
Planktonic Foraminiferal Biostratigraphy of the Upper Cretaceous of the Central European Basin


1
Institute of Paleobiology, Polish Academy of Sciences, Twarda 51/55, 00-818 Warszawa, Poland
2
Faculty of Geology, University of Warsaw, Al. Żwirki i Wigury 93, 02-089 Warszawa, Poland
3
Polish Geological Institute-National Research Institute, Rakowiecka 4, 00-975 Warszawa, Poland
*
Author to whom correspondence should be addressed.
Geosciences 2022, 12(1), 22; https://doi.org/10.3390/geosciences12010022
Submission received: 22 November 2021
/
Revised: 26 December 2021
/
Accepted: 30 December 2021
/
Published: 5 January 2022
(This article belongs to the Special Issue Application of Foraminifera in Biochronology)
Abstract
:Planktonic foraminifera are one of the most stratigraphically important groups of organisms for the Cretaceous system. However, standard foraminiferal zonations based mostly on species from the Tethyan bioprovince are hardly applicable in temperate regions where warm-water taxa are scarce or lacking. We propose a foraminiferal zonation based on foraminiferal events recognized in the northern Foraminiferal Transitional Bioprovince, which likely has a high correlation potential at least at a regional scale. Fifteen planktonic foraminiferal zones are distinguished from the upper Albian up to the uppermost Maastrichtian strata in extra-Carpathian Poland and western Ukraine. From the bottom to the top, Thalmanninella appenninica, Th. globotruncanoides, Th. reicheli, Rotalipora cushmani, Whiteinella archaeocretacea, Helvetoglobotruncana helvetica, Marginotruncana coronata, M. sinuosa, Pseudotextularia nuttalli, Globotruncana linneiana, G. arca, Contusotruncana plummerae, Rugoglobigerina pennyi, Globotruncanella petaloidea and Guembelitria cretacea. These zones are calibrated by macrofaunal zonations.
1. Introduction
Planktonic foraminifera are widely recognized as one of the most stratigraphically important groups of organisms for the Late Cretaceous period with a high correlation potential. However, standard foraminiferal zonations [1,2,3,4] mostly utilize tropical/subtropical taxa and, therefore, are hardly applicable in temperate and boreal regions where index taxa are scarce or lacking.
The distribution of planktonic foraminifera in the global ocean is controlled by many physical and biological factors, such as water temperature, salinity, water depth, nutrient composition, water transparency, turbidity, hydrodynamics of water masses, palaeogeography, the occurrence of migratory routes and mutual relations of all listed factors [5,6]. Recent assemblages of planktonic foraminifera have occurred in a few latitudinally distributed major faunal provinces, reflecting the general variation of the latitudinal sea surface temperature gradient and progressive loss of biodiversity from low to high latitudes. In addition, the boundaries of foraminiferal provinces are altered regularly by the seasonal variability of foraminiferal occurrences. This horizontal distributional pattern has also been recorded in Cretaceous planktonic foraminiferal assemblages with five generalized bioprovinces (Austral, Tethyan, Boreal and two Transitional provinces located between them) [1,7,8]. In addition, due to palaeoenvironmental changes during the Cretaceous period, such as climate swings and changes in ocean circulation or sea-level fluctuations, the biogeography of planktonic foraminiferal assemblages has varied markedly through time [9,10]. All these factors limit the application of standard planktonic foraminiferal zonation, established mainly for the Tethyan bioprovince, within higher latitudinal domains. Biozonation schemes constructed for individual/particular latitude regions appear to be more successful for precise biostratigraphic correlations.
The studies on planktonic foraminifera conducted by us and our collaborators [9,11,12,13,14,15,16,17,18,19,20,21,22,23,24,25,26,27,28,29,30,31,32,33,34] enable us to update and refine the foraminiferal zonations proposed for extra-Carpathian Poland. Accordingly, in this paper we present a revised comprehensive planktonic foraminiferal zonation of the Upper Cretaceous period based on the foraminiferal distribution in a set of natural and artificial exposures of macrofaunal well-dated strata from extra-Carpathian Poland and western Ukraine. During the Late Cretaceous period, this area was located in the central part of the European epicontinental sea [35,36,37] and belonged to the North Transitional Foraminiferal Bioprovince [1,7], characterized only by the sporadic immigration of typical Tethyan taxa. The proposed biozonation scheme has a high potential of applicability in a large part of the latitudinally expanded European Basin.
2. Materials and Methods
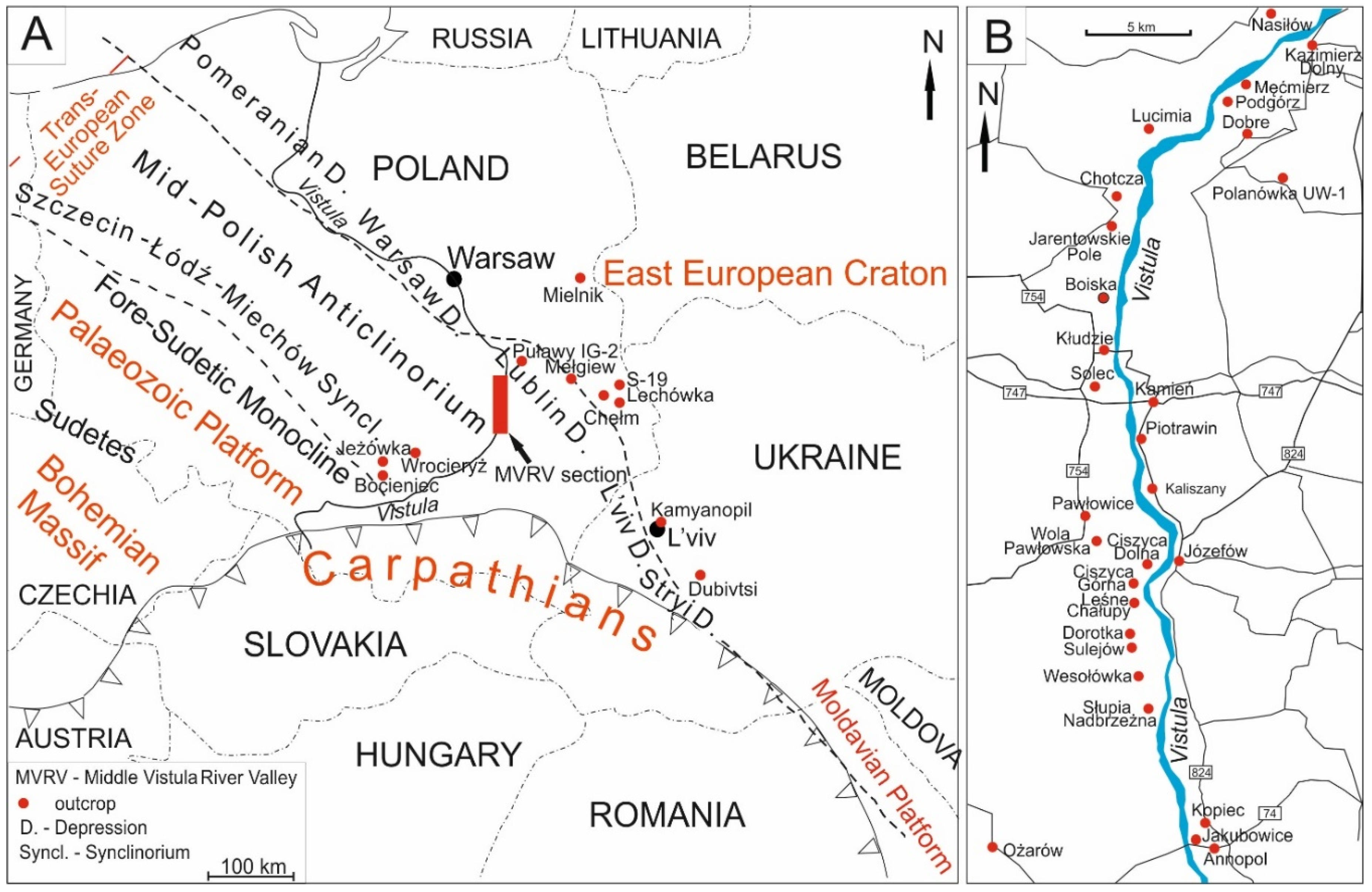
The planktonic foraminiferal assemblages that were used to construct the proposed zonation came from the Upper Cretaceous sections cropping out in a series of small to large natural exposures, several quarries and three boreholes located in central, eastern and south-eastern Poland and western Ukraine (Figure 1). In total, several hundred samples from 34 sections of the Upper Cretaceous of extra-Carpathian Poland and western Ukraine (Figure 1) were analysed by us. In addition to the re-evaluation of the previously studied localities, one borehole section (Polanówka UW-1 borehole), which had not been subject to our previous investigation, was additionally studied for the purpose of this paper.
The studied Cretaceous strata represent the southern parts of the Szczecin–Łódź–Miechów Synclinorium (Bocieniec [38], Jeżówka [29,39], Wrocieryż [21]), the Mid-Polish Anticlinorium (the so-called Middle Vistula River section representing middle Albian through Danian successions cropping out along both banks of the Middle Vistula River between the towns of Zawichost and Puławy [11,18,21,40,41,42,43]), and the SW margin of the East European Platform, within a tectonic unit called the Border Synclinorium [44], and, specifically, within the Lublin Syncline (Mielnik [29] and Chełm [23,45] quarries, Lechówka [31] and Mełgiew [25] outcrops and the Puławy IG-2 [17] and S-19 [28] boreholes) and the L’viv–Stryi Syncline [37,46] (Kamyanopil outcrop [25], Dubivtsi quarries [9,24,27,32]).
A new section published here for the first time is the Polanówka UW-1 borehole succession, situated 10 km south of Kazimierz Dolny, at the right bank of the Vistula River (Figure 1B) and 6–7 km east of two outcrops—Chotcza and Jarentowskie Pole—located at the left bank of the river. The borehole encompasses about an 80 m thick succession of upper Maastrichtian marly chalk. Samples for planktonic foraminiferal studies were collected every 5 m on average.
All samples were disaggregated in Glauber’s salt and then sieved at 63 µm. The 63 µm size fractions, containing about 300 specimens per sample, were used for foraminiferal analysis.
3. Results
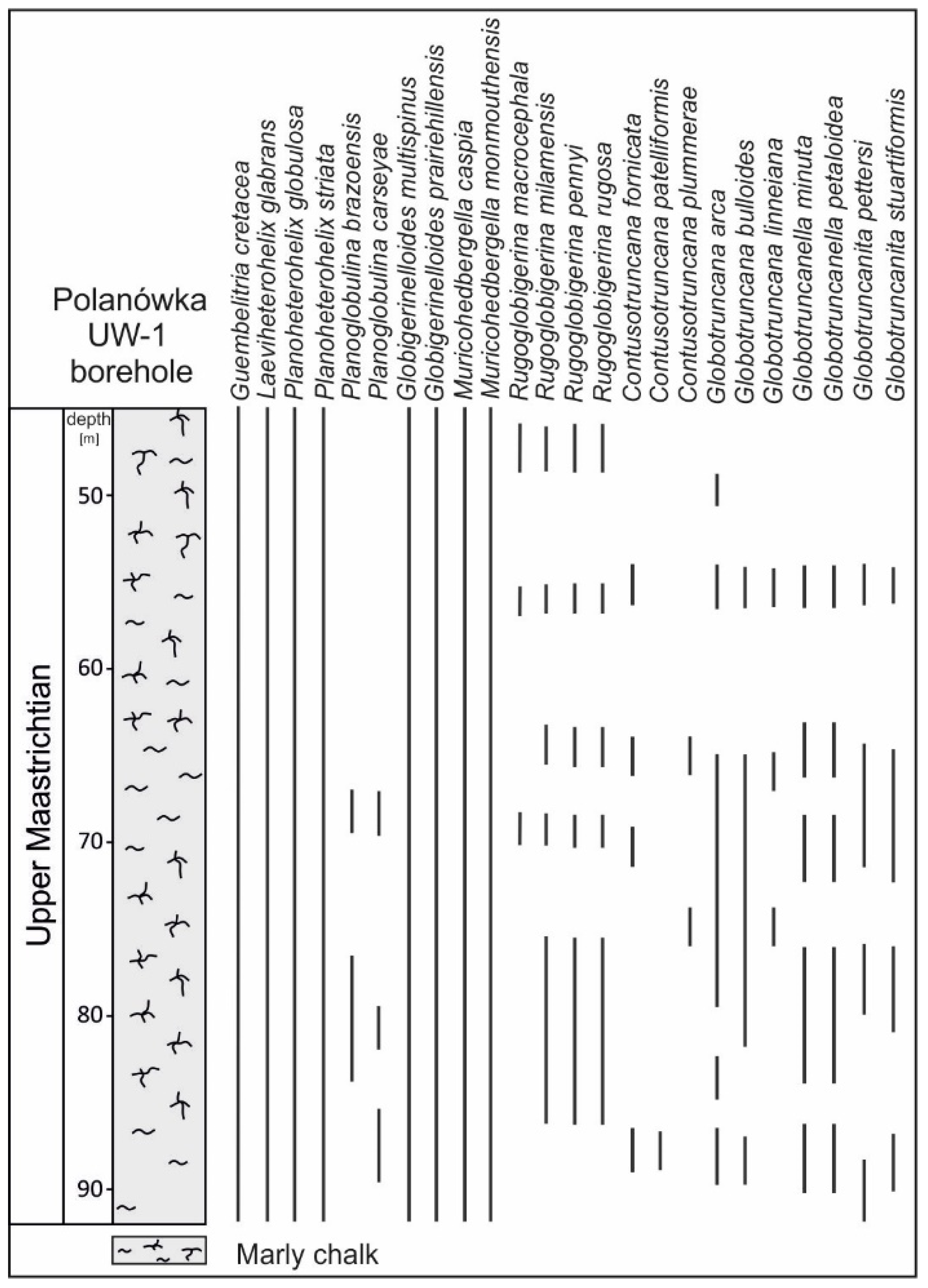
The taxonomic composition of planktonic foraminiferal assemblages in the newly studied Polanówka UW-1 borehole succession varied significantly. Shallow-water taxa, such as small, biserial heterohelicids (Planoheterohelix striata, P. globulosa, Laeviheterohelix glabrans), triserial guembelitriids (Guembelitria cretacea) and globigerinelloidids (Globigerinelloides multispinus, G. prairiehillensis), occurred throughout the section. Rugoglobigerina (R. pennyi, R. milamensis, R. rugosa, R. macrocephala) occurred abundantly in the lower part of the core between a depth of 86 and 75 m. The deep-dwelling keeled foraminifera (common Contusotruncana fornicata, C. patelliformis, C. plummerae, Globotruncana arca, G. linneiana, G. bulloides and very scarce Globotruncanita pettersi and G. stuartiformis) were recorded between a depth of 90 and 75 m, as well as in a short interval at ca. 55 m depth. Complex heterohelicids (Planoglobulina brazoensis, P. carseyae) were observed in the interval between 68 and 90 m. A detailed distribution of the recorded species is shown in Figure 2.
The taxonomic composition of the planktonic foraminiferal assemblages from the studied interval of the Polanówka UW-1 borehole was very similar to the ones found in the nearby two outcrops—Chotcza and Jarentowskie Pole—whose stratigraphical position was well documented by ammonites and belemnites as upper Maastrichtian (Figure 3).
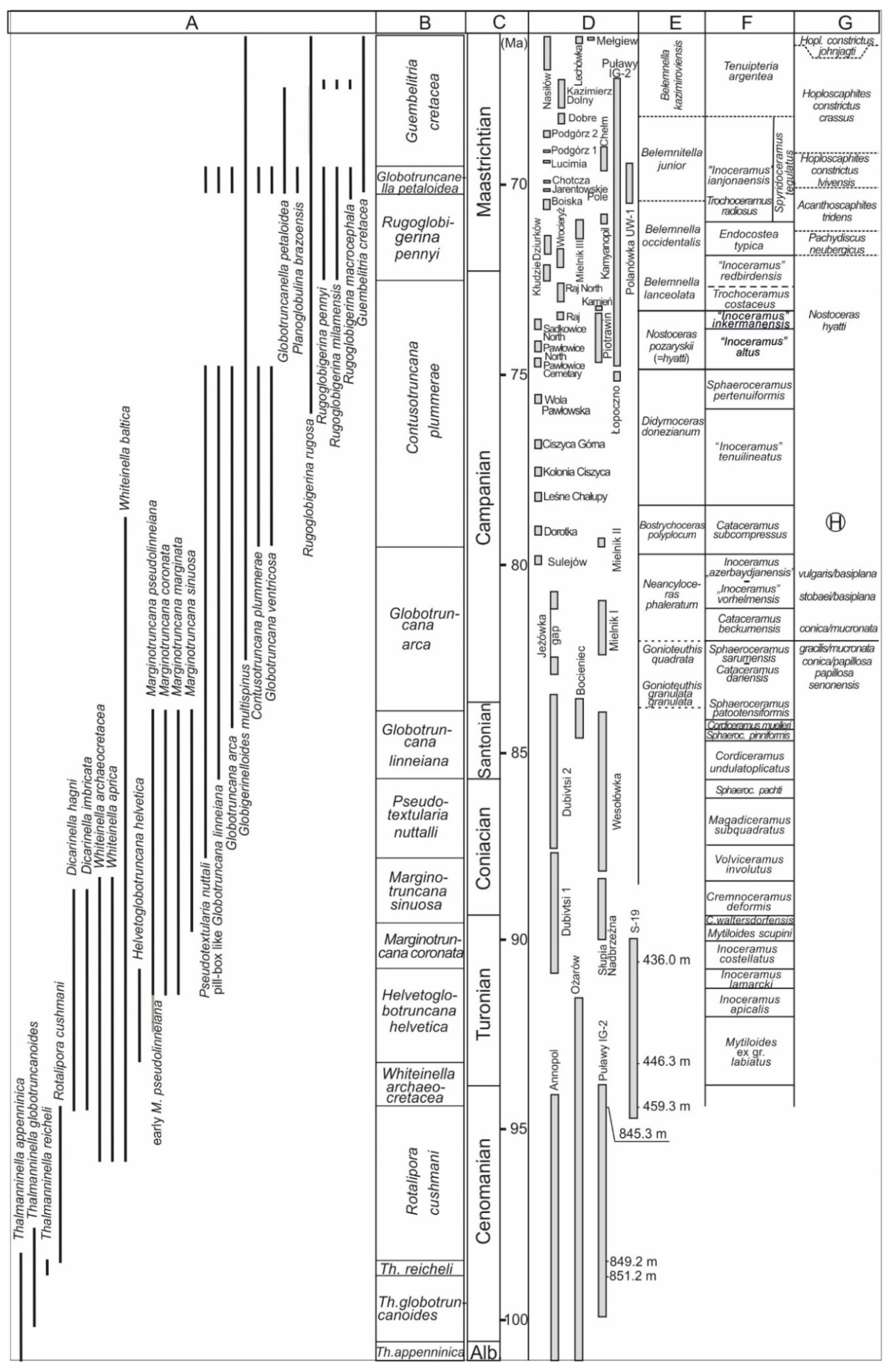
Figure 3 shows ranges of zonal markers in the proposed zonation against macrofossil zonations established in the study area. Figure 4 presents a correlation of proposed zones with those of [3,4]. Selected species of planktonic foraminifera from the Albian through to the Maastrichtian succession of the study area are illustrated in Figure 5, Figure 6 and Figure 7.
In the entire upper Albian–Maastrichtian interval, planktonic foraminifera were consistently present. The upper Albian through to the middle Turonian planktonic foraminiferal assemblages were well diversified and, generally, their taxonomic composition was similar to the one from the Tethyan area. Starting from late Turonian until the Maastrichtian, they were dominated by cosmopolitan taxa, more tolerant of cooler sea water at higher latitudes of the Transitional Foraminiferal Bioprovince. The absence or sporadic occurrences of Tethyan taxa used as zonal markers in the standard planktonic foraminiferal zonations caused us to develop a local zonation that, when calibrated by macrofossil zonations, could be applied for regional or even inter-regional correlations.
Fifteen planktonic foraminiferal zones were distinguished from the upper Albian to the uppermost Maastrichtian of the study area. In the interval from the upper Albian through to the middle Turonian, the zones were the same as in standard zonations, based on warm-water deep-dwelling species of planktonic foraminifera, i.e., Thalmanninella appenninica, Th. globotruncanoides, Th. reicheli, Rotalipora cushmani, Whiteinella archaeocretacea and Helvetoglobotruncana helvetica (e.g., [1,3,4]). The biozonation of the upper Turonian through Maastrichtian was based on more cosmopolitan planktonic foraminifera and comprised the following zones: Marginotruncana coronata, M. sinuosa, Pseudotextularia nuttalli, Globotruncana linneiana, G. arca, Contusotruncana plummerae, Rugoglobigerina pennyi, Globotruncanella petaloidea and Guembelitria cretacea.
Reference list of planktonic foraminifera and macrofossils mentioned in the text is shown in Appendix A.
4. Planktonic Foraminiferal Zonation
- Thalmanninella appenninica Interval Zone
- Author: Sigal [47].
- Definition: The body of strata from the lowest occurrence of Thalmanninella appenninica to the lowest occurrence of Thalmanninella globotruncanoides (Figure 3).
- Chronostratigraphy: Upper Albian.Figure 3. Summary of the planktonic foraminiferal ranges and the proposed biozonation and macrofaunal biozonations for the upper Albian to Maastrichtian of extra-Carpathian Poland and western Ukraine. (A) Vertical ranges of index species of planktonic foraminifera; (B) proposed biozonation scheme; (C) stages and ages after [48]; (D) stratigraphical intervals covered by studied successions in several outcrops, quarries and boreholes; (E) cephalopod zonation after [49]; (F) inoceramid zonation after [18,32,42,43]; (G) ammonite zonation after [50,51]; (H) belemnite/echinoid zonation after [52]. Genera abbreviations: Th.—Thalmanninella; Sphaeroc.—Sphaeroceramus; C.—Cremnoceramus; Hopl.—Hoploscaphites.Figure 3. Summary of the planktonic foraminiferal ranges and the proposed biozonation and macrofaunal biozonations for the upper Albian to Maastrichtian of extra-Carpathian Poland and western Ukraine. (A) Vertical ranges of index species of planktonic foraminifera; (B) proposed biozonation scheme; (C) stages and ages after [48]; (D) stratigraphical intervals covered by studied successions in several outcrops, quarries and boreholes; (E) cephalopod zonation after [49]; (F) inoceramid zonation after [18,32,42,43]; (G) ammonite zonation after [50,51]; (H) belemnite/echinoid zonation after [52]. Genera abbreviations: Th.—Thalmanninella; Sphaeroc.—Sphaeroceramus; C.—Cremnoceramus; Hopl.—Hoploscaphites.
![Geosciences 12 00022 g003]() Figure 4. Correlation of the proposed zonation with the standard planktonic foraminiferal zonations [3,4] based on Tethyan species; (A)— stages and ages after [48]; (B)— planktonic foraminiferal zonation (this paper); (C,D)—standard planktonic foraminiferal zonations: (C)—after [3]; (D)—after [4]. Genera abbreviations: Th.—Thalmanninella; Marg.—Marginotruncana; Glob.—Globotruncana; Gl-nella—Globotruncanella; Ps-guemb.—Pseudoguembelina; Pseudotext.—Pseudotextularia; Plum.—Plummerita.Figure 4. Correlation of the proposed zonation with the standard planktonic foraminiferal zonations [3,4] based on Tethyan species; (A)— stages and ages after [48]; (B)— planktonic foraminiferal zonation (this paper); (C,D)—standard planktonic foraminiferal zonations: (C)—after [3]; (D)—after [4]. Genera abbreviations: Th.—Thalmanninella; Marg.—Marginotruncana; Glob.—Globotruncana; Gl-nella—Globotruncanella; Ps-guemb.—Pseudoguembelina; Pseudotext.—Pseudotextularia; Plum.—Plummerita.
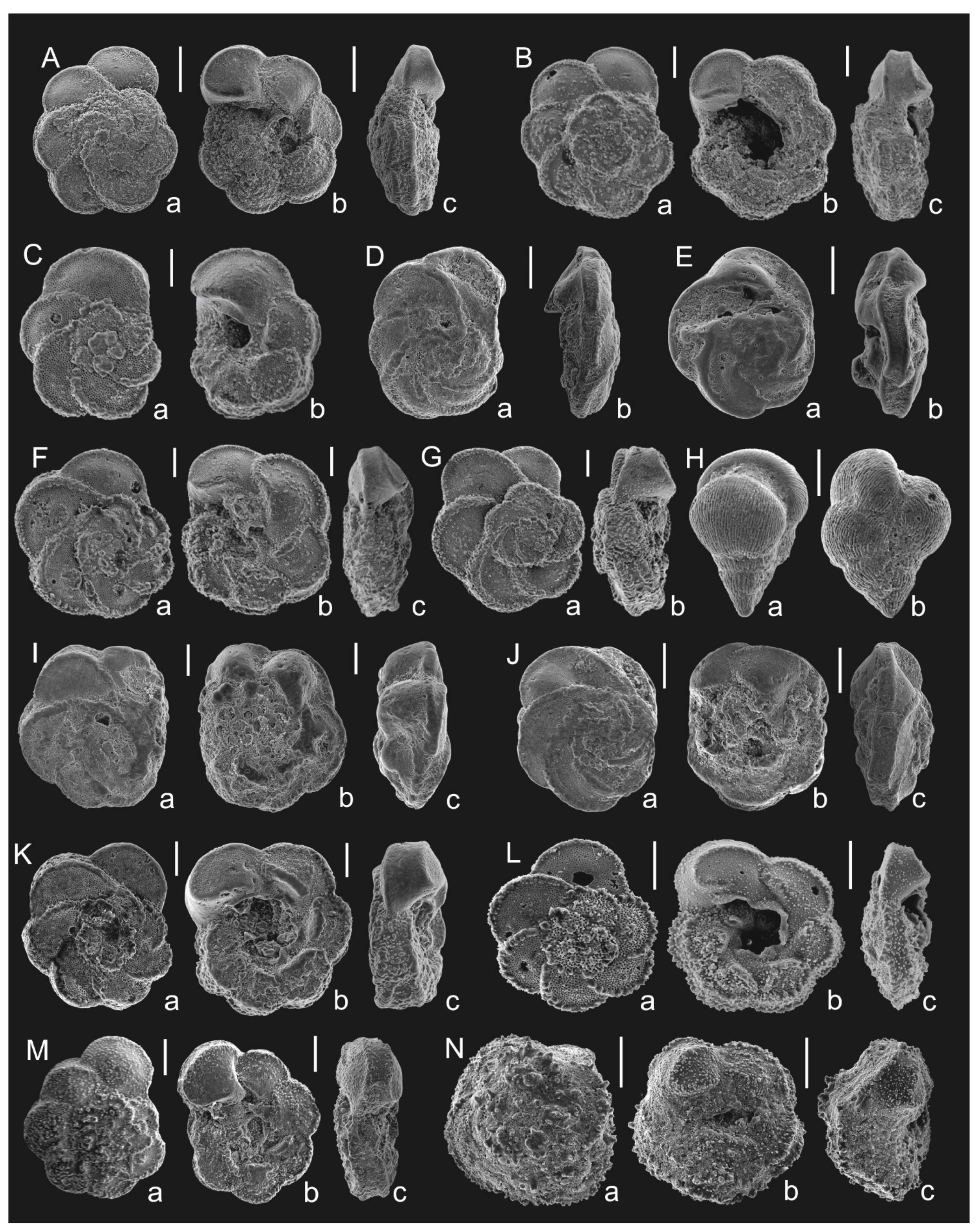
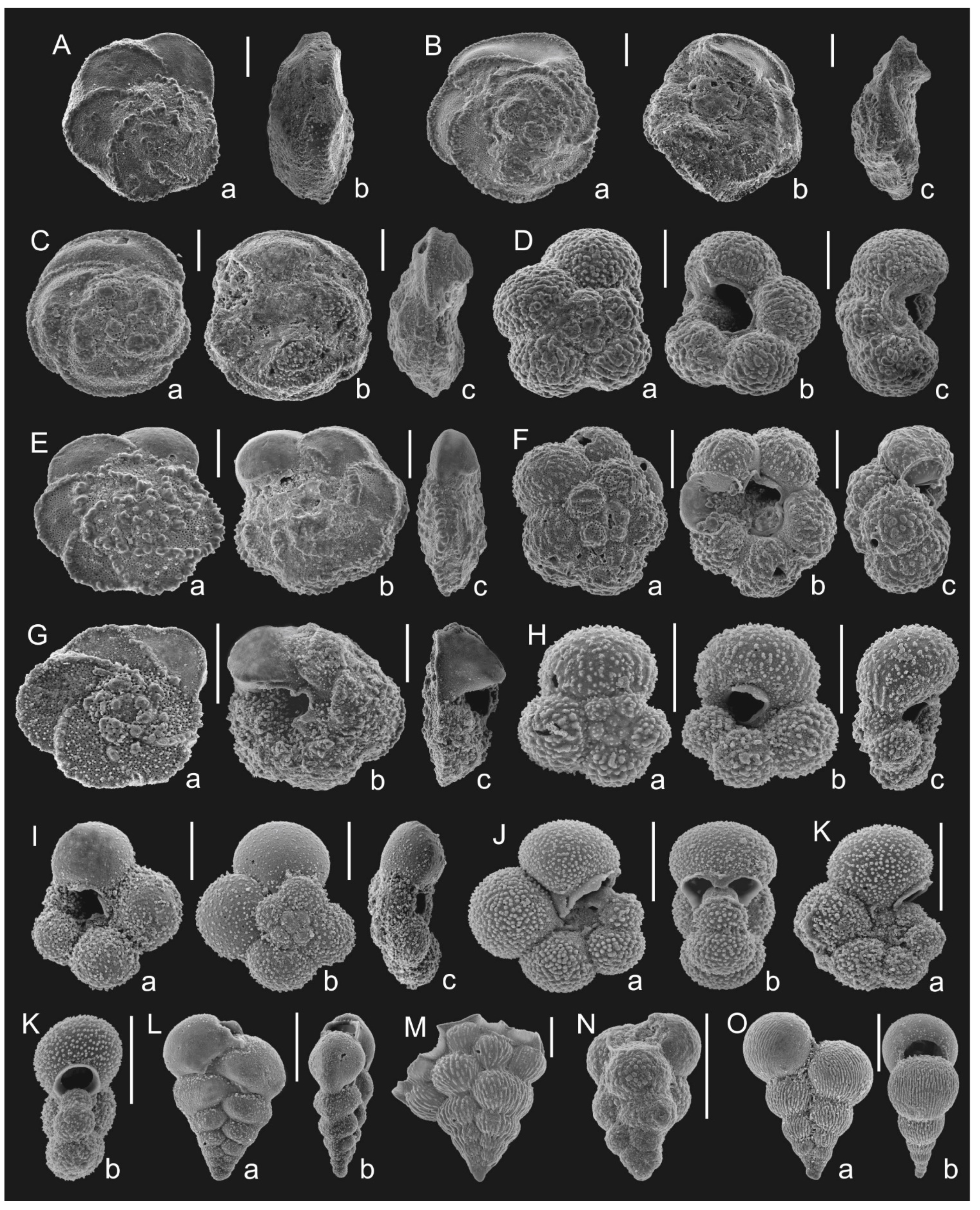
Figure 4. Correlation of the proposed zonation with the standard planktonic foraminiferal zonations [3,4] based on Tethyan species; (A)— stages and ages after [48]; (B)— planktonic foraminiferal zonation (this paper); (C,D)—standard planktonic foraminiferal zonations: (C)—after [3]; (D)—after [4]. Genera abbreviations: Th.—Thalmanninella; Marg.—Marginotruncana; Glob.—Globotruncana; Gl-nella—Globotruncanella; Ps-guemb.—Pseudoguembelina; Pseudotext.—Pseudotextularia; Plum.—Plummerita.Figure 4. Correlation of the proposed zonation with the standard planktonic foraminiferal zonations [3,4] based on Tethyan species; (A)— stages and ages after [48]; (B)— planktonic foraminiferal zonation (this paper); (C,D)—standard planktonic foraminiferal zonations: (C)—after [3]; (D)—after [4]. Genera abbreviations: Th.—Thalmanninella; Marg.—Marginotruncana; Glob.—Globotruncana; Gl-nella—Globotruncanella; Ps-guemb.—Pseudoguembelina; Pseudotext.—Pseudotextularia; Plum.—Plummerita.![Geosciences 12 00022 g004]() Figure 5. Planktonic foraminifera from the Puławy IG-2 (A,B) and S-19 (C–E,H,I,M) boreholes, Dubivtsi 1 quarry (F,J,K,N) and Słupia Nadbrzeżna outcrop (G,L); A(a–c). Thalmanninella globotruncanoides, depth 855.8 m; B(a–c). Thalmanninella reicheli, depth 850.6 m; C(a–c). Rotalipora cushmani, depth 460.5 m; D(a,b). Thalmanninella greenhornensis; depth 460.5 m; E(a,b). Whiteinella archaeocretacea, depth 458.8 m; F(a–c). Dicarinella canaliculata; G(a,b). Dicarinella hagni; H(a–c). Dicarinella imbricata, depth 458.2 m; I(a,b). Dicarinella hagni, depth 457.4 m; J(a,b). Helvetoglobotuncana helvetica; K(a–c). Falsotruncana sp.; L(a,b). Dicarinella concavata; M(a,b). Helvetoglobotuncana helvetica, depth 444.4 m; N(a–c). Whiteinella brittonensis. Scale bar = 100 µm.Figure 5. Planktonic foraminifera from the Puławy IG-2 (A,B) and S-19 (C–E,H,I,M) boreholes, Dubivtsi 1 quarry (F,J,K,N) and Słupia Nadbrzeżna outcrop (G,L); A(a–c). Thalmanninella globotruncanoides, depth 855.8 m; B(a–c). Thalmanninella reicheli, depth 850.6 m; C(a–c). Rotalipora cushmani, depth 460.5 m; D(a,b). Thalmanninella greenhornensis; depth 460.5 m; E(a,b). Whiteinella archaeocretacea, depth 458.8 m; F(a–c). Dicarinella canaliculata; G(a,b). Dicarinella hagni; H(a–c). Dicarinella imbricata, depth 458.2 m; I(a,b). Dicarinella hagni, depth 457.4 m; J(a,b). Helvetoglobotuncana helvetica; K(a–c). Falsotruncana sp.; L(a,b). Dicarinella concavata; M(a,b). Helvetoglobotuncana helvetica, depth 444.4 m; N(a–c). Whiteinella brittonensis. Scale bar = 100 µm.
Figure 5. Planktonic foraminifera from the Puławy IG-2 (A,B) and S-19 (C–E,H,I,M) boreholes, Dubivtsi 1 quarry (F,J,K,N) and Słupia Nadbrzeżna outcrop (G,L); A(a–c). Thalmanninella globotruncanoides, depth 855.8 m; B(a–c). Thalmanninella reicheli, depth 850.6 m; C(a–c). Rotalipora cushmani, depth 460.5 m; D(a,b). Thalmanninella greenhornensis; depth 460.5 m; E(a,b). Whiteinella archaeocretacea, depth 458.8 m; F(a–c). Dicarinella canaliculata; G(a,b). Dicarinella hagni; H(a–c). Dicarinella imbricata, depth 458.2 m; I(a,b). Dicarinella hagni, depth 457.4 m; J(a,b). Helvetoglobotuncana helvetica; K(a–c). Falsotruncana sp.; L(a,b). Dicarinella concavata; M(a,b). Helvetoglobotuncana helvetica, depth 444.4 m; N(a–c). Whiteinella brittonensis. Scale bar = 100 µm.Figure 5. Planktonic foraminifera from the Puławy IG-2 (A,B) and S-19 (C–E,H,I,M) boreholes, Dubivtsi 1 quarry (F,J,K,N) and Słupia Nadbrzeżna outcrop (G,L); A(a–c). Thalmanninella globotruncanoides, depth 855.8 m; B(a–c). Thalmanninella reicheli, depth 850.6 m; C(a–c). Rotalipora cushmani, depth 460.5 m; D(a,b). Thalmanninella greenhornensis; depth 460.5 m; E(a,b). Whiteinella archaeocretacea, depth 458.8 m; F(a–c). Dicarinella canaliculata; G(a,b). Dicarinella hagni; H(a–c). Dicarinella imbricata, depth 458.2 m; I(a,b). Dicarinella hagni, depth 457.4 m; J(a,b). Helvetoglobotuncana helvetica; K(a–c). Falsotruncana sp.; L(a,b). Dicarinella concavata; M(a,b). Helvetoglobotuncana helvetica, depth 444.4 m; N(a–c). Whiteinella brittonensis. Scale bar = 100 µm.![Geosciences 12 00022 g005]() Figure 6. Planktonic foraminifera from the Dubivtsi 1 quarry (A), Słupia Nadbrzeżna outcrop (B–G), Ciszyca Górna (H), Dubivtsi 2 quarry (I–K,M) and Polanówka UW-1 borehole (L,N); A(a–c). Marginotruncana coronata; B(a–c). Marginotruncana marginata; C(a,b). Marginotruncana caronae; D(a,b), E(a,b). Marginotruncana sinuosa; F(a–c). Marginotruncana renzi; G(a,b). Marginotruncana pseudolinneiana; H(a,b). Pseudotextularia nuttali; I(a–c). Marginotruncana paraconcavata; J(a–c). Marginotruncana undulata; K(a–c). Globotruncana linneiana; L(a–c). Globotruncana arca; M(a–c). Globotruncana bulloides; N(a–c). Contusotruncana patelliformis. Scale bar = 100 µm.Figure 6. Planktonic foraminifera from the Dubivtsi 1 quarry (A), Słupia Nadbrzeżna outcrop (B–G), Ciszyca Górna (H), Dubivtsi 2 quarry (I–K,M) and Polanówka UW-1 borehole (L,N); A(a–c). Marginotruncana coronata; B(a–c). Marginotruncana marginata; C(a,b). Marginotruncana caronae; D(a,b), E(a,b). Marginotruncana sinuosa; F(a–c). Marginotruncana renzi; G(a,b). Marginotruncana pseudolinneiana; H(a,b). Pseudotextularia nuttali; I(a–c). Marginotruncana paraconcavata; J(a–c). Marginotruncana undulata; K(a–c). Globotruncana linneiana; L(a–c). Globotruncana arca; M(a–c). Globotruncana bulloides; N(a–c). Contusotruncana patelliformis. Scale bar = 100 µm.
Figure 6. Planktonic foraminifera from the Dubivtsi 1 quarry (A), Słupia Nadbrzeżna outcrop (B–G), Ciszyca Górna (H), Dubivtsi 2 quarry (I–K,M) and Polanówka UW-1 borehole (L,N); A(a–c). Marginotruncana coronata; B(a–c). Marginotruncana marginata; C(a,b). Marginotruncana caronae; D(a,b), E(a,b). Marginotruncana sinuosa; F(a–c). Marginotruncana renzi; G(a,b). Marginotruncana pseudolinneiana; H(a,b). Pseudotextularia nuttali; I(a–c). Marginotruncana paraconcavata; J(a–c). Marginotruncana undulata; K(a–c). Globotruncana linneiana; L(a–c). Globotruncana arca; M(a–c). Globotruncana bulloides; N(a–c). Contusotruncana patelliformis. Scale bar = 100 µm.Figure 6. Planktonic foraminifera from the Dubivtsi 1 quarry (A), Słupia Nadbrzeżna outcrop (B–G), Ciszyca Górna (H), Dubivtsi 2 quarry (I–K,M) and Polanówka UW-1 borehole (L,N); A(a–c). Marginotruncana coronata; B(a–c). Marginotruncana marginata; C(a,b). Marginotruncana caronae; D(a,b), E(a,b). Marginotruncana sinuosa; F(a–c). Marginotruncana renzi; G(a,b). Marginotruncana pseudolinneiana; H(a,b). Pseudotextularia nuttali; I(a–c). Marginotruncana paraconcavata; J(a–c). Marginotruncana undulata; K(a–c). Globotruncana linneiana; L(a–c). Globotruncana arca; M(a–c). Globotruncana bulloides; N(a–c). Contusotruncana patelliformis. Scale bar = 100 µm.![Geosciences 12 00022 g006]() Figure 7. Planktonic foraminifera from the Jeżówka (A), Ciszyca Górna (B,C), Kłudzie (D,F), and Sulejów (E) outcrops, Polanówka UW-1 borehole (G,H,J–M), Chełm quarry (I,O) and Nasiłów outcrop (N); A(a,b). Globotruncana ventricosa; B(a–c). Contusotruncana fornicata; C(a–c). Contusotruncana plummerae; D(a–c). Rugoglobigerina rugosa; E(a–c). Globotruncana rugosa; F(a–c). Rugoglobigerina pennyi; G(a–c). Globotruncanita pettersi, depth 90.3 m; H(a–c). Globotruncanella minuta, depth 79.6 m; I(a–c). Globotruncanella petaloidea; J(a,b). Globigerinelloides multispinus, depth 74.85 m; K(a,b). Globigerinelloides prairiehillensis, depth 86.4 m; L(a,b). Laeviheterohelic glabrans, depth 74.9 m; M. Planoglobulina brazoensis, depth 81.1 m; N. Guembelitria cretacea; O(a,b). Planoheterohelix striata. Scale bar = 100 µm.Figure 7. Planktonic foraminifera from the Jeżówka (A), Ciszyca Górna (B,C), Kłudzie (D,F), and Sulejów (E) outcrops, Polanówka UW-1 borehole (G,H,J–M), Chełm quarry (I,O) and Nasiłów outcrop (N); A(a,b). Globotruncana ventricosa; B(a–c). Contusotruncana fornicata; C(a–c). Contusotruncana plummerae; D(a–c). Rugoglobigerina rugosa; E(a–c). Globotruncana rugosa; F(a–c). Rugoglobigerina pennyi; G(a–c). Globotruncanita pettersi, depth 90.3 m; H(a–c). Globotruncanella minuta, depth 79.6 m; I(a–c). Globotruncanella petaloidea; J(a,b). Globigerinelloides multispinus, depth 74.85 m; K(a,b). Globigerinelloides prairiehillensis, depth 86.4 m; L(a,b). Laeviheterohelic glabrans, depth 74.9 m; M. Planoglobulina brazoensis, depth 81.1 m; N. Guembelitria cretacea; O(a,b). Planoheterohelix striata. Scale bar = 100 µm.
Figure 7. Planktonic foraminifera from the Jeżówka (A), Ciszyca Górna (B,C), Kłudzie (D,F), and Sulejów (E) outcrops, Polanówka UW-1 borehole (G,H,J–M), Chełm quarry (I,O) and Nasiłów outcrop (N); A(a,b). Globotruncana ventricosa; B(a–c). Contusotruncana fornicata; C(a–c). Contusotruncana plummerae; D(a–c). Rugoglobigerina rugosa; E(a–c). Globotruncana rugosa; F(a–c). Rugoglobigerina pennyi; G(a–c). Globotruncanita pettersi, depth 90.3 m; H(a–c). Globotruncanella minuta, depth 79.6 m; I(a–c). Globotruncanella petaloidea; J(a,b). Globigerinelloides multispinus, depth 74.85 m; K(a,b). Globigerinelloides prairiehillensis, depth 86.4 m; L(a,b). Laeviheterohelic glabrans, depth 74.9 m; M. Planoglobulina brazoensis, depth 81.1 m; N. Guembelitria cretacea; O(a,b). Planoheterohelix striata. Scale bar = 100 µm.Figure 7. Planktonic foraminifera from the Jeżówka (A), Ciszyca Górna (B,C), Kłudzie (D,F), and Sulejów (E) outcrops, Polanówka UW-1 borehole (G,H,J–M), Chełm quarry (I,O) and Nasiłów outcrop (N); A(a,b). Globotruncana ventricosa; B(a–c). Contusotruncana fornicata; C(a–c). Contusotruncana plummerae; D(a–c). Rugoglobigerina rugosa; E(a–c). Globotruncana rugosa; F(a–c). Rugoglobigerina pennyi; G(a–c). Globotruncanita pettersi, depth 90.3 m; H(a–c). Globotruncanella minuta, depth 79.6 m; I(a–c). Globotruncanella petaloidea; J(a,b). Globigerinelloides multispinus, depth 74.85 m; K(a,b). Globigerinelloides prairiehillensis, depth 86.4 m; L(a,b). Laeviheterohelic glabrans, depth 74.9 m; M. Planoglobulina brazoensis, depth 81.1 m; N. Guembelitria cretacea; O(a,b). Planoheterohelix striata. Scale bar = 100 µm.![Geosciences 12 00022 g007]()





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
- Remarks: Planktonic foraminiferal assemblages yielded mostly shallow-water foraminifera of simple morphology, e.g., hedbergellids (Muricohedbergella planispira, M. delrioensis and Clavihedbergella simplex); heterohelicids (Planoheterohelix moremani), guembelitriids (Guembelitria cenomana) and globigerinelloidids (Globigerinelloides bentonensis, G. ultramicrus) were common. Rare occurrences of Praeglobotruncana delrioensis and rotaliporids (Thalmanninella appenninica, Th. Tehamaensis and Th. gandolfii) were also recorded.
- Stratigraphic distribution: Annopol outcrop, Ożarów quarry.
- Thalmanninella globotruncanoides Interval Zone
- Author: Robaszynski and Caron [3].
- Definition: The body of strata from the lowest occurrence of Thalmanninella globotruncanoides to the lowest occurrence of Thalmanninella reicheli (Figure 3).
- Chronostratigraphy: Lower Cenomanian.
- Remarks: Planktonic foraminifera assemblages of the zone were also dominated by hedbergellids, accompanied by less common guembelitriids, globigerinelloidids and heterohelicids. New species in the zone, besides the zonal marker, were Clavihedbergella simplicissima and Praeglobotruncana stephani.
- Stratigraphic distribution: Ożarów quarry, Puławy IG-2 borehole (from a depth of 851.2 m downward).
- Thalmanninella reicheli Taxon-Range Zone
- Author: Bolli [53].
- Definition: The body of strata with the total range of the zonal marker (Figure 3).
- Chronostratigraphy: Middle Cenomanian.
- Remarks: The zonal marker was rare but was consistently present.
- Other species were the same as in the previous zone, besides Thalmannitella brotzeni, which has not previously been recorded in this zone.
- Stratigraphic distribution: Puławy IG-2 borehole (depth 849.2–851.2 m).
- Rotalipora cushmani Taxon-Range Zone
- Author: Borsetti [54].
- Definition: The body of strata containing the total range of the nominate taxon (Figure 3).
- Chronostratigraphy: Upper middle to uppermost Cenomanian.
- Remarks: The zonal marker was consistently present, but nowhere abundant. In addition to the index fossil, Praeglobotruncana stephani and Whiteinella brittonensis appeared in this zone. Globigerinelloides bentonensis had its highest occurrence. Muricohedbergella delrioensis and M. planispira were transit species, i.e., species occurring both in a given zone and in adjacent zones.
- Stratigraphic distribution: Ożarów quarry, Puławy IG-2 borehole (depth 845.3–849.2 m); S-19 borehole (depth from 459.3 m downward).
- Whiteinella archaeocretacea Partial Range Zone
- Author: Bolli [53].
- Definition: The body of strata with Whiteinella archaeocretacea from the highest occurrence of Rotalipora cushmani to the lowest occurrence of Helvetoglobotruncana helvetica (Figure 3).
- Chronostratigraphy: Uppermost Cenomanian through to lower Turonian.
- Remarks: Besides quite rare occurrences of Whiteinella archaeocretacea, there were also Dicarinella (D. imbricata, D. algeriana, D. hagni and D. longoriai) and Praeglobotruncana (P. stephani, P. oraviensis and P. gibba) abundant in the zone; common occurrences of Whiteinella (W. brittonensis, Wh. Aprica and W. baltica) and Muricohedbergella delrioensis were also observed.
- Stratigraphic distribution: Ożarów quarry, Puławy IG-2 borehole; S-19 borehole (depth 446.3–459.3 m).
- Correlation: The Whiteinella archaeocretacea zone corresponds to the lower part of the Inoceramus ex. gr. labiatus and Neocardioceras juddii zones distinguished in extra-Carpathian Poland (Figure 3).
- Helvetoglobotruncana helvetica Taxon-Range Zone
- Author: Sigal [55].
- Definition: The body of strata containing the total range of Helvetoglobotruncana helvetica (Figure 3).
- Chronostratigraphy: Middle Turonian.
- Remarks: The zonal marker was present consistently, but nowhere abundantly. Within the zone, the first marginotruncanids with a still weakly developed double keel appeared. “True” Marginotruncana with strongly beaded keels appeared in the uppermost part of the zone. Whiteinella (W. brittonensis, W. aprica and W. baltica) and Muricohedbergella delrioensis were transit species. Planoheterohelix reussi and P. globulosa were present in the assemblages. The last occurrences of Clavihedbergella simplicissima were noted within this zone.
- Stratigraphic distribution: Dubivtsi 1 quarry, Ożarów quarry, S-19 borehole (depth 436.0–446.3 m).
- Marginotruncana coronata Partial Range Zone
- Author: Peryt [11], amended here.
- Definition: The body of strata with Marginotruncana coronata from the highest occurrence of Helvetoglobotruncana helvetica to the lowest occurrence of Marginotruncana sinuosa (Figure 3).
- Chronostratigraphy: Lower upper Turonian.
- Remarks: The Marginotruncanacoronata zone was originally defined as the Marginotruncana coronate and Globotruncana lapparenti lowest occurrence interval zone representing the upper Turonian and Coniacian.
- Characteristic assemblages: In the lower part of the zone, foraminiferal assemblages were dominated by Marginotruncana: M. pseudolinneiana, M. coronata, M. marginata, M. paraconcavata, M. renzi and M. caronae; Falsotruncana maslakovae occurred sporadically. In the middle part of this zone, a decline of all keeled forms was observed, e.g., Dicarinella and Marginotruncana. Heterohelicids (Planoheterohelix reussi and P. globulosa), Globigerinelloides ultramicrus and Whiteinella spp. occurred in the entire zone, although they were very rare. In the uppermost part of the zone, keeled planktonic foraminifera reappeared.
- Stratigraphic distribution: Dubivtsi 1 quarry, Słupia Nadbrzeżna outcrop, S-19 borehole (depth from 436.0 m upward).
- Correlation: The Marginotruncana coronata zone ranges from the lowermost part of the I. costellatus, through Mytilodes scupini, up to the lower part of the Cremnoceramus waltersdorfensis zones (Figure 3). The Marginotruncana coronata zone may be correlated with the Marginotruncana schneegansi and the lowermost part of the Dicarinella concavata zones of [3] and Dicarinella primitiva–Marginotruncana sigali and the lowermost part of the D. concavata zones of [4].
- Marginotruncana sinuosa Interval Zone
- Definition: The body of strata from the lowest occurrence of Marginotruncana sinuosa to the lowest occurrence of Pseudotextularia nuttalli (Figure 3).
- Chronostratigraphy: Uppermost Turonian through lower Coniacian.
- Remarks: The Marginotruncana sinuosa zone was distinguished informally as the body of strata between the lowest occurrences of Marginotruncana sinuosa and Globotruncana linneiana [18]. Nishi et al. [56] defined the zone as a stratigraphic interval between the first occurrences of Marginotruncana sinuosa and Contusotruncana fornicata.
- Characteristic assemblages: Besides the zonal marker, Marginotruncana paraconcavata and Contusotruncana fornicata appeared at the base of the zone. Sporadic occurrences of Dicarinella concavata were recorded. Common were also Planoheterohelix and Globigerinelloides.
- Stratigraphic distribution: Dubivtsi 1 quarry, Słupia Nadbrzeżna outcrop.
- Correlation: The Marginotruncana sinuosa zone corresponds to the uppermost part of the inoceramid zones of Cremnoceramus waltersdorfensis, Cr. deformis, and the lower part of the Volviceramus involutus. The Marginotruncana sinuosa zone may be correlated with the lower part of the Dicarinella concavata zone of the Tethyan area [3,4] (Figure 4).
- Pseudotextularia nuttalli Interval Zone
- Author: Nederbragt [57], amended here.
- Definition: The body of strata from the lowest occurrence of Pseudotextularia nuttalli to the lowest occurrence of pill-box-like morphotypes of Globotruncana linneiana (Figure 3).
- Chronostratigraphy: Middle through to upper Coniacian.
- Remarks: The Pseudotextularia nuttalli zone was originally defined as an interval between the lowest occurrences of Pseudotextularia nuttalli and Sigalia carpatica. Because of the absence of Sigalia carpatica in the study area, we selected Globotruncana linneiana to define the upper boundary of the Pseudotextularia nuttalli zone.
- Characteristic assemblages: Besides Pseudotextularia nuttalli, Marginotruncana (M. pseudolinneiana, M. coronata, M. marginata, M. paraconcavata, M. renzi, M. caronae, M. sinuosa and M. undulata) and Contusotruncana (C. fornicata and C. morozovae) also occurred abundantly in the zone. In western Ukraine, the disappearance of rare Tethyan planktonic foraminifera (Dicarinella concavata, Marginotruncana paraconcavata, Marginotruncana sinuosa, M. undulata and Contusotruncana morozovae) was observed at the zonal boundary.
- Stratigraphic distribution: Dubivtsi 2 quarry; Wesołówka outcrop.
- Correlation: The Pseudotextularia nuttalli zone corresponds to the upper part of the Volviceramus involutus, Magadiceramus subquadratus and Sphaeroceramus pachti inoceramid zones. The zone may be correlated with the upper part of the Dicarinella concavata of [3] and the upper part of the Dicarinella concavata and the lower part of the Dicarinella asymetrica zones of [4].
- Globotruncana linneiana Concurrent-Range Zone
- Author: Peryt [18], amended here.
- Definition: The body of strata from the lowest occurrence of pill-box-like morphotypes of Globotruncana linneiana to the highest occurrence of Marginotruncana marginata (Figure 3).
- Chronostratigraphy: Santonian.
- Remarks: The Globotruncana linneiana zone was originally defined as an interval between the lowest occurrences of nominal taxon and Contusotruncana fornicata.
- Characteristic assemblages: In the lower and middle part of the zone, planktonic foraminiferal assemblages were dominated by a few cosmopolitan species, such as Globotruncana linneiana, G. bulloides, Marginotruncana pseudolinneiana and M. marginata, and abundant heterohelicids and hedbergellids. In the upper Santonian, the first Globotruncana arca was recorded, while, at the Santonian/Campanian boundary, Marginotruncana disappeared.
- Stratigraphic distribution: Dubivtsi 2 quarry, Bocieniec and Wesołówka outcrops.
- Correlation: The Globotruncana linneiana zone corresponds to the inoceramid zones of Cordiceramus undulatoplicatus, Sphaeroceramus pinniformis and Cordiceramus muelleri, and the lower part of the Sphaeroceramus patootensiformis. The zone may be correlated with the Dicarinella asymetrica zone established for the Mediterranean [3] and the upper part of the D. asymetrica zone distinguished in the Gubbio section [4].
- Globotruncana arca Partial Range Zone
- Author: Salaj and Samuel [58], amended here.
- Definition: The body of strata with Globotruncana arca from the highest occurrence of Marginotruncana marginata to the lowest occurrence of Contusotruncana plummerae (Figure 3).
- Chronostratigraphy: Lower Campanian.
- Remarks: The Globotruncana arca zone was originally defined as an interval zone between the lowest occurrences of Globotruncana arca and Globotruncana rugosa.
- Characteristic assemblages: In addition to the index species, Globotruncana linneiana, G. bulloides, G. rugosa and Contusotruncana fornicata dominated the assemblages. Archaeoglobigerina bosquensis, A. cretacea, Planoheterohelix globulosa, P. moremani and P. reussi were also common.
- Stratigraphic distribution: Dubivtsi 2 quarry, Bocieniec outcrop, Jeżówka outcrop, Mielnik I quarry, Sulejów outcrop.
- Correlation: The Globotruncana arca zone corresponds to the upper part of the Sphaeroceramus patootensiformis, Sphaeroceramus sarumensis-Cataceramus dariensis, Cataceramus beckumensis and, Inoceramus “azerbaydjanensis”-“Inoceramus” vorhelmensis inoceramid zones; to the Gonioteuthis granulata, G. quadrata and Neancyloceras phaleratum cephalopod zones; to the vulgaris/basiplana, stobaei/basiplana, conica/mucronata, gracilis/mucronata, conica/papillosa, papillosa, senonensis cephalopod/echinoid zones. The Globotruncana arca zone correlates with the Globotruncanita elevata zone established for the Tethyan area [3,4] (Figure 4).
- Contusotruncana plummerae Interval Zone
- Author: Petrizzo et al. [59], amended here.
- Definition: The body of strata from the lowest occurrence of Contusotruncana plummerae to the lowest occurrence of Rugoglobigerina pennyi (Figure 3).
- Chronostratigraphy: Middle through to upper Campanian.
- Remarks: The Contusotruncana plummerae zone was originally defined as the stratigraphic interval from the lowest occurrence of the nominal taxon and the lowest occurrence of Radotruncana calcarata [59]. The absence of Radotruncana calcarata, Globotruncanella havanensis, Globotruncana aegyptiaca and Gansserina gansseri in the study area caused that the top of the zone was placed at the lowest occurrence of Rugoglobigerina pennyi, just below the Campanian/Maastrichtian boundary.
- Characteristic assemblages: The planktonic foraminiferal assemblages in the lower part of the zone were well-diversified and dominated by keeled forms: Contusotruncana (C. plummerae and C. fornicata) and Globotruncana (G. arca, G. rugosa, G. mariae, G. bulloides and G. linneiana); very rare occurrences of G. ventricosa were recorded. In the upper part of the zone, globotruncanids disappeared almost completely. Low-diversity planktonic foraminiferal assemblages dominated by Planoheterohelix and Globigerinelloides characterized this interval. In the upper part of the zone, Rugoglobigerina rugosa appeared.
- Stratigraphic distribution: Mielnik II quarry, Middle Vistula River outcrops (Dorotka, Leśne Chałupy, Kolonia Ciszyca, Ciszyca Górna, Wola Pawłowska, Pawłowice Cemetary, Łopoczno, Piotrawin, Kamień, Sadkowice North, Raj and Raj North), Puławy IG-2 borehole.
- Correlation: The Contusotruncana plummerae zone corresponds to the Bostrychoceras polyplocum, Didymoceramus donezianum, Nostoceras pozaryskii (=hyatti) cephalopod zones, Belemnella lanceolata zone and to the inoceramid zones of Cataceramus subcompressus, “Inoceramus” tenuilineatus, Sphaeroceramus pertenuiformis, “Inoceramus” altus, “Inoceramus” inkermanensis and Trochoceramus costaceus. The Contusotruncana plummerae zone represents the longest stratigraphic interval of the Upper Cretaceous in extra-Carpathian Poland and western Ukraine. Its lower part correlates with the Globotruncana ventricosa [3] or Contusotruncana plummerae zones [4], its middle part with the Globotruncanita calcarata, Globotruncanella havanensis and Globotruncana aegyptiaca zones [3,4] and its upper part may be correlated with the lower part of the Gansserina gansseri zone of [3] and [4] (Figure 4).
- Rugoglobigerina pennyi Interval Zone
- Author: Peryt [11].
- Definition: The body of strata from the lowest occurrence of Rugoglobigerina pennyi to the lowest occurrence of Globotruncanella petaloidea (Figure 3).
- Chronostratigraphy: Lower Maastrichtian.
- Remarks: The planktonic foraminiferal assemblages of the zone were dominated by Rugoglobigerina. At the base, in addition to the index species, Rugoglobigerina milamensis, R. rugosa and R. hexacamerata dominated assemblages. Planoheterohelix and Globigerinelloides were also common components of the assemblages. Keeled forms were almost completely absent.
- Stratigraphic distribution: Middle Vistula River outcrops (Kłudzie, Dziurków and Boiska), Wrocieryż outcrop, Mielnik III quarry, Kamyanopil outcrop, Puławy IG-2 borehole.
- Correlation: The Rugoglobigerina pennyi zone corresponds to the Belemnella occidentalis Zone and to the inoceramid zones of “Inoceramus” redbirdensis, Endocostea typica and Trochoceramus radiosus, and the ammonite zones of Pachydiscus neubergicus and Acantoscaphites tridens. The zone may be correlated with the middle part of the Gansserina gansseri zone of [3] and the upper part of the G. gansseri zone distinguished by [4] (Figure 4).
- Globotruncanella petaloidea Assemblage Zone
- Author: Peryt et al. (this paper).
- Definition: The body of strata containing Globotruncanella petaloidea, Globotruncana linneiana, Contusotruncana plummerae and Planoglobulina brazoensis (Figure 3).
- Chronostratigraphy: Middle Maastrichtian.
- Remarks: Abundant and well-diversified planktonic foraminiferal assemblages, dominated by keeled planktonic foraminifera (Globotruncana arca, G. linneiana, G. bulloides, Contusotruncana plummerae, C. fornicata and C. patelliformis), were recorded in the zone. They reappeared after their absence in the upper part of the Contusotruncana plummerae zone and the entire Rugoglobigerina pennyi zone. At the same level, also large, biserial and multiserial heterohelicids (Pseudotextularia deformis, Planoglobulina brazoensis and Racemiguembelina powelli) as well as triserial guembelitriids (Guembelitria cretacea) appeared, although in small numbers. Globotruncanella petaloidea, Planoheterohelix and Globigerinelloides were also common in the assemblages. On the other hand, Rugoglobigerina disappeared in the uppermost part of the zone.
- Stratigraphic distribution: Middle Vistula River outcrops (Jarentowskie Pole and Chotcza), Polanówka UW-1 borehole, Puławy IG-2 borehole, Chełm quarry.
- Correlation: The Globotruncanella petaloidea zone corresponds to the lower part of the Belemnitella junior belemnite zone, to the lower part of the “Inoceramus” ianjonaensis inoceramid zone, to the uppermost part of the Acanthoscaphites tridens and to the lower part of the Hoploscaphites constrictus lvivensis ammonite zones (Figure 3), and may be correlated with the upper part of the Gansserina gansseri and entire Contusotruncana contusa planktonic foraminiferal zones of the Mediterranean sections [3,4] (Figure 4).
- Guembelitria cretacea Concurrent-Range Zone
- Author: Peryt [11], amended here.
- Definition: The body of strata with Guembelitria cretacea between the highest occurrence of Contusotruncana plummerae and the highest occurrence of Globigerinelloides multispinus (Figure 3).
- Chronostratigraphy: Upper Maastrichtian.
- Remarks: Peryt [11] defined the Guembelitria cretacea zone as a total range of nominal species.
- Characteristic assemblages: The planktonic foraminiferal assemblages from the zone were dominated by heterohelicids (Planoheterohelixglobulosa, P. navarroensis, P. striata and P. vistulaensis) and Guembelitria cretacea; rare occurrences of Globigerinelloides (G. multispinus and G. prairiehillensis) and Archaeoglobigerina were also reported. A short-lived reappearance of Rugoglobina was recorded in the Belemnella kazimiroviensis zone, followed by an increase in the abundance of Guembelitria cretacea just before the Cretaceous/Paleogene boundary.
- Stratigraphic distribution: Middle Vistula River sections (Lucimia, Podgórz, Dobre, Kazimierz Dolny and Nasiłów), Chełm quarry, Lechówka outcrop, Mełgiew outcrop, Puławy IG-2 borehole.
- Correlation: The Guembelitria cretacea zone corresponds to the upper part of the Belemnitella junior and to Belemnella kazimiroviensis belemnite zones, to the upper part of the “Inoceramus” ianjonaensis and Tenuipteria argentea inoceramid zones and of the upper part of the Hoploscaphites constrictus lvivensis, H. constrictus crassus and H. constrictus johnjagti ammonite zones. The zone may be correlated with the almost entire Abathomphalus mayaroensis zone (without its lowermost part) of [3] and with the Abathomphalus mayaroensis zone (without its lowermost part), Pseudoguembelina hariaensis, Pseudotextularia elegans and Plummerita hantkeninnoides planktonic foraminiferal zones of [4] (Figure 4).
5. Discussion
The result of the mid-Cretaceous transgression and climate maximum in late Cenomanian to early Turonian times was manifested, among others, by the occurrence of wide epicontinental seas with tropical sea surface temperatures (SSTs) (possibly greater than 35 °C) [60]. These newly appeared ecological niches were inhabited by not only warm-water planktonic foraminifera, but also quite frequent deep-dwelling forms. Consequently, foraminiferal zones that have been established in Tethyan successions in the Cenomanian and lower Turonian could be recognized at higher latitudes, e.g., in the epicontinental Central Polish Basin. In the middle Turonian, when the Equatorial Atlantic Gateway was opened enough to inundate the deep North Atlantic with relatively cool polar waters of the Southern Ocean, the hot greenhouse climate ended. Then, a significant global cooling continued until the Maastrichtian, possibly additionally driven by declining pCO2 levels [61]. The Late Cretaceous climate cooling caused a narrowing of the latitudinal ranges of warm tropical/subtropical waters and a simultaneous decrease in the latitudinal extent of warm-water planktonic foraminifera ranges. In consequence, the Foraminiferal Transitional Bioprovince was characterized by the absence or very rare occurrences of warm-water species, which are usually the index taxa of standard foraminiferal zonations, e.g., marginotruncanids with very closely spaced keels (M. sigali—M. schneegansi group), umbiliconvex dicarinellids (Dicarinella primitiva–D. concavata–D. asymetrica lineage), Falsotruncana, Globotruncanita, Radotruncana, Gansserina, Abathomphalus, complex heterohelicids and Plummerita. Thus, the zonation we proposed for the upper Turonian through Maastrichtian was based mainly on the species of a wider geographical distribution, but usually also of longer stratigraphical ranges, e.g., double-keeled Marginotruncana, Globotruncana, Contusotruncana, Rugoglobigerina, Globotruncanella and heterohelicids (Pseudotextularia nuttalli) and Guembelitria cretacea (Figure 4).
The Late Cretaceous transgression encroached the area of the Polish Lowlands in the late Albian [62,63]. The first transgressive, shallow-water, siliciclastic deposits contained mostly shallow-water foraminifera of simple morphology and a rather insignificant biostratigraphic utility, e.g., low-trochospiral hedbergellids (Muricohedbergella planispira, M. delrioensis and Clavihedbergella simplex) and heterohelicids (Planoheterohelix moremani), guembelitriids (Guembelitria cenomana) and globigerinelloidids (Globigerinelloides ultramicrus) [13,14,33,64]. Deeper-dwelling foraminifera, such as Praeglobotruncana (P. delrioensis), occurred in these sediments sporadically. Along with the advancing transgression of deep-dwelling keeled thalmanninellids (Thalmanninella spp. and Rotalipora spp.) and the appearance of high-trochospiral hedbergellids (Praeglobotruncana stephani and P. gibba), a basis for the recognition of standard zones such as the Thalmanninella appenninica, Th. globotruncanoides, Th. reicheli and Rotalipora cushmani was established. Thalmanninella globotruncanoides was the boundary criterion for the GSSP of the Cenomanian stage with the Mont Risou stratotype section (southeast France) [65,66]. All of the standard zones, i.e., Th. globotruncanoides, Th. reicheli and R. cushmani, were present in the Puławy IG-2 borehole [17] (Figure 3 and Figure 4). In contrast, in the Annopol anticline, where the Cenomanian succession was strongly condensed (2 m thick) with stratigraphical gaps and mixing of microfauna by burrowers, only some of the foraminiferal zones were recognizable [33].
The GSSP for the base of the Turonian Stage was at the Rock Canyon Anticline, near Pueblo (Colorado, United States), and occurred within the Whiteinella archaeocretacea zone [48,67], which was followed by the Helvetogloglobotruncana helvetica zone. In the study area, both zones were recognized. In addition, this stratigraphic interval recorded some other events with a high stratigraphic potential. In the lower part of the W. archaeocretacea zone, the lowest occurrences of Dicarinella (D. imbricata, D. algeriana, D. longoriai and D. hagni) were recognized, while, in the middle part of the middle Turonian, i.e., within the Helvetogloglobotruncana helvetica zone, the first marginotruncananids with the still weakly developed keel structure appeared. “True” Marginotruncana with strongly beaded keels appeared in the middle–upper Turonian boundary interval, as elsewhere worldwide [68,69,70]. The Marginotruncana assemblage was dominated by double-keeled forms: M. pseudolinneiana, M. coronata and M. marginata—cosmopolitan species, known also from the Boreal Province (e.g., [71,72]). This deep-dwelling foraminiferal community was impoverished in single-keeled, warm-water species (M. schneegansi and M. sigali) which were mostly index taxa for the upper Turonian standard zonation. Another upper Turonian stratigraphically important species, but sporadically recorded in extra-Carpathian Poland, is Falsotruncana maslakovae, the index taxon for the standard F. maslakovae zone [70,73]. In consequence, the local Marginotruncana coronata zone was proposed for almost the entire upper Turonian in this region [11,18,24]. In the middle part of this zone (middle–upper Turonian) a decline of all keeled forms was observed [24], e.g., Dicarinella and Marginotruncana, which were believed to have been the deepest-dwelling planktonic foraminifera [2,5,74]. It was probably related to a considerable sea level fall that was not a local phenomenon and, apparently, corresponded to the late Turonian trough, well recognized in Western [75] and Eastern Europe [24], as well as to the two late Turonian second-order regressive peaks documented from the Western Interior Basin [76]. This event may be of correlation importance in the entire European Basin. The reappearance of the keeled forms, mainly Marginotruncana, in the study area was noted in the higher part of the upper Turonian (Mytiloides scupini zone) [22,24]. There were also sporadic occurrences of Dicarinella concavata, a species which is an index taxon of the Dicarinella concavata zone, commonly used in the standard zonal schemes [2,3]. The Turonian/Coniacian boundary was placed in the lower part of the Dicarinella concavata zone [46,77,78]. The GSSP for the Turonian stage was at Salzgitter-Salder (Germany), defined by the first occurrence of the inoceramid bivalve Cremnoceramis deformis erectus. One of the auxiliary sections was at Słupia Nadbrzeżna (Middle Vistula River Valley succession, central Poland) [77]. Because of the rarity of Dicarinella concavata in the extra-Carpathian area, this zone was not distinguished. However, the lowest occurrence of Marginotruncana sinuosa, a marker of the following zone in the proposed zonation [18,22,24], was recorded in the uppermost Turonian (C. waltersdorfensis waltersdorfensis zone) in the study area. Foraminiferal assemblages of the Marginotruncana sinuosa zone were characterized by the dominance of Marginotruncana, Planoheterohelix and Globigerinelloides [24,27].
The next important event recorded was the lowest occurrence of Pseudotextularia nuttalli placed in the middle Coniacian [24,27]. The event is recognized widely from tropical/subtropical to temperate latitudes [27,57,79,80,81,82,83]. However, some authors recorded the lowest occurrence of P. nuttalli slightly higher, namely, from the upper Coniacian calcareous nannoplankton zone UC10 [48]. The Coniacian/Santonian boundary was defined by the first occurrence of the inoceramid bivalve Platyceramus undulatoplicatus and the GSSP for the Santonian was at Olazagutía, Spain [73]. The first occurrence of the planktonic foraminifera Sigalia was accepted as a secondary marker for the basal Santonian [80]. However, this species was common in the Mediterranean region of the Tethys and was very rare or completely absent at higher latitudes. Therefore, we proposed a pill-box-like morphotype of Globotruncana linneiana as a marker of the proximity of the basal Santonian in extra-Carpathian Poland and western Ukraine, which showed a wider geographical distribution than S. carpatica, and appeared in the uppermost Coniacian, just below the first occurrence of P. undulatoplicatus in both the Olazagutía section and at higher latitudes. In the study area, the Coniacian/Santonian boundary was additionally marked by the disappearance of foraminifera possessing keels divided by a relatively narrow (Dicarinella concavata and Marginotruncana paraconcavata) and wavy peripheral band (Marginotruncana sinuosa, M. undulata and Contusotruncana morozovae) [9], which was interpreted as being related to local tectonics connected with the early stage of the Subhercynian phase of the Alpine orogeny [9,46,84,85]. In consequence, in the lower and middle Santonian in the region under discussion (Dubivtsi section), the planktonic foraminiferal assemblages were dominated by a few cosmopolitan species, such as Globotruncana linneiana, G. bulloides, Marginotruncana pseudolinneiana and M. marginata, and abundant heterohelicids and hedbergellids.
There were two events recorded in the upper Santonian: (1) the lowest occurrence of Globotruncana arca and (2) the disappearance of Marginotruncana at the Santonian/Campanian boundary [18,27,38]. The disappearance of Marginotruncana marginata indicated the lower boundary for the proposed Globotruncana arca partial-range zone. Because this event was recorded at the Santonian/Campanian boundary, worldwide it seemed to be a very good marker for the correlation between the Tethyan and the Transitional provinces. The GSSP for the base of the Campanian has not yet been established; however, the boundary was generally correlated with the extinction of planktonic foraminifera species Dicarinella asymetrica [48,86]. In the Bocieniec section (central southern Poland), which was proposed as a possible candidate for the GSSP for the base of the Santonian Stage [38], as well as in the entire extra-Carpathian Poland and western Ukraine, D. asymetrica was very rare and, therefore, was not used as a zonal marker.
The next foraminiferal events recognised in the study area were the LO’s of Contusotruncana plummerae and Globotruncana ventricosa in the middle Campanian [12,17,19]. Globotruncana ventricosa used as the zonal marker in standard zonations [87,88] occurred only sporadically in the study area [11,12,19,89], while C. plummerae was common. We proposed this species as a zonal marker in the upper Campanian of extra-Carpathian Poland and western Ukraine.
The planktonic foraminiferal assemblages in the middle Campanian were well-diversified and dominated by keeled forms: Contusotruncana (C. plummerae and C. fornicata) and Globotruncana (G. arca, G. rugosa, G. bulloides and G. linneiana). This assemblage indicated the existence of a quite high sea-level and open connections between the studied basin and the Tethys area and, apparently, correlated with the global Campanian transgressive peak well recognized in western Europe within the Belemnitella langei zone [90].
Upwards of the section, in the upper Campanian, globotruncanids disappeared almost completely. Low-diversity planktonic foraminiferal assemblages dominated by Planoheterohelix and Globigerinelloides characterized this interval. This taxonomic impoverishment probably reflected the latest Campanian–earliest Maastrichtian eustatic sea level fall [25,91]. In the upper Campanian, Rugoglobigerina rugosa appeared, and just below the Campanian/Maastrichtian boundary in the Middle Vistula River Valley succession, an abundant and taxonomically more diverse occurrence of Rugoglobigerina began [11,16,21,25]. The lowest occurrence of Rugoglobigerina pennyi marked the lower boundary of the Rugoglobigerina pennyi zone [11]. The predominance of Rugoglobigerina in the planktonic foraminiferal assemblages was recorded at high-latitude sections: Poland, Ukraine [19,25] and Russia [92,93]. In the GSSP for the Maastrichtian stage fixed at Level 115.2 on Platform IV of the geological site at Tercis les Bains (Landes, France) [94], the Rugoglobigerina assemblage common at high-latitude sections (R. rugosa, R. hexacamerata and R. pennyi) appeared below the Campanian/Maastrichtian boundary [95]; the same was also recorded in northern Spain [96]. Rugoglobigerina scotti, which correlates with the base of the Maastrichtian at Tercis les Baines, was a Tethyan species absent at higher latitudes.
Two important events were identified at the lower/upper Maastrichtian boundary: the disappearance of Rugoglobigerina and the reappearance of keeled planktonic foraminifera (Globotruncana arca, G. linneiana, G. bulloides, Contusotruncana plummerae, C. fornicata and C. patelliformis); at the same level, also large, biserial and multiserial heterohelicids (Pseudotextularia deformis, Planoglobulina brazoensis and Racemiguembelina powelli) appeared for a short time, although in small numbers [11,16,26]. Globotruncanella petaloidea, Planoheterohelix and Globigerinelloides were also recorded in the lower–upper Maastrichtian assemblages of the region. Those changes in planktonic foraminiferal assemblages were interpreted as being due to a significant sea level rise corresponding to the transgressive peak No. 4 of Hancock [90], recognized in north-western Europe [26]. This part of the succession represented the proposed Globotruncanella petaloidea zone. The planktonic foraminiferal assemblages from the upper Maastrichtian were dominated by Planoheterohelix and Guembelitria; rare occurrences of Globigerinelloides were also reported. A short-lived reappearance of Rugoglobigerina was recorded in the Belemnella kazimiroviensis zone, followed by an increase in the abundance of Guembelitria cretacea just before the Cretaceous/Paleogene boundary [11,16]. The uppermost part of the succession was included in the Guembelitria cretacea zone.
6. Concluding Remarks
We studied the planktonic foraminiferal assemblages from the uppermost Albian through to the Maastrichtian, in macrofaunal well-dated strata cropping out in a series of small to large natural exposures, several quarries and three boreholes, located in central, eastern and south-eastern Poland and western Ukraine, aiming to update and refine foraminiferal zonations proposed for extra-Carpathian Poland.
During the Late Cretaceous, the study area was located in the central part of the European epicontinental sea and belonged to the North Transitional Foraminiferal Bioprovince.
The absence or sporadic occurrences of Tethyan taxa used as zonal markers in the standard planktonic foraminiferal zonations helped us develop a local zonation which, when calibrated by macrofossil zonations, could be applied for regional or even inter-regional correlations.
We distinguished 15 planktonic foraminiferal zones from the upper Albian up to the uppermost Maastrichtian. In the interval from the upper Albian through to the middle Turonian, the zones were the same as in the standard zonations; they were based on warm-water deep-dwelling species of planktonic foraminifera, i.e., Thalmanninella appenninica, Th. globotruncanoides, Th. reicheli, Rotalipora cushmani, Whiteinella archaeocretacea and Helvetoglobotruncana helvetica. The biozonation of the upper Turonian through to the Maastrichtian, based on more cosmopolitan planktonic foraminifera, comprised the following zones: Marginotruncana coronata, M. sinuosa, Pseudotextularia nuttalli, Globotruncana linneiana, G. arca, Contusotruncana plummerae, Rugoglobigerina pennyi, Globotruncanella petaloidea and Guembelitria cretacea.
The proposed biozonation was calibrated by macrofaunal biozonations developed in the study area and correlated with the standard planktonic foraminiferal zonations.
Author Contributions
Conceptualization, D.P. and Z.D.; methodology, D.P., Z.D. and W.W.; formal analysis, D.P., Z.D. and W.W.; investigation, D.P., Z.D. and W.W.; writing—original draft preparation, D.P. and Z.D.; writing—review and editing, D.P. and Z.D. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Polish Geological Institute—National Research Institute, grant no. 62.9012.2027.00.0 to W.W., and grant no. 2017/27/B/ST10/00687 of the National Science Centre, Poland, to Z.D.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We would like to thank the three anonymous reviewers for their valuable comments, which have contributed substantially to the improvement of this work. We thank K. Leszczyński for improving the English text.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A
Faunal reference list of planktonic foraminifera and macrofossils mentioned in the text.
PLANKTONIC FORAMINIFERA:
Abathomphalus mayaroensis (Bolli, 1951);
Archaeoglobigerina bosquensis (Pessagno, 1967);
Archaeoglobigerina cretacea (d’Orbigny, 1840);
Clavihedbergella simplex (Morrow, 1934);
Clavigedbergella simplicissima (Magne and Sigal, 1954);
Contusotruncana contusa (Cushman, 1926);
Contusotruncana fornicata (Plummer, 1931) (Figure 7B(a–c));
Contusotruncanamorozovae (Vasilenko, 1961);
Contusotruncana patelliformis (Gandolfi, 1955) (Figure 6N(a–c));
Contusotruncana plummerae (Gandolfi, 1955) (Figure 7C(a–c));
Dicarinella algeriana (Caron, 1966);
Dicarinella asymetrica (Sigal, 1952);
Dicarinella canaliculata (Reuss, 1854) (Figure 5F(a–c));
Dicarinella concavata (Brotzen, 1934) (Figure 5L(a,b));
Dicarinella hagni (Scheibnerova, 1962) (Figure 5G(a,b),I(a,b));
Dicarinella imbricata (Mornod, 1950) (Figure 5H(a–c));
Dicarinella longoriai (Peryt, 1980);
Dicarinella primitiva (Dalbiez, 1955);
Falsotruncana maslakovae (Caron, 1981);
Falsotruncana sp. (Figure 5K(a–c));
Gansserina gansseri (Bolli, 1951);
Globigerinelloides bentonensis (Morrow, 1934);
Globigerinelloides multispinus (Lalicker, 1948) (Figure 7J(a,b));
Globigerinelloides prairiehillensis (Pessagno, 1967) (Figure 7K(a,b));
Globigerinelloides ultramicrus (Subbotina, 1949);
Globotruncana aegyptiaca (Nakkady, 1950);
Globotruncana arca (Cushman, 1926) (Figure 6L(a–c));
Globotruncana bulloides (Vogler, 1941) (Figure 6M(a–c));
Globotruncana linneiana (d’Orbigny, 1839) (Figure 6K(a–c));
Globotruncana rugosa (Marie, 1941) (Figure 7E(a–c));
Globotruncana ventricosa (White, 1928) (Figure 7A(a,b));
Globotruncanella havanensis (Voorwijk, 1937);
Globotruncanella minuta (Caron and Gonzalez Donoso in Robaszynski et al., 1984) (Figure 7H(a–c));
Globotruncanella petaloidea (Gandolfi, 1955) (Figure 7I(a–c));
Globotruncanita elevata (Brotzen, 1934);
Globotruncanita pettersi (Gandolfi, 1955) (Figure 7G(a–c));
Globotruncanita stuartiformis (Dalbiez, 1955);
Guembelitria cenomana (Keller, 1935);
Guembelitria cretacea (Cushman, 1933) (Figure 7N);
Helvetoglobotruncana helvetica (Bolli, 1945) (Figure 5J(a,b),M(a,b));
Laeviheterohelix glabrans (Cushman, 1938) (Figure 7L(a,b));
Marginotruncana caronae (Peryt, 1980) (Figure 6C(a,b));
Marginotruncana coronata (Bolli, 1945) (Figure 6A(a–c));
Marginotruncana marginata (Reuss, 1845) (Figure 6B(a–c));
Marginotruncana paraconcavata (Porthault, in Donze et al., 1970) (Figure 6I(a–c));
Marginotruncana pseudolinneiana (Pessagno, 1967) (Figure 6G(a,b));
Marginotruncana schneegansi (Sigal, 1952);
Marginotruncana sigali (Reichel, 1950);
Marginotruncana sinuosa (Porthault, in Donze et al., 1970) (Figure 6D(a,b),E(a,b));
Marginotruncana renzi (Gandolfi, 1942) (Figure 6F(a–c));
Marginotruncana undulata (Lehmann, 1963) (Figure 6J(a–c));
Muricohedbergella delrioensis (Carsey, 1926);
Muricohedbergella planispira (Tappan, 1940);
Planoglobulina brazoensis (Martin, 1972) (Figure 7M);
Planoglobulina carseyae (Plummer, 1931);
Planoheterohelix globulosa (Ehrenberg, 1840);
Planoheterohelix moremani (Cushman, 1938);
Planoheterohelix navarroenis (Loeblich);
Planoheterohelix reussi (Cushman, 1938);
Planoheterohelix striata (Ehrenberg, 1840) (Figure 7O(a,b));
Planoheterohelix vistulaensis (Peryt, 1980);
Plummerita hantkeninoides (Bronnimann, 1952);
Praeglobotruncana delrioensis (Plummer, 1931);
Praeglobotruncana gibba (Klaus, 1960);
Praeglobotruncana oraviensis (Scheibnerova, 1960);
Praeglobotruncana stephani (Gandolfi, 1942);
Pseudoguembelina hariaensis (Nederbragt, 1991);
Pseudotextularia elegans (Rzehak, 1891);
Pseudotextularia nuttalli (Voorwijk, 1937) (Figure 6H(a,b));
Racemiguembelina powelli (Smith and Pessagno, 1973);
Radotruncana calcarata (Cushman, 1927);
Rotalipora cushmani (Morrow, 1934) (Figure 5C(a–c));
Rugoglobigerinahexacamerata (Brönnimann, 1952);
Rugoglobigerinamacrocephala (Brönnimann, 1952);
Rugoglobigerinamilamensis (Smith and Pessagno 1973);
Rugoglobigerina pennyi (Brönnimann, 1952) (Figure 7F(a–c));
Rugoglobigerina rugosa (Plummer, 1927) (Figure 7D(a–c));
Rugoglobigerina scotti (Brönnimann 1952);
Sigalia carpatica (Salaj and Samuel, 1963);
Thalmanninella appenninica (Renz, 1936);
Thalmanninella gandolfii (Luterbacher and Premoli Silva, 1962);
Thalmanninella globotruncanoides (Sigal, 1948) (Figure 5A(a–c));
Thalmanninella greenhornensis (Morrow, 1934) (Figure 5D(a,b));
Thalmanninella reicheli (Mornod, 1950) (Figure 5B(a–c));
Thalmanninella tehamaensis (Marianos and Zingula, 1966);
Whiteinella baltica (Douglas and Rankin, 1969);
Whiteinella aprica (Loeblich and Tappan, 1961);
Whiteinella archaeocretacea (Pessagno, 1967) (Figure 5E(a,b));
Whiteinella brittonensis (Loeblich and Tappan, 1961) (Figure 5N(a–c)).
INOCERAMIDS:
Cataceramus beckumensis (Giers, 1964);
Cataceramus dariensis (Dobrov and Pavlova, 1959);
Cataceramus subcompressus (Meek and Hayden, 1860);
Cremnoceramus deformis (Meek, 1871);
Cremnoceramus waltersdorfensis (Andert, 1911);
Cordiceramus muelleri (Petrascheck, 1906);
Cordiceramus undulatoplicatus (Roemer, 1852);
Endocostea typica (Whitfield, 1877);
“Inoceramus” altus (Meek, 1871);
Inoceramus apicalis (Woods, 1912);
Inoceramus “azerbaydjanensis” (Aliev, 1939);
Inoceramus costellatus (Woods, 1912);
„Inoceramus” ianjonaensis (Sornay, 1973);
“Inoceramus” inkermanensis (Dobrov and Pavlova, 1959);
Inoceramus ex. gr. labiatus (Schlotheim, 1813);
Inoceramus lamarckiParkinson (1819);
“Inoceramus” redbirdensis (Walaszczyk, Cobban and Harries, 2001);
Inoceramus tenuilineatus (Hall and Meek, 1856);
“Inoceramus” vorhelmensis (Walaszczyk, 1997);
Magadiceramus subquadratus (Schlüter, 1887);
Mytiloides scupini (Heinz, 1930);
Sphaeroceramus patootensiformis (Seitz, 1965);
Sphaeroceramus pertenuiformis (Walaszczyk, Cobban and Harries, 2001);
Sphaeroceramus pinniformis (Willet, 1871);
Sphaeroceramus sarumensis (Woods, 1912);
Spyridoceramus tegulatus (Hagenow, 1842);
Tenuipteria argentea (Conrad, 1858);
Trochoceramus costaceus (Khalafova, 1966);
Trochoceramus radiosus (Quaas, 1902);
Volviceramus involutus (Sowerby, 1828).
BELEMNITES:
Belemnitella junior (Nowak, 1913);
Belemnella lanceolata (von Schlotheim, 1813);
Belemnella kazimiroviensis (Skolozdrowna, 1932);
Belemnitella mucronata (Schlotheim, 1813);
Belemnella occidentalis (Birkelund, 1957);
Gonioteuthis granulata (Blainville, 1827);
Gonioteuthis quadrata gracilis (Stolley, 1892).
AMMONITES:
Acantoscaphites tridens (Kner, 1848);
Bostrychoceras polyplocum (Roemer, 1841);
Didymoceramus donezianum (Michailov, 1951);
Hoploscaphites constrictus crassus (Łopuski, 1911);
Hoploscaphites constrictus johnjagti (Machalski, 2005);
Hoploscaphites constrictus lvivensis (Machalski, 2005);
Neocardioceras juddii (Barrois and Guerne, 1878);
Neancyloceras phaleratum (Griepenkerl, 1889);
Nostoceras hyatti (Stephenson, 1941);
Nostoceras pozaryskii (Błaszkiewicz, 1980);
Pachydiscus neubergicus (Hauer, 1858);
Pachydiscus (Patagiosites) stobaei (Nilsson, 1827).
ECHINOIDS
Echinocorys conica (Agassiz, 1847);
Galeolabasiplana (Ernst, 1971);
Galeola papillosa (Leske, 1778);
Galerites vulgaris (Leske, 1778).
References
- Caron, M. Cretaceous planktic foraminifera. In Plankton Stratigraphy; Bolli, H.M., Saunders, J.B., Perch-Nielsen, K., Eds.; Cambridge University Press: Cambridge, UK, 1985; pp. 17–86. [Google Scholar]
- Premoli Silva, I.; Sliter, W.V. Cretaceous planktonic foraminiferal biostratigraphy and evolutionary trends from the Bottaccione Section, Gubbio, Italy. Palaeontogr. Ital. 1994, 81, 2–90. [Google Scholar]
- Robaszynski, F.; Caron, M. Foraminifères planctoniques du Crétacé: Commentaire de la zonation Europe Méditerranée. Bull. Soc. Géol. France 1995, 166, 681–692. [Google Scholar]
- Coccioni, R.; Silva, I.P. Revised Upper Albian–Maastrichtian planktonic foraminiferal biostratigraphy and magnetostratigraphy of the classical Tethyan Gubbio section (Italy). Newslett. Stratigr. 2015, 48, 47–90. [Google Scholar] [CrossRef]
- Bé, A.W.H. An ecological, zoogeographic and taxonomic review of recent planktonic foraminifera. In Oceanic Micropaleontology; Ramsey, T.S., Ed.; Academic Press: London, UK, 1977; pp. 1–100. [Google Scholar]
- Hemleben, C.; Spindler, M.; Anderson, O.R. Modern Planktonic Foraminifera; Springer: Heidelberg, Germany, 1989. [Google Scholar]
- Pożaryska, K.; Peryt, D. The Late Cretaceous and Early Paleocene foraminiferal “Transitional Province” in Poland. In Aspekte der Kreide Europas; Wiedmann, J., Ed.; IUGS, Series A; Schweizerbart: Stuttgart, Germany, 1979; Volume 6, pp. 293–303. [Google Scholar]
- Petrizzo, M.R.; Huber, B.T.; Falzoni, F.; MacLeod, K.G. Changes in biogeographic distribution patterns of southern mid-to high latitude planktonic foraminifera during the Late Cretaceous hot to cool greenhouse climate transition. Cret. Res. 2020, 115, 104547. [Google Scholar] [CrossRef]
- Dubicka, Z.; Peryt, D.; Szuszkiewicz, M. Foraminiferal evidence for paleogeographic and paleoenvironmental changes across the Coniacian–Santonian boundary in western Ukraine. Palaeogeogr. Palaeoclim. Palaeoecol. 2014, 401, 43–56. [Google Scholar] [CrossRef]
- Petrizzo, M.R.; Watkins, D.K.; MacLeod, K.G.; Hasegawa, T.; Huber, B.T.; Batenburg, S.J.; Kato, T. Exploring the paleoceanographic changes registered by planktonic foraminifera across the Cenomanian-Turonian boundary interval and Oceanic Anoxic Event 2 at southern high latitudes in the Mentelle Basin (SE Indian Ocean). Glob. Planet. Chang. 2021, 206, 103595. [Google Scholar] [CrossRef]
- Peryt, D. Planktic Foraminifera zonation of the Upper Cretaceous in the Middle Vistula River valley, Poland. Palaeont. Pol. 1980, 41, 3–101. [Google Scholar]
- Peryt, D. Planktonic foraminifers and the age of chalk from Mielnik (East Poland). Bull. Acad. Pol. Sci. Ser. Sci. Terre 1981, 29, 137–142. [Google Scholar]
- Peryt, D. Mid-Cretaceous microbiostratigraphy and foraminifers of the NE margins of the Świętokrzyskie (Holy Cross) Mts., Poland. Acta Palaeont. Pol. 1983, 28, 417–466. [Google Scholar]
- Peryt, D. Planktonic foraminiferal zonation of Mid-Cretaceous of the Annopol Anticline (Central Poland). Zitteliana 1983, 10, 575–583. [Google Scholar]
- Peryt, D. Paleoecology of middle and late Cretaceous foraminifers from the Lublin Upland (SE Poland). Rév. Paléobiol. 1988, 2, 311–321. [Google Scholar]
- Peryt, D. Maastrichtian extinctions of planktonic foraminifera in central and eastern Poland. Rev. Esp. Paleont. 1988, 3, 105–115. [Google Scholar]
- Peryt, D.; Wyrwicka, K. The Cenomanian/Turonian boundary event in Central Poland. Palaeogeogr. Palaeoclim. Palaeoecol. 1993, 104, 185–197. [Google Scholar] [CrossRef]
- Walaszczyk, I.; Peryt, D. Inoceramid-foraminiferal biostratigraphy of the Turonian through Santonian deposits of the Middle Vistula Section, Central Poland. Zbl. Paläont. Teil I 1996, 11/12, 1501–1513. [Google Scholar]
- Peryt, D.; Witek, A.; Jaskuła, I. Foraminifers and ostracods from the Campanian of the Jeżówka quarry, the Miechów Trough, southern Poland: Paleoecological implications. In 9th Paleontological Conference, Warszawa, 10–11 October 2008; Pisera, A., Bitner, M.A., Halamski, A.T., Eds.; Polish Academy of Sciences; Institute of Paleobiology: Warszawa, Poland, 2008; pp. 69–71. [Google Scholar]
- Peryt, D.; Dubicka, Z. Palaeoenvironmental changes in the early Maastrichtian of the Nida Trough (Southern Poland): Foraminiferal evidence. Grzybowski Found. Spec. Publ. 2009, 15, 101–108. [Google Scholar]
- Peryt, D.; Dubicka, Z. Foraminiferal bioevents in the Upper Campanian to lowest Maastrichtian of the Middle Vistula River section, Poland. Geol. Quart. 2015, 59, 814–830. [Google Scholar] [CrossRef] [Green Version]
- Walaszczyk, I.; Wood, C.J.; Lees, J.; Peryt, D.; Voigt, S.; Wiese, F. The Salzgitter-Salder Quarry (Lower Saxony, Germany) and Słupia Nadbrzeżna river cliff section (central Poland): A proposed candidate composite Global Boundary Stratotype Section and Point for the base of the Coniacian Stage (Upper Cretaceous). Acta Geol. Pol. 2010, 60, 445–477. [Google Scholar]
- Dubicka, Z.; Peryt, D. Integrated biostratigraphy of Upper Maastrichtian chalk at Chełm (SE Poland). Ann. Soc. Geol. Pol. 2011, 81, 185–197. [Google Scholar]
- Dubicka, Z.; Peryt, D. Foraminifers and stable isotope record of the Dubivtsi chalk (upper Turonian, Western Ukraine): Palaeoenvironmental implications. Geol. Quart. 2012, 56, 199–214. [Google Scholar]
- Dubicka, Z.; Peryt, D. Latest Campanian and Maastrichtian palaeoenvironmental changes: Implications from an epicontinental sea (SE Poland and western Ukraine). Cret. Res. 2012, 37, 272–284. [Google Scholar] [CrossRef]
- Dubicka, Z.; Peryt, D. The Lower/Upper Maastrichtian boundary inerval in the Lublin Syncline (SE Poland, Boreal realm): New insight into foraminiferal biostratigraphy. Newsl. Stratigr. 2012, 45, 139–150. [Google Scholar] [CrossRef] [Green Version]
- Dubicka, Z. Otwornice i Stratygrafia Osadów Górnej Kredy Okolic Halicza (Ukraina Zachodnia) [Foraminifers and Upper Cretaceous Stratigraphy of Halych Area (Western Ukraine)]. Ph.D. Thesis, Institute of Paleobiology, Warsaw, Poland, 2012. [Google Scholar]
- Dubicka, Z.; Bojanowski, M.; Peryt, D.; Barski, M. Biotic and isotopic vestiges of oligotrophy on continental shelves during Oceanic Anoxic Event 2. Glob. Biogeochem. Cycles 2021, 35, e2020GB006831. [Google Scholar] [CrossRef]
- Dubicka, Z. Benthic foraminiferal biostratigraphy of the lower and middle Campanian of the Polish Lowlands and its application for interregional correlation. Cret. Res. 2015, 56, 491–503. [Google Scholar] [CrossRef]
- Bojanowski, M.J.; Dubicka, Z.; Minoletti, F.; Olszewska-Nejbert, D.; Surowski, M. Stable C and O isotopic study of the Campanian chalk from the Mielnik section (eastern Poland): Signals from bulk rock, belemnites, benthic foraminifera, nannofossils and microcrystalline cements. Palaeogeogr. Palaeoclim. Palaeoecol. 2017, 465, 193–211. [Google Scholar] [CrossRef]
- Machalski, M.; Vellekoop, J.; Dubicka, Z.; Peryt, D.; Harasimiuk, M. Late Maastrichtian cephalopods, dinoflagellate cysts and foraminifera from the Cretaceous–Paleogene succession at Lechówka, southeast Poland: Stratigraphic and environmental implications. Cret. Res. 2016, 57, 208–227. [Google Scholar] [CrossRef]
- Walaszczyk, I.; Dubicka, Z.; Olszewska-Nejbert, D.; Remin, Z. Integrated biostratigraphy of the Santonian through Maastrichtian (Upper Cretaceous) of extra-Carpathian Poland. Acta Geol. Pol. 2016, 66, 313–358. [Google Scholar] [CrossRef] [Green Version]
- Dubicka, Z.; Machalski, M. Foraminiferal record in a condensed marine succession: A case study from the Albian and Cenomanian (mid-Cretaceous) of Annopol, Poland. Geol. Mag. 2017, 154, 399–418. [Google Scholar] [CrossRef]
- Remin, Z.; Cyglicki, M.; Barski, M.; Dubicka, Z.; Roszkowska-Remin, J. The K-Pg boundary section at Nasiłów, Poland: Stratigraphic reassessment based on foraminifers, dinoflagellate cysts and palaeomagnetism. Geol Quart. 2021, 65, 45. [Google Scholar] [CrossRef]
- Ziegler, P.A. Geological Atlas of Western and Central Europe; Geological Society Publishing House: Bath, UK, 1990. [Google Scholar]
- Dadlez, R.; Marek, S.; Pokorski, J. Palaeogeographical Atlas of the Epicontinental Permian and Mesozoic in Poland, 1:2 500 000; Państwowy Instytut Geologiczny: Warsaw, Poland, 1998. [Google Scholar]
- Świdrowska, J.; Hakenberg, M.; Poluhtovič, B.; Seghedi, A.; Višnâkov, I. Evolution of the Mesozoic basin on the southwestern edge of the East European Craton (Poland, Ukraine, Moldova, Romania). Stud. Geol. Pol. 2008, 130, 3–130. [Google Scholar]
- Dubicka, Z.; Jurkowska, A.; Thibault, N.; Razmjooei, M.J.; Wójcik, K.; Gorzelak, P.; Felisiak, I. An integrated stratigraphic study across the Santonian/Campanian boundary at Bocieniec, southern Poland: A new boundary stratotype candidate. Cret. Res. 2017, 80, 61–85. [Google Scholar] [CrossRef]
- Jurkowska, A.; Świerczewska-Gładysz, E.; Dubicka, Z.; Olszewska-Nejbert, D. Porosphaera globularis (Phillips, 1829) (Porifera, Calcarea) in the Campanian (Upper Cretaceous) of extra-Carpathian Poland. Acta Geol. Pol. 2015, 65, 121–139. [Google Scholar] [CrossRef] [Green Version]
- Pożaryski, W. Stratygrafia senonu w przełomie Wisły między Rachowem i Puławami. Biul. Państw. Inst. Geol. 1938, 6, 3–94. (In Polish) [Google Scholar]
- Pożaryska, K. Lagenidae du Crétacé supérieur de Pologne. Palaeont. Pol. 1957, 8, 3–261. [Google Scholar]
- Walaszczyk, I. Turonian through Santonian deposits of the Central Polish Uplands; their facies development, inoceramid paleontology and stratigraphy. Acta Geol. Pol. 1992, 42, 1–122. [Google Scholar]
- Walaszczyk, I. Inoceramids and inoceramid biostratigraphy of the Upper Campanian to basal Maastrichtian of the Middle Vistula River section, central Poland. Acta Geol. Pol. 2004, 54, 95–168. [Google Scholar]
- Pożaryski, W.; Brochwicz-Lewiński, W.; Brodowicz, Z.; Jaskowiak-Schoeneich, M.; Milewicz, J.; Sawicki, L.; Uberna, T. Geological Map of Poland and Adjoining Countries, without Cenozoic Formations (without Quaternary in the Carpathians); Wyd. Geol.: Warsaw, Poland, 1979. [Google Scholar]
- Machalski, M.; Jagt, J.W.M.; Dubicka, Z. Additional records of scaphitid ammonites from the basal upper Maastrichtian (Upper Cretaceous) of eastern Poland. Bull. Inst. Royal Sci. Natur. Belg. Sci. Terre 2008, 78, 261–268. [Google Scholar]
- Pasternak, S.I.; Sen’kovskyiy, M.; Gavrylyshyn, V.I. Volyno-Podillia u Kreydovomu Periodi; Naukova Dumka: Kiev, Ukraine, 1987. (In Ukrainian) [Google Scholar]
- Sigal, J. Essai de zonation du Crétacé méditerranéen à l’aide des foraminifères planctoniques. Geol. Mediter. 1977, 4, 99–108. [Google Scholar] [CrossRef]
- Gale, A.S.; Mutterlose, J.; Batenburg, S.; Gradstein, F.M.; Agterberg, F.P.; Ogg, J.G.; Petrizzo, M.R. Chapter 27, The Cretaceous Period. In Geologic Time Scale; Gradstein, F.M., Ogg, J.G., Schmitz, M.D., Ogg, G.M., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 1023–1086. [Google Scholar]
- Błaszkiewicz, A. Campanian and Maastrichtian ammonites of the Middle Vistula River valley, Poland: A stratigraphic and paleontological study. Prace Inst. Geol. 1980, 92, 3–63. [Google Scholar]
- Machalski, M. A new ammonite zonation for the Maastrichtian Stage in Poland. In The Maastrichtian Stage; the Current Concept; Jagt, J.W.M., Jagt-Yazykova, E.A., Eds.; Natuurhistorisch Museum Maastricht: Maastricht, The Netherlands, 2012; pp. 40–44. [Google Scholar]
- Remin, Z. Upper Coniacian, Santonian, and lowermost Campanian ammonites of the Lipnik–Kije section, central Poland, taxonomy, stratigraphy, and palaeogeographic significance. Cret. Res. 2010, 31, 154–180. [Google Scholar] [CrossRef]
- Jagt, J.W.M.; Walaszczyk, I.; Yazykova, E.A.; Zatoń, M. Linking southern Poland and Northern Germany: Campanian cephalopods, inoceramid bivalves and echinoids. Acta Geol. Pol. 2004, 54, 573–586. [Google Scholar]
- Bolli, H.M. Zonation of Cretaceous to Pliocene marine sediments based on planktonic foraminifera. Asoc. Venezolana Geol. Min. Petrol. Bol. Inform. 1966, 9, 2–32. [Google Scholar]
- Borsetti, A.M. Foraminiferi planctonici di una serie cretacea dei Dintorni di Piobbico (Prov. di Pesaro). Giorn. Geol. 1962, 29, 19–75. [Google Scholar]
- Sigal, J. Notes micropaléontologiques nord-africaines.du Cénomanien au Santonien: Zones et limites en faciès pélagique. C. R. Somm. Soc. Géol. Fr. 1955, 7–8, 157–160. [Google Scholar]
- Nishi, H.; Takashima, R.; Hatsugai, T.; Saito, T.; Moriya, K.; Ennyu, A.; Sakai, T. Planktonic foraminiferal zonation in the Cretaceous Yezo Group, Central Hokkaido, Japan. J. Asian Earth Sci. 2003, 21, 867–886. [Google Scholar] [CrossRef] [Green Version]
- Nederbragt, A.J. Biostratigraphy and Paleoceanographic Potential of the Cretaceous Planktic Foraminifera Heterohelicidae; Vrije University: Amsterdam, The Netherlands, 1990; 204p. [Google Scholar]
- Salaj, J.; Samuel, O. Foraminifera der Westkarpaten-Kreide; Geologicky ustav Dionyza Stura: Bratislava, Slovakia, 1966. [Google Scholar]
- Petrizzo, M.R.; Falzoni, F.; Premoli Silva, I. Identification of the base of the lower-to-middle Campanian Globotruncana ventricosa Zone: Comments on reliability and global correlations. Cret. Res. 2011, 32, 387–405. [Google Scholar] [CrossRef]
- Linnert, C.; Robinson, S.A.; Lees, J.A.; Bown, P.B.; Pérez-Rodríguez, I.; Petrizzo, M.R.; Falzoni, F.; Littler, K.; Arz, J.A.; Russel, E.E. Evidence for global cooling in the Late Cretaceous. Nature Comm. 2014, 5, 4194. [Google Scholar] [CrossRef] [Green Version]
- Friedrich, O.; Norris, R.D.; Erbacher, J. Evolution of middle to Late Cretaceous oceans—A 55 m.y. record of Earth’s temperature and carbon cycle. Geology 2012, 40, 107–110. [Google Scholar] [CrossRef]
- Cieśliński, S. Development of the Danish-Polish Furrow in the Góry Świętokrzyskie Region in the Albian, Cenomanian and Lower Turonian. Biul. Inst. Geol. 1976, 295, 249–271, (In Polish with English summary). [Google Scholar]
- Marcinowski, R.; Radwański, A. The mid-Cretaceous transgression onto the Central Polish Uplands (marginal part of the Central European Basin). Zitteliana 1983, 10, 65–96. [Google Scholar]
- Walaszczyk, I. Mid-Cretaceous events at the marginal part of the Central European Basin (Annopol-on-Vistula section, Central Poland). Acta Geol. Pol. 1987, 37, 61–74. [Google Scholar]
- Gale, A.S.; Kennedy, W.J.; Burnett, J.A.; Caron, M.; Kidd, B.E. The Late Albian to Early Cenomanian succession at Mont Risou near Rosans (Drȏme, SE France): An integrated study (ammonites, inoceramids, planktonic foraminifera, nannofossils, oxygen and carbon isotopes). Cret. Res. 1996, 17, 515–606. [Google Scholar] [CrossRef]
- Kennedy, W.J.; Gale, A.S.; Lees, J.A.; Caron, M. The Global Boundary Stratotype Section and Point (GSSP) for the base of the Cenomanian Stage, Mont Risou, Hautes-Alpes, France. Episodes 2004, 27, 21–32. [Google Scholar] [CrossRef] [Green Version]
- Caron, M.; Dall’Agnolo, S.; Accarie, H.; Barrera, E.; Kauffman, E.G.; Amédro, F.; Robaszynski, F. High-resolution stratigraphy of the Cenomanian-Turonian boundary interval at Pueblo (USA) and Wadi Bahloul (Tunisia): Stable isotopes and bioevents correlation. Géobios 2006, 39, 171–200. [Google Scholar] [CrossRef]
- Huber, B.T.; Petrizzo, M.R. Evolution and taxonomic study of the Cretaceous planktic foraminiferal genus Helvetoglobotruncana Reiss, 1957. J. Foraminifer. Res. 2014, 44, 40–57. [Google Scholar] [CrossRef]
- Haynes, S.J.; MacLeod, K.G.; Huber, B.T.; Warny, S.; Kaufman, A.J.; Pancost, R.D.; Berrocoso, A.J.; Petrizzo, M.R.; Watkins, D.K.; Zhelezinskaia, I. Southeastern Tanzania depositional environments, marine and terrestrial links, and exceptional microfossil preservation in the warm Turonian. GSA Bull. 2017, 129, 515–533. [Google Scholar] [CrossRef]
- Huber, B.T.; Petrizzo, M.R.; Watkins, D.K.; Haynes, S.J.; MacLeod, K.G. Correlation of Turonian continental margin and deep-sea sequences in the subtropical Indian Ocean sediments by integrated planktonic foraminiferal and calcareous nannofossil biostratigraphy. Newsl. Stratigr. 2017, 50, 141–185. [Google Scholar] [CrossRef]
- Douglas, R.G.; Rankin, C. Cretaceous planktonic foraminifera from Bornholm Island and their zoogeographic significance. Lethaia 1969, 2, 185–217. [Google Scholar] [CrossRef]
- Norling, E. The foraminiferal fauna. Medd. Dansk Geol. Foren. 1973, 22, 97–112. [Google Scholar]
- Petrizzo, M.R. Upper Turonian–lower Campanian planktonic foraminifera from southern mid–high latitudes (Exmouth Plateau, NW Australia): Biostratigraphy and taxonomic notes. Cret. Res. 2000, 21, 479–505. [Google Scholar] [CrossRef]
- Hart, M.B.; Bailey, H. The distribution of planktonic Foraminiferida in the mid-Cretaceous of NW Europe. In Aspekte der Kreide Europas; Wiedmann, J., Ed.; Series A; IUGS: Paris, France, 1979; Volume 6, pp. 527–542. [Google Scholar]
- Voigt, S.; Wiese, F. Evidence for late Cretaceous (Late Turonian) climate cooling from oxygen-isotope variations and palaeobiogeographic changes in Western and Central Europe. J. Geol. Soc. 2000, 157, 737–743. [Google Scholar] [CrossRef]
- Hancock, J.M.; Walaszczyk, I. Mid-Turonian to Coniacian changes of sea-level around Dallas, Texas. Cret. Res. 2004, 25, 459–471. [Google Scholar] [CrossRef]
- Ogg, J.G.; Agterberg, F.P.; Gradstein, F.M. The Cretaceous Period. In The Geologic Time Scale 2004; Gradstein, F.M., Ogg, J.G., Smith, A.G., Eds.; Cambridge University Press: Cambridge, UK, 2004; pp. 344–383. [Google Scholar]
- Walaszczyk, I.; Čech, S.; Crampton, J.C.; Dubicka, Z.; Ifrim, C.; Jarvis, I.; Kennedy, W.J.; Lees, J.A.; Lodowski, D.; Pearce, M.; et al. The Global Boundary Stratotype Section and Point (GSSP) for the base of the Coniacian Stage (Salzgitter-Salder, Germany) and its auxiliary sections (Słupia Nadbrzeżna, central Poland; Střeleč, Czech Republic; and El Rosario, NE Mexico). Episodes, 2021; Ahead of Print. [Google Scholar] [CrossRef]
- Lamolda, M.A.; Peryt, D.; Ion, J. Planktonic foraminiferal bioevents in the Coniacian/Santonian boundary interval at Olazagutia, Navarra province, Spain. Cret. Res. 2007, 28, 18–29. [Google Scholar] [CrossRef]
- Lamolda, M.A.; Paul, C.R.C.; Peryt, D.; Pons, J.M. The Global Boundary Stratotype and Section Point (GSSP) for the base of the Santonian Stage, “Cantera de Margas”, Olazagutia, northern Spain. Episodes 2014, 37, 2–13. [Google Scholar] [CrossRef] [Green Version]
- Elamri, Z.; Farouk, S.; Zaghib-Turki, D. Santonian planktonic foraminiferal biostratigraphy of the northern Tunisia. Geol. Croat. 2014, 67, 111–126. [Google Scholar] [CrossRef]
- Haynes, S.J.; Huber, B.T.; MacLeod, K.G. Evolution and phylogeny of mid-Cretaceous (Albian-Coniacian) biserial planktic foraminifera. J. Foraminifer. Res. 2015, 45, 42–81. [Google Scholar] [CrossRef]
- Petrizzo, M.R.; Jimenez Berrocoso, A.; Falzoni, F.; Huber, B.T.; MacLeods, K.G. The Coniacian–Santonian sedimentary record in southern Tanzania (Ruvuma Basin, East Africa): Planktonic foraminiferal evolutionary, geochemical and palaeoceanographic patterns. Sedimentology 2017, 64, 252–285. [Google Scholar] [CrossRef] [Green Version]
- Pasternak, S.I. Biostratygrafiya Kreydovykh Vidkladiv Volyno-Podilskoi Plyty; Vydavnytstvo Akademii Nauk Ukrainskoy RSR: Kiev, Ukraine, 1959. (In Ukrainian) [Google Scholar]
- Pożaryski, W. Outline of Paleozoic and Mesozoic tectonics of Polish Lowlands. Kwart. Geol. 1964, 8, 1–41, (In Polish with English summary). [Google Scholar]
- Miniati, F.; Petrizzo, M.R.; Falzoni, F.; Erba, E. Calcareous plankton biostratigraphy of the Santonian-Campanian boundary interval in the Bottaccione section (Umbria-Marche basin, central Italy). Riv. It. Paleont. Strat. 2020, 126, 783–801. [Google Scholar] [CrossRef]
- Dalbiez, F. The genus Globotruncana in Tunisia. Micropaleontology 1955, 1, 161–170. [Google Scholar] [CrossRef]
- Georgescu, M.D. Upper Cretaceous planktic foraminiferal biostratigraphy. Studia UBB Geologia 2017, 61, 5–20. [Google Scholar] [CrossRef] [Green Version]
- Witek, A. Otwornice Planktoniczne i Biostratygrafia Kampanu Okolic Wolbromia (Południowa Polska). Master’s Thesis, Silesian University, Faculty of Earth Sciences, Sosnowiec, Poland, 2006; 55p. (In Polish). [Google Scholar]
- Hancock, J.M. Transatlantic correlations in the Campanian-Maastrichtian stages by eustatic changes of sea-level. Geol. Soc. Spec. Publ. 1993, 70, 241–256. [Google Scholar] [CrossRef]
- Niechwedowicz, M.; Walaszczyk, I.; Barski, M. Phytoplankton response to palaeoenvironmental changes across the Campanian–Maastrichtian (Upper Cretaceous) boundary interval of the Middle Vistula River section, central Poland. Palaeogeogr. Palaeoclim. Palaeoecol. 2021, 577, 110558. [Google Scholar] [CrossRef]
- Olferev, A.G.; Beniamovski, V.N.; Vishnevskaya, V.S.; Ivanov, A.V.; Kopaevich, L.F.; Pervushov, E.M.; Seltser, V.B.; Tesakova, E.M.; Kharitonov, V.M.; Shcherbinina, E.A. Upper Cretaceous deposits in the north west of Saratov oblast, Part 1: Litho- and biostratigraphic analysis of the Vishnevoe section. Stratigr. Geol. Corr. 2007, 15, 610–655. [Google Scholar] [CrossRef]
- Beniamovskii, V.N.; Kopaevich, L. Late Cretaceous palaeobiogeography and migrations of Foraminifera in western Eurasia. Österr. Akad. Wiss., Schriftenr. Erdwiss. Komm. 2002, 15, 61–78. [Google Scholar]
- Odin, G.S.; Lamaurelle, M.A. The global Campanian–Maastrichtian stage boundary. Episodes 2001, 24, 229–238. [Google Scholar] [CrossRef]
- Arz, J.A.; Molina, E. Planktic foraminiferal quantitative analysis across the Campanian/Maastrichtian boundary at Tercis les Bains (France). Dev. Palaeont. Stratigr. 2001, 19, 338–348. [Google Scholar]
- Pérez-Rodríguez, I.; Lees, J.A.; Larrasoaña, J.C.; Arz, J.A.; Arenillas, I. Planktonic foraminiferal and calcareous nannofossil biostratigraphy and magnetostratigraphy of the uppermost Campanian and Maastrichtian at Zumaia, northern Spain. Cret. Res. 2012, 37, 100–126. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
(A) Tectonic map sketch of Central Europe showing the location of the studied sections (after [37]); (B) geographical location of the sections in the Middle Vistula River Valley.
Figure 1.
(A) Tectonic map sketch of Central Europe showing the location of the studied sections (after [37]); (B) geographical location of the sections in the Middle Vistula River Valley.

Figure 2.
Stratigraphic distribution of planktonic foraminifera in the Polanówka UW-1 borehole section.
Figure 2.
Stratigraphic distribution of planktonic foraminifera in the Polanówka UW-1 borehole section.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Peryt, D.; Dubicka, Z.; Wierny, W. Planktonic Foraminiferal Biostratigraphy of the Upper Cretaceous of the Central European Basin. Geosciences 2022, 12, 22. https://doi.org/10.3390/geosciences12010022
AMA Style
Peryt D, Dubicka Z, Wierny W. Planktonic Foraminiferal Biostratigraphy of the Upper Cretaceous of the Central European Basin. Geosciences. 2022; 12(1):22. https://doi.org/10.3390/geosciences12010022
Chicago/Turabian StylePeryt, Danuta, Zofia Dubicka, and Weronika Wierny. 2022. "Planktonic Foraminiferal Biostratigraphy of the Upper Cretaceous of the Central European Basin" Geosciences 12, no. 1: 22. https://doi.org/10.3390/geosciences12010022
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.