
Involvement of MicroRNA-27a-3p in the Licorice-Induced Alteration of Cd28 Expression in Mice

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Licorice Decoction
2.2. Animal Experiments
2.3. MiRNA Expression Profile
2.4. Transcriptome Analysis
2.5. MiRNA Target Gene Prediction
2.6. Identification of Key miRNAs
2.7. Identification of mmu-miR-27a-3p Target Genes
2.8. Construction of Dual-Luciferase Reporter Plasmid
2.9. Cell Culture and Transfection
2.10. Luciferase Reporter Assay
2.11. Flow Cytometry
2.12. Quantitative Real-Time PCR (qRT-PCR)
2.13. Statistical Analysis
3. Results
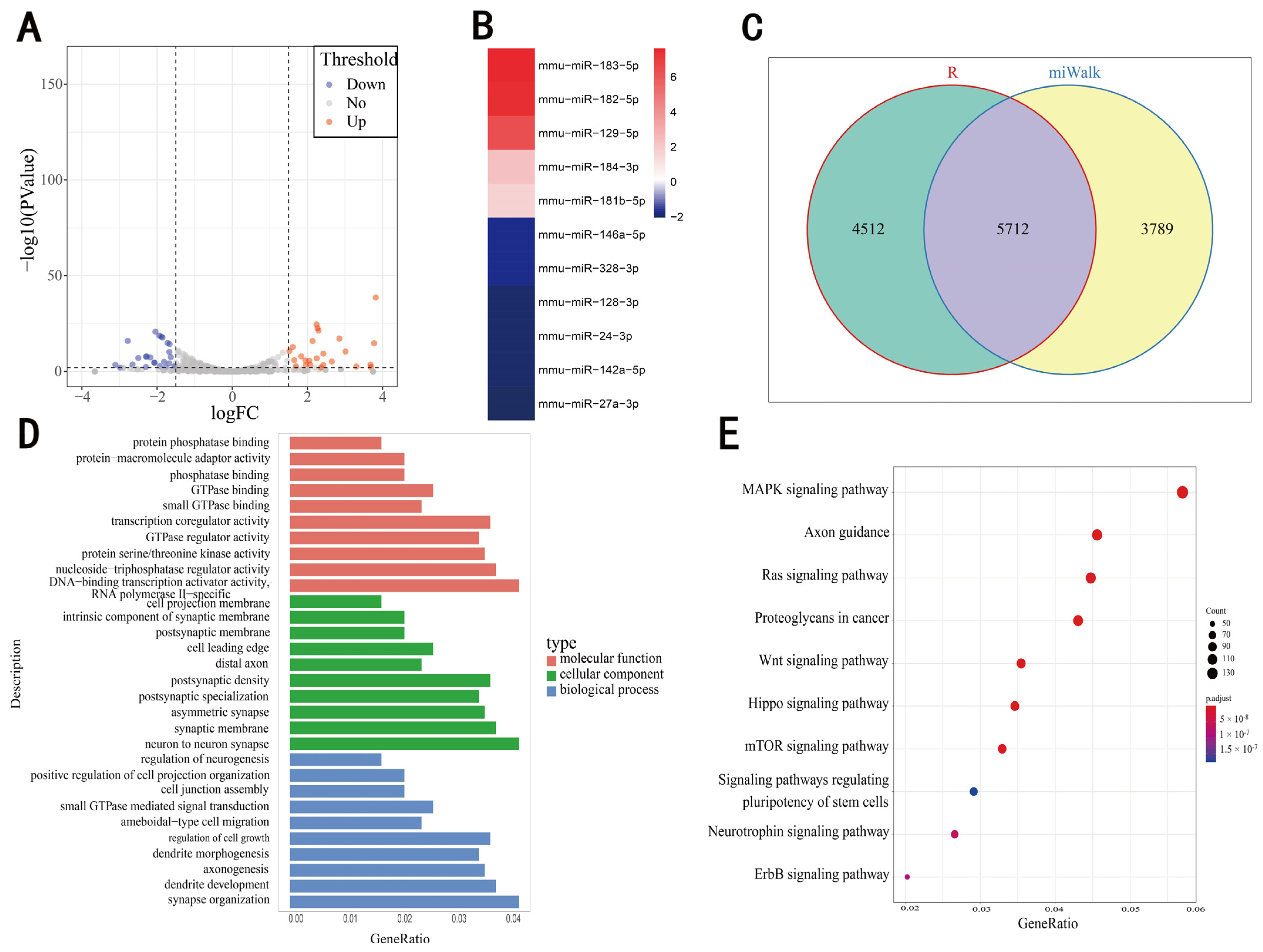
3.1. Screening of Differentially Expressed microRNAs and Prediction of Their Target Genes
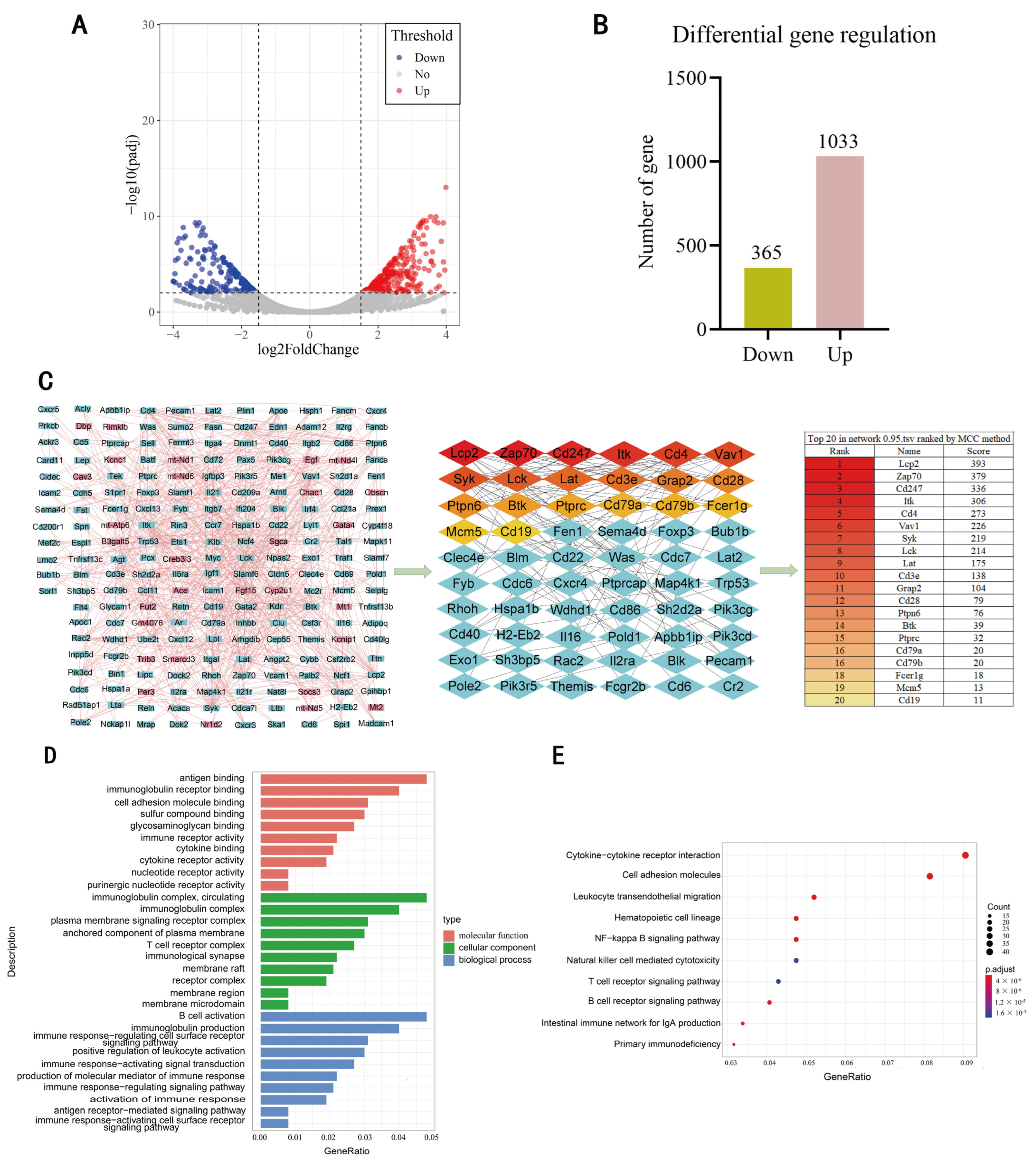
3.2. Screening of DEgenes Based on RNA-Seq
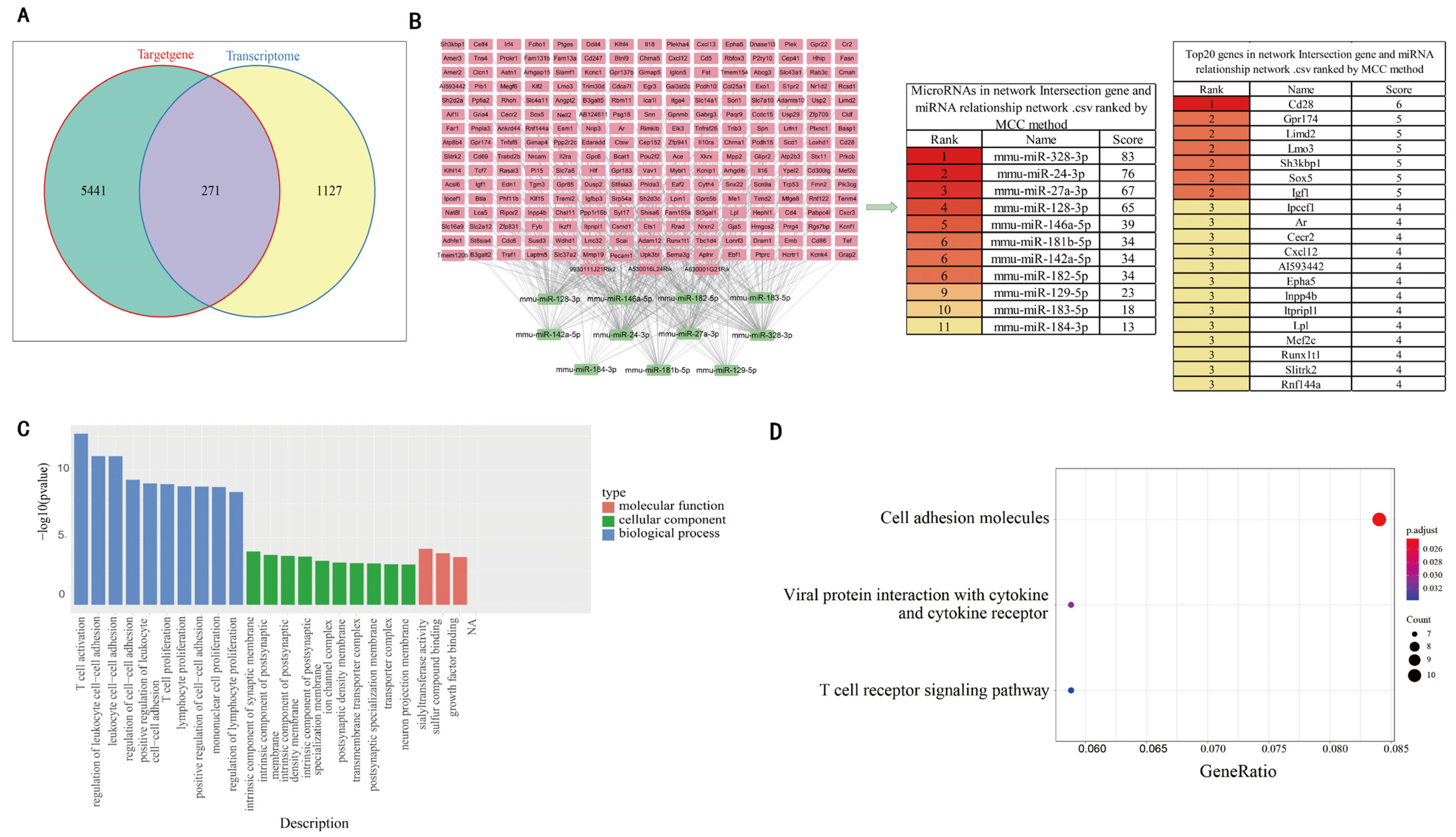
3.3. Comprehensive Analysis of DEgenes and Predicted Target Genes of DEmiRNAs
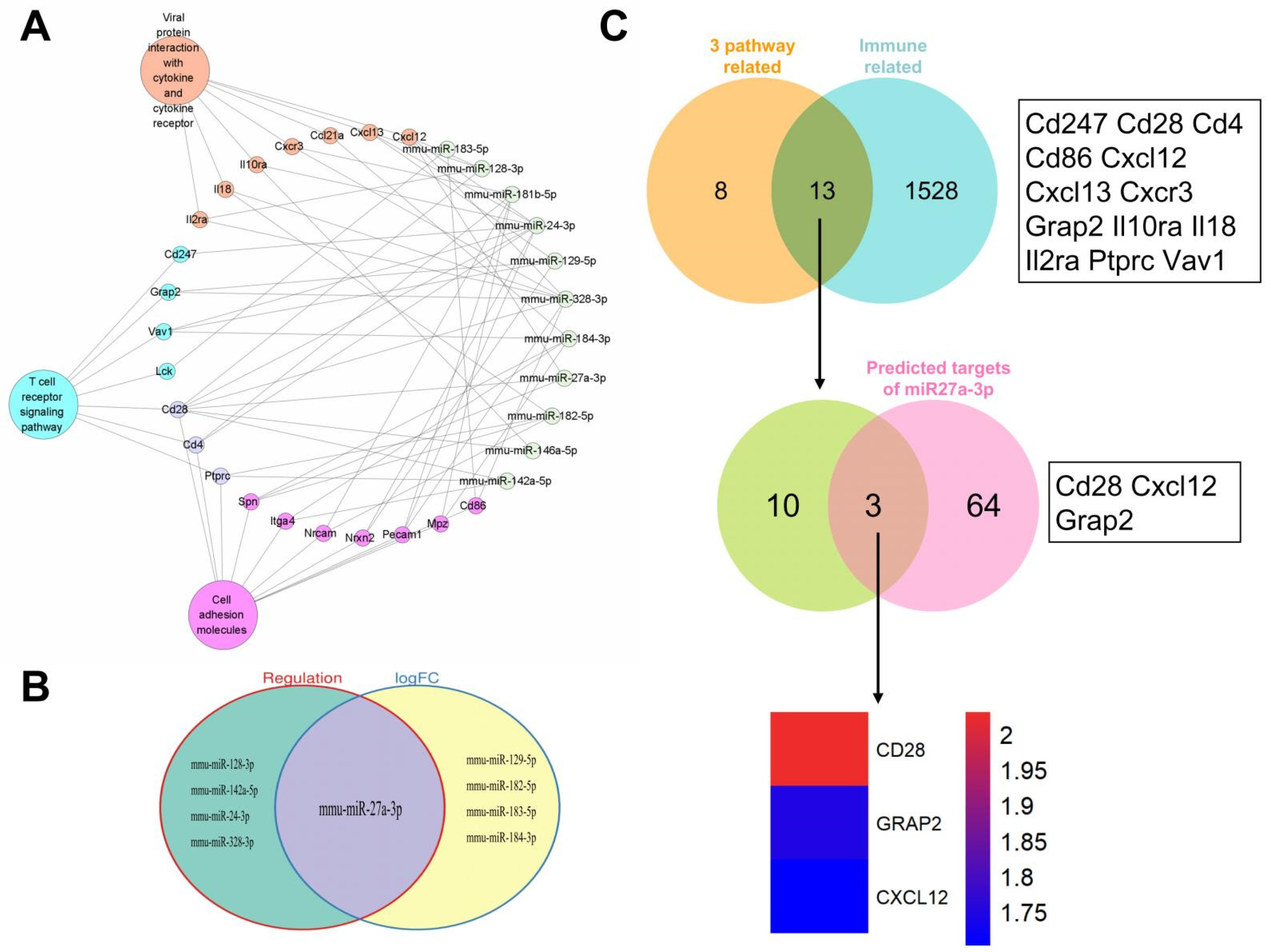
3.4. Identification of mmu-miR-27a-3p as a Key miRNA and Prediction of Its Target Genes
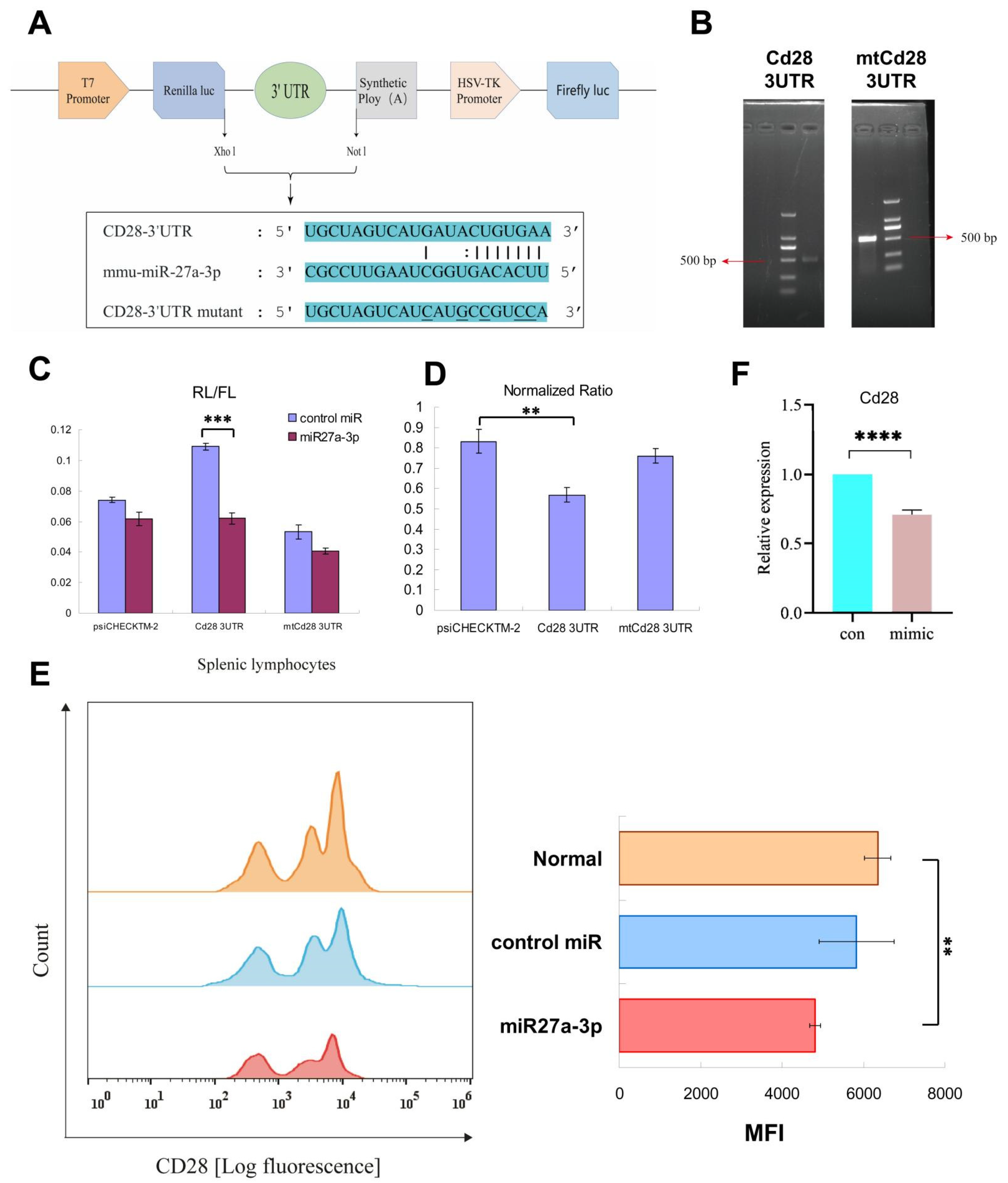
3.5. Regulation of Cd28 Gene Expression by mmu-miRNA-27a-3p
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Armanini, D.; Fiore, C.; Mattarello, M.J.; Bielenberg, J.; Palermo, M. History of the Endocrine Effects of Licorice. Exp. Clin. Endocrinol. Diabet. 2002, 110, 257–261. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Yuan, B.C.; Ma, Y.S.; Zhou, S.; Liu, Y. The Anti-Inflammatory Activity of Licorice, a Widely Used Chinese Herb. Pharm. Biol. 2017, 55, 5–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bi, X.L.; Gong, M.R.; Di, L.Q. Review on Prescription Compatibility of Shaoyao Gancao Decoction and Reflection on Pharmacokinetic Compatibility Mechanism of Traditional Chinese Medicine Prescription Based on In Vivo Drug Interaction of Main Efficacious Components. Evid.-Based Complement Altern. Med. 2014, 2014, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Upadhyay, S.; Mantha, A.K.; Dhiman, M. Glycyrrhiza Glabra (Licorice) Root Extract Attenuates Doxorubicin-Induced Cardiotoxicity via Alleviating Oxidative Stress and Stabilising the Cardiac Health in H9c2 Cardiomyocytes. J. Ethnopharmacol. 2020, 258, 13. [Google Scholar] [CrossRef]
- Yang, R.; Wang, L.Q.; Yuan, B.C.; Liu, Y. The Pharmacological Activities of Licorice. Planta Med. 2015, 81, 1654–1669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chinese Pharmacopoeia Commission. Pharmacopoeia of the People’s Republic of China, 1st ed.; Chemical Industry Press: Beijing, China, 2020; pp. 88–89. [Google Scholar]
- Chen, K.; Yang, R.; Shen, F.Q.; Zhu, H.L. Advances in Pharmacological Activities and Mechanisms of Glycyrrhizic Acid. Curr. Med. Chem. 2020, 27, 6219–6243. [Google Scholar] [CrossRef]
- Han, Y.X.; Jia, Q.J.; Yang, D.F.; Chai, W.G.; Zhang, X.M.; He, Q.L.; Liang, Z.S. Current Advances in Environmental Stimuli Regulating the Glycyrrhizic Acid Biosynthesis Pathway. Fitoterapia 2021, 151, 7. [Google Scholar] [CrossRef]
- Sabbadin, C.; Bordin, L.; Dona, G.; Manso, J.; Avruscio, G.; Armanini, D. Licorice: From Pseudohyperaldosteronism to Therapeutic Uses. Front. Endocrinol. 2019, 10, 6. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Hu, C.; Hood, M.; Zhang, X.; Zhang, L.; Kan, J.T.; Du, J. A Novel Combination of Vitamin C, Curcumin and Glycyrrhizic Acid Potentially Regulates Immune and Inflammatory Response Associated with Coronavirus Infections: A Perspective from System Biology Analysis. Nutrients 2020, 12, 17. [Google Scholar] [CrossRef]
- Katayama, M.; Fukuda, T.; Okamura, T.; Suzuki, E.; Tamura, K.; Shimizu, Y.; Suda, Y.; Suzuki, K. Effect of Dietary Addition of Seaweed and Licorice on the Immune Performance of Pigs. Anim. Sci. J. 2011, 82, 274–281. [Google Scholar] [CrossRef]
- Rashidi, N.; Khatibjoo, A.; Taherpour, K.; Akbari-Gharaei, M.; Shirzadi, H. Effects of Licorice Extract, Probiotic, Toxin Binder and Poultry Litter Biochar on Performance, Immune Function, Blood Indices and Liver Histopathology of Broilers Exposed to Aflatoxin-B-1. Poult. Sci. 2020, 99, 5896–5906. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Han, M.Y.; Fu, Z.Y.; Wang, Y.F.; Zhao, W.; Yu, G.; Ma, Z.H. Immune Responses of Asian Seabass Lates Calcarifer to Dietary Glycyrrhiza Uralensis. Animals 2020, 10, 12. [Google Scholar] [CrossRef] [PubMed]
- Reda, F.M.; El-Saadony, M.T.; El-Rayes, T.K.; Farahat, M.; Attia, G.; Alagawany, M. Dietary Effect of Licorice (Glycyrrhiza Glabra) on Quail Performance, Carcass, Blood Metabolites and Intestinal Microbiota. Poult. Sci. 2021, 100, 8. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Wang, C.; Cai, H.L.; Wen, J.; Fang, P.F. Licorice Extracts Attenuate Nephrotoxicity Induced by Brucine through Suppression of Mitochondria Apoptotic Pathway and Stat3 Activation. Curr. Med. Sci. 2019, 39, 890–898. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.P.; Lusvarghi, S.; Hsiao, S.H.; Liu, T.C.; Li, Y.Q.; Huang, Y.H.; Hung, T.H.; Ambudkar, S.V. Licochalcone a Selectively Resensitizes Abcg2-Overexpressing Multidrug-Resistant Cancer Cells to Chemotherapeutic Drugs. J. Nat. Prod. 2020, 83, 1461–1472. [Google Scholar] [CrossRef]
- Abdel-Wahab, A.H.A.; Effat, H.; Mahrous, E.A.; Ali, M.A.; Al-Shafie, T.A. A Licorice Roots Extract Induces Apoptosis and Cell Cycle Arrest and Improves Metabolism via Regulating Mirnas in Liver Cancer Cells. Nutr. Cancer 2021, 73, 1047–1058. [Google Scholar] [CrossRef]
- Tu, C.; Ma, Y.Z.; Song, M.Y.; Yan, J.Y.; Xiao, Y.F.; Wu, H. Liquiritigenin Inhibits Il-1 Beta-Induced Inflammation and Cartilage Matrix Degradation in Rat Chondrocytes. Eur. J. Pharmacol. 2019, 858, 7. [Google Scholar] [CrossRef]
- Chen, D.; Bellussi, L.M.; Cocca, S.; Wang, J.; Passali, G.C.; Ha, X.; Chew, L.; Passali, D. Glycyrrhetinic Acid Suppressed Hmgb1 Release by up-Regulation of Sirt6 in Nasal Inflammation. J. Biol. Regul. Homeost. Agents 2017, 31, 269–277. [Google Scholar]
- Li, X.P.; Meng, X.H.; Luo, K.; Luan, S.; Shi, X.L.; Cao, B.X.; Kong, J. The Identification of Micrornas Involved in the Response of Chinese Shrimp Fenneropenaeus Chinensis to White Spot Syndrome Virus Infection. Fish Shellfish Immunol. 2017, 68, 220–231. [Google Scholar] [CrossRef]
- Feng, S.W.; Luo, S.H.; Ji, C.C.; Shi, J. Mir-29c-3p Regulates Proliferation and Migration in Ovarian Cancer by Targeting Kif4a. World J. Surg. Oncol. 2020, 18, 8. [Google Scholar] [CrossRef]
- Chen, L.; Heikkinen, L.; Wang, C.L.; Yang, Y.; Sun, H.Y.; Wong, G. Trends in the Development of Mirna Bioinformatics Tools. Brief. Bioinform. 2019, 20, 1836–1852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, S.N.; Hu, S.; Shan, Y.P.; Li, L.Y.; Zhou, H.; Chen, J.S.; Yang, J.F.; Li, J.; Huang, Q.; Shen, C.P.; et al. Relevance Function of Microrna-708 in the Pathogenesis of Cancer. Cell. Signal. 2019, 63, 12. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Chen, X.M.; Zhong, J.; Ye, H.H.; Zhang, S.J.; Ge, D.Y.; Wang, X.D.; Wu, Y. The Effects of the Xijiao Dihuang Decoction Combined with Yinqiao Powder on Mirna-Mrna Profiles in Mice Infected with Influenza a Virus. BMC Complementary Med. Ther. 2020, 20, 19. [Google Scholar] [CrossRef] [PubMed]
- Song, K.H.; Kim, Y.H.; Kim, B.Y. Sho-Saiko-to, a Traditional Herbal Medicine, Regulates Gene Expression and Biological Function by Way of Micrornas in Primary Mouse Hepatocytes. BMC Complementary Altern. Med. 2014, 14, 16. [Google Scholar] [CrossRef] [Green Version]
- Zhou, S.Y.; Zhang, S.J.; Shen, H.Y.; Chen, W.; Xu, H.Z.; Chen, X.; Sun, D.W.; Zhong, S.L.; Zhao, J.H.; Tang, J.H. Curcumin Inhibits Cancer Progression through Regulating Expression of Micrornas. Tumor Biol. 2017, 39, 12. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.J.; Xiong, X.; Sui, Y.T. Isoliquiritigenin Attenuates Neuroinflammation in Traumatic Brain Injury in Young Rats. Neuroimmunomodulation 2019, 26, 102–109. [Google Scholar] [CrossRef]
- Zheng, Y.F.; Wei, J.H.; Fang, S.Q.; Tang, Y.P.; Cheng, H.B.; Wang, T.L.; Li, C.Y.; Peng, G.P. Hepatoprotective Triterpene Saponins from the Roots of Glycyrrhiza Inflata. Molecules 2015, 20, 6273–6283. [Google Scholar] [CrossRef]
- Yeh, C.F.; Wang, K.C.; Chiang, L.C.; Shieh, D.E.; Yen, M.H.; Chang, J.S. Water Extract of Licorice Had Anti-Viral Activity against Human Respiratory Syncytial Virus in Human Respiratory Tract Cell Lines. J. Ethnopharmacol. 2013, 148, 466–473. [Google Scholar]
- Khazraei-Moradian, S.; Ganjalikhani-Hakemi, M.; Andalib, A.; Yazdani, R.; Arasteh, J.; Kardar, G.A. The Effect of Licorice Protein Fractions on Proliferation and Apoptosis of Gastrointestinal Cancer Cell Lines. Nutr. Cancer 2017, 69, 330–339. [Google Scholar] [CrossRef]
- Lin, X.; Lin, Q. Mirna-495-3p Attenuates Tnf-Alpha Induced Apoptosis and Inflammation in Human Nucleus Pulposus Cells by Targeting Il5ra. Inflammation 2020, 43, 1797–1805. [Google Scholar] [CrossRef]
- Chen, J.Y.; Xu, L.F.; Hu, H.L.; Wen, Y.Q.; Chen, D.; Liu, W.H. Mirna-215-5p Alleviates the Metastasis of Prostate Cancer by Targeting Pgk1. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 639–646. [Google Scholar] [PubMed]
- Sur, S.; Steele, R.; Shi, X.Y.; Ray, R.B. Mirna-29b Inhibits Prostate Tumor Growth and Induces Apoptosis by Increasing Bim Expression. Cells 2019, 8, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.Y.; Pais, E.M.A.; Li, L.; Chen, J.R.; Miao, L.; Fordjour, P.A.; Fan, G.W. Microrna-155: A Novel Armamentarium against Inflammatory Diseases. Inflammation 2017, 40, 708–716. [Google Scholar]
- Liu, Y.; Liu, C.P.; Zhang, A.K.; Yin, S.C.; Wang, T.; Wang, Y.; Wang, M.M.; Liu, Y.X.; Ying, Q.H.; Sun, J.R.; et al. Down-Regulation of Long Non-Coding Rna Meg3 Suppresses Osteogenic Differentiation of Periodontal Ligament Stem Cells (Pdlscs) through Mir-27a-3p/Igf1 Axis in Periodontitis. Aging 2019, 11, 5334–5350. [Google Scholar] [CrossRef]
- Zhang, P.; Li, L.Q.; Zhang, D.; Shen, Y. Over-Expressed Mir-27a-3p Inhibits Inflammatory Response to Spinal Cord Injury by Decreasing Tlr4. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 5416–5423. [Google Scholar]
- Yao, X.L.; Tu, Y.; Xu, Y.L.; Guo, Y.Y.; Yao, F.; Zhang, X.H. Endoplasmic Reticulum Stress-Induced Exosomal Mir-27a-3p Promotes Immune Escape in Breast Cancer via Regulating Pd-L1 Expression in Macrophages. J. Cell. Mol. Med. 2020, 24, 9560–9573. [Google Scholar] [CrossRef]
- Fan, X.H.; Wang, J.Y.; Qin, T.T.; Zhang, Y.J.; Liu, W.T.; Jiang, K.T.; Huang, D.Z. Exosome Mir-27a-3p Secreted from Adipocytes Targets Icos to Promote Antitumor Immunity in Lung Adenocarcinoma. Thorac. Cancer 2020, 11, 1453–1464. [Google Scholar] [CrossRef]
- Dodson, L.F.; Boomer, J.S.; Deppong, C.M.; Shah, D.D.; Sim, J.; Bricker, T.L.; Russell, J.H.; Green, J.M. Targeted Knock-in Mice Expressing Mutations of Cd28 Reveal an Essential Pathway for Costimulation. Mol. Cell. Biol. 2009, 29, 3710–3721. [Google Scholar] [CrossRef] [Green Version]
- Friend, L.D.; Shah, D.D.; Deppong, C.; Lin, J.; Bricker, T.L.; Juehne, T.I.; Rose, C.M.; Green, J.M. A Dose-Dependent Requirement for the Proline Motif of Cd28 in Cellular and Humoral Immunity Revealed by a Targeted Knockin Mutant. J. Exp. Med. 2006, 203, 2121–2133. [Google Scholar] [CrossRef]
- van Wijk, F.; Nierkens, S.; de Jong, W.; Wehrens, E.J.M.; Boon, L.; van Kooten, P.; Knippels, U.M.J.; Pieters, R. The Cd28/Ctla-4-B7 Signaling Pathway Is Involved in Both Allergic Sensitization and Tolerance Induction to Orally Administered Peanut Proteins. J. Immunol. 2007, 178, 6894–6900. [Google Scholar] [CrossRef] [Green Version]
- Rozanski, C.H.; Utley, A.; Carlson, L.M.; Farren, M.R.; Murray, M.; Russell, L.M.; Nair, J.R.; Yang, Z.Y.; Brady, W.; Garrett-Sinha, L.A.; et al. Cd28 Promotes Plasma Cell Survival, Sustained Antibody Responses, and Blimp-1 Upregulation through Its Distal Pyap Proline Motif. J. Immunol. 2015, 194, 4717–4728. [Google Scholar] [CrossRef] [PubMed]
- Utley, A.; Chavel, C.; Lightman, S.; Holling, G.A.; Cooper, J.; Peng, P.; Liu, W.S.; Barwick, B.G.; Gavile, C.M.; Maguire, O.; et al. Cd28 Regulates Metabolic Fitness for Long-Lived Plasma Cell Survival. Cell Rep. 2020, 31, 21. [Google Scholar] [CrossRef] [PubMed]
- Yousefi, M.; Duplay, P. Cd28 Controls the Development of Innate-Like Cd8+ T Cells by Promoting the Functional Maturation of Nkt Cells. Eur. J. Immunol. 2013, 43, 3017–3027. [Google Scholar] [CrossRef] [PubMed]
- Boesteanu, A.C.; Katsikis, P.D. Memory T Cells Need Cd28 Costimulation to Remember. Semin. Immunol. 2009, 21, 69–77. [Google Scholar] [CrossRef] [Green Version]
- Greenfield, E.A.; Nguyen, K.A.; Kuchroo, V.K. Cd28/B7 Costimulation: A Review. Crit. Rev. Immunol. 1998, 18, 389–418. [Google Scholar] [CrossRef] [PubMed]
- Wakamatsu, E.; Omori, H.; Ohtsuka, S.; Ogawa, S.; Green, J.M.; Abe, R. Regulatory T Cell Subsets Are Differentially Dependent on Cd28 for Their Proliferation. Mol. Immunol. 2018, 101, 92–101. [Google Scholar] [CrossRef]
- Watanabe, M.; Lu, Y.; Breen, M.; Hodes, R.J. B7-Cd28 Co-Stimulation Modulates Central Tolerance via Thymic Clonal Deletion and Treg Generation through Distinct Mechanisms. Nat. Commun. 2020, 11, 14. [Google Scholar] [CrossRef]
- Jiang, R.C.; Gao, J.M.; Shen, J.Y.; Zhu, X.Q.; Wang, H.; Feng, S.Y.; Huang, C.; Shen, H.T.; Liu, H.L. Glycyrrhizic Acid Improves Cognitive Levels of Aging Mice by Regulating T/B Cell Proliferation. Front. Aging Neurosci. 2020, 12, 10. [Google Scholar] [CrossRef]
- Utsunomiya, T.; Kobayashi, M.; Herndon, D.N.; Pollard, R.B.; Suzuki, F. Glycyrrhizin (20 Beta-Carboxy-11-Oxo-30-Norolean-12-En-3 Beta-Yl-2-O-Beta-D-Glucopyranuronosyl-Alpha-D-Glucopyranosiduronic Acid) Improves the Resistance of Thermally Injured Mice to Opportunistic Infection of Herpes Simplex Virus Type 1. Immunol. Lett. 1995, 44, 59–66. [Google Scholar] [CrossRef]
- Fouladi, S.; Masjedi, M.; Ghasemi, R.; Hakemi, M.G.; Eskandari, N. The in Vitro Impact of Glycyrrhizic Acid on Cd4+T Lymphocytes through Ox40 Receptor in the Patients with Allergic Rhinitis. Inflammation 2018, 41, 1690–1701. [Google Scholar] [CrossRef]
- Guo, A.; He, D.M.; Xu, H.B.; Geng, C.A.; Zhao, J. Promotion of Regulatory T Cell Induction by Immunomodulatory Herbal Medicine Licorice and Its Two Constituents. Sci. Rep. 2015, 5, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, G.; Liang, G.; Zhang, Y.; Hu, J.; Zhou, C.; Li, J.; Zhang, W.; Shen, H.; Wu, F.; Tao, C.; et al. Involvement of MicroRNA-27a-3p in the Licorice-Induced Alteration of Cd28 Expression in Mice. Genes 2022, 13, 1143. https://doi.org/10.3390/genes13071143
Feng G, Liang G, Zhang Y, Hu J, Zhou C, Li J, Zhang W, Shen H, Wu F, Tao C, et al. Involvement of MicroRNA-27a-3p in the Licorice-Induced Alteration of Cd28 Expression in Mice. Genes. 2022; 13(7):1143. https://doi.org/10.3390/genes13071143
Chicago/Turabian StyleFeng, Gang, Guozheng Liang, Yaqian Zhang, Jicong Hu, Chuandong Zhou, Jiawen Li, Wenfeng Zhang, Han Shen, Fenglin Wu, Changli Tao, and et al. 2022. "Involvement of MicroRNA-27a-3p in the Licorice-Induced Alteration of Cd28 Expression in Mice" Genes 13, no. 7: 1143. https://doi.org/10.3390/genes13071143