska, M.; et al. Impact of storage conditions and storage time on silver nanoparticles’ physicochemical properties and implications for their biological effects. RSC Adv. 2015, 5, 84172–84185. [Google Scholar] [CrossRef] [Green Version]
- Mathema, V.B.; Thakuri, B.C.; Sillanpää, M. Bacterial mer operon-mediated detoxification of mercurial compounds: A short review. Arch. Microbiol. 2011, 193, 837–844. [Google Scholar] [CrossRef] [PubMed]
- Martinez, J.L.; Sánchez, M.B.; Martínez-Solano, L.; Hernandez, A.; Garmendia, L.; Fajardo, A.; Alvarez-Ortega, C. Functional role of bacterial multidrug efflux pumps in microbial natural ecosystems. FEMS Microbiol. Rev. 2009, 33, 430–449. [Google Scholar] [CrossRef] [PubMed]
- Piddock, L.J.V. Multidrug-resistance efflux pumps—Not just for resistance. Nat. Rev. Microbiol. 2006, 4, 629–636. [Google Scholar] [CrossRef] [PubMed]
- Fernando, D.M.; Kumar, A. Resistance-Nodulation-Division Multidrug Efflux Pumps in Gram-Negative Bacteria: Role in Virulence. Antibiotics 2013, 2, 163–181. [Google Scholar] [CrossRef] [PubMed]
- Delmar, J.A.; Su, C.-C.; Yu, E.W. Bacterial multidrug efflux transporters. Annu. Rev. Biophys. 2014, 43, 93–117. [Google Scholar] [CrossRef] [PubMed]
- Long, F.; Su, C.-C.; Zimmermann, M.T.; Boyken, S.E.; Rajashankar, K.R.; Jernigan, R.L.; Yu, E.W. Crystal structures of the CusA efflux pump suggest methionine-mediated metal transport. Nature 2010, 467, 484–488. [Google Scholar] [CrossRef] [PubMed]
- Su, C.-C.; Long, F.; Lei, H.-T.; Bolla, J.R.; Do, S.V.; Rajashankar, K.R.; Yu, E.W. Charged amino acids (R83, E567, D617, E625, R669 and K678) of CusA are required for metal ion transport in the Cus efflux system. J. Mol. Biol. 2012, 422, 429–441. [Google Scholar] [CrossRef] [PubMed]
- Su, C.-C.; Long, F.; Yu, E.W. The Cus efflux system removes toxic ions via a methionine shuttle. Protein Sci. 2011, 20, 6–18. [Google Scholar] [CrossRef] [PubMed]
- Chacón, K.N.; Mealman, T.D.; McEvoy, M.M.; Blackburn, N.J. Tracking metal ions through a Cu/Ag efflux pump assigns the functional roles of the periplasmic proteins. Proc. Natl. Acad. Sci. USA 2014, 111, 15373–15378. [Google Scholar] [CrossRef] [PubMed]
- Franke, S.; Grass, G.; Rensing, C.; Nies, D.H. Molecular analysis of the copper-transporting efflux system CusCFBA of Escherichia coli. J. Bacteriol. 2003, 185, 3804–3812. [Google Scholar] [CrossRef] [PubMed]
- Asiani, K.R.; Williams, H.; Bird, L.; Jenner, M.; Searle, M.S.; Hobman, J.L.; Scott, D.J.; Soultanas, P. SilE is an intrinsically disordered periplasmic “molecular sponge” involved in bacterial silver resistance. Mol. Microbiol. 2016, 101, 731–742. [Google Scholar] [CrossRef] [PubMed]
- Silver, S. Bacterial silver resistance: Molecular biology and uses and misuses of silver compounds. FEMS Microbiol. Rev. 2003, 27, 341–353. [Google Scholar] [CrossRef]
- Silver, S.; Gupta, A.; Matsui, K.; Lo, J.F. Resistance to Ag(I) cations in bacteria: Environments, genes and proteins. Met.-Based Drugs 1999, 6, 315–320. [Google Scholar] [CrossRef] [PubMed]
- Finley, P.J.; Norton, R.; Austin, C.; Mitchell, A.; Zank, S.; Durham, P. Unprecedented Silver Resistance in Clinically Isolated Enterobacteriaceae: Major Implications for Burn and Wound Management. Antimicrob. Agents Chemother. 2015, 59, 4734–4741. [Google Scholar] [CrossRef] [PubMed]
- Loh, J.V.; Percival, S.L.; Woods, E.J.; Williams, N.J.; Cochrane, C.A. Silver resistance in MRSA isolated from wound and nasal sources in humans and animals. Int. Wound J. 2009, 6, 32–38. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
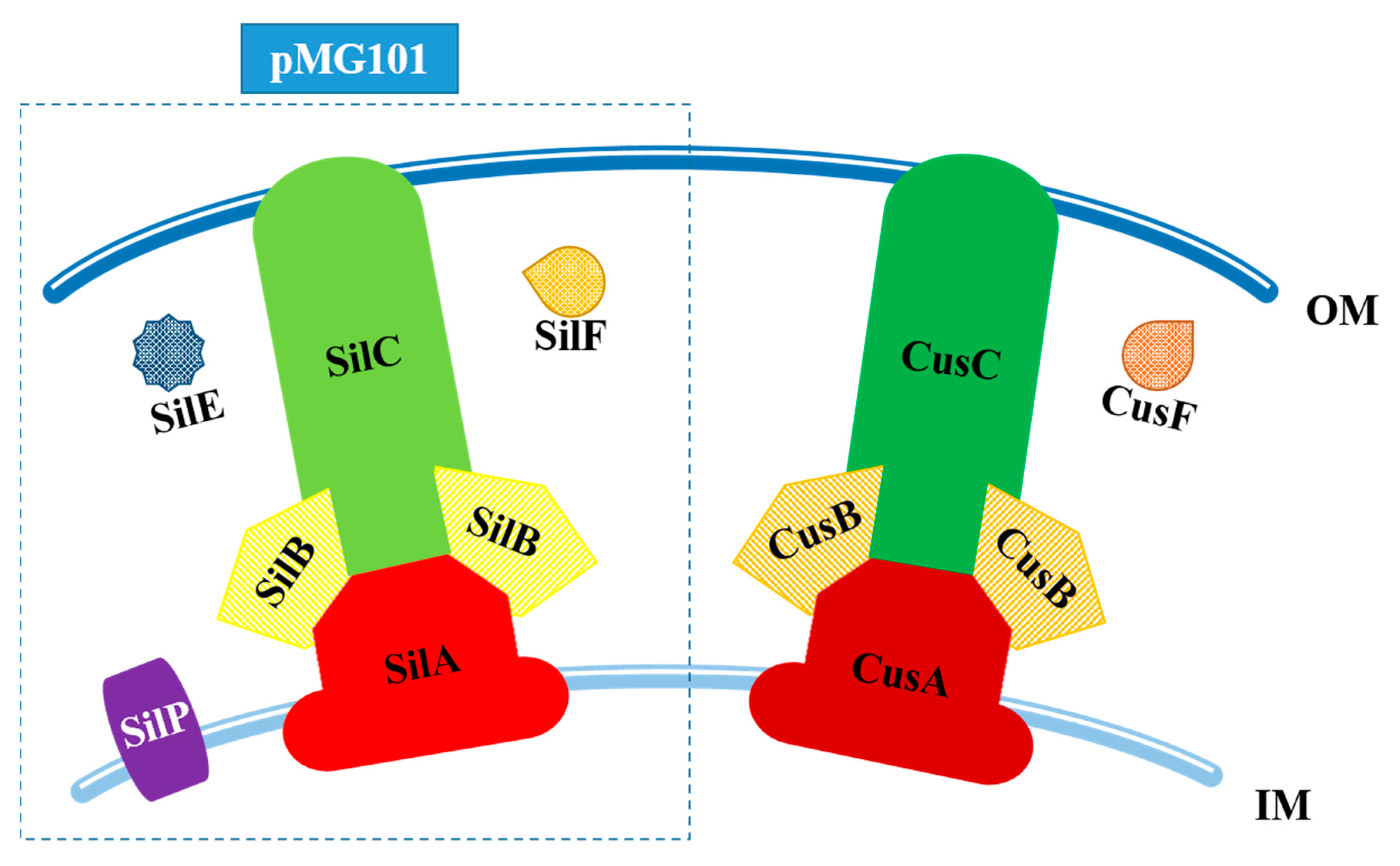{kind=link}
| Nanocomposite (Named According to the Reference) | Silver Nanoparticles Size | Silver Nanoparticles Shape | Silver Amount in Nanocomposites | Form of Compound (If Applicable) | Type of Synthesis | Antibacterial Activity | References |
|---|---|---|---|---|---|---|---|
| Silver nanoparticles | 10–15 nm | spherical, polyhedral | n/a | n/a | chemical | Antibacterial effect was dose-dependent. Tested silver nanoparticles were more effective against Gram-negative bacteria than Gram-positive; MoA: binding to the cell wall and penetrating it; modulation of cellular signalling | [18] |
| AgNPs | 5–30 nm | variable: most spherical | n/a | n/a | biological | Increased antibacterial activity of antibiotics in the presence of AgNPs; MoA: binding to proteins (by thiol groups) and DNA | [19] |
| Silver nanoparticles | 39 nm (spherical), 40 nm (triangular), 133–192 nm, diameter: 16 nm (rod-shaped) | variable: most spherical, triangular, rod-shaped | n/a | n/a | chemical | Inhibition of Escherichia coli growth on medium with silver nanoparticles; MoA: damage of bacterial cell membrane on multiple locations, formation of irregular pits | [20] |
| Nano-Ag | 9.3 ± 2.8 nm | spherical | n/a | n/a | chemical | Inhibition of E. coli growth at 6 µM concentration of Nano-Ag; MoA: changes in expression of genes encoding envelope proteins (accumulation of envelope protein precursors), destabilization of outer membrane, disturbance of proton motive force | [21] |
| Silver nanoparticles | 12 nm | spherical | n/a | n/a | chemical | Inhibition of E. coli growth at 50–60 µg/mL concentration of silver nanoparticles; MoA: damage of membranes, incorporation of silver nanoparticles into membranes, forming pits, disturbances in permeability | [22] |
| Silver nanoparticles | 16 ± 8 nm, 21 ± 18 nm | icosahedral, twinned, decahedral | n/a | agglomerated inside the carbon matrix | chemical | Inhibition of Gram-negative and Gram-positive bacteria growth at 75 μg/mL concentration of silver nanoparticles; MoA: binding to cell membrane, permeability changes, disturbances in respiration process, penetration of the bacterial membranes, interacting with DNA, releasing silver ions | [23] |
| Ti/Ag | not specified | not specified | 1.93–6.08% [m/m] | nanotexture, rutile, anatase | biological | Inhibition of S. aureus (MRSA) and E. coli growth at 15–75 µM; MoA: not specified | [24] |
| Nanosilver/diatomite | 1–20 nm | spherical particles | 0.537% [m/m] | not specified | chemical | 0.5 g nanosilver/diatomite kills above 99% of E. coli within 30 min; MoA: not specified | [25] |
| Chitosan-AgNps | 8–28 nm | spherical | 1% [m/m] | chitosan/alginate nanofibers | chemical | Inhibition of E. coli, Klebsiella pneumoniae, Bacillus pumilus and Staphylococcus aureus growth; MoA: not specified | [26] |
| AgNps | 15–160 nm (mean diameter 60 ± 10 nm) | spherical and irregular | n/a | not specified | biological | Inhibition of multidrug (MDR) pathogens: Acinetobacter baumannii, E. coli, Pseudomonas aeruginosa and Salmonella enterica growth at 25–50 µg concentration; MoA: not specified | [27] |
| GO-l-cys-AgNps | 31.5–42 nm (mean diameter 35.34 ± 0.2 nm) | spherical | not specified | graphene sheets functionalized with l-cysteine | chemical | Inhibition of E. coli growth MoA: damages of the cell membrane | [28] |
| AgNPs | 6–26 nm, 4.24–23.22 nm | spherical | n/a | foam | biological | Inhibition of the Gram-positive and Gram-negative bacteria growth at 676.9 mg/L concentration; MoA: not specified | [29] |
| Ag-NPs | 100 nm, 30 nm diameter 200–300 length | Spherical, rod-like | n/a | oil microemulsion | chemical | Inhibition of E. coli and S. aureus growth at 0.05 mg/L; MoA: not specified | [30] |
| AgNPs | 15 nm | spherical | n/a | n/a | biological | Inhibition of E. coli, S. aureus, P. aeruginosa growth at 50 µg/mL; MoA: not specified | [31] |
| AgNPs | 5–40 nm | variable: spherical or rod-like | n/a | n/a | biological | Increased antibacterial activity of ampicillin, erythromycin and chloramphenicol in the presence of AgNPs (E. coli, Salmonella Typhi, S. aureus, Micrococcus luteus); MoA: not specified | [32] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kędziora, A.; Speruda, M.; Krzyżewska, E.; Rybka, J.; Łukowiak, A.; Bugla-Płoskońska, G. Similarities and Differences between Silver Ions and Silver in Nanoforms as Antibacterial Agents. Int. J. Mol. Sci. 2018, 19, 444. https://doi.org/10.3390/ijms19020444
Kędziora A, Speruda M, Krzyżewska E, Rybka J, Łukowiak A, Bugla-Płoskońska G. Similarities and Differences between Silver Ions and Silver in Nanoforms as Antibacterial Agents. International Journal of Molecular Sciences. 2018; 19(2):444. https://doi.org/10.3390/ijms19020444
Chicago/Turabian StyleKędziora, Anna, Mateusz Speruda, Eva Krzyżewska, Jacek Rybka, Anna Łukowiak, and Gabriela Bugla-Płoskońska. 2018. "Similarities and Differences between Silver Ions and Silver in Nanoforms as Antibacterial Agents" International Journal of Molecular Sciences 19, no. 2: 444. https://doi.org/10.3390/ijms19020444