
Vegfa Impacts Early Myocardium Development in Zebrafish


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
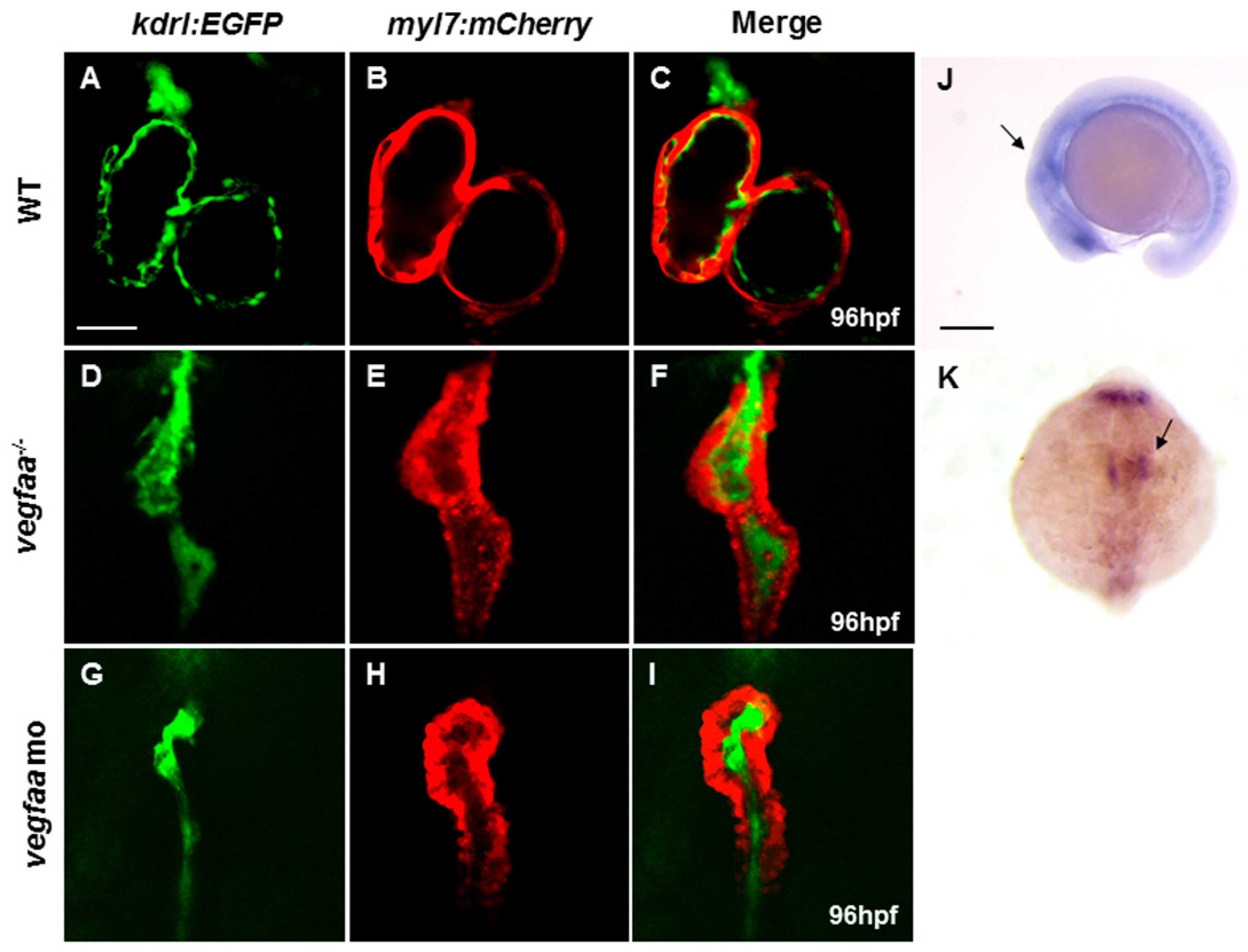
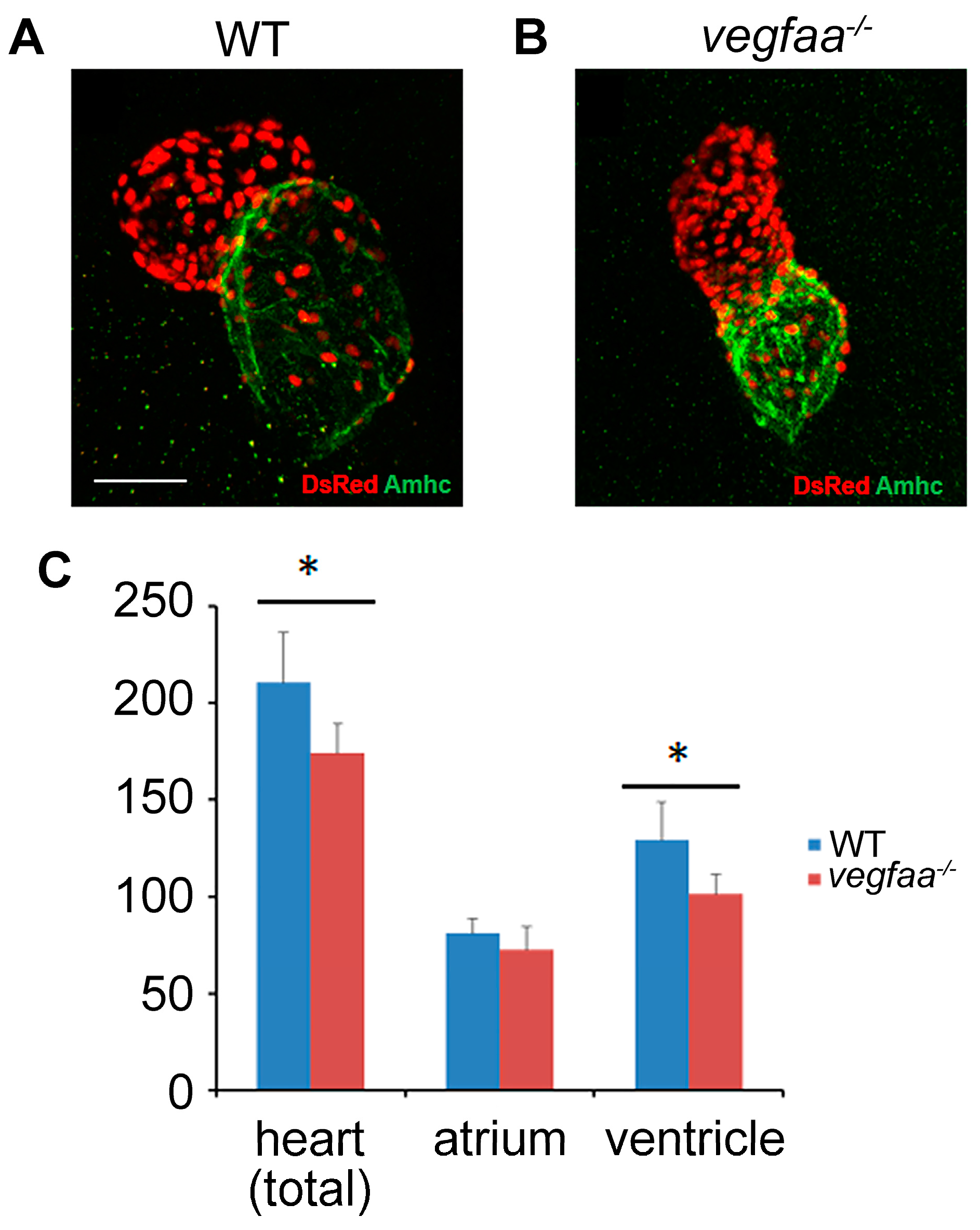
2.1. Vegfaa Deficiency Disrupts Myocardial and Endocardial Morphogenesis
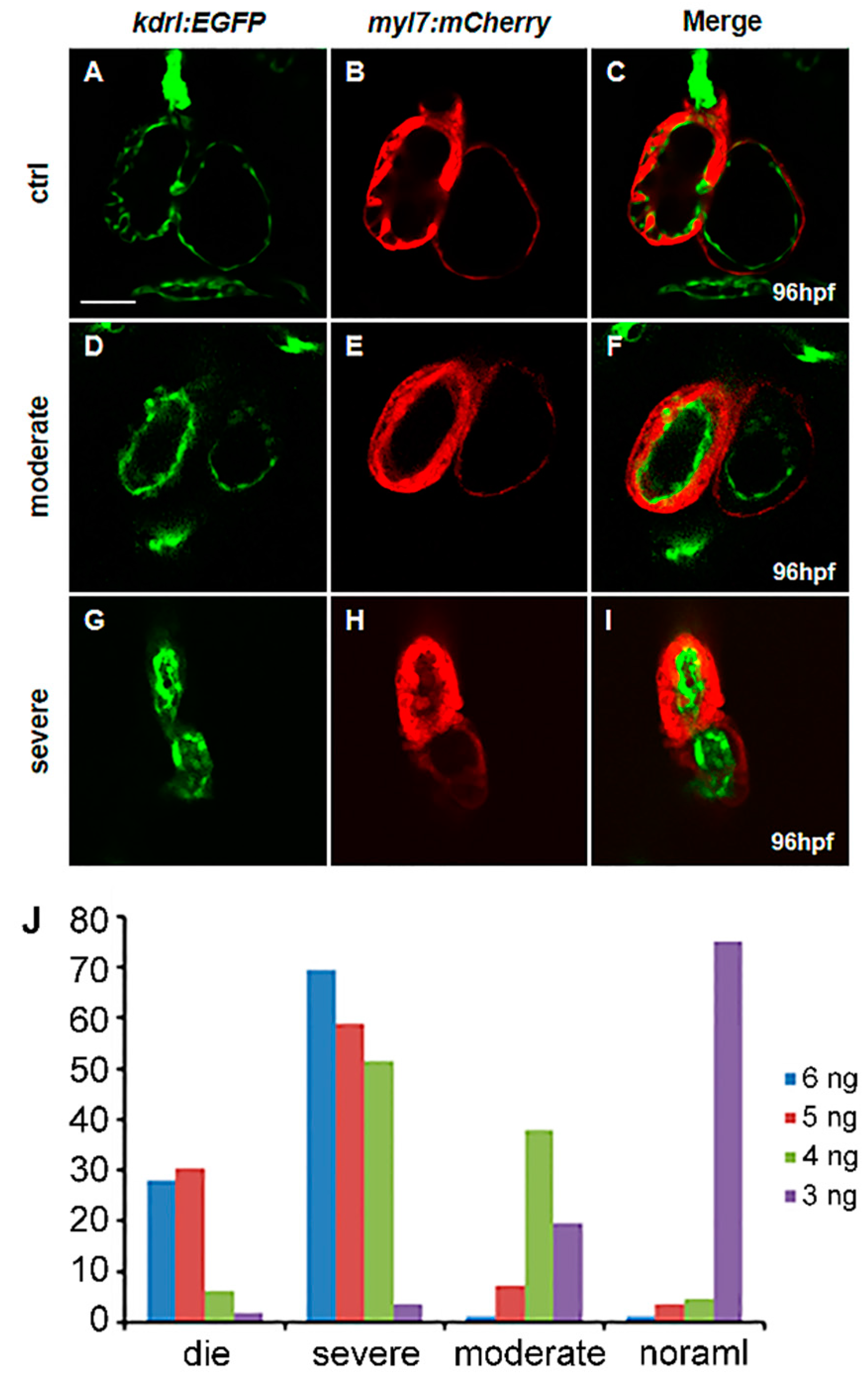
2.2. Vegfaa Regulates Heart Development in a Strict Dose-Dependent Manner
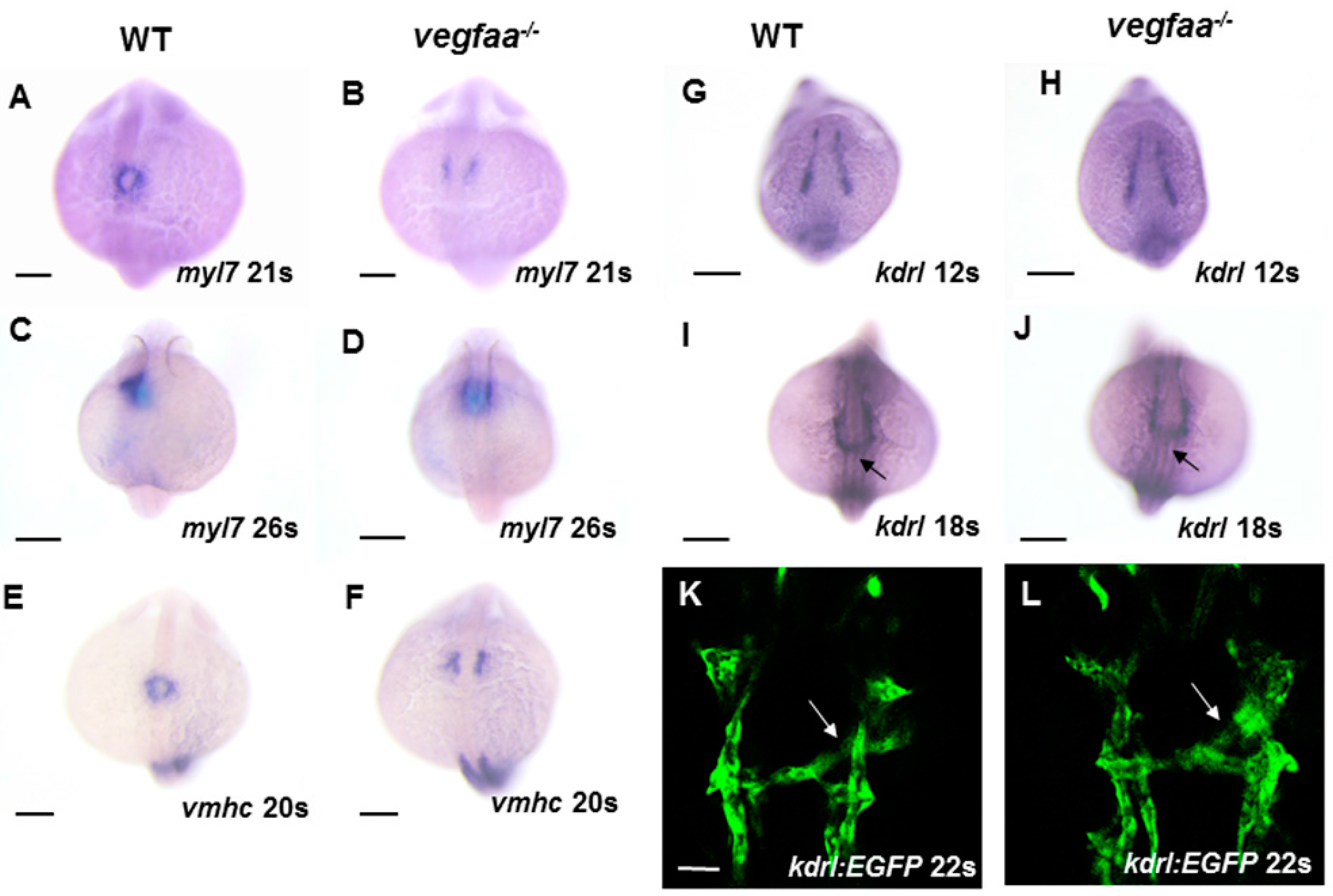
2.3. Early Myocardial/Endocardial Development in the Vegfaa Mutant
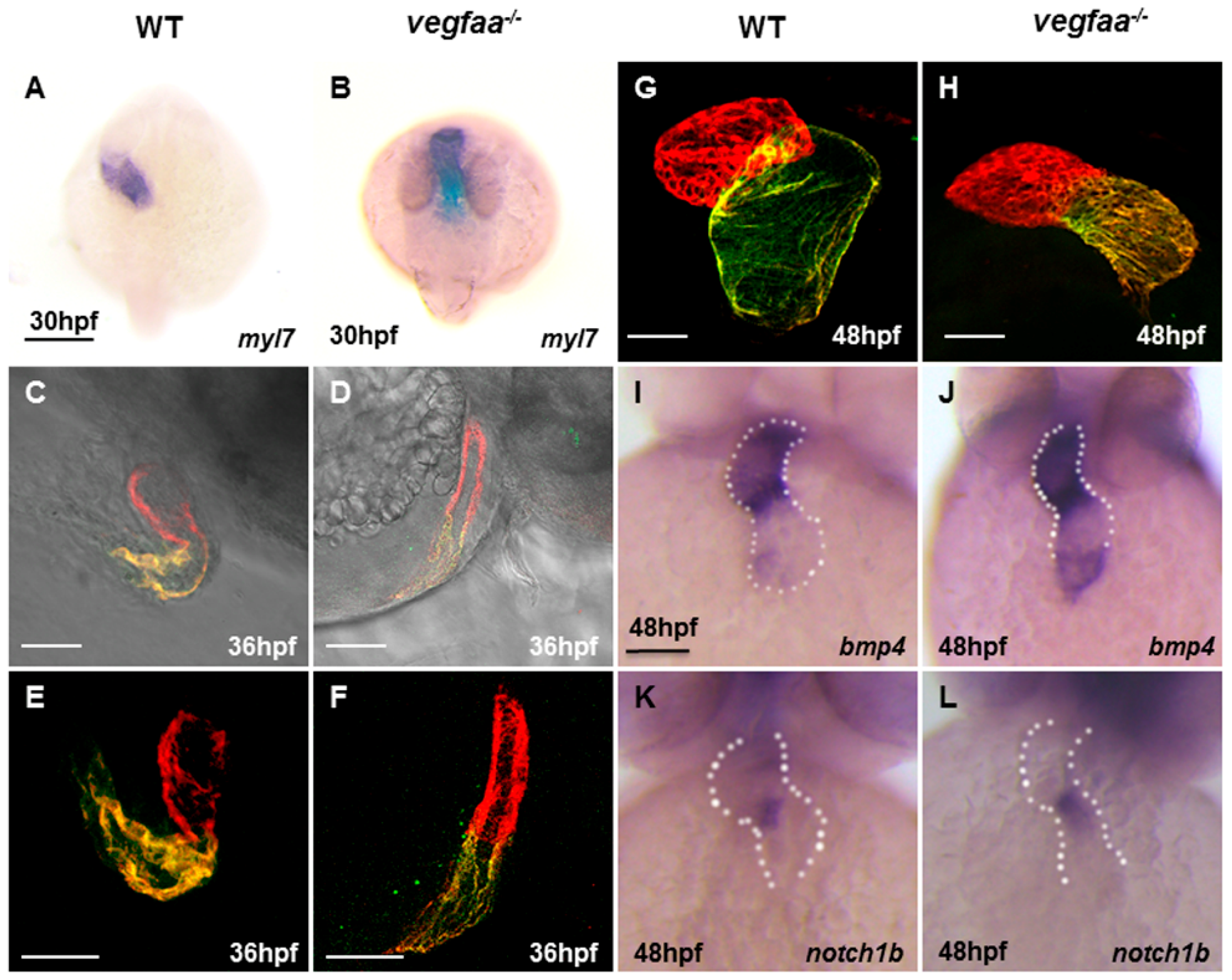
2.4. Vegfaa Deficiency Disrupts Cardiac Looping and Chamber Formation
2.5. Vegfaa Promotes Cardiomyocytes Proliferation
3. Discussion
4. Materials and Methods
4.1. Zebrafish Lines and Maintenance
4.2. Morpholino Knockdown
4.3. In Situ Hybridization and Immunohistochemistry
4.4. Cardiomyocyte Counting
4.5. Imaging
4.6. Statistical Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| CHD | Congenital heart diseases |
| ALPM | Anterior lateral plate mesoderm |
| AVC | Atrio-ventricular canal |
| MO | Morpholino oligonucleotides |
| TALEN | Transcription activator-like (TAL) effector nuclease |
| WISH | Whole-mount in situ hybridizations |
| DSHB | Developmental Studies Hybridoma Bank |
References
- Pierpont, M.E.; Basson, C.T.; Benson, D.W., Jr.; Gelb, B.D.; Giglia, T.M.; Goldmuntz, E.; McGee, G.; Sable, C.A.; Srivastava, D.; Webb, C.L. Genetic basis for congenital heart defects: Current knowledge: A scientific statement from the american heart association congenital cardiac defects committee, council on cardiovascular disease in the young: Endorsed by the american academy of pediatrics. Circulation 2007, 115, 3015–3038. [Google Scholar] [CrossRef] [PubMed]
- Fahed, A.C.; Gelb, B.D.; Seidman, J.G.; Seidman, C.E. Genetics of congenital heart disease: The glass half empty. Circ. Res. 2013, 112, 707–720. [Google Scholar] [CrossRef] [PubMed]
- Schoenebeck, J.J.; Keegan, B.R.; Yelon, D. Vessel and blood specification override cardiac potential in anterior mesoderm. Dev. Cell 2007, 13, 254–267. [Google Scholar] [CrossRef] [PubMed]
- Bruneau, B.G. Signaling and transcriptional networks in heart development and regeneration. Cold Spring Harb. Perspect. Biol. 2013, 5, a008292. [Google Scholar] [CrossRef] [PubMed]
- Ferrara, N.; Carver-Moore, K.; Chen, H.; Dowd, M.; Lu, L.; O’Shea, K.S.; Powell-Braxton, L.; Hillan, K.J.; Moore, M.W. Heterozygous embryonic lethality induced by targeted inactivation of the VEGF gene. Nature 1996, 380, 439–442. [Google Scholar] [CrossRef] [PubMed]
- Carmeliet, P.; Ferreira, V.; Breier, G.; Pollefeyt, S.; Kieckens, L.; Gertsenstein, M.; Fahrig, M.; Vandenhoeck, A.; Harpal, K.; Eberhardt, C.; et al. Abnormal blood vessel development and lethality in embryos lacking a single vegf allele. Nature 1996, 380, 435–439. [Google Scholar] [CrossRef] [PubMed]
- Stankunas, K.; Ma, G.K.; Kuhnert, F.J.; Kuo, C.J.; Chang, C.P. VEGF signaling has distinct spatiotemporal roles during heart valve development. Dev. Biol. 2010, 347, 325–336. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.P.; Neilson, J.R.; Bayle, J.H.; Gestwicki, J.E.; Kuo, A.; Stankunas, K.; Graef, I.A.; Crabtree, G.R. A field of myocardial-endocardial NFAT signaling underlies heart valve morphogenesis. Cell 2004, 118, 649–663. [Google Scholar] [CrossRef] [PubMed]
- Stalmans, I.; Lambrechts, D.; de Smet, F.; Jansen, S.; Wang, J.; Maity, S.; Kneer, P.; von der Ohe, M.; Swillen, A.; Maes, C.; et al. VEGF: A modifier of the del22q11 (DiGeorge) syndrome? Nat. Med. 2003, 9, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.M.; Cope, J.J.; Ackermann, G.E.; Goishi, K.; Armstrong, E.J.; Paw, B.H.; Bischoff, J. Vascular endothelial growth factor receptor signaling is required for cardiac valve formation in zebrafish. Dev. Dyn. 2006, 235, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Combs, M.D.; Yutzey, K.E. VEGF and RANKL regulation of NFATc1 in heart valve development. Circ. Res. 2009, 105, 565–574. [Google Scholar] [CrossRef] [PubMed]
- Johnson, E.N.; Lee, Y.M.; Sander, T.L.; Rabkin, E.; Schoen, F.J.; Kaushal, S.; Bischoff, J. NFATc1 mediates vascular endothelial growth factor-induced proliferation of human pulmonary valve endothelial cells. J. Biol. Chem. 2003, 278, 1686–1692. [Google Scholar] [CrossRef] [PubMed]
- Bahary, N.; Goishi, K.; Stuckenholz, C.; Weber, G.; Leblanc, J.; Schafer, C.A.; Berman, S.S.; Klagsbrun, M.; Zon, L.I. Duplicate VegfA genes and orthologues of the KDR receptor tyrosine kinase family mediate vascular development in the zebrafish. Blood 2007, 110, 3627–3636. [Google Scholar] [CrossRef] [PubMed]
- Liang, D.; Chang, J.R.; Chin, A.J.; Smith, A.; Kelly, C.; Weinberg, E.S.; Ge, R. The role of vascular endothelial growth factor (VEGF) in vasculogenesis, angiogenesis, and hematopoiesis in zebrafish development. Mech. Dev. 2001, 108, 29–43. [Google Scholar] [CrossRef]
- Zhu, D.; Jin, D.; Fang, Y.; Chen, Y.; Pan, W.; Liu, D.; Li, F.; Zhong, T.P. Vegfa signaling regulates diverse artery/vein formation in vertebrate vasculatures. J. Genet. Genom. 2017. under review. [Google Scholar]
- Nasevicius, A.; Larson, J.; Ekker, S.C. Distinct requirements for zebrafish angiogenesis revealed by a VEGF—A morphant. Yeast 2000, 17, 294–301. [Google Scholar] [CrossRef]
- Chen, J.N.; Fishman, M.C. Zebrafish tinman homolog demarcates the heart field and initiates myocardial differentiation. Development 1996, 122, 3809–3816. [Google Scholar] [PubMed]
- Yelon, D.; Ticho, B.; Halpern, M.E.; Ruvinsky, I.; Ho, R.K.; Silver, L.M.; Stainier, D.Y. The bHLH transcription factor hand2 plays parallel roles in zebrafish heart and pectoral fin development. Development 2000, 127, 2573–2582. [Google Scholar] [PubMed]
- Walsh, E.C.; Stainier, D.Y. UDP-glucose dehydrogenase required for cardiac valve formation in zebrafish. Science 2001, 293, 1670–1673. [Google Scholar] [CrossRef] [PubMed]
- Wong, K.S.; Rehn, K.; Palencia-Desai, S.; Kohli, V.; Hunter, W.; Uhl, J.D.; Rost, M.S.; Sumanas, S. Hedgehog signaling is required for differentiation of endocardial progenitors in zebrafish. Dev. Biol. 2012, 361, 377–391. [Google Scholar] [CrossRef] [PubMed]
- Sehnert, A.J.; Huq, A.; Weinstein, B.M.; Walker, C.; Fishman, M.; Stainier, D.Y. Cardiac troponin T is essential in sarcomere assembly and cardiac contractility. Nat. Genet. 2002, 31, 106–110. [Google Scholar] [CrossRef] [PubMed]
- Fish, J.E.; Wythe, J.D.; Xiao, T.; Bruneau, B.G.; Stainier, D.Y.; Srivastava, D.; Woo, S. A Slit/miR-218/Robo regulatory loop is required during heart tube formation in zebrafish. Development 2011, 138, 1409–1419. [Google Scholar] [CrossRef] [PubMed]
- Damert, A.; Miquerol, L.; Gertsenstein, M.; Risau, W.; Nagy, A. Insufficient VEGFA activity in yolk sac endoderm compromises haematopoietic and endothelial differentiation. Development 2002, 129, 1881–1892. [Google Scholar] [PubMed]
- Holtzman, N.G.; Schoenebeck, J.J.; Tsai, H.J.; Yelon, D. Endocardium is necessary for cardiomyocyte movement during heart tube assembly. Development 2007, 134, 2379–2386. [Google Scholar] [CrossRef] [PubMed]
- Stankunas, K.; Hang, C.T.; Tsun, Z.Y.; Chen, H.; Lee, N.V.; Wu, J.I.; Shang, C.; Bayle, J.H.; Shou, W.; Iruela-Arispe, M.L.; et al. Endocardial Brg1 represses ADAMTS1 to maintain the microenvironment for myocardial morphogenesis. Dev. Cell 2008, 14, 298–311. [Google Scholar] [CrossRef] [PubMed]
- Palencia-Desai, S.; Rost, M.S.; Schumacher, J.A.; Ton, Q.V.; Craig, M.P.; Baltrunaite, K.; Koenig, A.L.; Wang, J.; Poss, K.D.; Chi, N.C.; et al. Myocardium and BMP signaling are required for endocardial differentiation. Development 2015, 142, 2304–2315. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.W.; Beis, D.; Mitchell, T.; Chen, J.N.; Stainier, D.Y. Cellular and molecular analyses of vascular tube and lumen formation in zebrafish. Development 2005, 132, 5199–5209. [Google Scholar] [CrossRef] [PubMed]
- Palencia-Desai, S.; Kohli, V.; Kang, J.; Chi, N.C.; Black, B.L.; Sumanas, S. Vascular endothelial and endocardial progenitors differentiate as cardiomyocytes in the absence of Etsrp/Etv2 function. Development 2011, 138, 4721–4732. [Google Scholar] [CrossRef] [PubMed]
- Mably, J.D.; Mohideen, M.A.; Burns, C.G.; Chen, J.N.; Fishman, M.C. Heart of glass regulates the concentric growth of the heart in zebrafish. Curr. Biol. 2003, 13, 2138–2147. [Google Scholar] [CrossRef] [PubMed]
- Thisse, C.; Thisse, B. High-resolution in situ hybridization to whole-mount zebrafish embryos. Nat. Protoc. 2008, 3, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Alexander, J.; Stainier, D.Y.; Yelon, D. Screening mosaic F1 females for mutations affecting zebrafish heart induction and patterning. Dev. Genet. 1998, 22, 288–299. [Google Scholar] [CrossRef]





© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, D.; Fang, Y.; Gao, K.; Shen, J.; Zhong, T.P.; Li, F. Vegfa Impacts Early Myocardium Development in Zebrafish. Int. J. Mol. Sci. 2017, 18, 444. https://doi.org/10.3390/ijms18020444
Zhu D, Fang Y, Gao K, Shen J, Zhong TP, Li F. Vegfa Impacts Early Myocardium Development in Zebrafish. International Journal of Molecular Sciences. 2017; 18(2):444. https://doi.org/10.3390/ijms18020444
Chicago/Turabian StyleZhu, Diqi, Yabo Fang, Kun Gao, Jie Shen, Tao P. Zhong, and Fen Li. 2017. "Vegfa Impacts Early Myocardium Development in Zebrafish" International Journal of Molecular Sciences 18, no. 2: 444. https://doi.org/10.3390/ijms18020444