Investigating the Role of Gold Nanoparticle Shape and Size in Their Toxicities to Fungi




Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Synthesis of Colloidal GNPs
2.3. Characterisation of GNPs
2.4. Toxicity Test of GNPs on Fungi
2.5. Statistical Analysis
3. Results
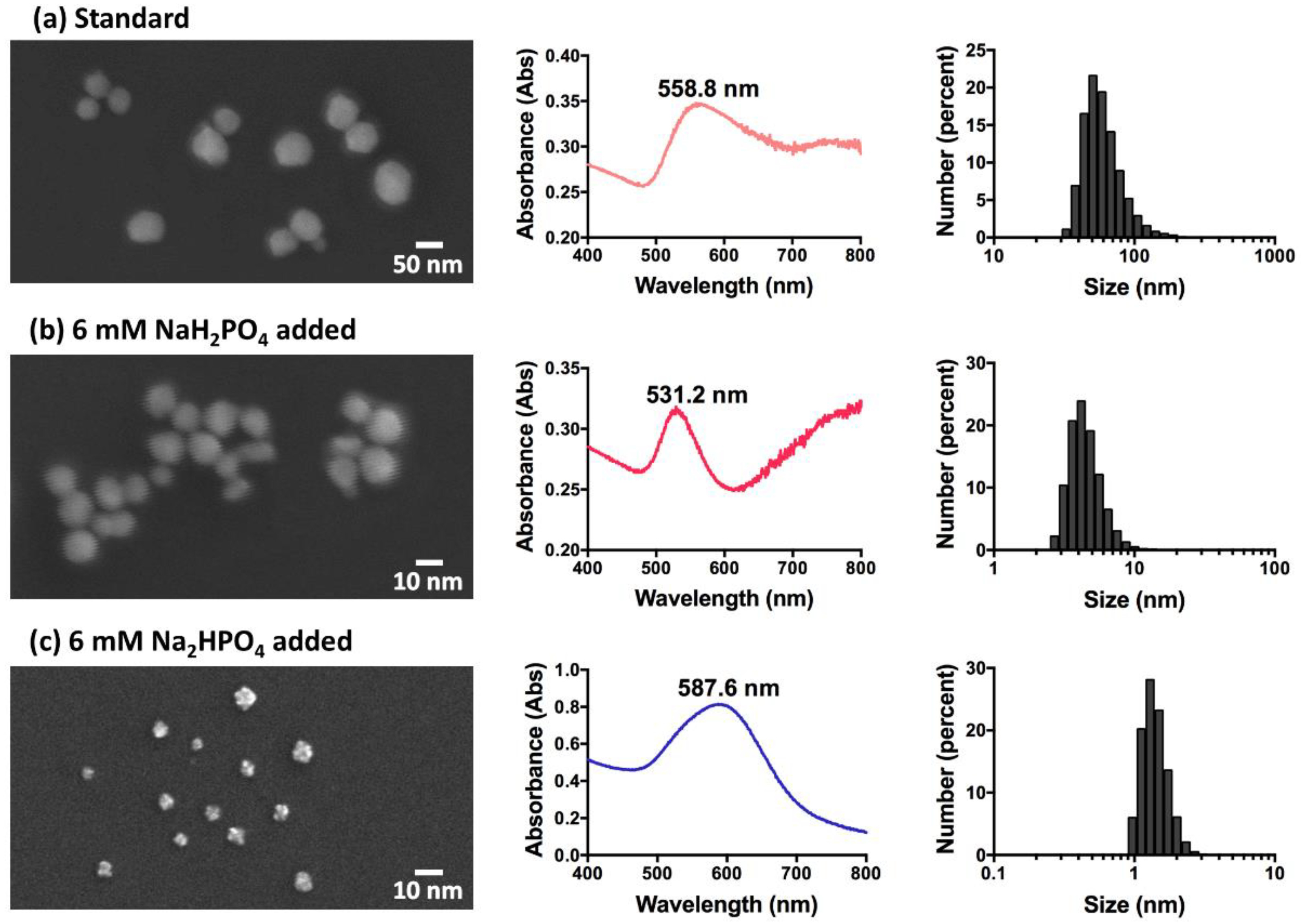
3.1. Characterisation of GNPs
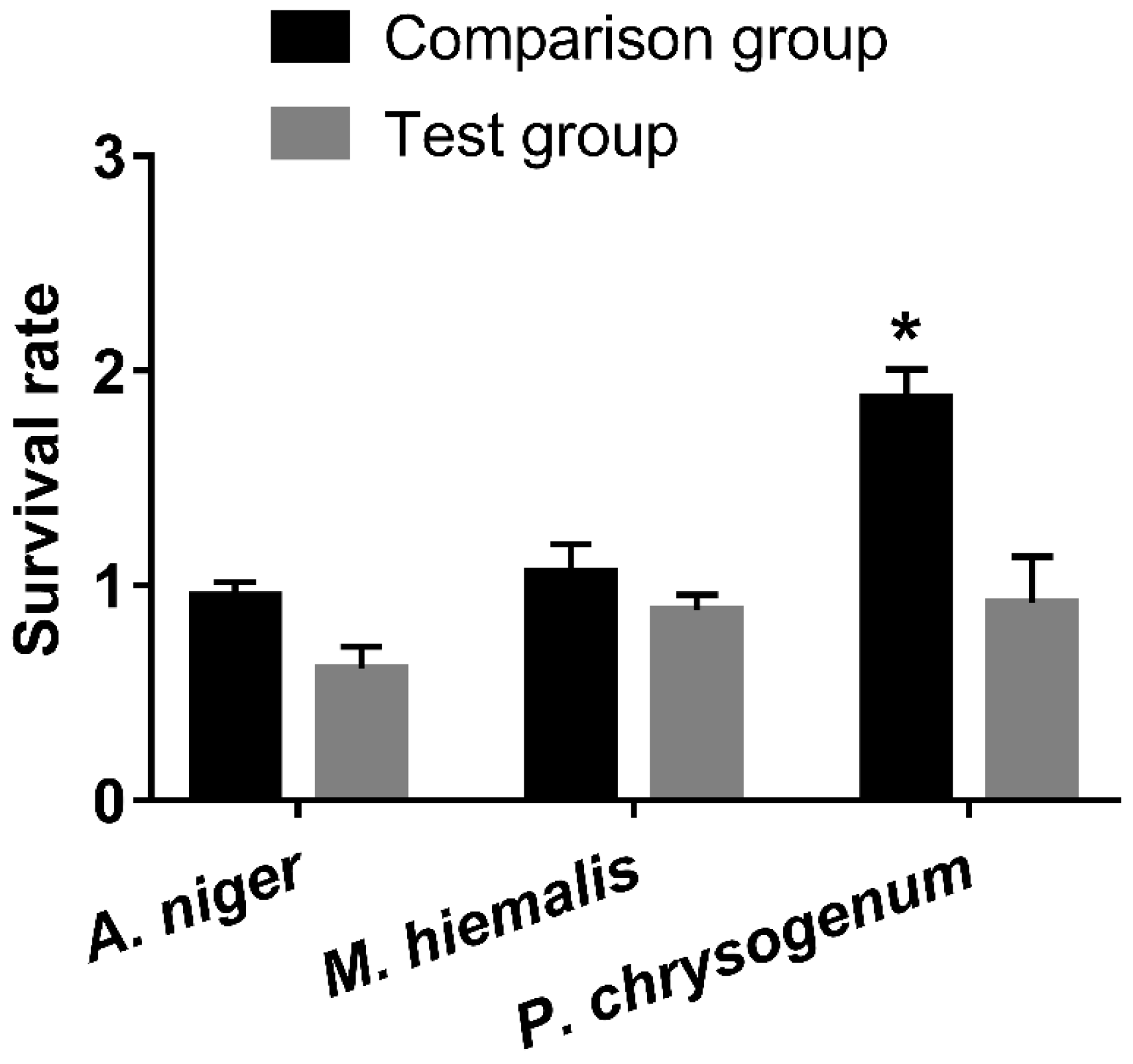
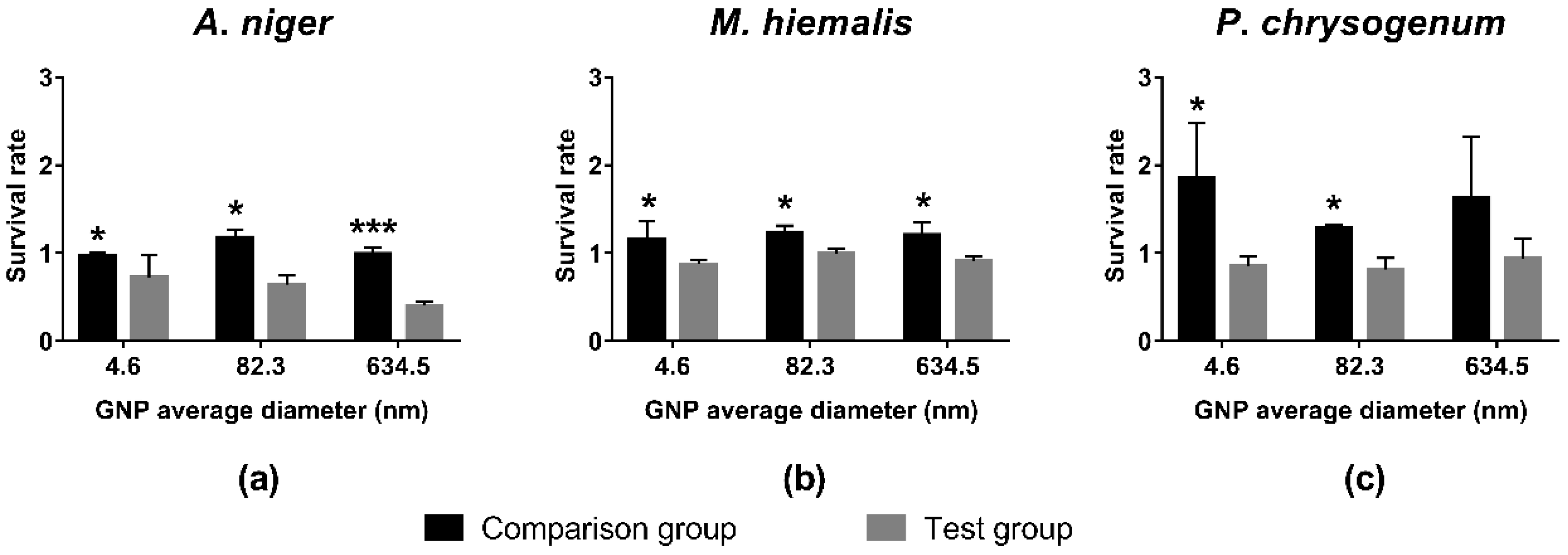
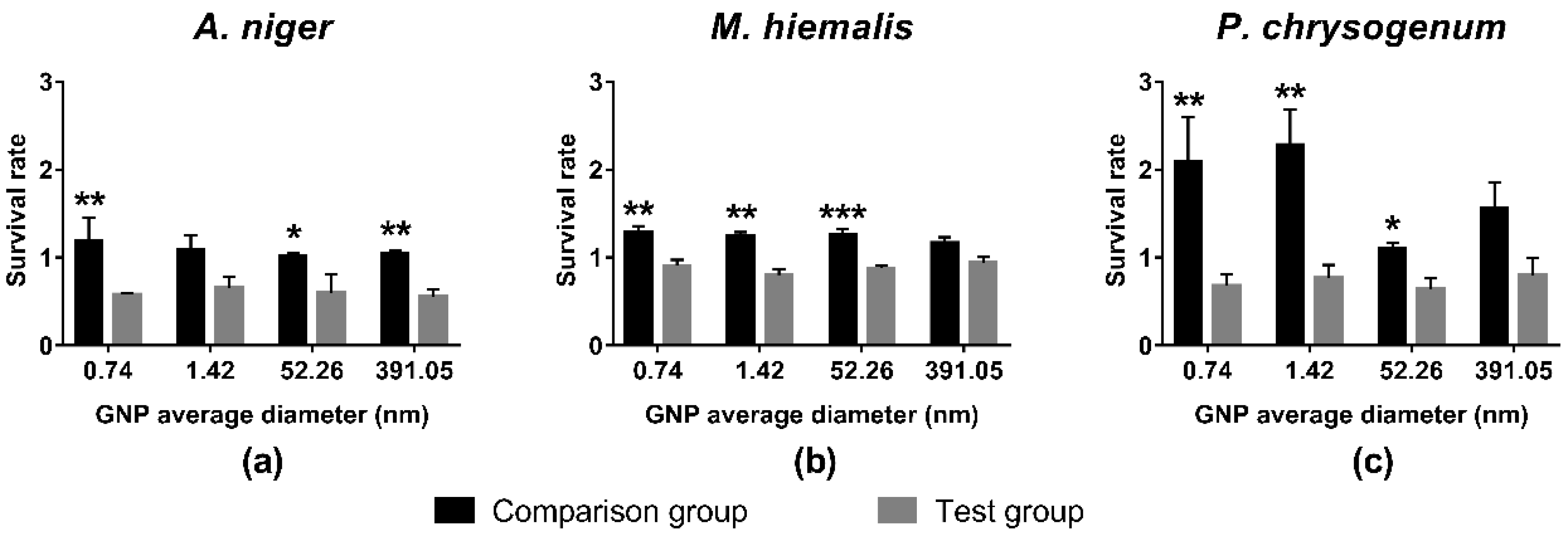
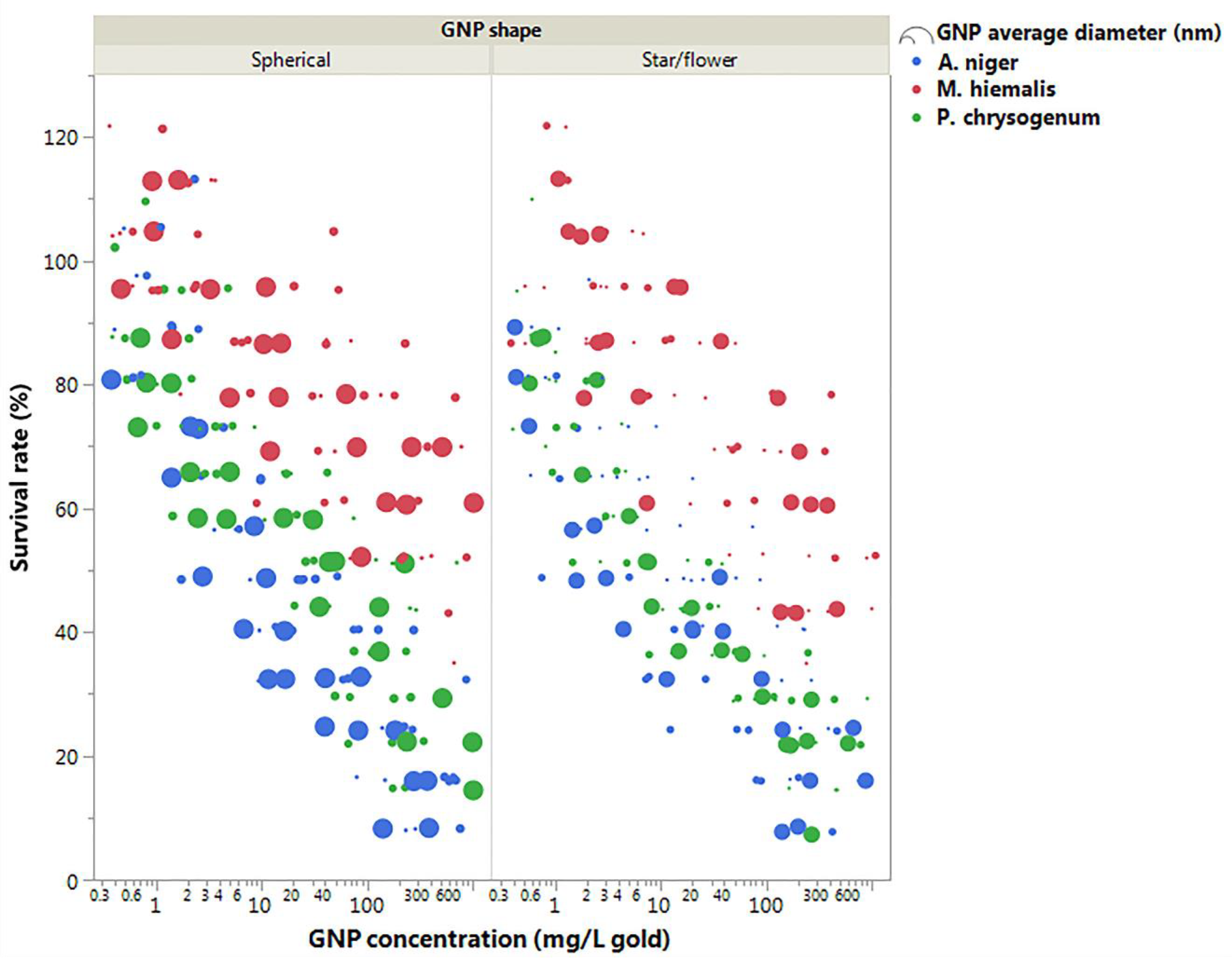
3.2. Toxicity Test of GNPs with Different Sizes and Shapes at Concentration of 19.697 mg/L Gold
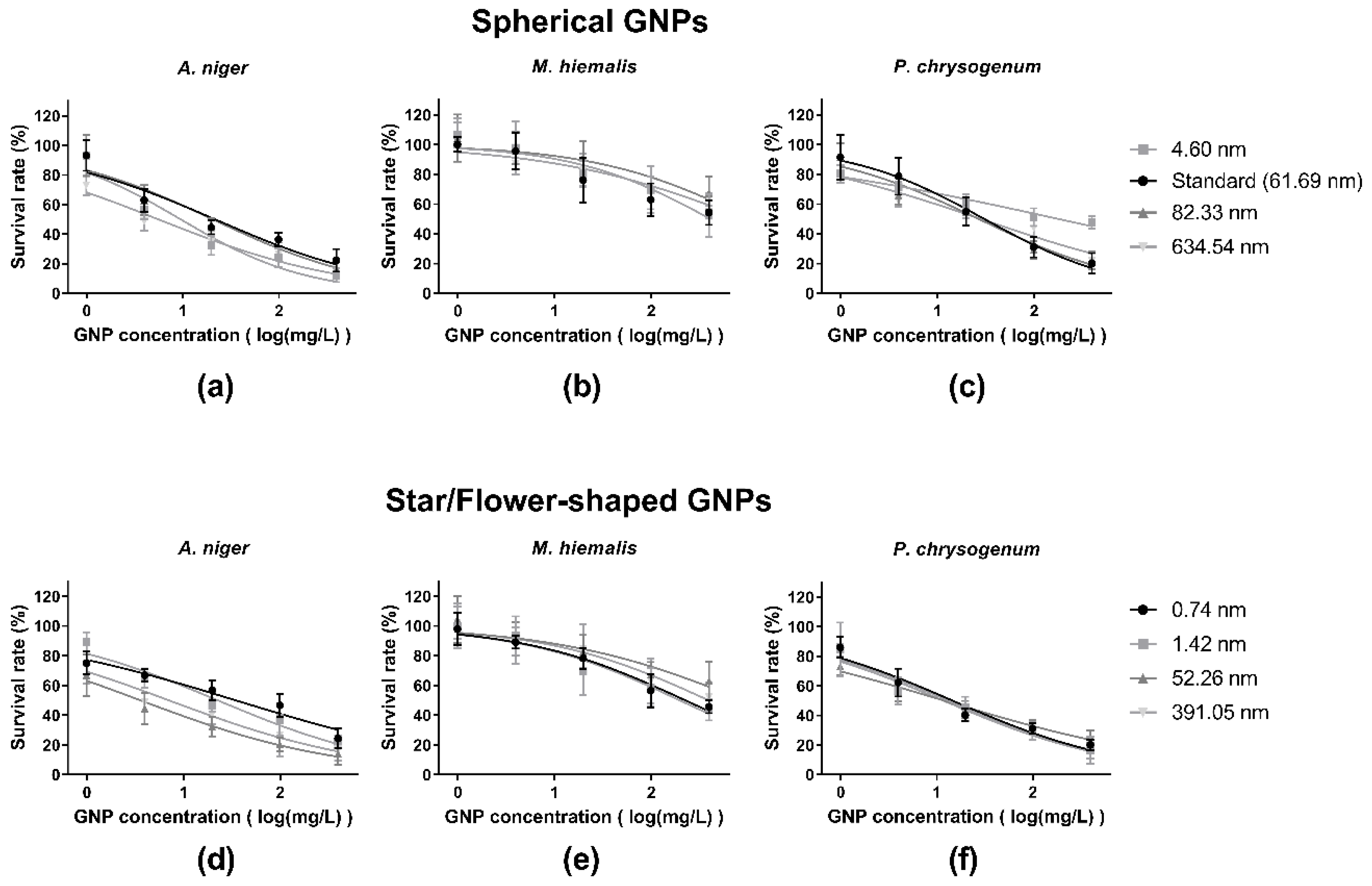
3.3. Dose–Response Curves
3.4. Statistical Analysis Results
4. Discussion
4.1. Ensuring Reproducibility of GNP Mass Concentrations
4.2. Confirming Toxicity of GNPs
4.3. Comparing the Role of Fungi Species and GNP Size and Shape on GNP Toxicity
4.3.1. Fungi Species
4.3.2. GNP Size
4.3.3. GNP Shape
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Toshima, N. Nanoscale Materials; Liz-Marzan, L.M., Kamat, P.V., Eds.; Kluwer Academic Publishers: London, UK, 2003; pp. 79–96. [Google Scholar]
- Rotello, V.M. Nanoparticles: Building Blocks for Nanotechnology; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2004. [Google Scholar]
- Rogach, A.; Talapin, D.; Weller, H.; Caruso, F. Colloids and Colloid Assemblies; Caruso, F., Ed.; Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2004. [Google Scholar]
- Schmid, G. Nanoparticles: From Theory to Application; John Wiley & Sons: Hoboken, NJ, USA, 2011. [Google Scholar]
- Sreeprasad, T.S.; Pradeep, T. Noble metal nanoparticles. In Springer Handbook of Nanomaterials; Springer: Berlin/Heidelberg, Germany, 2013; pp. 303–388. [Google Scholar]
- Willets, K.A.; Van Duyne, R.P. Localized surface plasmon resonance spectroscopy and sensing. Annu. Rev. Phys. Chem. 2007, 58, 267–297. [Google Scholar] [CrossRef] [PubMed]
- Petryayeva, E.; Krull, U.J. Localized surface plasmon resonance: Nanostructures, bioassays and biosensing—A review. Anal. Chim. Acta 2011, 706, 8–24. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Rycenga, M.; Skrabalak, S.E.; Wiley, B.; Xia, Y. Chemical synthesis of novel plasmonic nanoparticles. Annu. Rev. Phys. Chem. 2009, 60, 167–192. [Google Scholar] [CrossRef] [PubMed]
- Cassano, D.; Pocoví-Martínez, S.; Voliani, V. Ultrasmall-in-nano approach: Enabling the translation of metal nanomaterials to clinics. Bioconjugate Chem. 2017, 29, 4–16. [Google Scholar] [CrossRef] [PubMed]
- Morones, J.R.; Elechiguerra, J.L.; Camacho, A.; Holt, K.; Kouri, J.B.; Ramírez, J.T.; Yacaman, M.J. The bactericidal effect of silver nanoparticles. Nanotechnology 2005, 16, 2346. [Google Scholar] [CrossRef] [PubMed]
- Jain, P.; Pradeep, T. Potential of silver nanoparticle-coated polyurethane foam as an antibacterial water filter. Biotechnol. Bioeng. 2005, 90, 59–63. [Google Scholar] [CrossRef] [PubMed]
- Ahamed, M.; AlSalhi, M.S.; Siddiqui, M. Silver nanoparticle applications and human health. Clin. Chim. Acta 2010, 411, 1841–1848. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Lawson, R.; Ray, P.C.; Yu, H. Toxic effects of gold nanoparticles on salmonella typhimurium bacteria. Toxicol. Ind. Health 2011, 27, 547–554. [Google Scholar] [CrossRef] [PubMed]
- Shah, V.; Belozerova, I. Influence of metal nanoparticles on the soil microbial community and germination of lettuce seeds. Water Air Soil Pollut. 2009, 197, 143–148. [Google Scholar] [CrossRef]
- Zhao, Y.; Tian, Y.; Cui, Y.; Liu, W.; Ma, W.; Jiang, X. Small molecule-capped gold nanoparticles as potent antibacterial agents that target gram-negative bacteria. J. Am. Chem. Soc. 2010, 132, 12349–12356. [Google Scholar] [CrossRef] [PubMed]
- Tian, F.; Clift, M.J.; Casey, A.; del Pino, P.; Pelaz, B.; Conde, J.; Byrne, H.J.; Rothen-Rutishauser, B.; Estrada, G.; de la Fuente, J.M. Investigating the role of shape on the biological impact of gold nanoparticles in vitro. Nanomedicine 2015, 10, 2643–2657. [Google Scholar] [CrossRef] [PubMed]
- Chithrani, B.D.; Ghazani, A.A.; Chan, W.C. Determining the size and shape dependence of gold nanoparticle uptake into mammalian cells. Nano Lett. 2006, 6, 662–668. [Google Scholar] [CrossRef] [PubMed]
- Alkilany, A.M.; Murphy, C.J. Toxicity and cellular uptake of gold nanoparticles: What we have learned so far? J. Nanopart. Res. 2010, 12, 2313–2333. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Lu, W.; Tovmachenko, O.; Rai, U.S.; Yu, H.; Ray, P.C. Challenge in understanding size and shape dependent toxicity of gold nanomaterials in human skin keratinocytes. Chem. Phys. Lett. 2008, 463, 145–149. [Google Scholar] [CrossRef] [PubMed]
- Nowack, B.; Bucheli, T.D. Occurrence, behavior and effects of nanoparticles in the environment. Environ. Pollut. 2007, 150, 5–22. [Google Scholar] [CrossRef] [PubMed]
- Dunphy Guzman, K.A.; Taylor, M.R.; Banfield, J.F. Environmental Risks of Nanotechnology: National Nanotechnology Initiative Funding, 2000–2004; ACS Publications: Washington, DC, USA, 2006. [Google Scholar]
- Maynard, A.D. A research strategy for addressing risk. Nanotechnol. Woodrow Wilson Int. Cent. Sch. 2006, 444, 267–269. [Google Scholar]
- Oberdörster, G.; Oberdörster, E.; Oberdörster, J. Nanotoxicology: An emerging discipline evolving from studies of ultrafine particles. Environ. Health Perspect. 2005, 113, 823–839. [Google Scholar] [CrossRef] [PubMed]
- Navarro, E.; Baun, A.; Behra, R.; Hartmann, N.B.; Filser, J.; Miao, A.-J.; Quigg, A.; Santschi, P.H.; Sigg, L. Environmental behavior and ecotoxicity of engineered nanoparticles to algae, plants, and fungi. Ecotoxicology 2008, 17, 372–386. [Google Scholar] [CrossRef] [PubMed]
- Colvin, V.L. The potential environmental impact of engineered nanomaterials. Nat. Biotechnol. 2003, 21, 1166–1170. [Google Scholar] [CrossRef] [PubMed]
- Cheetham, N.W. Introducing Biological Energetics: How Energy and Information Control the Living World; Oxford University Press: Oxford, UK, 2010. [Google Scholar]
- Moore, B.; Duncan, J.; Burgess, J. Fungal bioaccumulation of copper, nickel, gold and platinum. Miner. Eng. 2008, 21, 55–60. [Google Scholar] [CrossRef]
- Samson, R.A.; Houbraken, J.; Summerbell, R.C.; Flannigan, B.; Miller, J.D. Common and important species of fungi and actinomycetes in indoor environments. In Microorganisms in Home and Indoor Work Environments: Diversity, Health Impacts, Investigation and Control; CRC Press: Boca Raton, FL, USA, 2002; pp. 285–473. [Google Scholar]
- Saleh, Y.; Mayo, M.; Ahearn, D. Resistance of some common fungi to gamma irradiation. Appl. Environ. Microbiol. 1988, 54, 2134–2135. [Google Scholar] [PubMed]
- Young, J.C.; Fulcher, R. Mycotoxins in grains: Causes, consequences, and cures. Cereal Foods World 1984, 29, 725–728. [Google Scholar]
- Chen, R.; Wu, J.; Li, H.; Cheng, G.; Lu, Z.; Che, C.-M. Fabrication of gold nanoparticles with different morphologies in hepes buffer. Rare Met. 2010, 29, 180–186. [Google Scholar] [CrossRef]
- Habib, A.; Tabata, M.; Wu, Y.G. Formation of gold nanoparticles by good’s buffers. Bull. Chem. Soc. Jpn. 2005, 78, 262–269. [Google Scholar] [CrossRef]
- Goldman, E.; Green, L.H. Practical Handbook of Microbiology; CRC Press: Boca Raton, FL, USA, 2015. [Google Scholar]
- He, Z.; Liu, K.; Manaloto, E.; Casey, A.; Cribaro, G.P.; Byrne, H.J.; Tian, F.; Barcia, C.; Conway, G.E.; Cullen, P.J. Cold atmospheric plasma induces ATP-dependent endocytosis of nanoparticles and synergistic u373mg cancer cell death. Sci. Rep. 2018, 8, 5298. [Google Scholar] [CrossRef] [PubMed]
- Balasubramanian, S.K.; Yang, L.; Yung, L.-Y.L.; Ong, C.-N.; Ong, W.-Y.; Liya, E.Y. Characterization, purification, and stability of gold nanoparticles. Biomaterials 2010, 31, 9023–9030. [Google Scholar] [CrossRef] [PubMed]
- Haiss, W.; Thanh, N.T.; Aveyard, J.; Fernig, D.G. Determination of size and concentration of gold nanoparticles from UV–VIS spectra. Anal. Chem. 2007, 79, 4215–4221. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Zhang, Q.; Lee, J.Y.; Wang, D.I. The synthesis of sers-active gold nanoflower tags for in vivo applications. ACS Nano 2008, 2, 2473–2480. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Lee, J.Y.; Wang, D.I. Seedless, surfactantless, high-yield synthesis of branched gold nanocrystals in hepes buffer solution. Chem. Mater. 2007, 19, 2823–2830. [Google Scholar] [CrossRef]
- Maiorano, G.; Rizzello, L.; Malvindi, M.A.; Shankar, S.S.; Martiradonna, L.; Falqui, A.; Cingolani, R.; Pompa, P.P. Monodispersed and size-controlled multibranched gold nanoparticles with nanoscale tuning of surface morphology. Nanoscale 2011, 3, 2227–2232. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.; Li, N.; Astruc, D. State of the art in gold nanoparticle synthesis. Coord. Chem. Rev. 2013, 257, 638–665. [Google Scholar] [CrossRef]
- Good, N.E.; Winget, G.D.; Winter, W.; Connolly, T.N.; Izawa, S.; Singh, R.M. Hydrogen ion buffers for biological research. Biochemistry 1966, 5, 467–477. [Google Scholar] [CrossRef] [PubMed]
- Foster, J.W. The heavy metal nutrition of fungi. Bot. Rev. 1939, 5, 207–239. [Google Scholar] [CrossRef]
- Mann, T. Studies on the metabolism of mould fungi: 1. Phosphorus metabolism in moulds. Biochem. J. 1944, 38, 339–345. [Google Scholar] [CrossRef] [PubMed]
- Gadd, G. Fungi and Yeast for Metal Accumulation in Microbial Mineral Recovery; Ehrlich, H.L., Bierley, C.L., Eds.; McGraw Hill: New York, NY, USA, 1990. [Google Scholar]
- Spinti, M.; Zhuang, H.; Trujillo, E.M. Evaluation of immobilized biomass beads for removing heavy metals from wastewaters. Water Environ. Res. 1995, 67, 943–952. [Google Scholar] [CrossRef]
- Ashida, J. Adaptation of fungi to metal toxicants. Annu. Rev. Phytopathol. 1965, 3, 153–174. [Google Scholar] [CrossRef]
- GADD, G.M. Interactions of fungip with toxic metals. New Phytol. 1993, 124, 25–60. [Google Scholar] [CrossRef]
- Fomina, M.; Charnock, J.; Bowen, A.D.; Gadd, G.M. X-ray absorption spectroscopy (XAS) of toxic metal mineral transformations by fungi. Environ. Microbiol. 2007, 9, 308–321. [Google Scholar] [CrossRef] [PubMed]
- Zafar, S.; Aqil, F.; Ahmad, I. Metal tolerance and biosorption potential of filamentous fungi isolated from metal contaminated agricultural soil. Bioresour. Technol. 2007, 98, 2557–2561. [Google Scholar] [CrossRef] [PubMed]
- Price, M.S.; Classen, J.J.; Payne, G.A. Aspergillus niger absorbs copper and zinc from swine wastewater. Bioresour. Technol. 2001, 77, 41–49. [Google Scholar] [CrossRef]
- Tahir, A. Resistant fungal biodiversity of electroplating effluent and their metal tolerance index. In Electroplating; InTech: London, UK, 2012. [Google Scholar]
- Fourest, E.; Canal, C.; Roux, J.-C. Improvement of heavy metal biosorption by mycelial dead biomasses (Rhizopus arrhizus, Mucor miehei and Penicillium chrysogenum): Ph control and cationic activation. FEMS Microbiol. Rev. 1994, 14, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Mehra, R.K.; Winge, D.R. Metal ion resistance in fungi: Molecular mechanisms and their regulated expression. J. Cell. Biochem. 1991, 45, 30–40. [Google Scholar] [CrossRef] [PubMed]
- Habib, A.; Tabata, M. Oxidative DNA damage induced by hepes (2-[4-(2-hydroxyethyl)-1-piperazinyl] ethanesulfonic acid) buffer in the presence of Au (III). J. Inorg. Biochem. 2004, 98, 1696–1702. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample 1 | Shape | Concentration of Phosphate Added (mM) | LSPR Peak (nm) | Diameter (nm ± SD) | Zeta Potential (mV ± SD) | Average Equivalent Spherical Total Surface Area 2 Per 1 mL (cm2) |
|---|---|---|---|---|---|---|
| Standard | Spherical | 0 | 558.8 | 61.69 ± 22.35 | −23.50 ± 0.21 | 1.98 |
| NaH2PO4 added | Spherical | 2 | 565.0 | 82.33 ± 35.86 | −26.43 ± 0.13 | 1.49 |
| 6 | 531.2 | 4.60 ± 1.32 | −29.67 ± 0.13 | 26.60 | ||
| 40 | 532.2 | 634.54 ± 224.30 | −36.87 ± 0.45 | 0.19 | ||
| Na2HPO4 added | Star-shaped/Flower-shaped | 1 | 625.8 | 52.26 ± 14.94 | −40.80 ± 0.24 | 2.34 |
| 6 | 587.6 | 1.42 ± 0.32 | −34.97 ± 0.35 | 86.09 | ||
| 50 | 543.4 | 0.74 ± 0.25 | −23.73 ± 0.60 | 165.29 | ||
| 240 | 664.0 | 391.05 ± 153.47 | −22.13 ± 0.27 | 0.31 |
| GNP Shape | Diameter Mean | A. niger | M. hiemalis | P. chrysogenum | |||
|---|---|---|---|---|---|---|---|
| IC50 | 95% Confidence Interval | IC50 | 95% Confidence Interval | IC50 | 95% Confidence Interval | ||
| Spherical | 61.69 (Standard) | 21.55 | 13.34–34.80 | N/A 2 | N/A | 30.39 | 20.04–46.09 |
| 4.60 | 9.46 | 6.242–14.34 | 408.61 | 168.0–994.1 | 177.25 | 102.9–305.3 | |
| 82.33 | 20.40 | 12.62–32.97 | 1077.59 | 220.5–5267 | 26.51 | 18.15–38.71 | |
| 634.54 | 5.54 | 3.928–7.814 | 950.40 | 269.3–3354 | 27.82 | 18.92–40.91 | |
| Star-shaped/Flower-shaped | 0.74 | 34.42 | 21.69–54.62 | 224.66 | 140.2–360.0 | 14.58 | 10.15–20.94 |
| 1.42 | 22.42 | 15.10–33.30 | 196.65 | 94.35–409.9 | 13.16 | 7.729–22.40 | |
| 52.26 | 3.56 | 2.078–6.114 | 904.56 | 214.7–3810 | 11.92 | 8.335–17.05 | |
| 391.05 | 7.01 | 3.947–12.44 | 391.57 | 155.1–988.4 | 11.65 | 8.073–16.80 | |
| Source of Variation | Analysis Subject | A. niger | M. hiemalis | P. chrysogenum | |||
|---|---|---|---|---|---|---|---|
| Spherical GNP | Star/Flower-Shaped GNP | Spherical GNP | Star/Flower-Shaped GNP | Spherical GNP | Star/Flower-Shaped GNP | ||
| GNP size | F value | 14.82 | 29.85 | 2.099 | 2.183 | 9.069 | 0.2945 |
| p value | <0.0001 | <0.0001 | 0.1137 | 0.1031 | <0.0001 | 0.8292 | |
| p value summary 2 | **** | **** | ns | ns | **** | ns | |
| GNP size × GNP concentration 1 | F value | 1.397 | 1.99 | 0.7349 | 0.5058 | 6.309 | 1.131 |
| p value | 0.2029 | 0.0481 | 0.7103 | 0.8999 | <0.0001 | 0.3606 | |
| p value summary 2 | ns | * | ns | ns | **** | ns | |
| Source of Variation | Degrees of Freedom | F Value | p Value | p Value Summary 2 |
|---|---|---|---|---|
| GNP shape | 1 | 11.1958 | 0.0009 | *** |
| Fungi species | 2 | 184.7116 | <0.0001 | **** |
| GNP concentration | 1 | 447.8957 | <0.0001 | **** |
| GNP shape × Fungi species 1 | 2 | 2.7142 | 0.0673 | ns |
| GNP shape × GNP concentration | 1 | 0.1539 | 0.6950 | ns |
| Fungi species × GNP concentration | 2 | 1.1037 | 0.3325 | ns |
| GNP shape × Fungi species × GNP concentration | 2 | 0.6936 | 0.5003 | ns |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, K.; He, Z.; Byrne, H.J.; Curtin, J.F.; Tian, F. Investigating the Role of Gold Nanoparticle Shape and Size in Their Toxicities to Fungi. Int. J. Environ. Res. Public Health 2018, 15, 998. https://doi.org/10.3390/ijerph15050998
Liu K, He Z, Byrne HJ, Curtin JF, Tian F. Investigating the Role of Gold Nanoparticle Shape and Size in Their Toxicities to Fungi. International Journal of Environmental Research and Public Health. 2018; 15(5):998. https://doi.org/10.3390/ijerph15050998
Chicago/Turabian StyleLiu, Kangze, Zhonglei He, Hugh J. Byrne, James F. Curtin, and Furong Tian. 2018. "Investigating the Role of Gold Nanoparticle Shape and Size in Their Toxicities to Fungi" International Journal of Environmental Research and Public Health 15, no. 5: 998. https://doi.org/10.3390/ijerph15050998