Keratinocyte Growth Factor-1 Protects Radioiodine-Induced Salivary Gland Dysfunction in Mice


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
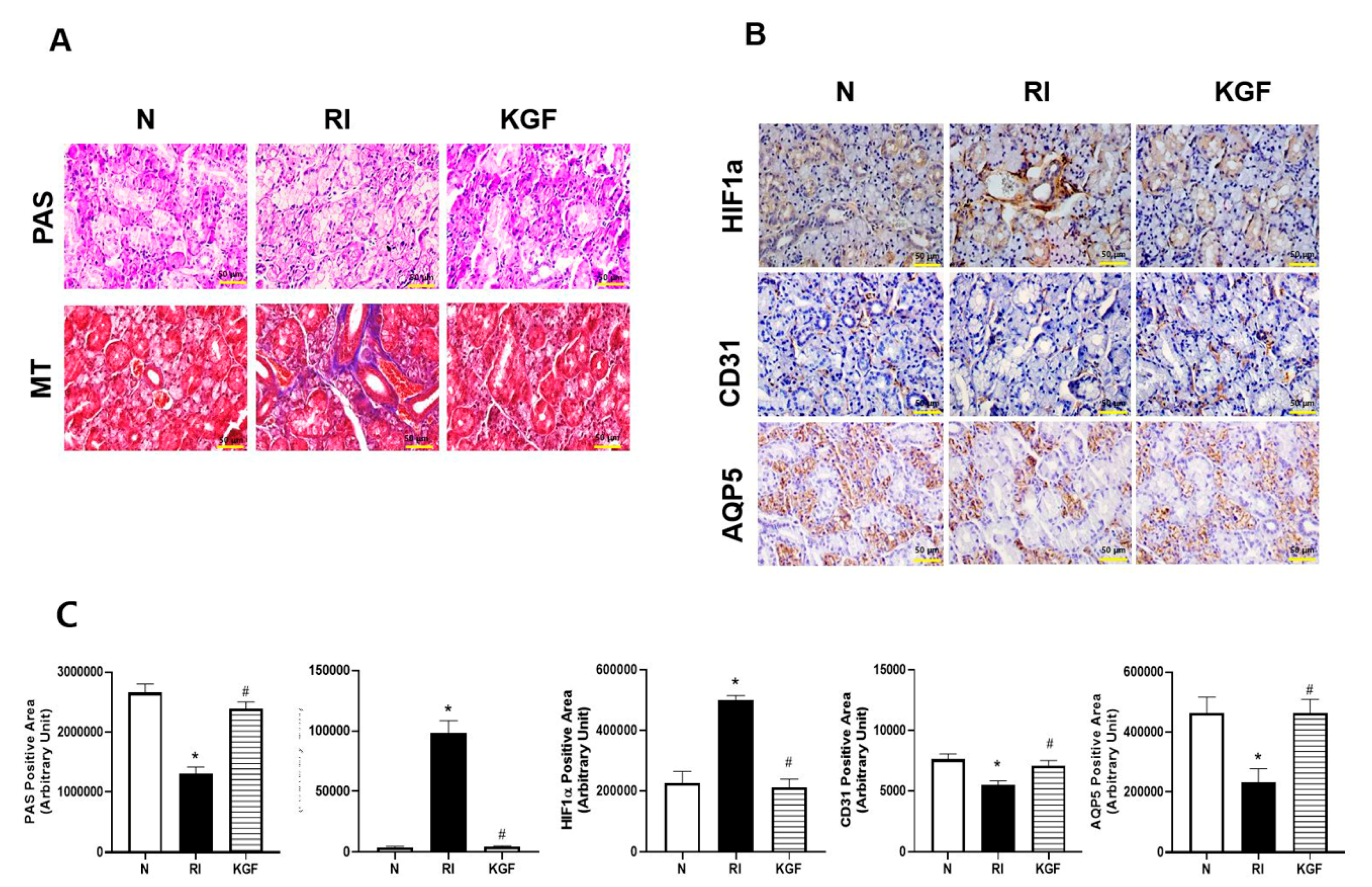
2. Materials and Methods
2.1. Animal Studies
2.2. Measurements of Body and SG Weights, Salivary Flow Rates, and Lag Times
2.3. SPECT Image Analysis
2.4. Histopathology
2.5. Immunohistochemistry
2.6. Detection of Amylase Activity and Expression in Saliva
2.7. Detection of Apoptosis and Related Proteins
2.8. Western Blotting
2.9. Statistical Analysis
3. Results
3.1. Preventive Effect of KGF-1 on RI Induced Body and SG Weight Losses
3.2. Protective Effect of KGF-1 on RI-Induced SG Dysfunction
3.3. Enhanced SG Secretory Function by KGF-1
3.4. Salivary Cell Protective Effect of KGF-1
3.5. Anti-Apoptotic Effect of KGF-1 in SGs
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Choi, J.-S.; An, H.-Y.; Park, I.S.; Kim, S.-K.; Kim, Y.-M.; Lim, J.-Y. Radioprotective Effect of Epigallocatechin-3-Gallate on Salivary Gland Dysfunction after Radioiodine Ablation in a Murine Model. Clin. Exp. Otorhinolaryngol. 2016, 9, 244–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.W.; Kim, J.-M.; Kim, S.-K.; Kim, Y.M.; Choi, J. Protective Effect of Ginseng on Salivary Dysfunction Following Radioiodine Therapy in a Mouse Model. Thyroid 2018, 28, 1034–1041. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; An, H.-Y.; Shin, H.-S.; Kim, Y.M.; Lim, J.-Y. Enhanced tissue remodelling efficacy of adipose-derived mesenchymal stem cells using injectable matrices in radiation-damaged salivary gland model. J. Tissue Eng. Regen. Med. 2017, 12, e695–e706. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.-S.; Shin, H.-S.; An, H.-Y.; Kim, Y.-M.; Lim, J.-Y. Radioprotective effects of Keratinocyte Growth Factor-1 against irradiation-induced salivary gland hypofunction. Oncotarget 2017, 8, 13496–13508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, H.-Y.; Shin, H.-S.; Choi, J.-S.; Kim, H.J.; Lim, J.-Y.; Kim, Y.-M. Adipose Mesenchymal Stem Cell Secretome Modulated in Hypoxia for Remodeling of Radiation-Induced Salivary Gland Damage. PLoS ONE 2015, 10, e0141862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rotolo, S.; Ceccarelli, S.; Romano, F.; Frati, L.; Marchese, C.; Angeloni, A. Silencing of Keratinocyte Growth Factor Receptor Restores 5-Fluorouracil and Tamoxifen Efficacy on Responsive Cancer Cells. PLoS ONE 2008, 3, e2528. [Google Scholar] [CrossRef] [PubMed]
- Takeoka, M.; Ward, W.F.; Pollack, H.; Kamp, D.W.; Panos, R.J. KGF facilitates repair of radiation-induced DNA damage in alveolar epithelial cells. Am. J. Physiol. 1997, 272, L1174–L1180. [Google Scholar] [CrossRef]
- Zeeh, J.; Procaccino, F.; Hoffmann, P.; Aukerman, S.; McRoberts, J.; Soltani, S.; Pierce, G.; Lakshmanan, J.; Lacey, D.; Eysselein, V. Keratinocyte growth factor ameliorates mucosal injury in an experimental model of colitis in rats. Gastroenterology 1996, 110, 1077–1083. [Google Scholar] [CrossRef]
- Brizel, D.M.; Murphy, B.A.; Rosenthal, D.; Pandya, K.J.; Glück, S.; Brizel, H.E.; Meredith, R.F.; Berger, D.; Chen, M.-G.; Mendenhall, W. Phase II Study of Palifermin and Concurrent Chemoradiation in Head and Neck Squamous Cell Carcinoma. J. Clin. Oncol. 2008, 26, 2489–2496. [Google Scholar] [CrossRef]
- Brake, R.; Starnes, C.; Lu, J.; Chen, D.; Yang, S.; Radinsky, R.; Borges, L. Effects of palifermin on antitumor activity of chemotherapeutic and biological agents in human head and neck and colorectal carcinoma xenograft models. Mol. Cancer Res. 2008, 6, 1337–1346. [Google Scholar] [CrossRef] [Green Version]
- Zheng, C.; Cotrim, A.P.; Rowzee, A.M.; Swaim, W.; Sowers, A.; Mitchell, J.B.; Baum, B.J. Prevention of radiation-induced salivary hypofunction following hKGF gene delivery to murine submandibular glands. Clin. Cancer Res. 2011, 17, 2842–2851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maier, H.; Bihl, H. Effect of radioactive iodine therapy on parotid gland function. Acta Otolaryngol. 1987, 103, 318–324. [Google Scholar] [CrossRef] [PubMed]
- Dohan, O.; Carrasco, N. Advances in Na(+)/I(−) symporter (NIS) research in the thyroid and beyond. Mol. Cell Endocrinol. 2003, 213, 59–70. [Google Scholar] [CrossRef]
- La Perle, K.M.; Kim, D.C.; Hall, N.C.; Bobbey, A.; Shen, D.H.; Nagy, R.S.; Wakely, P.E.; Lehman, A.; Jarjoura, D.; Jhiang, S.M. Modulation of Sodium/Iodide Symporter Expression in the Salivary Gland. Thyroid 2013, 23, 1029–1036. [Google Scholar] [CrossRef] [PubMed]
- Klubo-Gwiezdzinska, J.; Costello, J.; Jensen, K.; Patel, A.; Tkavc, R.; Van Nostrand, D.; Burman, K.D.; Wartofsky, L.; Vasko, V.V. Amifostine does not protect thyroid cancer cells in DNA damaging in vitro models. Endocr. Connect. 2017, 6, 469–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marmary, Y.; Adar, R.; Gaska, S.; Wygoda, A.; Maly, A.; Cohen, J.; Eliashar, R.; Mizrachi, L.; Orfaig-Geva, C.; Baum, B.J.; et al. Radiation-Induced Loss of Salivary Gland Function Is Driven by Cellular Senescence and Prevented by IL6 Modulation. Cancer Res. 2016, 76, 1170–1180. [Google Scholar] [CrossRef] [Green Version]
- Tai, G.; Zhang, H.; Du, J.; Chen, G.; Huang, J.; Yu, J.; Cai, J.; Liu, F. TIGAR overexpression diminishes radiosensitivity of parotid gland fibroblast cells and inhibits IR-induced cell autophagy. Int. J. Clin. Exp. Pathol. 2015, 8, 4823–4829. [Google Scholar]
- Zheng, X.; Wang, X.; Ma, Z.; Sunkari, V.G.; Botusan, I.; Takeda, T.; Bjorklund, A.; Inoue, M.; Catrina, S.B.; Brismar, K.; et al. Acute hypoxia induces apoptosis of pancreatic beta-cell by activation of the unfolded protein response and upregulation of CHOP. Cell Death Dis. 2012, 3, e322. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Ma, C.; Li, R.; Xue, J.; Liu, L.; Liu, P. Hypoxia Induces Apoptosis through HIF-1α Signaling Pathway in Human Uterosacral Ligaments of Pelvic Organ Prolapse. BioMed Res. Int. 2017, 2017, 8316094. [Google Scholar] [CrossRef] [Green Version]
- Cai, Y.; Wang, W.; Qiu, Y.; Yu, M.; Yin, J.; Yang, H.; Mei, J. KGF inhibits hypoxia-induced intestinal epithelial cell apoptosis by upregulating AKT/ERK pathway-dependent E-cadherin expression. Biomed. Pharmacother. 2018, 105, 1318–1324. [Google Scholar] [CrossRef]
- Kojima, T.; Kanemaru, S.-I.; Hirano, S.; Tateya, I.; Suehiro, A.; Kitani, Y.; Kishimoto, Y.; Ohno, S.; Nakamura, T.; Ito, J. The protective efficacy of basic fibroblast growth factor in radiation-induced salivary gland dysfunction in mice. Laryngoscope 2011, 121, 1870–1875. [Google Scholar] [CrossRef] [PubMed]
- Fang, D.; Hu, S.; Liu, Y.; Quan, V.-H.; Seuntjens, J.; Tran, S.D. Identification of the active components in Bone Marrow Soup: A mitigator against irradiation-injury to salivary glands. Sci. Rep. 2015, 5, 16017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, V.T.; Dawson, P.; Zhang, Q.; Harris, Z.; Limesand, K.H. Administration of growth factors promotes salisphere formation from irradiated parotid salivary glands. PLoS ONE 2018, 13, e0193942. [Google Scholar] [CrossRef] [PubMed]
- Kojima, A.; Horiguchi, N.; Kakizaki, S.; Takayama, H.; Mori, M. Hepatocyte growth factor promotes lung carcinogenesis in transgenic mice treated with diethylnitrosamine. Anticancer. Res. 2013, 33, 895–901. [Google Scholar]
- Yura, Y.; Yoshioka, Y.; Yamamoto, S.; Kusaka, J.; Bando, T.; Yoshida, H.; Sato, M. Enhancing effects of fibroblast growth factor on the proliferation of salivary gland carcinoma cells and salivary gland carcinogenesis. J. Oral Pathol. Med. 2001, 30, 159–167. [Google Scholar] [CrossRef]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, J.M.; Choi, M.E.; Kim, S.-K.; Kim, J.W.; Kim, Y.-M.; Choi, J.-S. Keratinocyte Growth Factor-1 Protects Radioiodine-Induced Salivary Gland Dysfunction in Mice. Int. J. Environ. Res. Public Health 2020, 17, 6322. https://doi.org/10.3390/ijerph17176322
Kim JM, Choi ME, Kim S-K, Kim JW, Kim Y-M, Choi J-S. Keratinocyte Growth Factor-1 Protects Radioiodine-Induced Salivary Gland Dysfunction in Mice. International Journal of Environmental Research and Public Health. 2020; 17(17):6322. https://doi.org/10.3390/ijerph17176322
Chicago/Turabian StyleKim, Jeong Mi, Mi Eun Choi, Seok-Ki Kim, Ji Won Kim, Young-Mo Kim, and Jeong-Seok Choi. 2020. "Keratinocyte Growth Factor-1 Protects Radioiodine-Induced Salivary Gland Dysfunction in Mice" International Journal of Environmental Research and Public Health 17, no. 17: 6322. https://doi.org/10.3390/ijerph17176322