
Coat Color-Facilitated Efficient Generation and Analysis of a Mouse Model of Down Syndrome Triplicated for All Human Chromosome 21 Orthologous Regions


Abstract
:1. Introduction
2. Materials and Methods
2.1. Mice
2.2. Open Field Test
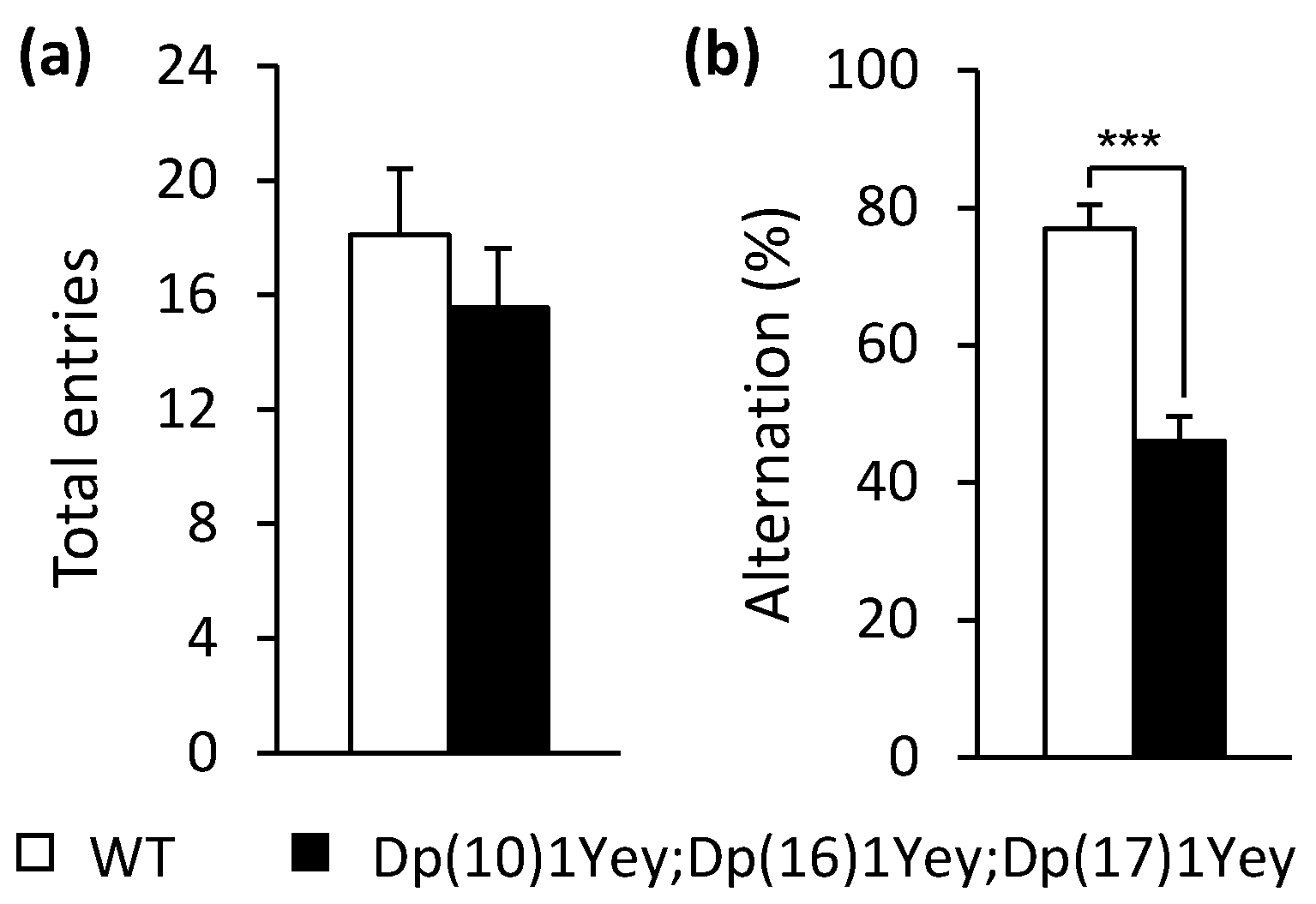
2.3. T-Maze Spontaneous Alternation Test
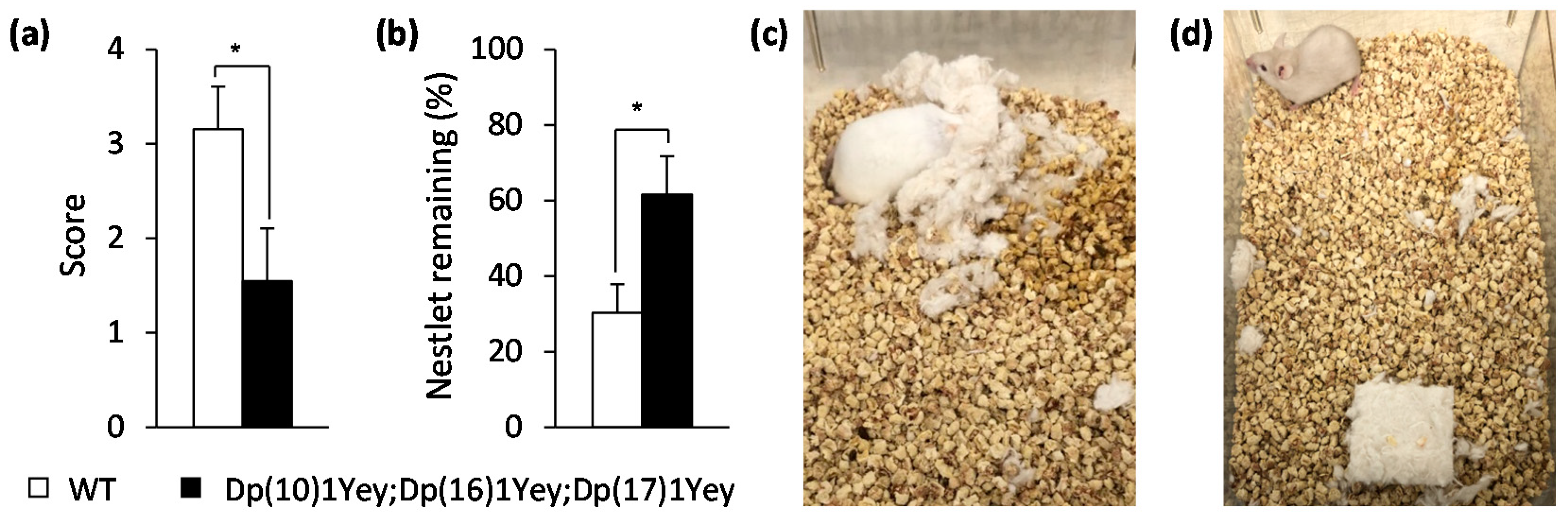
2.4. Nesting Test
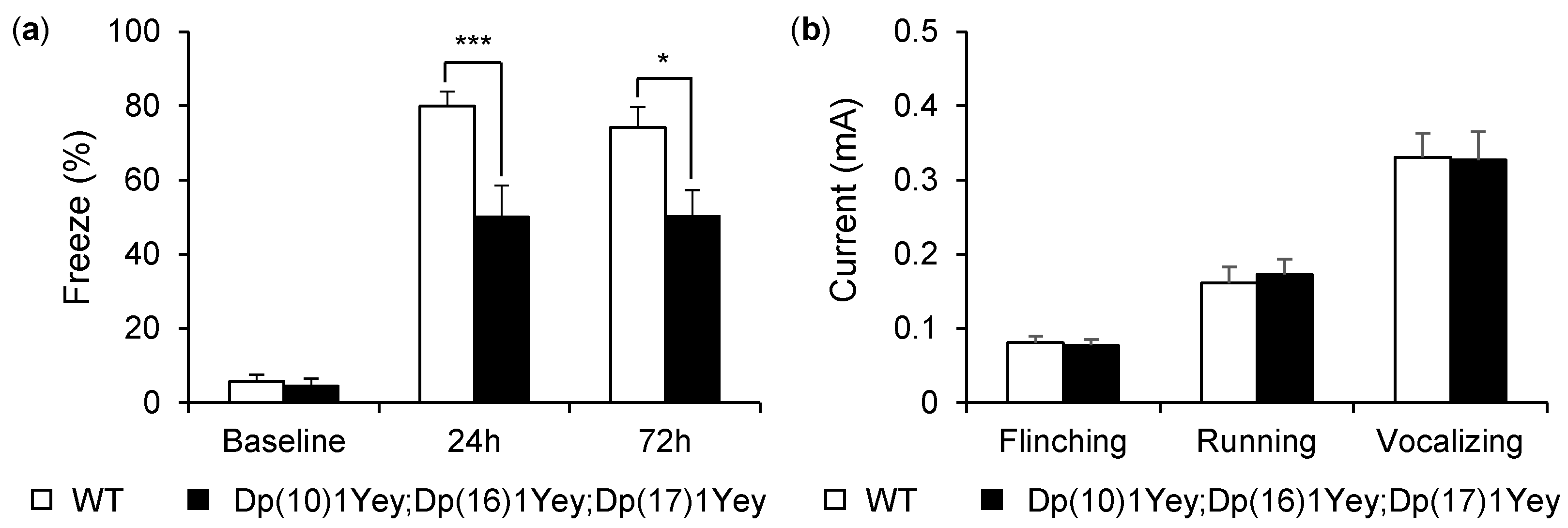
2.5. Contextual Fear-Conditioning Test
2.6. Foot-Shock Sensitivity Test
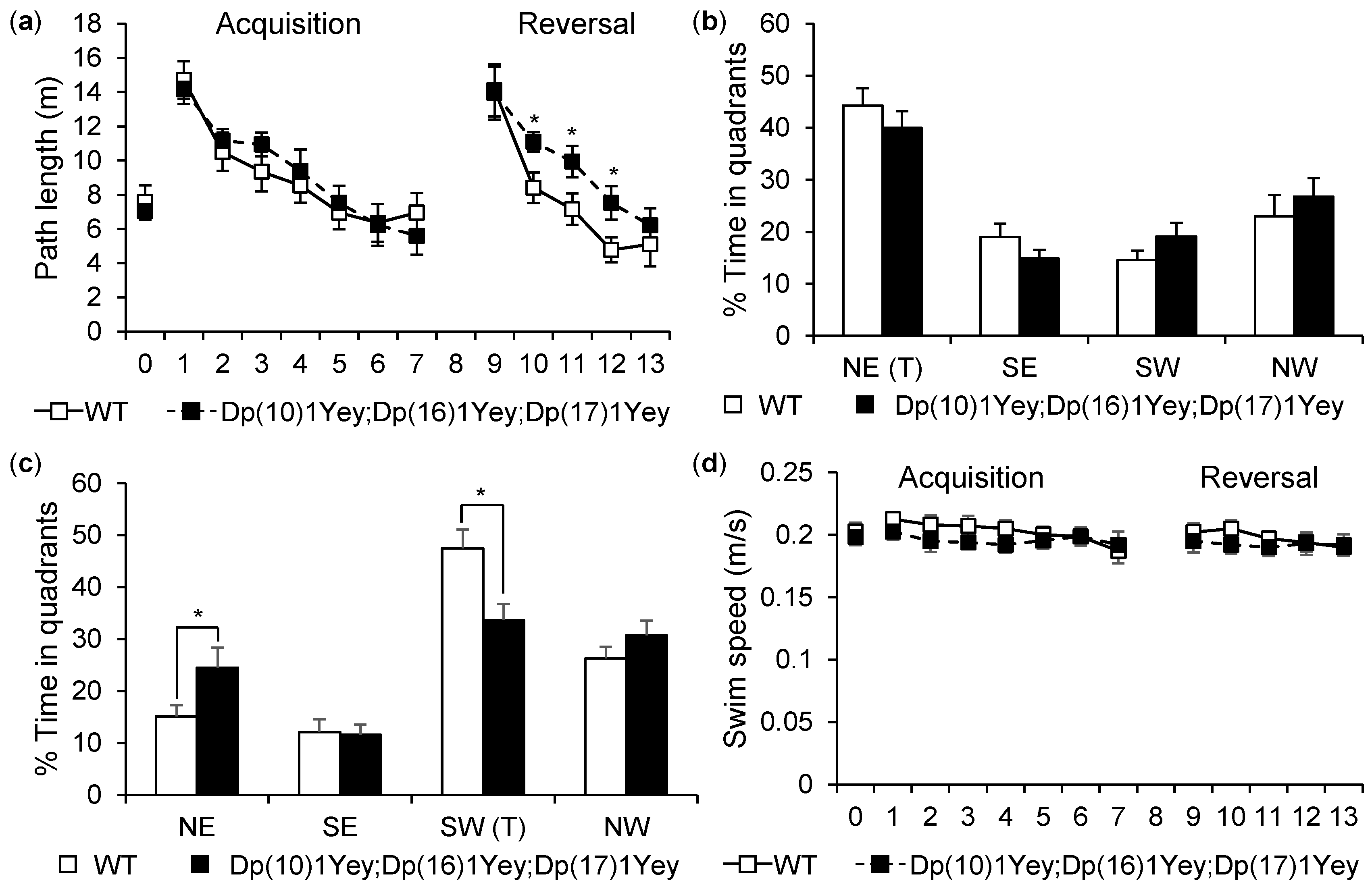
2.7. Morris Water Maze Test
2.8. Sample Size and Statistical Methods
3. Results
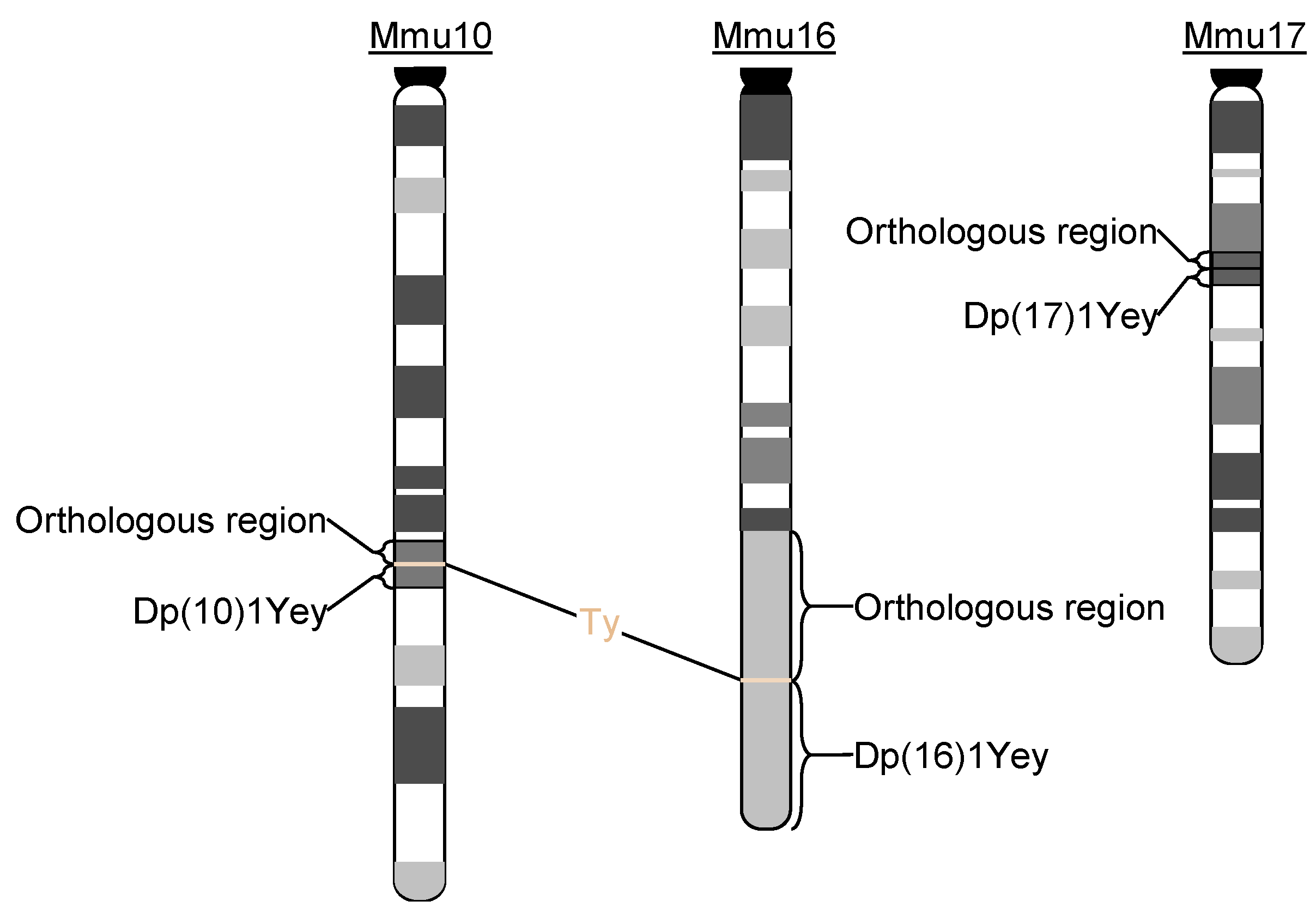
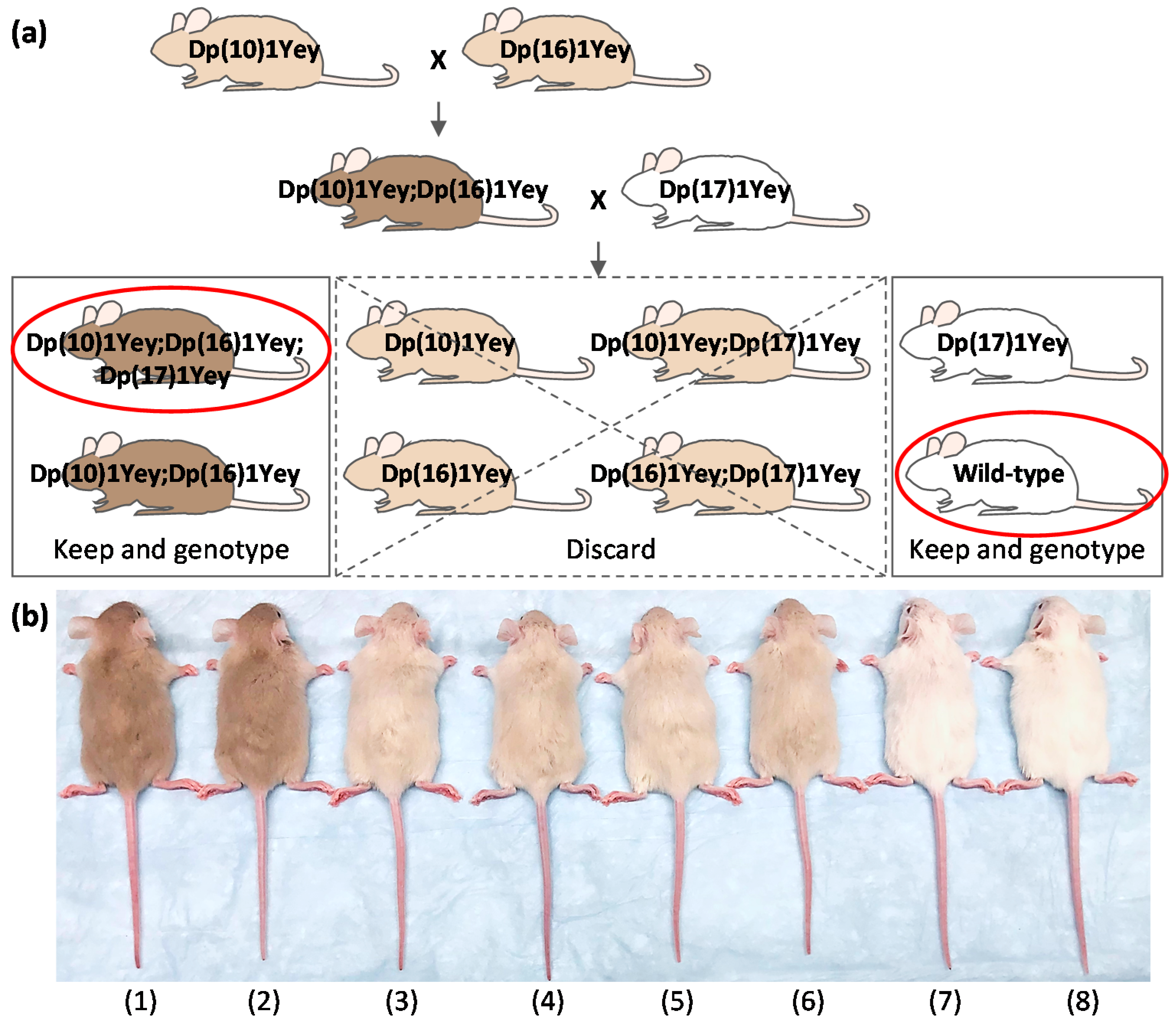
3.1. Efficient Generation of Dp(10)1Yey;Dp(16)1Yey;Dp(17)1Yey Mice Facilitated by Coat Colors
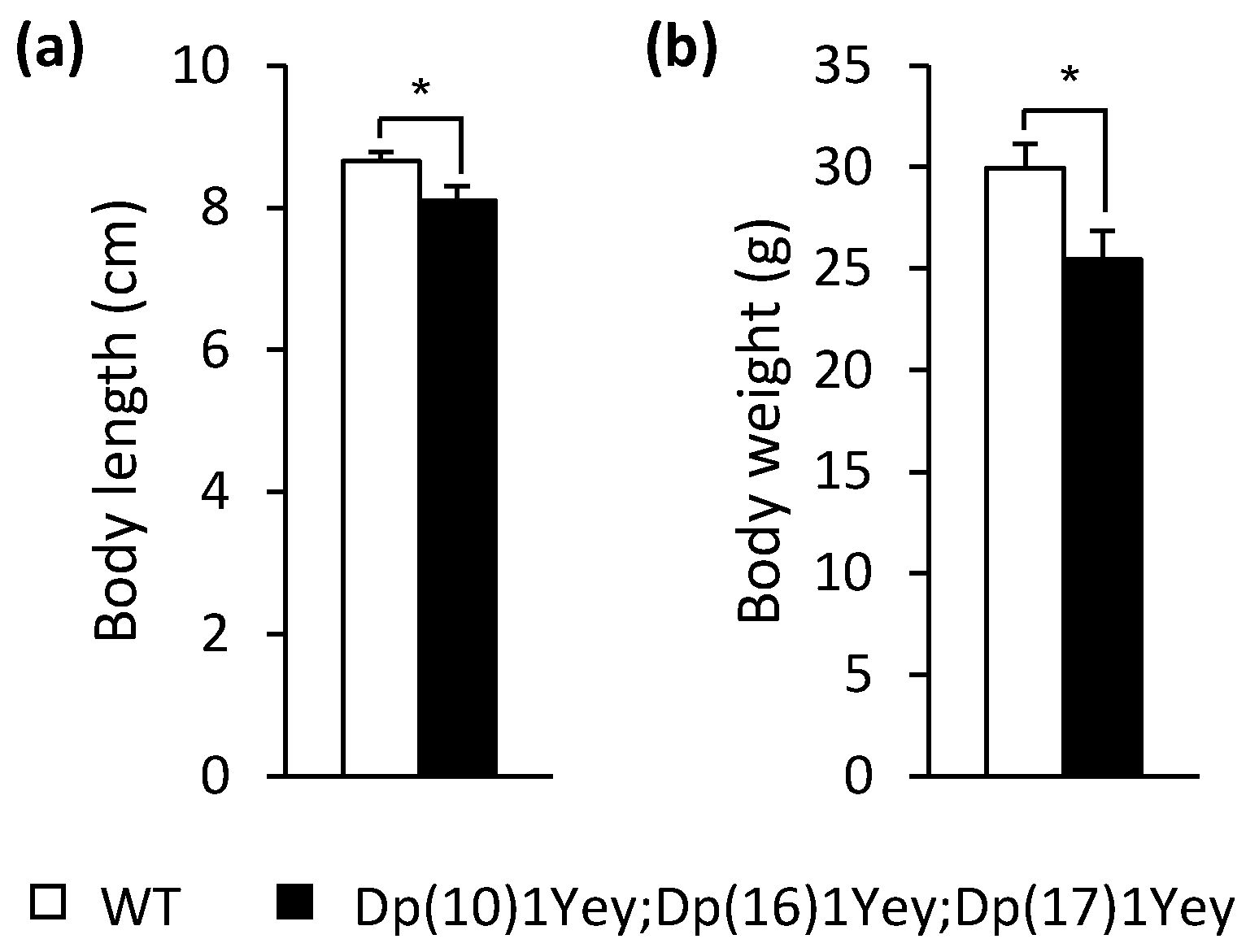
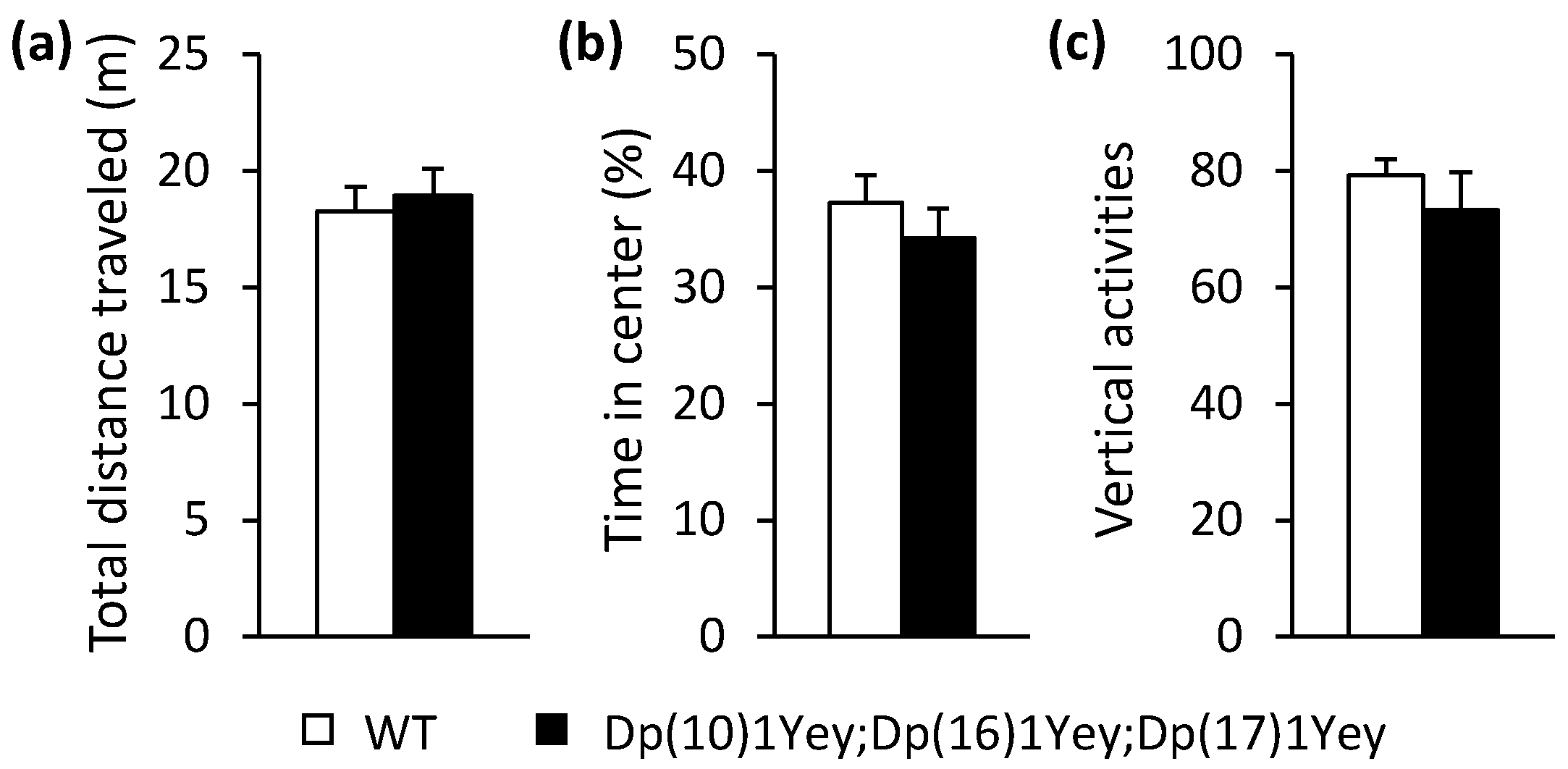
3.2. Dp(10)1Yey;Dp(16)1Yey;Dp(17)1Yey Mice Identified via the Coat Color-Facilitated Strategy Exhibit Cognitive Deficits
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bull, M.J. Down syndrome. N. Engl. J. Med. 2020, 382, 2344–2352. [Google Scholar] [CrossRef]
- Antonarakis, S.E.; Skotko, B.G.; Rafii, M.S.; Strydom, A.; Pape, S.E.; Bianchi, D.W.; Sherman, S.L.; Reeves, R.H. Down syndrome. Nat. Rev. Dis. Primers 2020, 6, 9. [Google Scholar] [CrossRef] [PubMed]
- Dierssen, M. Down syndrome: The brain in trisomic mode. Nat. Rev. Neurosci. 2012, 13, 844–858. [Google Scholar] [CrossRef]
- Korenberg, J.R.; Chen, X.N.; Schipper, R.; Sun, Z.; Gonsky, R.; Gerwehr, S.; Carpenter, N.; Daumer, C.; Dignan, P.; Disteche, C.; et al. Down syndrome phenotypes: The consequences of chromosomal imbalance. Proc. Natl. Acad. Sci. USA 1994, 91, 4997–5001. [Google Scholar] [CrossRef] [Green Version]
- Davisson, M.T.; Schmidt, C.; Akeson, E.C. Segmental trisomy of murine chromosome 16: A new model system for studying Down syndrome. Prog. Clin. Biol. Res. 1990, 360, 263–280. [Google Scholar] [PubMed]
- Reeves, R.H.; Irving, N.G.; Moran, T.H.; Wohn, A.; Kitt, C.; Sisodia, S.S.; Schmidt, C.; Bronson, R.T.; Davisson, M.T. A mouse model for Down syndrome exhibits learning and behaviour deficits. Nat. Genet. 1995, 11, 177–184. [Google Scholar] [CrossRef]
- Sago, H.; Carlson, E.J.; Smith, D.J.; Kilbridge, J.; Rubin, E.M.; Mobley, W.C.; Epstein, C.J.; Huang, T.T. Ts1Cje, a partial trisomy 16 mouse model for Down syndrome, exhibits learning and behavioral abnormalities. Proc. Natl. Acad. Sci. USA 1998, 95, 6256–6261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xing, Z.; Li, Y.; Pao, A.; Bennett, A.S.; Tycko, B.; Mobley, W.C.; Yu, Y.E. Mouse-based genetic modeling and analysis of Down syndrome. Br. Med. Bull. 2016, 120, 111–122. [Google Scholar] [CrossRef] [Green Version]
- Duchon, A.; Raveau, M.; Chevalier, C.; Nalesso, V.; Sharp, A.J.; Herault, Y. Identification of the translocation breakpoints in the Ts65Dn and Ts1Cje mouse lines: Relevance for modeling Down syndrome. Mamm. Genome 2011, 22, 674–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reinholdt, L.G.; Ding, Y.; Gilbert, G.J.; Czechanski, A.; Solzak, J.P.; Roper, R.J.; Johnson, M.T.; Donahue, L.R.; Lutz, C.; Davisson, M.T. Molecular characterization of the translocation breakpoints in the Down syndrome mouse model Ts65Dn. Mamm. Genome 2011, 22, 685–691. [Google Scholar] [CrossRef] [Green Version]
- Kazuki, Y.; Gao, F.J.; Li, Y.; Moyer, A.J.; Devenney, B.; Hiramatsu, K.; Miyagawa-Tomita, S.; Abe, S.; Kazuki, K.; Kajitani, N.; et al. A non-mosaic transchromosomic mouse model of down syndrome carrying the long arm of human chromosome 21. eLife 2020, 9, e56223. [Google Scholar] [CrossRef] [PubMed]
- O’Doherty, A.; Ruf, S.; Mulligan, C.; Hildreth, V.; Errington, M.L.; Cooke, S.; Sesay, A.; Modino, S.; Vanes, L.; Hernandez, D.; et al. An aneuploid mouse strain carrying human chromosome 21 with Down syndrome phenotypes. Science 2005, 309, 2033–2037. [Google Scholar] [CrossRef] [Green Version]
- Gribble, S.M.; Wiseman, F.K.; Clayton, S.; Prigmore, E.; Langley, E.; Yang, F.; Maguire, S.; Fu, B.; Rajan, D.; Sheppard, O.; et al. Massively parallel sequencing reveals the complex structure of an irradiated human chromosome on a mouse background in the Tc1 model of Down syndrome. PLoS ONE 2013, 8, e60482. [Google Scholar] [CrossRef]
- Reeves, R.H. Down syndrome mouse models are looking up. Trends Mol. Med. 2006, 12, 237–240. [Google Scholar] [CrossRef]
- Herault, Y.; Delabar, J.M.; Fisher, E.M.C.; Tybulewicz, V.L.J.; Yu, E.; Brault, V. Rodent models in Down syndrome research: Impact and future opportunities. Dis. Model. Mech. 2017, 10, 1165–1186. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Yu, T.; Morishima, M.; Pao, A.; LaDuca, J.; Conroy, J.; Nowak, N.; Matsui, S.; Shiraishi, I.; Yu, Y. Duplication of the entire 22.9-Mb human chromosome 21 syntenic region on mouse chromosome 16 causes cardiovascular and gastrointestinal abnormalities. Hum. Mol. Genet. 2007, 16, 1359–1366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olson, L.E.; Richtsmeier, J.T.; Leszl, J.; Reeves, R.H. A chromosome 21 critical region does not cause specific Down syndrome phenotypes. Science 2004, 306, 687–690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, P.L.; Magnol, L.; Sahun, I.; Brault, V.; Duchon, A.; Prandini, P.; Gruart, A.; Bizot, J.C.; Chadefaux-Vekemans, B.; Deutsch, S.; et al. A new mouse model for the trisomy of the Abcg1-U2af1 region reveals the complexity of the combinatorial genetic code of down syndrome. Hum. Mol. Genet. 2009, 18, 4756–4769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watson-Scales, S.; Kalmar, B.; Lana-Elola, E.; Gibbins, D.; La Russa, F.; Wiseman, F.; Williamson, M.; Saccon, R.; Slender, A.; Olerinyova, A.; et al. Analysis of motor dysfunction in Down Syndrome reveals motor neuron degeneration. PLoS Genet. 2018, 14, e1007383. [Google Scholar] [CrossRef]
- Yu, T.; Li, Z.; Jia, Z.; Clapcote, S.J.; Liu, C.; Li, S.; Asrar, S.; Pao, A.; Chen, R.; Fan, N.; et al. A mouse model of Down syndrome trisomic for all human chromosome 21 syntenic regions. Hum. Mol. Genet. 2010, 19, 2780–2791. [Google Scholar] [CrossRef] [Green Version]
- Duchon, A.; Del Mar Muniz Moreno, M.; Lorenzo, S.M.; de Souza, M.P.S.; Chevalier, C.; Nalesso, V.; Meziane, H.; de Sousa, P.L.; Noblet, V.; Armspach, J.P.; et al. Multi-influential genetic interactions alter behaviour and cognition through six main biological cascades in Down syndrome mouse models. Hum. Mol. Genet. 2021. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Liu, C.; Yu, T.; Zhang, L.; Meng, K.; Xing, Z.; Belichenko, P.V.; Kleschevnikov, A.M.; Pao, A.; Peresie, J.; et al. Genetic dissection of the Down syndrome critical region. Hum. Mol. Genet. 2015, 24, 6540–6551. [Google Scholar] [CrossRef] [PubMed]
- Ma’ayan, A.; Gardiner, K.; Iyengar, R. The cognitive phenotype of Down syndrome: Insights from intracellular network analysis. NeuroRx 2006, 3, 396–406. [Google Scholar] [CrossRef] [PubMed]
- Gupta, M.; Dhanasekaran, A.R.; Gardiner, K.J. Mouse models of Down syndrome: Gene content and consequences. Mamm. Genome 2016, 27, 538–555. [Google Scholar] [CrossRef]
- Deacon, R.M.; Rawlins, J.N. T-maze alternation in the rodent. Nat. Protoc. 2006, 1, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Deacon, R.M. Assessing nest building in mice. Nat. Protoc. 2006, 1, 1117–1119. [Google Scholar] [CrossRef]
- Clapcote, S.J.; Lazar, N.L.; Bechard, A.R.; Roder, J.C. Effects of the rd1 mutation and host strain on hippocampal learning in mice. Behav. Genet. 2005, 35, 591–601. [Google Scholar] [CrossRef]
- Clapcote, S.J.; Roder, J.C. Survey of embryonic stem cell line source strains in the water maze reveals superior reversal learning of 129S6/SvEvTac mice. Behav. Brain Res. 2004, 152, 35–48. [Google Scholar] [CrossRef]
- D’Hooge, R.; Nagels, G.; Franck, F.; Bakker, C.E.; Reyniers, E.; Storm, K.; Kooy, R.F.; Oostra, B.A.; Willems, P.J.; De Deyn, P.P. Mildly impaired water maze performance in male Fmr1 knockout mice. Neuroscience 1997, 76, 367–376. [Google Scholar] [CrossRef]
- Yu, T.; Clapcote, S.J.; Li, Z.; Liu, C.; Pao, A.; Bechard, A.R.; Carattini-Rivera, S.; Matsui, S.; Roder, J.C.; Baldini, A.; et al. Deficiencies in the region syntenic to human 21q22.3 cause cognitive deficits in mice. Mamm. Genome 2010, 21, 258–267. [Google Scholar] [CrossRef] [Green Version]
- Brown, W.P.; Morgan, K.T. Age, breed designation, coat color, and coat pattern influenced the length of stay of cats at a no-kill shelter. J. Appl. Anim. Welf. Sci. 2015, 18, 169–180. [Google Scholar] [CrossRef]
- Buggia, L.B. Questions about coat color and aggression in cats. J. Am. Vet. Med. Assoc. 2011, 239, 1289. [Google Scholar]
- Jacobs, L.N.; Staiger, E.A.; Albright, J.D.; Brooks, S.A. The MC1R and ASIP Coat Color Loci May Impact Behavior in the Horse. J. Hered. 2016, 107, 214–219. [Google Scholar] [CrossRef] [PubMed]
- Perez-Guisado, J.; Munoz-Serrano, A.; Lopez-Rodriguez, R. Evaluation of the Campbell test and the influence of age, sex, breed, and coat color on puppy behavioral responses. Can. J. Vet. Res. 2008, 72, 269–277. [Google Scholar]
- Stelow, E.A.; Bain, M.J.; Kass, P.H. The Relationship Between Coat Color and Aggressive Behaviors in the Domestic Cat. J. Appl. Anim. Welf. Sci. 2016, 19, 1–15. [Google Scholar] [CrossRef]
- Turner, J.W., Jr.; Carbonell, C. A relationship between frequency of display of territorial marking behavior and coat color in male Mongolian gerbils. Lab. Anim. Sci. 1984, 34, 488–490. [Google Scholar] [PubMed]
- Webb, A.A.; Cullen, C.L. Coat color and coat color pattern-related neurologic and neuro-ophthalmic diseases. Can. Vet. J. 2010, 51, 653–657. [Google Scholar]
- Yamamuro, Y.; Shiraishi, A. Genotype-dependent participation of coat color gene loci in the behavioral traits of laboratory mice. Behav. Process. 2011, 88, 81–87. [Google Scholar] [CrossRef]
- Lu, Y.M.; Jia, Z.; Janus, C.; Henderson, J.T.; Gerlai, R.; Wojtowicz, J.M.; Roder, J.C. Mice lacking metabotropic glutamate receptor 5 show impaired learning and reduced CA1 long-term potentiation (LTP) but normal CA3 LTP. J. Neurosci. 1997, 17, 5196–5205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marechal, D.; Lopes Pereira, P.; Duchon, A.; Herault, Y. Dosage of the Abcg1-U2af1 region modifies locomotor and cognitive deficits observed in the Tc1 mouse model of Down syndrome. PLoS ONE 2015, 10, e0115302. [Google Scholar] [CrossRef] [Green Version]
- Sturgeon, X.; Le, T.; Ahmed, M.M.; Gardiner, K.J. Pathways to cognitive deficits in Down syndrome. Prog. Brain Res. 2012, 197, 73–100. [Google Scholar] [CrossRef]
- Zhang, L.; Meng, K.; Jiang, X.; Liu, C.; Pao, A.; Belichenko, P.V.; Kleschevnikov, A.M.; Josselyn, S.; Liang, P.; Ye, P.; et al. Human chromosome 21 orthologous region on mouse chromosome 17 is a major determinant of Down syndrome-related developmental cognitive deficits. Hum. Mol. Genet. 2014, 23, 578–589. [Google Scholar] [CrossRef] [Green Version]
- Threadgill, D.W.; Dlugosz, A.A.; Hansen, L.A.; Tennenbaum, T.; Lichti, U.; Yee, D.; LaMantia, C.; Mourton, T.; Herrup, K.; Harris, R.C.; et al. Targeted disruption of mouse EGF receptor: Effect of genetic background on mutant phenotype. Science 1995, 269, 230–234. [Google Scholar] [CrossRef] [PubMed]
- Genik, P.C.; Bielefeldt-Ohmann, H.; Liu, X.; Story, M.D.; Ding, L.; Bush, J.M.; Fallgren, C.M.; Weil, M.M. Strain background determines lymphoma incidence in Atm knockout mice. Neoplasia 2014, 16, 129–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, K.R.; Zheng, Q.Y.; Noben-Trauth, K. Strain background effects and genetic modifiers of hearing in mice. Brain Res. 2006, 1091, 79–88. [Google Scholar] [CrossRef] [Green Version]
- Han, H.J.; Allen, C.C.; Buchovecky, C.M.; Yetman, M.J.; Born, H.A.; Marin, M.A.; Rodgers, S.P.; Song, B.J.; Lu, H.C.; Justice, M.J.; et al. Strain background influences neurotoxicity and behavioral abnormalities in mice expressing the tetracycline transactivator. J. Neurosci. 2012, 32, 10574–10586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailey, R.M.; Howard, J.; Knight, J.; Sahara, N.; Dickson, D.W.; Lewis, J. Effects of the C57BL/6 strain background on tauopathy progression in the rTg4510 mouse model. Mol. Neurodegener. 2014, 9, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, S.; Sanford, U.R.; Vargas, J.C.; Xu, H.; Groen, A.; Paulusma, C.C.; Grenert, J.P.; Pawlikowska, L.; Sen, S.; Elferink, R.P.; et al. Strain background modifies phenotypes in the ATP8B1-deficient mouse. PLoS ONE 2010, 5, e8984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, S.W.; Roberts, R.A.; Robbins, G.R.; Perry, J.L.; Kai, M.P.; Chen, K.; Bo, T.; Napier, M.E.; Ting, J.P.; Desimone, J.M.; et al. Nanoparticle clearance is governed by Th1/Th2 immunity and strain background. J. Clin. Investig. 2013, 123, 3061–3073. [Google Scholar] [CrossRef] [Green Version]
- Carvalhaes, L.S.; Gervasio, O.L.; Guatimosim, C.; Heljasvaara, R.; Sormunen, R.; Pihlajaniemi, T.; Kitten, G.T. Collagen XVIII/endostatin is associated with the epithelial-mesenchymal transformation in the atrioventricular valves during cardiac development. Dev. Dyn. 2006, 235, 132–142. [Google Scholar] [CrossRef]
- Izzo, A.; Manco, R.; Bonfiglio, F.; Cali, G.; De Cristofaro, T.; Patergnani, S.; Cicatiello, R.; Scrima, R.; Zannini, M.; Pinton, P.; et al. NRIP1/RIP140 siRNA-mediated attenuation counteracts mitochondrial dysfunction in Down syndrome. Hum. Mol. Genet. 2014, 23, 4406–4419. [Google Scholar] [CrossRef] [PubMed]
- Izzo, A.; Nitti, M.; Mollo, N.; Paladino, S.; Procaccini, C.; Faicchia, D.; Cali, G.; Genesio, R.; Bonfiglio, F.; Cicatiello, R.; et al. Metformin restores the mitochondrial network and reverses mitochondrial dysfunction in Down syndrome cells. Hum. Mol. Genet. 2017, 26, 1056–1069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jagadeesh, A.; Maroun, L.E.; Van Es, L.M.; Millis, R.M. Autoimmune Mechanisms of Interferon Hypersensitivity and Neurodegenerative Diseases: Down Syndrome. Autoimmune Dis. 2020, 2020, 6876920. [Google Scholar] [CrossRef] [PubMed]
- Karousou, E.; Stachtea, X.; Moretto, P.; Viola, M.; Vigetti, D.; D’Angelo, M.L.; Raio, L.; Ghezzi, F.; Pallotti, F.; De Luca, G.; et al. New insights into the pathobiology of Down syndrome--hyaluronan synthase-2 overexpression is regulated by collagen VI alpha2 chain. FEBS J. 2013, 280, 2418–2430. [Google Scholar] [CrossRef]
- Locke, A.E.; Dooley, K.J.; Tinker, S.W.; Cheong, S.Y.; Feingold, E.; Allen, E.G.; Freeman, S.B.; Torfs, C.P.; Cua, C.L.; Epstein, M.P.; et al. Variation in folate pathway genes contributes to risk of congenital heart defects among individuals with Down syndrome. Genet. Epidemiol. 2010, 34, 613–623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheng, L.; Leshchyns’ka, I.; Sytnyk, V. Neural Cell Adhesion Molecule 2 (NCAM2)-Induced c-Src-Dependent Propagation of Submembrane Ca2+ Spikes Along Dendrites Inhibits Synapse Maturation. Cereb. Cortex 2019, 29, 1439–1459. [Google Scholar] [CrossRef]
- Zheng, Q.; Li, G.; Wang, S.; Zhou, Y.; Liu, K.; Gao, Y.; Zhou, Y.; Zheng, L.; Zhu, L.; Deng, Q.; et al. Trisomy 21-induced dysregulation of microglial homeostasis in Alzheimer’s brains is mediated by USP25. Sci. Adv. 2021, 7. [Google Scholar] [CrossRef]
- Aziz, N.M.; Guedj, F.; Pennings, J.L.A.; Olmos-Serrano, J.L.; Siegel, A.; Haydar, T.F.; Bianchi, D.W. Lifespan analysis of brain development, gene expression and behavioral phenotypes in the Ts1Cje, Ts65Dn and Dp(16)1/Yey mouse models of Down syndrome. Dis. Model. Mech. 2018, 11. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype Group | (1) | (2) | (3) | (4) | (5) | (6) | (7) | (8) |
|---|---|---|---|---|---|---|---|---|
| Coat colors | Tan | Tan | Cream | Cream | Cream | Cream | White | White |
| Progeny # | 23 | 25 | 31 | 23 | 28 | 24 | 34 | 31 |
| Ratio | 10.50% | 11.42% | 14.16% | 10.50% | 12.79% | 10.96% | 15.53% | 14.16% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Xing, Z.; Yu, T.; Pao, A.; Daadi, M.; Yu, Y.E. Coat Color-Facilitated Efficient Generation and Analysis of a Mouse Model of Down Syndrome Triplicated for All Human Chromosome 21 Orthologous Regions. Genes 2021, 12, 1215. https://doi.org/10.3390/genes12081215
Li Y, Xing Z, Yu T, Pao A, Daadi M, Yu YE. Coat Color-Facilitated Efficient Generation and Analysis of a Mouse Model of Down Syndrome Triplicated for All Human Chromosome 21 Orthologous Regions. Genes. 2021; 12(8):1215. https://doi.org/10.3390/genes12081215
Chicago/Turabian StyleLi, Yichen, Zhuo Xing, Tao Yu, Annie Pao, Marcel Daadi, and Y. Eugene Yu. 2021. "Coat Color-Facilitated Efficient Generation and Analysis of a Mouse Model of Down Syndrome Triplicated for All Human Chromosome 21 Orthologous Regions" Genes 12, no. 8: 1215. https://doi.org/10.3390/genes12081215