
Parasites of Selected Freshwater Snails in the Eastern Murray Darling Basin, Australia

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
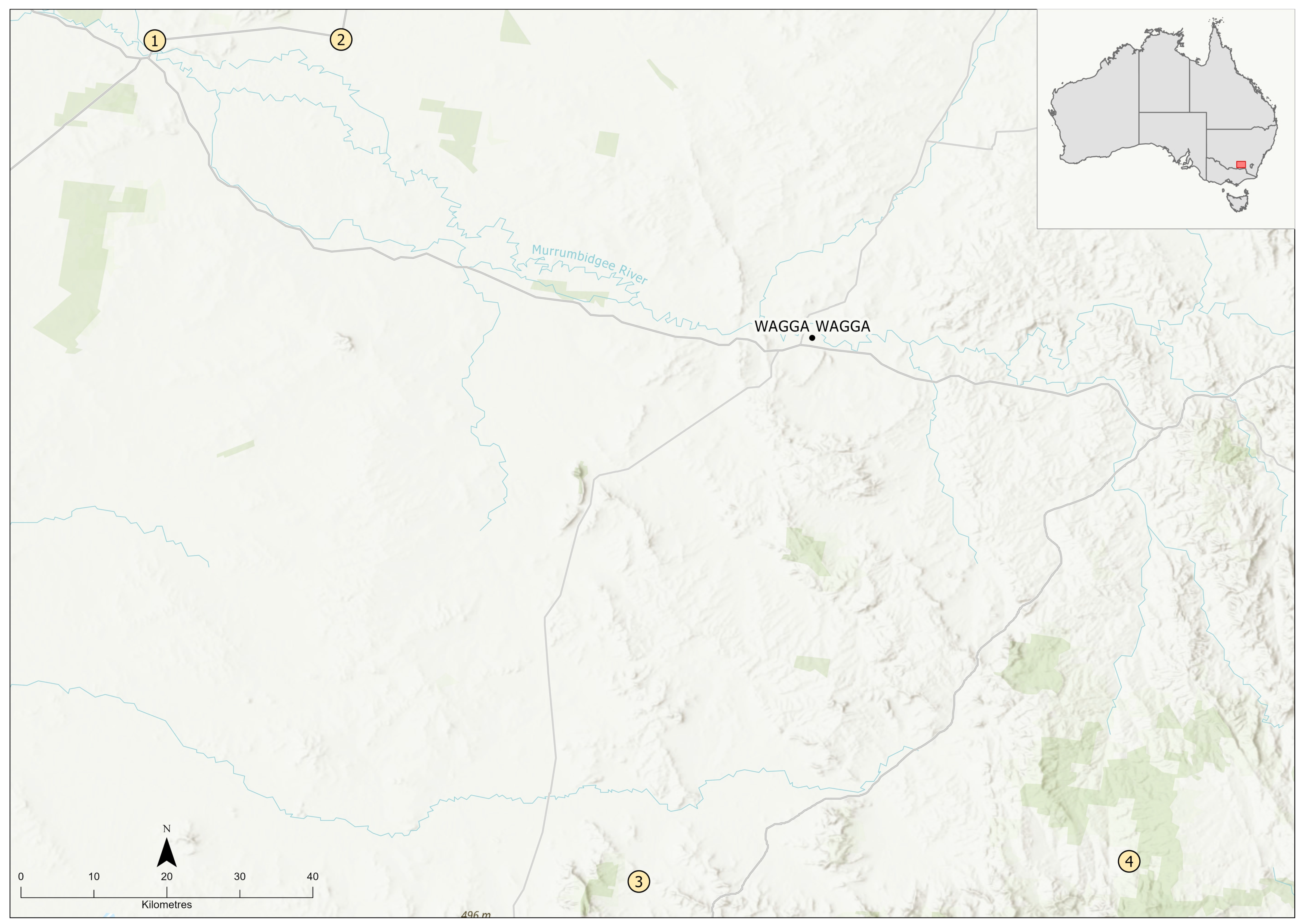
2.1. Sample Collection
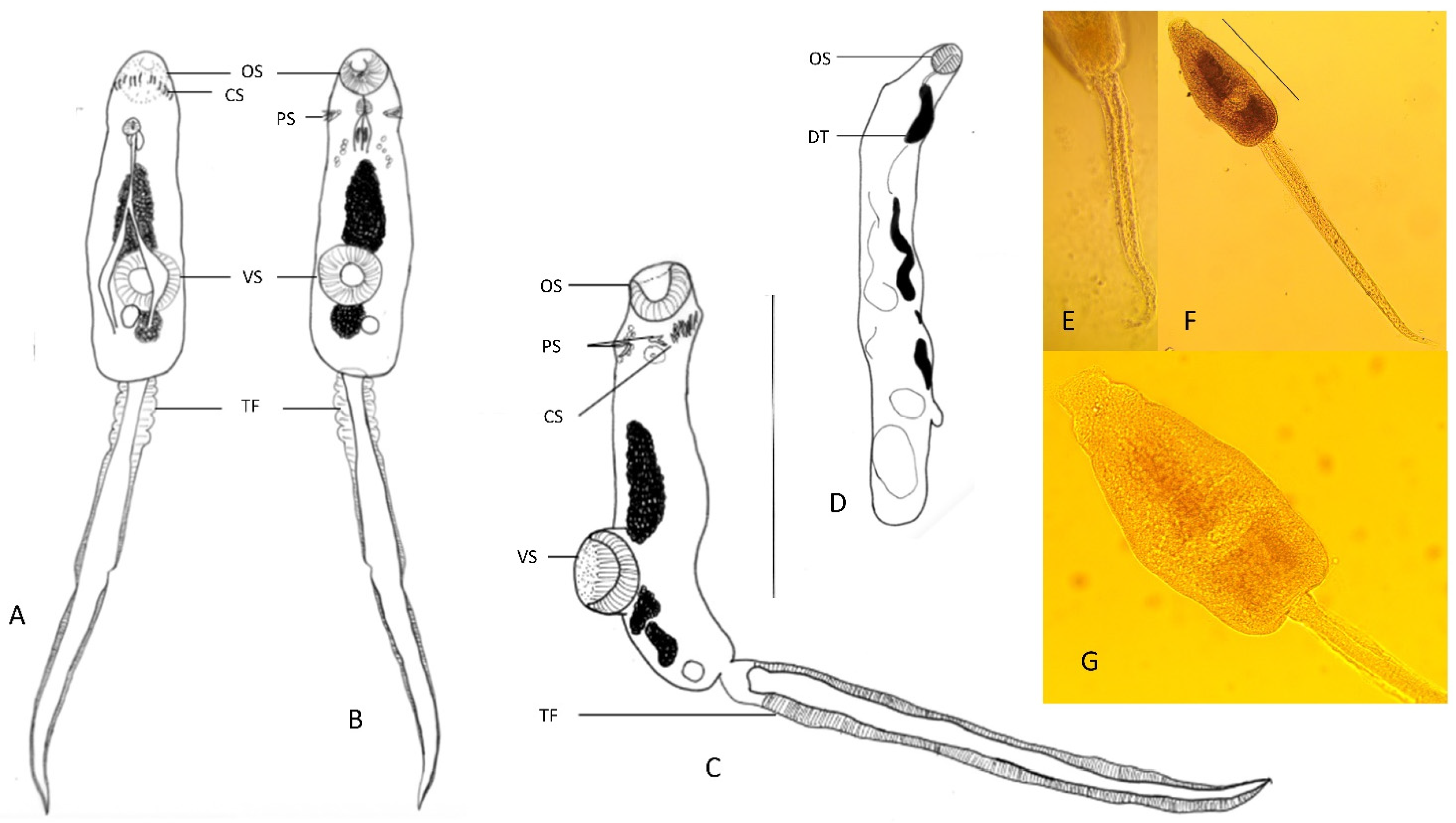
2.2. Morphology of Parasites
2.3. Molecular Diagnostics of Parasites
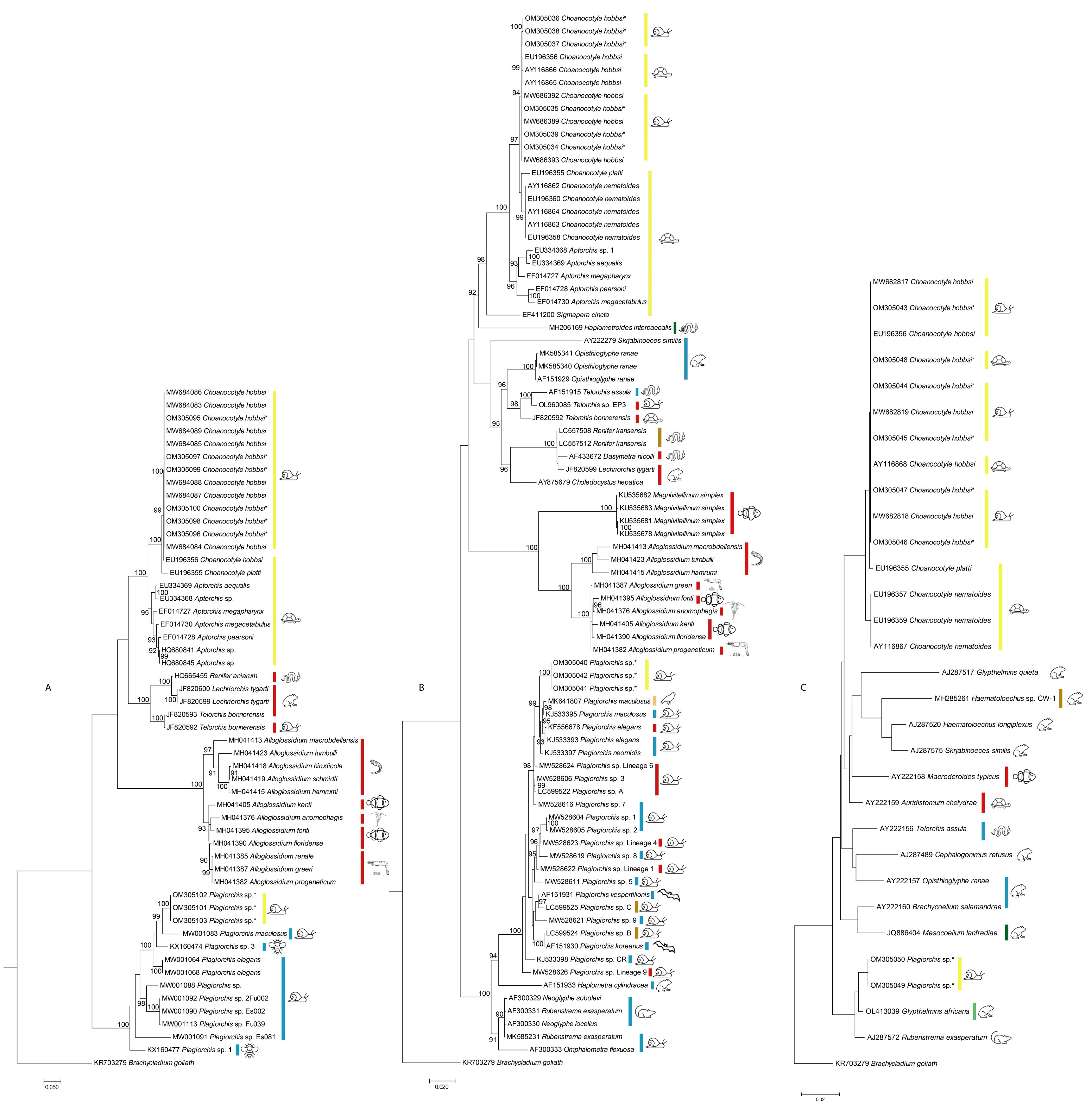
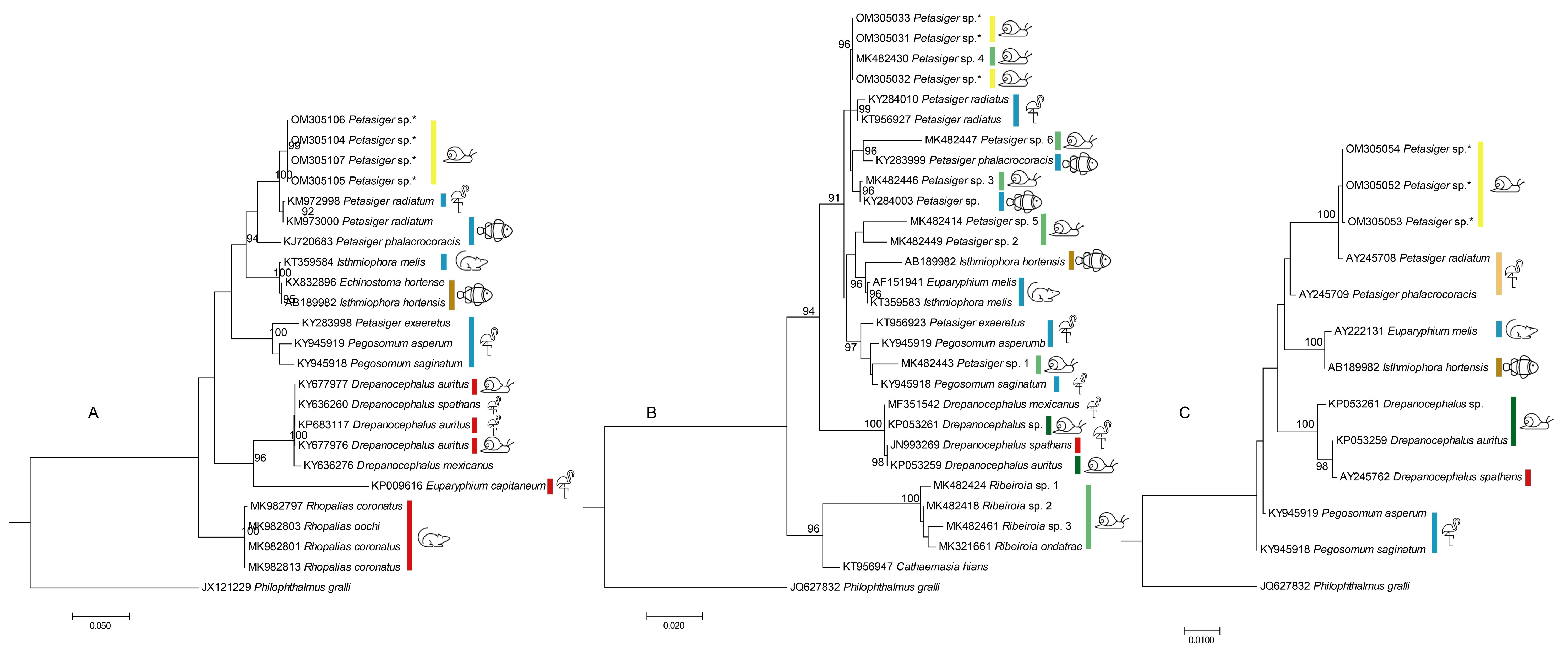
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Smith, B.J. Field Guide to the Non-Marine Molluscs of South Eastern Australia; Australian National University Press: Canberra, NSW, Australia, 1979. [Google Scholar]
- Johnson, P.D.; Bogan, A.E.; Brown, K.M.; Burkhead, N.M.; Cordeiro, J.R.; Garner, J.T.; Hartfield, P.D.; Lepitzki, D.A.; Mackie, G.L.; Pip, E. Conservation status of freshwater gastropods of Canada and the United States. Fisheries 2013, 38, 247–282. [Google Scholar] [CrossRef]
- Narr, C.F.; Krist, A.C. Improving estimates of richness, habitat associations, and assemblage characteristics of freshwater gastropods. Aquat. Conserv. Mar. Freshw. Ecosyst. 2020, 30, 131–143. [Google Scholar] [CrossRef]
- Rossini, R.A.; Fensham, R.J.; Walter, G.H. Different species requirements within a heterogeneous spring complex affects patch occupancy of threatened snails in Australian desert springs. Water 2020, 12, 2942. [Google Scholar] [CrossRef]
- Ponder, W.F. Conservation status, threats and habitat requirements of Australian terrestrial and freshwater Mollusca. Mem. Mus. Vic. 1997, 56, 421–430. [Google Scholar] [CrossRef] [Green Version]
- Zukowski, S.; Walker, K.F. Freshwater snails in competition: Alien Physa acuta (Physidae) and native Glyptophysa gibbosa (Planorbidae) in the River Murray, South Australia. Mar. Freshw. Res. 2009, 60, 999–1005. [Google Scholar] [CrossRef]
- Boray, J. The potential impact of exotic Lymnaea spp. on fascioliasis in Australasia. Vet. Prasitol. 1978, 4, 127–141. [Google Scholar] [CrossRef]
- Ponder, W. The occurrence of Lymnaea (pseudosuccinea) columella, an intermediate host of Fasciola hepatica, in Australia. Aust. Vet. J. 1975, 51, 494–495. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, D.; Baker, A.; Porter, M.; Shamsi, S.; Barton, D. Wild fallow deer (Dama dama) as definitive hosts of Fasciola hepatica (liver fluke) in alpine New South Wales. Aust. Vet. J. 2020, 98, 546–549. [Google Scholar] [CrossRef]
- Ponder, W.; Hallan, A.; Shea, M.; Clark, S.; Richards, K.; Klunzinger, M.; Kessner, V. Australian Freshwater Molluscs. Revision 1. 2020. Available online: https://keys.lucidcentral.org/keys/v3/freshwater_molluscs/ (accessed on 1 February 2020).
- Ebbs, E.T.; Loker, E.S.; Brant, S.V. Phylogeography and genetics of the globally invasive snail Physa acuta Draparnaud 1805, and its potential to serve as an intermediate host to larval digenetic trematodes. BMC Evol. Biol. 2018, 18, 103. [Google Scholar] [CrossRef]
- Taylor, D.W. Introduction to Physidae (Gastropoda: Hygrophila); biogeography, classification, morphology. Rev. Biol. Trop. 2003, 51 (Suppl. S1), 265–287. [Google Scholar]
- Shamsi, S.; Nuhoglu, A.; Zhu, X.; Barton, D.P. Genetic characterisation of cercarial stages of Choanocotyle Jue Sue and Platt, 1998 (Digenea: Choanocotylidae) in a native Australian freshwater snail, Isidorella hainesii (Tryon). Int. J. Parasitol. Parasites Wildl. 2021, 16, 48–51. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Huelsenbeck, J. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A. FigTree v1.4.2, A Graphical Viewer of Phylogenetic Trees. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 1 August 2020).
- Kasl, E.L.; Font, W.F.; Criscione, C.D. Resolving evolutionary changes in parasite life cycle complexity: Molecular phylogeny of the trematode genus Alloglossidium indicates more than one origin of precociousness. Mol. Phylogenet. Evol. 2018, 126, 371–381. [Google Scholar] [CrossRef]
- Tkach, V.V.; Snyder, S.D. Aptorchis glandularis n. sp. (Digenea: Plagiorchioidea) from the northwestern red-faced turtle, Emydura australis (Pleurodira: Chelidae) in the Kimberley, Western Australia. J. Parasitol. 2008, 94, 918–924. [Google Scholar] [CrossRef]
- Snyder, S.D.; Tkach, V.V. Aptorchis kuchlingi n. sp.(Digenea: Plagiorchioidea) from the oblong turtle, Chelodina oblonga (Pleurodira: Chelidae), in western Australia. Comp. Parasitol. 2011, 78, 280–285. [Google Scholar] [CrossRef]
- Tkach, V.V.; Snyder, S.D. Aptorchis megacetabulus n. sp. (Platyhelminthes: Digenea) from the northern long-necked turtle, Chelodina rugosa (Pleurodira: Chelidae), in Australia. J. Parasitol. 2007, 93, 404–408. [Google Scholar] [CrossRef]
- Olson, P.; Cribb, T.; Tkach, V.; Bray, R.; Littlewood, D. Phylogeny and classification of the Digenea (Platyhelminthes: Trematoda). Int. J. Parasitol. 2003, 33, 733–755. [Google Scholar] [CrossRef]
- Briscoe, A.G.; Bray, R.A.; Brabec, J.; Littlewood, D. The mitochondrial genome and ribosomal operon of Brachycladium goliath (Digenea: Brachycladiidae) recovered from a stranded minke whale. Parasitol. Int. 2016, 65, 271–275. [Google Scholar] [CrossRef]
- Tkach, V.V.; Kudlai, O.; Kostadinova, A. Molecular phylogeny and systematics of the Echinostomatoidea Looss, 1899 (Platyhelminthes: Digenea). Int. J. Parasitol. 2016, 46, 171–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cribb, T.H.; Bray, R.A.; Olson, P.D.; Pichelin, S.P.; Herniou, E.A. The Digenea. In Interrelationships of the Platyhelminthes; Littlewood, D.T.J., Bray, R.A., Eds.; Taylor & Francis: London, UK, 2001; pp. 168–185. [Google Scholar]
- Tkach, V.V.; Snyder, S.D. Choanocotyle platti sp. nov. from the northern long-necked turtle, Chelodina rugosa (Pleurodira, Chelidae) in Australia. Acta Parasitol. 2007, 52, 318–324. [Google Scholar] [CrossRef] [Green Version]
- Platt, T.R.; Tkach, V.V. Two new species of Choanocotyle Jue Sue and Platt, 1998 (Digenea: Choanocotylidae) from an Australian freshwater turtle (Testudines: Pleurodira: Chelidae). J. Parasitol. 2003, 89, 145–150. [Google Scholar] [CrossRef]
- Razo-Mendivil, U.J.; León-Regagnon, V.; Pérez-Ponce de León, G. Description of two new species of Glypthelmins Stafford, 1905 (Digenea: Macroderoididae) in Rana spp. from Mexico, based on morphology and mtDNA and rDNA sequences. Syst. Parasitol. 2004, 59, 199–210. [Google Scholar] [CrossRef]
- Tkach, V.; Grabda-Kazubska, B.; Swiderski, Z. Systematic position and phylogenetic relationships of the family Omphalometridae (Digenea, Plagiorchiida) inferred from partial lsrDNA sequences. Int. J. Parasitol. 2001, 31, 81–85. [Google Scholar] [CrossRef]
- Pinto, H.A.; Griffin, M.J.; Quiniou, S.M.; Ware, C.; Melo, A.L. Biomphalaria straminea (Mollusca: Planorbidae) as an intermediate host of Drepanocephalus spp. (Trematoda: Echinostomatidae) in Brazil: A morphological and molecular study. Parasitol. Res. 2016, 115, 51–62. [Google Scholar] [CrossRef]
- Kudlai, O.; Kostadinova, A.; Pulis, E.E.; Tkach, V.V. A new species of Drepanocephalus Dietz, 1909 (Digenea: Echinostomatidae) from the double-crested cormorant Phalacrocorax auritus (Lesson)(Aves: Phalacrocoracidae) in North America. Syst. Parasitol. 2015, 90, 221–230. [Google Scholar] [CrossRef]
- Alberson, N.R.; Rosser, T.G.; Buddenborg, S.K.; Khoo, L.H.; Loker, E.S.; Richardson, T.D.; Woodyard, E.T.; Wise, D.J.; Pote, L.M.; Griffin, M.J. North and South American haplotypes of Drepanocephalus auritus (Digenea: Echinostomatidae) are released from Biomphalaria havanensis (Mollusca: Planorbidae) inhabiting catfish aquaculture ponds in Mississippi, USA. Comp. Parasitol. 2017, 84, 87–101. [Google Scholar] [CrossRef]
- Hernández-Cruz, E.; Hernández-Orts, J.S.; Sereno-Uribe, A.; de León, G.P.-P.; García-Varela, M. Multilocus phylogenetic analysis and morphological data reveal a new species composition of the genus Drepanocephalus Dietz, 1909 (Digenea: Echinostomatidae), parasites of fish-eating birds in the Americas. J. Helminthol. 2018, 92, 572–595. [Google Scholar] [CrossRef]
- Griffin, M.J.; Khoo, L.H.; Quiniou, S.M.; O’Hear, M.M.; Pote, L.M.; Greenway, T.E.; Wise, D.J. Genetic sequence data identifies the cercaria of Drepanocephalus spathans (Digenea: Echinostomatidae), a parasite of the double-crested cormorant (Phalacrocorax auritus), with notes on its pathology in juvenile channel catfish (Ictalurus punctatus). J. Parasitol. 2012, 98, 967–972. [Google Scholar] [CrossRef]
- Qiu, J.-H.; Zhang, Y.; Zhang, X.-X.; Gao, Y.; Li, Q.; Chang, Q.-C.; Wang, C.-R. Metacercaria infection status of fishborne zoonotic trematodes, except for Clonorchis sinensis in fish from the Heilongjiang Province, China. Foodborne Pathog. Dis. 2017, 14, 440–446. [Google Scholar] [CrossRef]
- Kudlai, O.; Tkach, V.V.; Pulis, E.E.; Kostadinova, A. Redescription and phylogenetic relationships of Euparyphium capitaneum Dietz, 1909, the type-species of Euparyphium Dietz, 1909 (Digenea: Echinostomatidae). Syst. Parasitol. 2015, 90, 53–65. [Google Scholar] [CrossRef]
- Tkach, V.; Pawlowski, J.; Mariaux, J. Phylogenetic analysis of the suborder Plagiorchiata (Platyhelminthes, Digenea) based on partial lsrDNA sequences. Int. J. Parasitol. 2000, 30, 83–93. [Google Scholar] [CrossRef]
- Sinsch, U.; Balczun, C.; Scheid, P.; Dehling, J.M. Component endoparasite communities mirror life-history specialization in syntopic reed frogs (Hyperolius spp.). Diversity 2021, 13, 669. [Google Scholar] [CrossRef]
- Müller, M.I.; Morais, D.H.; da Silva, R.J. Molecular phylogenetic position of Haplometroides intercaecalis (Digenea, Plagiorchiidae). Acta Parasitol. 2018, 63, 522–526. [Google Scholar] [CrossRef]
- Sato, H.; Suzuki, K. Gastrointestinal helminths of feral raccoons (Procyon lotor) in Wakayama Prefecture, Japan. J. Vet. Med. Sci. 2006, 68, 311–318. [Google Scholar] [CrossRef] [Green Version]
- Hildebrand, J.; Adamczyk, M.; Laskowski, Z.; Zaleśny, G. Host-dependent morphology of Isthmiophora melis (Schrank, 1788) Luhe, 1909 (Digenea, Echinostomatinae)–morphological variation vs. molecular stability. Parasites Vectors 2015, 8, 481. [Google Scholar] [CrossRef] [Green Version]
- Pulis, E.E.; Tkach, V.V.; Newman, R.A. Helminth parasites of the wood frog, Lithobates sylvaticus, in prairie pothole wetlands of the Northern Great Plains. Wetlands 2011, 31, 675–685. [Google Scholar] [CrossRef]
- Hernández-Mena, D.I.; Mendoza-Garfias, B.; Ornelas-García, C.P.; Pérez-Ponce de León, G. Phylogenetic position of Magnivitellinum Kloss, 1966 and Perezitrema Baruš & Moravec, 1967 (Trematoda: Plagiorchioidea: Macroderoididae) inferred from partial 28S rDNA sequences, with the establishment of Alloglossidiidae n. fam. Syst. Parasitol. 2016, 93, 525–538. [Google Scholar] [CrossRef]
- Gomes, T.F.; Melo, F.T.; Giese, E.G.; Furtado, A.P.; Gonçalves, E.C.; Santos, J.N. A new species of Mesocoelium (Digenea: Mesocoeliidae) found in Rhinella marina (Amphibia: Bufonidae) from Brazilian Amazonia. Memórias Inst. Oswaldo Cruz 2013, 108, 186–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molnar, K.; Gibson, D.I.; Cech, G.; Papp, M.; Deak-Paulus, P.; Juhasz, L.; Toth, N.; Szekely, C. The occurrence of metacercariae of Petasiger (Digenea: Echinostomatidae) in an unusual site, within the lateral line scales of cyprinid fishes. Folia Parasitol. 2015, 62, 198–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dzikowski, R.; Levy, M.G.; Poore, M.F.; Flowers, J.R.; Paperna, I. Clinostomum complanatum and Clinostomum marginatum (Rudolphi, 1819) (Digenea:Clinostomidae) are separate species based on differences in ribosomal DNA. J. Parasitol. 2004, 90, 413–414. [Google Scholar] [CrossRef] [PubMed]
- Cech, G.; Molnár, K.; Székely, C. Molecular biological studies of adult and metacercarial stages of Petasiger exaeretus Dietz, 1909 (Digenea: Echinostomatidae). Acta Vet. Hung. 2017, 65, 198–207. [Google Scholar] [CrossRef] [Green Version]
- Cech, G.; Sándor, D.; Molnár, K.; Varga, Á.; Caffara, M.; Fioravanti, M.L.; Buchmann, K.; Székely, C. Digenean trematodes in Hungarian freshwater aquacultures. Food Waterborne Parasitol. 2021, 22, e00101. [Google Scholar] [CrossRef]
- Laidemitt, M.R.; Brant, S.V.; Mutuku, M.W.; Mkoji, G.M.; Loker, E.S. The diverse echinostomes from East Africa: With a focus on species that use Biomphalaria and Bulinus as intermediate hosts. Acta Trop. 2019, 193, 38–49. [Google Scholar] [CrossRef]
- Heneberg, P.; Rojas, A.; Bizos, J.; Kocková, L.; Malá, M.; Rojas, D. Focal Philophthalmus gralli infection possibly persists in Melanoides tuberculata over two years following the definitive hosts’ removal. Parasitol. Int. 2014, 63, 802–807. [Google Scholar] [CrossRef]
- Literák, I.; Heneberg, P.; Sitko, J.; Wetzel, E.J.; Callirgos, J.M.C.; Čapek, M.; Basto, D.V.; Papoušek, I. Eye trematode infection in small passerines in Peru caused by Philophthalmus lucipetus, an agent with a zoonotic potential spread by an invasive freshwater snail. Parasitol. Int. 2013, 62, 390–396. [Google Scholar] [CrossRef]
- Zikmundová, J.; Georgieva, S.; Faltýnková, A.; Soldánová, M.; Kostadinova, A. Species diversity of Plagiorchis Lühe, 1899 (Digenea: Plagiorchiidae) in lymnaeid snails from freshwater ecosystems in central Europe revealed by molecules and morphology. Syst. Parasitol. 2014, 88, 37–54. [Google Scholar] [CrossRef]
- Duan, Y.; Al-Jubury, A.; Kania, P.W.; Buchmann, K. Trematode diversity reflecting the community structure of Danish freshwater systems: Molecular clues. Parasites Vectors 2021, 14, 1–15. [Google Scholar] [CrossRef]
- Suleman; Ma, J.; Khan, M.S.; Tkach, V.V.; Muhammad, N.; Zhang, D.; Zhu, X.-Q. Characterization of the complete mitochondrial genome of Plagiorchis maculosus (Digenea, Plagiorchiidae), representative of a taxonomically complex digenean family. Parasitol. Int. 2019, 71, 99–105. [Google Scholar] [CrossRef]
- Grabner, D.S. Hidden diversity: Parasites of stream arthropods. Freshw. Biol. 2017, 62, 52–64. [Google Scholar] [CrossRef]
- Kudlai, O.; Pantoja, C.; O’Dwyer, K.; Jouet, D.; Skírnisson, K.; Faltýnková, A. Diversity of Plagiorchis (Trematoda: Digenea) in high latitudes: Species composition and snail host spectrum revealed by integrative taxonomy. J. Zool. Syst. Evol. Res. 2021, 59, 937–962. [Google Scholar] [CrossRef]
- Nakao, M.; Sasaki, M. Trematode diversity in freshwater snails from a stopover point for migratory waterfowls in Hokkaido, Japan: An assessment by molecular phylogenetic and population genetic analyses. Parasitol. Int. 2021, 83, 102329. [Google Scholar] [CrossRef]
- Santoro, M.; Tkach, V.V.; Mattiucci, S.; Kinsella, J.M.; Nascetti, G. Renifer aniarum (Digenea: Reniferidae), an introduced North American parasite in grass snakes Natrix natrix in Calabria, southern Italy. Dis. Aquat. Organ. 2011, 95, 233–240. [Google Scholar] [CrossRef] [Green Version]
- Iwaki, T.; Sata, N.; Hasegawa, H.; Matsuo, K.; Na, T. Ochetosoma kansense (Plagiochiida: Ochetosomatidae) from native snake species in Japan. Jpn. J. Zoo Wildl. Med. 2020, 25, 129–134. [Google Scholar] [CrossRef]
- López-Caballero, J.; Mata-López, R.; de León, G.P.-P. Molecular data reveal a new species of Rhopalias Stiles & Hassall, 1898 (Digenea, Echinostomatidae) in the Common opossum, Didelphis marsupialis L.(Mammalia, Didelphidae) in the Yucatán Peninsula, Mexico. ZooKeys 2019, 854, 145. [Google Scholar]
- Kostadinova, A.; Gibson, D.I. Isthmiophora Lühe, 1909 and Euparyphium Dietz, 1909 (Digenea: Echinostomatidae) re-defined, with comments on their nominal species. Syst. Parasitol. 2002, 52, 205–217. [Google Scholar] [CrossRef]
- Kostadinova, A.; Jones, A. Superfamily echinostomatoidea Looss, 1899. In Keys to the Trematoda; Jones, A., Bray, R.A., Gibson, D.I., Eds.; CABI Publishing: Wallingford, UK, 2005; pp. 5–8. [Google Scholar]
- Chalkowski, K.; Lepczyk, C.A.; Zohdy, S. Parasite ecology of invasive species: Conceptual framework and new hypotheses. Trends Parasitol. 2018, 34, 655–663. [Google Scholar] [CrossRef]
- Poulin, R.; Paterson, R.A.; Townsend, C.R.; Tompkins, D.M.; Kelly, D.W. Biological invasions and the dynamics of endemic diseases in freshwater ecosystems. Freshw. Biol. 2011, 56, 676–688. [Google Scholar] [CrossRef]
- Chessman, B.C. Declines of freshwater turtles associated with climatic drying in Australia’s Murray–Darling Basin. Wildl. Res. 2011, 38, 664–671. [Google Scholar] [CrossRef]
- Shamsi, S.; Stoddart, A.; Smales, L.; Wassens, S. Occurrence of Contracaecum bancrofti larvae in fish in the Murray–Darling Basin. J. Helminthol. 2019, 93, 574–579. [Google Scholar] [CrossRef]
- Shamsi, S.; Turner, A.; Wassens, S. Description and genetic characterization of a new Contracaecum larval type (Nematoda: Anisakidae) from Australia. J. Helminthol. 2017, 92, 216–222. [Google Scholar] [CrossRef]
- Smales, L.; Miller, A.; Obendorf, D. Parasites of the Water Rat, Hydromys chrysogaster, from Victoria and South-Australia. Aust. J. Zool. 1989, 37, 657–663. [Google Scholar] [CrossRef]
- Dybing, N.A.; Fleming, P.A.; Adams, P.J. Environmental conditions predict helminth prevalence in red foxes in Western Australia. Int. J. Parasitol. Parasites Wildl. 2013, 2, 165–172. [Google Scholar] [CrossRef] [Green Version]
- Johnston, T.H.; Angel, L.M. Life history of the trematode Petasiger australis n. sp. Trans. R. Soc. S. Aust. 1941, 65, 285–291. [Google Scholar]
- Angel, M. An account of Plagiorchis maculosus (Rud.), its synonymy and its life history in South Australia. Trans. R. Soc. S. Aust. 1959, 82, 265–281. [Google Scholar]
- Faltynkova, A.; Gibson, D.I.; Kostadinova, A. A revision of Petasiger Dietz, 1909 (Digenea: Echinostomatidae) and a key to its species. Syst. Parasitol. 2008, 71, 1–40. [Google Scholar] [CrossRef]
- Mawson, P.M.; Angel, M.; Edmonds, S.J. A checklist of helminths from Australian birds. Rec. S. Aust. Mus. 1986, 19, 219–325. [Google Scholar]
- Johnston, T.H. The metacercaria stage of Australian species of Clinostomum. Rec. S. Aust. Mus. 1942, 7, 187–191. [Google Scholar]
- Našincová, V.; Scholz, T.; Moravec, F.T. The life cycle of Paryphostomum radiatum (Dujardin, 1845) (Trematoda: Echinostomatidae), a parasite of cormorants. Folia Parasitol. 1993, 40, 193–201. [Google Scholar]
- Morgan, J.; Blair, D. Relative merits of nuclear ribosomal internal transcribed spacers and mitochondrial CO1 and ND1 genes for distinguishing among Echinostoma species (Trematoda). Parasitology 1998, 116, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Rochat, E.C.; Blasco-Costa, I.; Scholz, T.; Unmack, P.J. High diversity of metazoan parasites in carp gudgeons (Eleotridae: Hypseleotris spp.) from Eastern Australia. J. Helminthol. 2020, 94, e146. [Google Scholar] [CrossRef] [PubMed]
- Shamsi, S.; Day, S.; Zhu, X.; McLellan, M.; Barton, D.P.; Dang, M.; Nowak, B.F. Wild fish as reservoirs of parasites on Australian Murray cod farms. Aquaculture 2021, 539, 736584. [Google Scholar] [CrossRef]
- Brönmark, C. Freshwater snail diversity: Effects of pond area, habitat heterogeneity and isolation. Oecologia 1985, 67, 127–131. [Google Scholar] [CrossRef]

 , snails;
, snails;  , turtles;
, turtles;  , snakes;
, snakes;  , frogs and toads;
, frogs and toads;  , leeches;
, leeches;  , fishes;
, fishes;  , Daphnia;
, Daphnia;  , freshwater prawns;
, freshwater prawns;  , insects;
, insects;  , bats;
, bats;  , mammals other than bats;
, mammals other than bats;  , swallow). The hosts are those listed in Table 1 and include hosts from which parasites/sequences were obtained. Some of these hosts are intermediate/paratenic and some are definitive hosts.
, snails; , turtles; , snakes; , frogs and toads; , leeches; , fishes; , Daphnia; , freshwater prawns; , insects; , bats; , mammals other than bats; , swallow). The hosts are those listed in Table 1 and include hosts from which parasites/sequences were obtained. Some of these hosts are intermediate/paratenic and some are definitive hosts.
, swallow). The hosts are those listed in Table 1 and include hosts from which parasites/sequences were obtained. Some of these hosts are intermediate/paratenic and some are definitive hosts.
, snails; , turtles; , snakes; , frogs and toads; , leeches; , fishes; , Daphnia; , freshwater prawns; , insects; , bats; , mammals other than bats; , swallow). The hosts are those listed in Table 1 and include hosts from which parasites/sequences were obtained. Some of these hosts are intermediate/paratenic and some are definitive hosts.
 , snails; , fishes; , mammals other than bats;
, snails; , fishes; , mammals other than bats;  , fish-eating birds). The hosts are those listed in Table 1 and include hosts from which parasites/sequences were obtained. Some of these hosts are intermediate/paratenic and some are definitive hosts.
, snails; , fishes; , mammals other than bats; , fish-eating birds). The hosts are those listed in Table 1 and include hosts from which parasites/sequences were obtained. Some of these hosts are intermediate/paratenic and some are definitive hosts.
, fish-eating birds). The hosts are those listed in Table 1 and include hosts from which parasites/sequences were obtained. Some of these hosts are intermediate/paratenic and some are definitive hosts.
, snails; , fishes; , mammals other than bats; , fish-eating birds). The hosts are those listed in Table 1 and include hosts from which parasites/sequences were obtained. Some of these hosts are intermediate/paratenic and some are definitive hosts.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trematode Species | Trematode Family | GenBank Accession No | Host | Parasite Development Stage | Locality | Reference |
|---|---|---|---|---|---|---|
| Alloglossidium anomaphagis | Alloglossidiidae | MH041376 | Daphnia obtusa | Adult | USA | [19] |
| Alloglossidium floridense | Alloglossidiidae | MH041390 | Noturus gyrinus | Adult | USA | [19] |
| Alloglossidium fonti | Alloglossidiidae | MH041395 | Ameiurus melas | Adult | USA | [19] |
| Alloglossidium greeri | Alloglossidiidae | MH041387 | Cambarellus shufeldtii | Adult | USA | [19] |
| Alloglossidium hamrumi | Alloglossidiidae | MH041415 | Macrobdella decora | Adult | USA | [19] |
| Alloglossidium hirudicola | Alloglossidiidae | MH041418 | Macrobdella decora | Adult | USA | [19] |
| Alloglossidium kenti | Alloglossidiidae | MH041405 | Ictalurus punctatus | Adult | USA | [19] |
| Alloglossidium macrobdellensis | Alloglossidiidae | MH041413 | Macrobdella decora | Adult | USA | [19] |
| Alloglossidium progeneticum | Alloglossidiidae | MH041382 | Procambarus spiculifer | Adult | USA | [19] |
| Alloglossidium renale | Alloglossidiidae | MH041385 | Palaemonetes kadiakensis | Adult | USA | [19] |
| Alloglossidium schmidti | Alloglossidiidae | MH041419 | Haemopis grandis | Adult | Canada | [19] |
| Alloglossidium turnbulli | Alloglossidiidae | MH041423 | Haemopis grandis | Adult | USA | [19] |
| Aptorchis aequalis | Plagiorchiidae | EU334369 | Emydura krefftii | Adult | Australia | [20] |
| Aptorchis glandularis | Plagiorchiidae | EU334368 | Emydura australis | Adult | Australia | [20] |
| Aptorchis kuchlingi | Plagiorchiidae | HQ680841, HQ680845 | Chelodina oblonga | Adult | Australia | [21] |
| Aptorchis megacetabulus | Plagiorchiidae | EF014730 | Chelodina rugosa | Adult | Australia | [22] |
| Aptorchis megapharynx | Plagiorchiidae | EF014727 | Chelodina longicollis | Adult | Australia | [22] |
| Aptorchis pearsoni | Plagiorchiidae | EF014728 | Chelodina expansa | Adult | Australia | [22] |
| Auridostomum chelydrae | Auridistomidae | AY222159 | Chelydra serpentina | Adult | USA | [23] |
| Brachycladium goliath (OUTGROUP) | Brachycladiidae | KR703279 | Balaenoptera acutorostrata | Adult | UK | [24] |
| Brachycoelium salamandrae | Brachycoeliidae | AY222160 | Salamandra salamandra | Adult | Ukraine | [23] |
| Cathaemasia hians | Echinostomatidae | KT956947 | Planorbis planorbis | Cercaria | Czech Republic | [25] |
| Cephalogonimus retusus | Cephalogonimidae | AJ287489 | Rana ridibunda | Adult | - | [26] |
| Choanocotylehobbsi | Choanocotylidae | EU196356 | Chelodina oblonga | Adult | Australia | [27] |
| Choanocotyle hobbsi | Choanocotylidae | MW682817-MW682819 | Isidorella hainesii | Cercaria | Australia | [13] |
| Choanocotyle hobbsi | Choanocotylidae | MW684083-MW684089 | Isidorella hainesii | Cercaria | Australia | [13] |
| Choanocotyle hobbsi | Choanocotylidae | MW686389, MW686392-MW686393 | Isidorella hainesii | Cercaria | Australia | [13] |
| Choanocotyle hobbsi | Choanocotylidae | OM305034-OM305039 | Haitia acuta | Cercaria | Australia | This study |
| Choanocotyle hobbsi | Choanocotylidae | OM305043-OM305048 | Haitia acuta | Cercaria | Australia | This study |
| Choanocotyle hobbsi | Choanocotylidae | OM305095-OM305100 | Haitia acuta | Cercaria | Australia | This study |
| Choanocotyle nematoides | Choanocotylidae | AY116862-AY116864, AY116867 | Chelodina oblonga | Adult | Australia | [28] |
| Choanocotyle nematoides | Choanocotylidae | EU196357-EU196358 | Emydura krefftii | Adult | Australia | [27] |
| Choanocotyle nematoides | Choanocotylidae | EU196359-EU196360 | Emydura macquarii | Adult | Australia | [27] |
| Choanocotyle platti | Choanocotylidae | EU196355 | Chelodina rugosa | Adult | Australia | [27] |
| Choledocystus hepatica | Plagiorchiidae | AY875679 | Rhinella marina | Adult | Mexico | [29] |
| Dasymetra nicolli | Reniferidae | AF433672 | Nerodia rhombifer | Adult | USA | [30] |
| Drepanocephalus auritus | Echinostomatidae | KP053259 | Biomphalaria stramina | Cercaria | Brazil | [31] |
| Drepanocephalus auritus | Echinostomatidae | KP683117 | Phalacrocorax auritus | Adult | USA | [32] |
| Drepanocephalus auritus | Echinostomatidae | KY677976, KY677977 | Biomphalaria havanensis | Cercaria | USA | [33] |
| Drepanocephalus mexicanus | Echinostomatidae | KY636276 | Nannopterum brasilianus | Adult | - | [34] |
| Drepanocephalus mexicanus | Echinostomatidae | MF351542 | Nannopterum brasilianus | Adult | - | [34] |
| Drepanocephalus sp. | Echinostomatidae | KP053261 | Biomphalaria stramina | Cercaria | Brazil | [31] |
| Drepanocephalus spathans | Echinostomatidae | AY245762 | Not stated | Not stated | Not stated | Unpublished |
| Drepanocephalus spathans | Echinostomatidae | JN993269 | Phalacrocorax auritus | Adult | USA | [35] |
| Drepanocephalus spathans | Echinostomatidae | KY636260 | Nannopterum brasilianus | Adult | - | [34] |
| Echinostoma hortense a | Echinostomatidae | KX832896 | Misgurnus anguillicaudatus | Metacercariae | China | [36] |
| Euparyphium capitaneum | Echinostomatidae | KP009616 | Anhinga anhinga | Adult | USA | [37] |
| Euparyphium melis b | Echinostomatidae | AF151941 | Nyctereutes procyonoides | Adult | Ukraine | [38] |
| Euparyphium melis b | Echinostomatidae | AY222131 | Nyctereutes procyonoides | Adult | Ukraine | [23] |
| Glypthelmins africana | Glypthelminthidae | OL413039 | Hyperolius viridiflavus | Adult | Rwanda | [39] |
| Glypthelmins quieta | Glypthelminthidae | AJ287517 | Rana catesbeiana | Adult | - | [26] |
| Haematoleochus longiplexus | Haematoleochidae | AJ287520 | Rana catesbeiana | Adult | - | [26] |
| Haematoleochus sp. | Haematoleochidae | MH285261 | Odorrana grahami | Adult | China | Unpublished |
| Haplometra cylindracea | Plagiorchiidae | AF151933 | Rana arvalis | Adult | Ukraine | [38] |
| Haplometroides intercaecalis | Plagiorchiidae | MH206169 | Phalotris matogrossensis | Adult | Brazil | [40] |
| Isthmiophora hortensis | Echinostomatidae | AB189982 | Misgurnus anguillicaudatus | Adult | Japan | [41] |
| Isthmiophora melis | Echinostomatidae | KT359583-KT359584 | Apodemus agrariu | Adult | Poland | [42] |
| Lechriorchis tygarti | Reniferidae | JF820599-JF62600 | Lithobates sylvaticus | Metacercaria | USA | [43] |
| Macroderoides typicus | Macroderoididae | AY222158 | Lepisosteus platostomus | Adult | USA | [23] |
| Magnivitellinum simplex | Alloglossidiidae | KU535678, KU535681-KU535683 | Astyanax mexicanus | Adult | Mexico | [44] |
| Mesocoelium lanfrediae | Brachycoeliidae | JQ886404 | Rhinella marina | Adult | Brazil | [45] |
| Neoglyphe locellus | Omphalometridae | AF300330 | Sorex araneus | Adult | Ukraine | [30] |
| Neoglyphe sobolesi | Omphalometridae | AF300329 | Sorex araneus | Adult | Ukraine | [30] |
| Omphalometra flexuosa | Omphalometridae | AF300333 | Planorbis planorbis | Cercaria | Poland | [30] |
| Opisthioglyphe ranae | Telorchiidae | AF151929 | Rana arvalis | Adult | Ukraine | [38] |
| Opisthioglyphe ranae | Telorchiidae | AY222157 | Rana arvalis | Adult | Ukraine | [23] |
| Opisthioglyphe ranae | Telorchiidae | MK585340-MK585341 | Pelophylax ridibundus | Metacercaria | Russia | Unpublished |
| Paryphostomum radiatum c | Echinostomatidae | KM972998, KM973000 | Phalacrocorax carbo | Adult | Hungary | [46] |
| Paryphystomum radiatium c | Echinostomatidae | AY245708 | Phalacrocorax carbo | Adult | Israel | [47] |
| Pegosomum asperum | Echinostomatidae | KY945919 | Ardea alba | Adult | Germany | Unpublished |
| Pegosomum saginatum | Echinostomatidae | KY945918 | Ardea alba | Adult | Germany | Unpublished |
| Petasiger exaeretus | Echinostomatidae | KT956923 | Phalacrocorax carbo | Adult | Ukraine | [25] |
| Petasiger exaeretus | Echinostomatidae | KY283998 | Phalacrocorax carbo | Adult | Hungary | [48] |
| Petasiger phalacrocoracis | Echinostomatidae | AY245709 | Phalacrocorax carbo | Adult | Israel | [47] |
| Petasiger phalacrocoracis | Echinostomatidae | KJ720683 | Rutilus rutilus | Metacercaria | Hungary | [46] |
| Petasiger phalacrocoracis | Echinostomatidae | KY283999 | Rutilus rutilus | Metacercaria | Hungary | [49] |
| Petasiger radiatus | Echinostomatidae | KJ956927 | Phalacrocorax carbo | Adult | Ukraine | [25] |
| Petasiger radiatus | Echinostomatidae | KY284010 | Phalacrocorax carbo | Adult | Hungary | [49] |
| Petasiger sp. | Echinostomatidae | KY284003 | Rutilus rutilus | Metacercaria | Hungary | [49] |
| Petasiger sp. | Echinostomatidae | OM305031-OM305033 | Isidorella hainesii | Cercaria | Australia | This study |
| Petasiger sp. | Echinostomatidae | OM305052-OM305054 | Isidorella hainesii | Cercaria | Australia | This study |
| Petasiger sp. | Echinostomatidae | OM305104-OM305107 | Isidorella hainesii | Cercaria | Australia | This study |
| Petasiger sp. 1 | Echinostomatidae | MK482443 | Radix natalensis | Cercaria | Kenya | [50] |
| Petasiger sp. 2 | Echinostomatidae | MK482449 | Bulinus globosus | Cercaria | Kenya | [50] |
| Petasiger sp. 3 | Echinostomatidae | MK482446 | Radix natalensis | Cercaria | Kenya | [50] |
| Petasiger sp. 4 | Echinostomatidae | MK482430 | Biomphalaria pfeifferi | Cercaria | Kenya | [50] |
| Petasiger sp. 5 | Echinostomatidae | MK482414 | Bulinus sp. | Cercaria | Kenya | [50] |
| Petasiger sp. 6 | Echinostomatidae | MK482447 | Bulinus sp. | Cercaria | Kenya | [50] |
| Philophthalmus gralli (OUTGROUP) | Philophthalmidaae | JQ627832 | Tachuris rubrigastra | Adult | Peru | [51] |
| Philophthalmus gralli (OUTGROUP) | Philophthalmidaae | JX121229 | Tachuris rubrigastra | Adult | Peru | [52] |
| Plagiorchiselegans | Plagiorchiidae | KF556678 | Lymnaea stagnalis | Cercaria | USA | [35] |
| Plagiorchiselegans | Plagiorchiidae | KJ533393 | Lymnaea stagnalis | Cercaria | Czech Republic | [53] |
| Plagiorchiselegans | Plagiorchiidae | MW001064, MW001068 | Lymnaea stagnalis | Cercaria | Denmark | [54] |
| Plagiorchis koreanus | Plagiorchiidae | AF151930 | Nyctalus noctula | Adult | Ukraine | [38] |
| Plagiorchis maculosus | Plagiorchiidae | KJ533395 | Lymnaea stagnalis | Cercaria | Czech Republic | [53] |
| Plagiorchis maculosus | Plagiorchiidae | MK641807 | Hirundo rustica | Adult | Pakistan | [55] |
| Plagiorchis maculosus | Plagiorchiidae | MW001083 | Lymnaea stagnalis | Cercaria | Denmark | [54] |
| Plagiorchis neomidis | Plagiorchiidae | KJ533397 | Lymnaea stagnalis | Cercaria | Czech Republic | [53] |
| Plagiorchis sp. | Plagiorchiidae | KJ533398 | Lymnaea stagnalis | Cercaria | Czech Republic | [53] |
| Plagiorchis sp. | Plagiorchiidae | MW001088 | Lymnaea stagnalis | Cercaria | Denmark | [54] |
| Plagiorchis sp. | Plagiorchiidae | MW001090 | Stagnicola palustris | Cercaria | Denmark | [54] |
| Plagiorchis sp. | Plagiorchiidae | MW001091 | Ampullaceana balthica | Cercaria | Denmark | [54] |
| Plagiorchis sp. | Plagiorchiidae | MW001113 | Lymnaea stagnalis | Cercaria | Denmark | [54] |
| Plagiorchis sp. | Plagiorchiidae | OM305040-OM305042 | Bullastra lessoni | Cercaria and Sporocysts | Australia | This study |
| Plagiorchis sp. | Plagiorchiidae | OM305049-OM305050 | Bullastra lessoni | Cercaria and Sporocysts | Australia | This study |
| Plagiorchis sp. | Plagiorchiidae | OM305101-OM305103 | Bullastra lessoni | Cercaria and Sporocysts | Australia | This study |
| Plagiorchis sp. 1 | Plagiorchiidae | KX160477 | Hydropsyche sp. | Metacercaria | Germany | [56] |
| Plagiorchis sp. 1 | Plagiorchiidae | MW528604 | Ampullaceana balthica | Cercaria | Iceland | [57] |
| Plagiorchis sp. 2 | Plagiorchiidae | MW001092 | Stagnicola palustris | Cercaria | Denmark | [54] |
| Plagiorchis sp. 2 | Plagiorchiidae | MW528605 | Radix balthica | Cercaria | Iceland | [57] |
| Plagiorchis sp. 3 | Plagiorchiidae | KX160474 | Lepidostematus sp. | Metacercaria | Germany | [56] |
| Plagiorchis sp. 3 | Plagiorchiidae | MW528606 | Radix balthica | Cercaria | Ireland | [57] |
| Plagiorchis sp. 5 | Plagiorchiidae | MW528611 | Radix balthica | Cercaria | Ireland | [57] |
| Plagiorchis sp. 7 | Plagiorchiidae | MW528616 | Radix balthica | Cercaria | Ireland | [57] |
| Plagiorchis sp. 8 | Plagiorchiidae | MW528619 | Radix balthica | Cercaria | Ireland | [57] |
| Plagiorchis sp. 9 | Plagiorchiidae | MW528621 | Stagnicola fuscus | Cercaria | Ireland | [57] |
| Plagiorchis sp. A | Plagiorchiidae | LC599522 | Radix auricularia | Daughter Sporocyst | Japan | [58] |
| Plagiorchis sp. B | Plagiorchiidae | LC599524 | Radix auricularia | Daughter Sporocyst | Japan | [58] |
| Plagiorchis sp. C | Plagiorchiidae | LC599525 | Radix auricularia | Daughter Sporocyst | Japan | [58] |
| Plagiorchis sp. Lineage 1 | Plagiorchiidae | MW528622 | Stagnicola elodes | Cercaria | USA | [57] |
| Plagiorchis sp. Lineage 4 | Plagiorchiidae | MW528623 | Stagnicola elodes | Cercaria | USA | [57] |
| Plagiorchis sp. Lineage 6 | Plagiorchiidae | MW528624 | Stagnicola elodes | Cercaria | USA | [57] |
| Plagiorchis sp. Lineage 9 | Plagiorchiidae | MW528626 | Stagnicola elodes | Cercaria | USA | [57] |
| Plagiorchis vespertilionis | Plagiorchiidae | AF151931 | Myotis daubentoni | Adult | Ukraine | [38] |
| Renifer aniarum | Reniferidae | HQ665459 | Nerodia rhombifer | Adult | USA | [59] |
| Renifer kansensis | Reniferidae | LC557508, LC557512 | Elaphe quadrivirgata | Adult | Japan | [60] |
| Rhopalias coronatus | Echinostomatidae | MK982797, MK982801, MK982813 | Didelphismarsupialis virginiana | Adult | Mexico | [61] |
| Rhopalias oochi | Echinostomatidae | MK982803 | Didelphismarsupialis marsupialis | Adult | Mexico | [61] |
| Ribeiroia ondatrae | Echinostomatidae | MK321661 | Biomphalaria sudanica | Cercaria | Kenya | [50] |
| Ribeiroia sp. 1 | Echinostomatidae | MK482424 | Biomphalaria sudanica | Cercaria | Kenya | [50] |
| Ribeiroia sp. 2 | Echinostomatidae | MK482418 | Biomphalaria sudanica | Cercaria | Kenya | [50] |
| Ribeiroia sp. 3 | Echinostomatidae | MK482461 | Biomphalaria sudanica | Cercaria | Kenya | [50] |
| Rubenstrema exasperatum | Omphalometridae | AF300331 | Sorex araneus | Adult | Ukraine | [30] |
| Rubenstrema exasperatum | Omphalometridae | AJ287572 | Crocidura leucodon | - | - | [26] |
| Rubenstrema exasperatum | Omphalometridae | MK585231 | Planorbarius corneus | Metacercaria | Russia | Unpublished |
| Sigmapera cincta | Plagiorchiidae | EF411200 | Emydura kreffti | Not stated | Australia | Unpublished |
| Skrjabinoeces similis | Plagiorchiidae | AJ287575 | Rana ridibunda | - | - | [26] |
| Skrjabinoeces similis | Plagiorchiidae | AY222279 | Pelophylax ridibundus | Adult | Bulgaria | [23] |
| Telorchis assula | Telorchiidae | AF151915 | Natrix natrix | Adult | Ukraine | [38] |
| Telorchis assula | Telorchiidae | AY222156 | Natrix natrix | Adult | Ukraine | [23] |
| Telorchis bonnerensis | Telorchiidae | JF820591 | Ambystoma tigrinum | Adult | USA | [43] |
| Telorchis bonnerensis | Telorchiidae | JF820593 | Lithobates sylvaticus | Metacercaria | USA | [43] |
| Telorchis sp. | Telorchiidae | OL960085 | Planorbella trivolvis | Not stated | USA |
| Snail Species | No. Examined (No. Infected) | Locality | Provisional Parasite Identification (Groups/Morphotype) | Parasite Species Found | Infected Snail Code | No. of Sporocysts | No. of Redia | No. of Cercaria | Genetic ID (Y/N) |
|---|---|---|---|---|---|---|---|---|---|
| Bullastra lessoni | 11 (1) | 1 | A | Plagiorchis sp. | 11 | >100 | 0 | >100 | Y |
| Haitia acuta | 88 (4) | 2 | B | Choanocotyle hobbsi | 47, 123, 124, 126 | 0, 0, 0, 0 | 0, 0, 0, 0 | 5, 1, 1, 2 | N |
| 11 (0) | 4 | - | - | - | - | - | - | - | |
| 8 (1) | 3 | B | Choanocotyle hobbsi | 34 | 10–50 | 0 | 50–100 | Y | |
| Isidorella hainesii | 150 (2) | 2 | C | Petasiger sp. | 94, 85 | 0 | >100 | 50–100 | Y |
| 4 (0) | 4 | - | - | - | - | - | - | - | |
| 3 (0) | 3 | - | - | - | - | - | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barton, D.P.; Zhu, X.; Nuhoglu, A.; Pearce, L.; McLellan, M.; Shamsi, S. Parasites of Selected Freshwater Snails in the Eastern Murray Darling Basin, Australia. Int. J. Environ. Res. Public Health 2022, 19, 7236. https://doi.org/10.3390/ijerph19127236
Barton DP, Zhu X, Nuhoglu A, Pearce L, McLellan M, Shamsi S. Parasites of Selected Freshwater Snails in the Eastern Murray Darling Basin, Australia. International Journal of Environmental Research and Public Health. 2022; 19(12):7236. https://doi.org/10.3390/ijerph19127236
Chicago/Turabian StyleBarton, Diane P., Xiaocheng Zhu, Alara Nuhoglu, Luke Pearce, Matthew McLellan, and Shokoofeh Shamsi. 2022. "Parasites of Selected Freshwater Snails in the Eastern Murray Darling Basin, Australia" International Journal of Environmental Research and Public Health 19, no. 12: 7236. https://doi.org/10.3390/ijerph19127236