The Profiles of Low Molecular Nitrogen Compounds and Fatty Acids in Wort and Beer Obtained with the Addition of Quinoa (Chenopodium quinoa Willd.), Amaranth (Amaranthus cruentus L.) or Maltose Syrup

 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Raw Materials
2.2. Yeasts
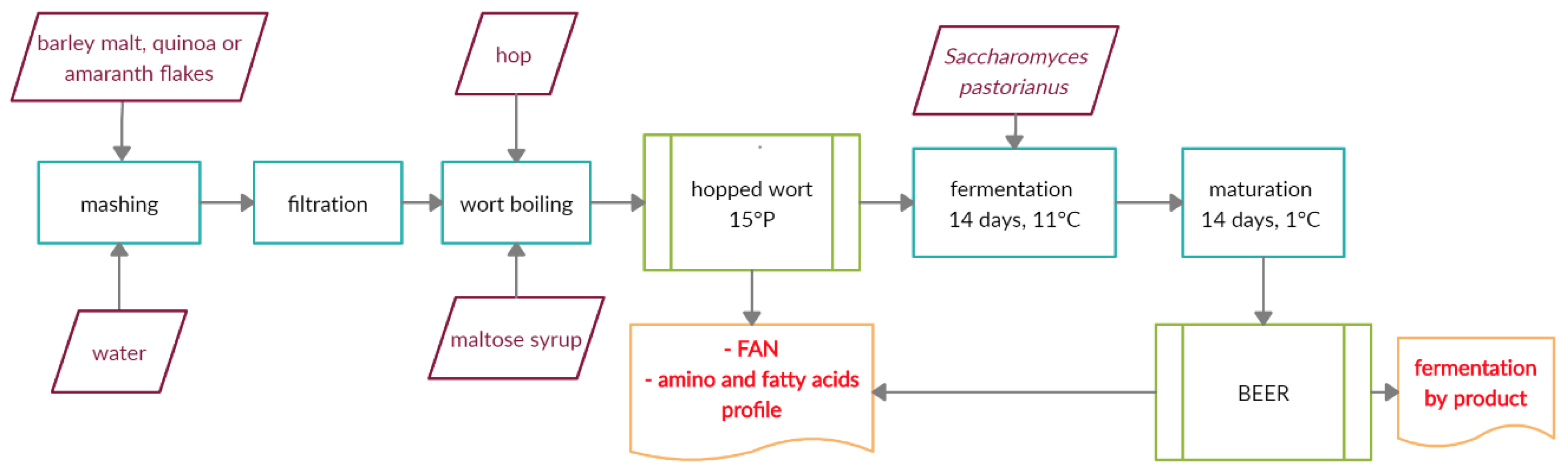
2.3. The Production of Worts and Beers
2.4. The Fatty Acids Determination in Worts and Beers
2.5. Determination of Low Molecular Nitrogen Compounds and Amino Acids
2.6. Determination of Fermentation by Products
2.7. Statistics
3. Results and Discussion
3.1. Fatty Acid Profiles in Worts
3.2. Fatty Acid Profiles in Beers
3.3. The Content of Free Amino Nitrogen in Worts and Beers Supplemented with Pseudocereals
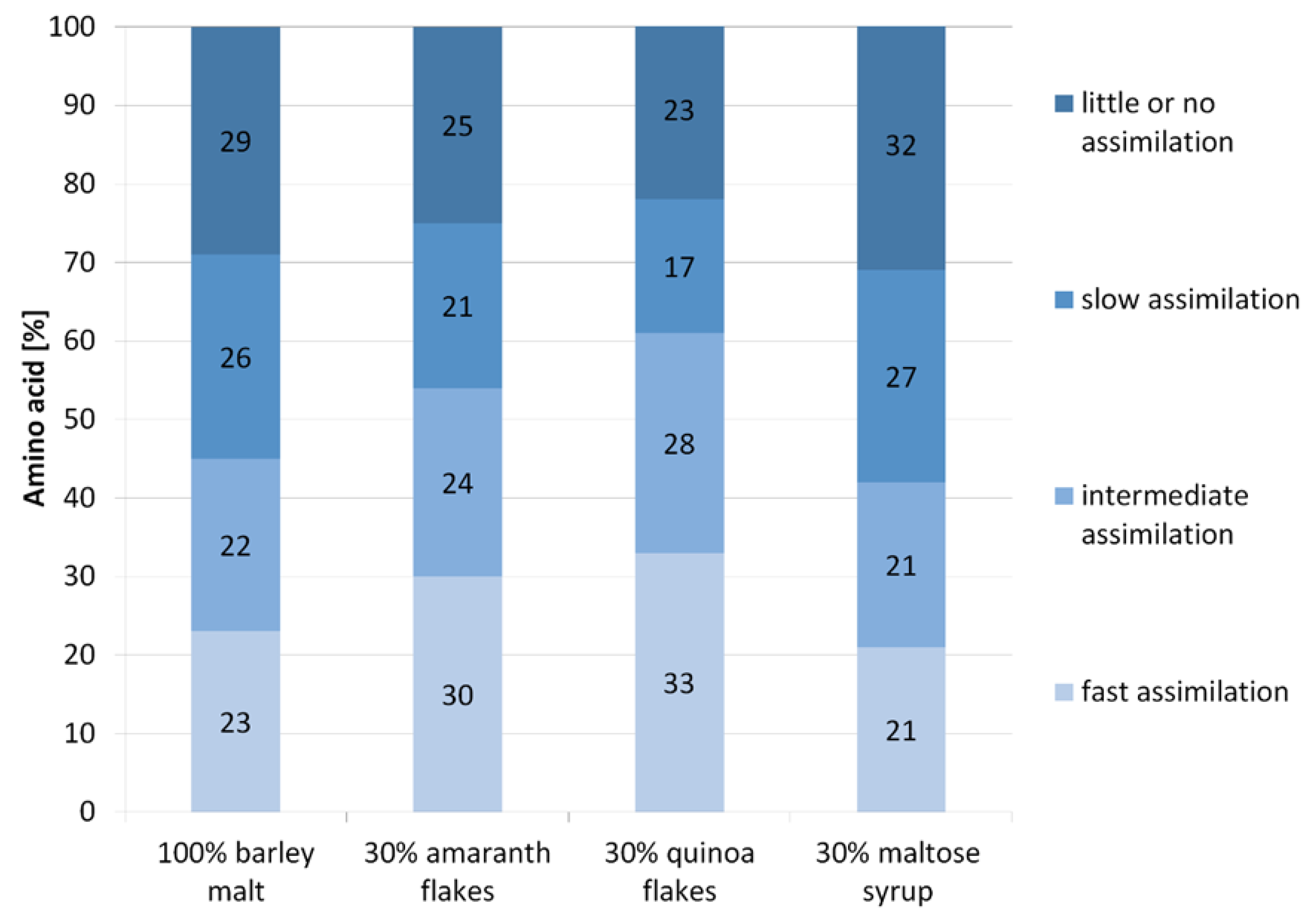
3.4. The Amino Acids Profile in Wort Obtained with Addition of Adjuncts
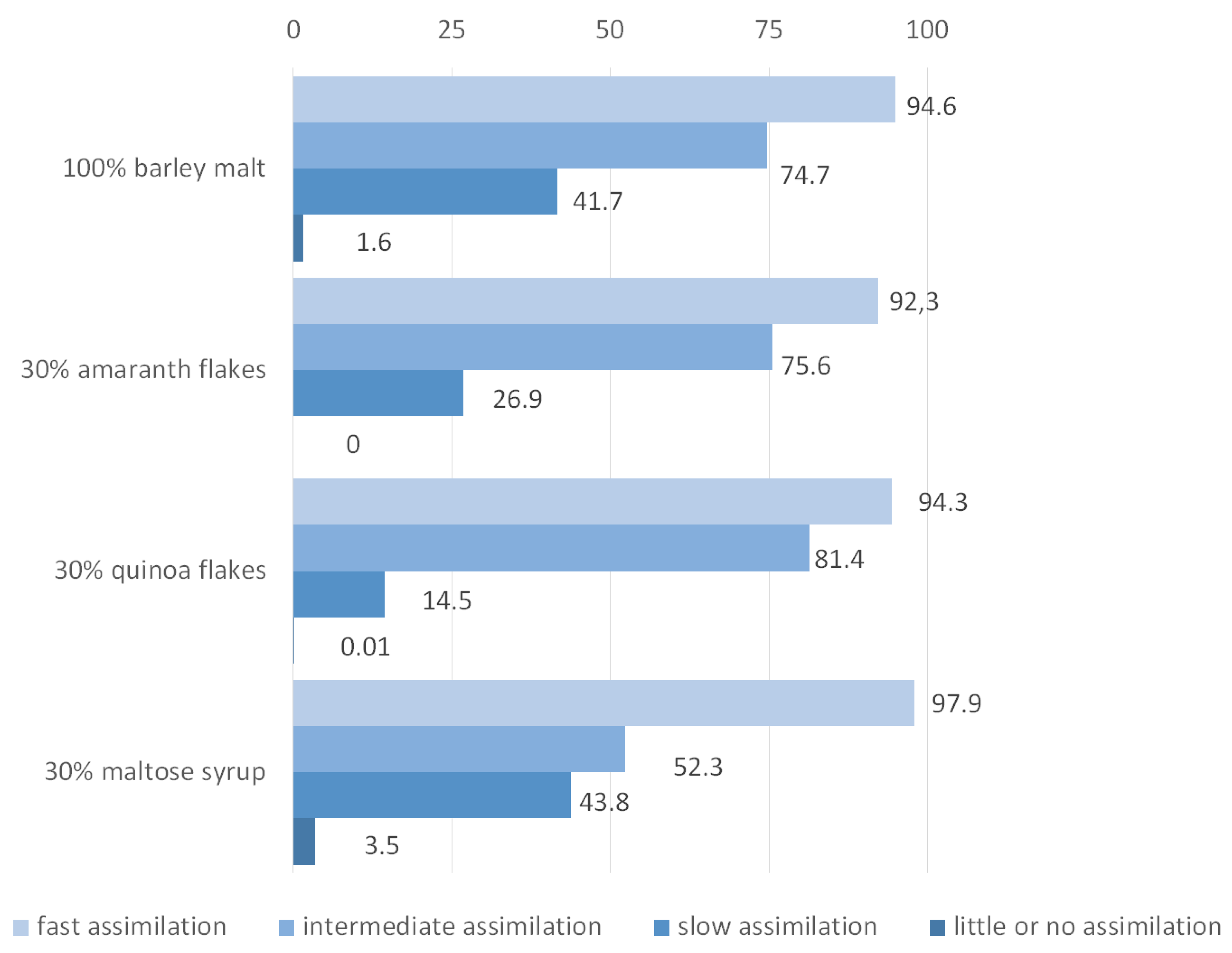
3.5. The Use of Amino Acids by Yeast during Fermentation
3.6. The Effect of Amino Acid Profile in Worts on the Synthesis of Fermentation Byproducts by Yeast
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Stewart, G.G.; Hill, A.E.; Russell, I. 125th anniversary review: Developments in brewing and distilling yeast strains. J. Inst. Brew. 2013, 119, 202–220. [Google Scholar] [CrossRef]
- Ferreira, I.F.M.; Guido, L.F. Impact of wort amino acids on beer flavour: A review. Fermentation 2018, 4, 23. [Google Scholar] [CrossRef] [Green Version]
- Vanderhaegen, B.; Neven, H.; Verachtert, H.; Derdelinckx, G. The chemistry of beer aging-A critical review. Food Chem. 2009, 95, 357–381. [Google Scholar] [CrossRef]
- Bravi, E.; Benedetti, P.; Marconi, O.; Perretti, G. Determination of free fatty acids in beer wort. Food Chem. 2014, 151, 374–378. [Google Scholar] [CrossRef] [PubMed]
- Lei, H.; Zhao, H.; Yu, Z.; Zhao, M. Effects of wort gravity and nitrogen level on fermentation performance of brewer’s yeast and the formation of flavor volatiles. Appl. Biochem. Biotechnol. 2012, 166, 1562–1574. [Google Scholar] [CrossRef] [PubMed]
- Santos Mathias, T.R.; de Mello, P.P.M.; Sérvulo, E.F.C. Nitrogen compounds in brewing wort and beer. J. Brew. Distill. 2014, 5, 10–17. [Google Scholar]
- Bogdan, P.; Kordialik-Bogacka, E. Alternatives to malt in brewing. Trends Food Sci. Technol. 2017, 65, 1–9. [Google Scholar] [CrossRef]
- Piddocke, M.P.; Kreisz, S.; Heldt-Hansen, H.P.; Nielsen, K.F.; Olsson, L. Physiological characterization of brewer’s yeast in high-gravity beer fermentations with glucose or maltose syrups as adjuncts. Appl. Microbiol. Biotechnol. 2009, 84, 453–464. [Google Scholar] [CrossRef] [PubMed]
- Torres-Vargas, O.L.; García-Salcedo, Á.J.; Calderón, H.A. Physical-chemical characterization of quinoa (Chenopodium quinoa Willd.), amaranth (Amaranthus caudatus L.), and chia (Salvia hispanica L.) flours and seeds. Acta Agron. 2018, 67, 215–222. [Google Scholar] [CrossRef]
- Chandra, S.; Dwivedi, P.; Baig, M.M.V.; Shinde, L.P. Importance of quinoa and amaranth in food security. J. Agric. Ecol. 2018, 5, 26–37. [Google Scholar]
- Valcárcel-Yamani, B.; Lannes, S.C. Applications of quinoa (Chenopodium Quinoa Willd.) and amaranth (Amaranthus Spp.) and their influence in the nutritional value of cereal based foods. Am. J. Public Health 2012, 6, 265–275. [Google Scholar]
- Schoenlechner, R.; Sibenhandl, S.; Berghofer, E. Pseudocereals. In Gluten-Free Cereal Products and Beverages; Arendt, E.K., Bello, F.D., Eds.; Academic Press: Cambridge, MA, USA, 2008; pp. 149–190. [Google Scholar]
- Alvarez-Jubete, L.; Arendt, E.K.; Gallagher, E. Nutritive value of pseudocereals and their increasing use as functional gluten-free ingredients. Trends Food Sci. Technol. 2010, 21, 106–113. [Google Scholar] [CrossRef]
- Kordialik-Bogacka, E.; Bogdan, P.; Pielech-Przybylska, K.; Michałowska, D. Suitability of unmalted quinoa for beer production. J. Sci. Food Agric. 2018, 98, 5027–5036. [Google Scholar] [CrossRef] [PubMed]
- Budryn, G.; Żyżelewicz, D.; Nebesny, E.; Oracz, J.; Krysiak, W. Influence of addition of green tea and green coffee extracts on the properties of fine yeast pastry fried products. Food Res. Int. 2013, 50, 149–160. [Google Scholar] [CrossRef]
- Dudkiewicz, M.; Berlowska, J.; Kregiel, D. Acid whey as a medium for cultivation of conventional and non-conventional yeasts. Biotechnol. Food Sci. 2016, 80, 75–82. [Google Scholar]
- Balcerek, M.; Pielech-Przybylska, K. Effect of supportive enzymes on chemical composition and viscosity of rye mashes obtained by PSL method and efficiency of their fermentation. Eur. Food Res. Technol. 2009, 229, 141–151. [Google Scholar] [CrossRef]
- Cozzolino, D.; Degner, S. An overview on the role of lipids and fatty acids in barley grain and their products during beer brewing. Food Res. Int. 2016, 81, 114–121. [Google Scholar] [CrossRef]
- Gibson, B. 125th Anniversary Review: Improvement of higher gravity brewery fermentation via wort enrichment and supplementation. J. Inst. Brew. 2011, 117, 268–284. [Google Scholar] [CrossRef]
- Kühbeck, F.; Schütz, M.; Thiele, F.; Krottenthaler, M.; Back, W. Influence of lauter turbidity and hot trub on wort composition, fermentation, and beer quality. J. Am. Soc. Brew. Chem. 2006, 64, 16–28. [Google Scholar] [CrossRef]
- Schisler, D.O.; Ruocco, J.J.; Mabee, M.S. Wort trub content and its effects on fermentation and beer flavour. J. Am. Soc. Brew. Chem. 1982, 40, 57–61. [Google Scholar]
- Thomas, D.S.; Hossack, J.A.; Rose, A.H. Plasma-Membrane lipid composition and ethanol tolerance inSaccharomyces cerevisiae. Arch. Microbiol. 1978, 117, 239–245. [Google Scholar] [CrossRef]
- Stewart, G.G.; Martin, S.A. Wort clarity: Effects on fermentation. Master Brew. Assoc. Am. 2004, 41, 18–26. [Google Scholar]
- Moonjai, N.; Verstrepen, K.J.; Shen, H.-Y.; Derdelinckx, G.; Verachtert, H.; Delvaux, F.R. Linoleic acid supplementation of a cropped brewing lager strain: Effects on subsequent fermentation performance with serial repitching. J. Inst. Brew. 2003, 109, 262–272. [Google Scholar] [CrossRef]
- Hull, G. Hull olive oil addition to yeast as an alternative to wort aeration. Tech. Q. 2008, 45, 17–23. [Google Scholar] [CrossRef] [Green Version]
- Moonjai, N.; Verstrepen, K.J.; Delvaux, F.R.; Derdelinckx, G.; Verachtert, H. The effects if linoleic acid supplementation of cropped yeast on its subsequent fermentation performance and acetate ester synthesis. J. Instit. Brew. 2006, 108, 227–235. [Google Scholar] [CrossRef]
- Thurston, P.A.; Quain, D.E.; Tubb, R.S. Lipid metabolism and the regulation of volatile ester synthesis in Saccharomyces cerevisiae. J. Inst. Brew. 1972, 88, 90–94. [Google Scholar] [CrossRef]
- Kalmokoff, M.L.; Ingledew, W.M. Evaluation of ethanol tolerance in selected Saccharomyces strains. J. Am. Soc. Brew. Chem. 1985, 43, 189–196. [Google Scholar] [CrossRef]
- Chen, E.C.-H. Utilization of wort fatty acids by yeast during fermentation. J. Am. Soc. Brew. Chem. 1980, 38, 148–153. [Google Scholar] [CrossRef]
- Lekkas, C.; Stewart, G.G.; Hill, A.E.; Taidi, B.; Hodgson, J. Elucidation of the role of nitrogenous wort components in yeast fermentation. J. Inst. Brew. 2007, 113, 3–8. [Google Scholar] [CrossRef]
- Kordialik-Bogacka, E.; Bogdan, P.; Diowksz, A. Malted and unmalted oats in brewing. J. Inst. Brew. 2014, 120, 390–398. [Google Scholar] [CrossRef]
- Aastrup, S. Beer from 100% barley. Scand. Brew. Rev. 2010, 67, 28–33. [Google Scholar]
- Tenge, C. Yeast. In Handbook of Brewing; Wiley: Hoboken, NJ, USA, 2009; pp. 119–145. [Google Scholar]
- Thomas, K.C.; Ingledew, W.M. Relationship of low lysine and high arginine concentrations to efficient ethanolic fermentation of wheat mash. Can. J. Microbiol. 1992, 38, 626–634. [Google Scholar] [CrossRef] [PubMed]
- Olaniran, A.O.; Hiralal, L.; Mokoena, M.P.; Pillay, B. Flavour-active volatile compounds in beer: Production, regulation and control. J. Inst. Brew. 2017, 123, 13–23. [Google Scholar] [CrossRef] [Green Version]
- Hazelwood, L.A.; Daran, J.-M.; Van Maris, A.J.A.; Pronk, J.T.; Dickinson, J.R. The Ehrlich pathway for fusel alcohol production: A century of research on Saccharomyces cerevisiae metabolism. Appl. Environ. Microbiol. 2008, 74, 2259–2266. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Fatty Acid (mg/L) | 100% Barley Malt | 30% Amaranth Flakes | 30% Quinoa Flakes | 30% Maltose Syrup | ||||
|---|---|---|---|---|---|---|---|---|
| Wort | Beer | Wort | Beer | Wort | Beer | Wort | Beer | |
| Caproic (C6:0) | 0.20 ± 0.01 a | 0.15 ± 0.01 1 | 0.22 ± 0.02 b | 0.18 ± 0.01 2 | 0.24 ± 0.02 b | 0.16 ± 0.02 1 | 0.17 ± 0.01 c | 0.12 ± 0.01 4 |
| Caprylic (C8:0) | 0.24 ± 0.02 a | 0.20 ± 0.01 1 | 0.30 ± 0.01 b | 0.28 ± 0.01 2 | 0.26 ± 0.02 a | 0.24 ± 0.02 3 | 0.17 ± 0.01 c | 0.16 ± 0.01 4 |
| Capric (C10:0) | 0.22 ± 0.02 a | 0.20 ± 0.00 1 | 0.25 ± 0.01 b | 0.23 ± 0.00 2 | 0.23 ± 0.01 a | 0.23 ± 0.00 2 | 0.15 ± 0.01 c | 0.14 ± 0.01 3 |
| Lauric (C12:0) | 0.22 ± 0.01 a | 0.20 ± 0.00 1 | 0.26 ± 0.00 b | 0.22 ± 0.01 2 | 0.26 ± 0.01 c | 0.20 ± 0.00 1 | 0.16 ± 0.01 d | 0.13 ± 0.01 3 |
| Myristic (C14:0) | 0.23 ± 0.01 a | 0.10 ± 0.00 1 | 0.15 ± 0.02 b | 0.10 ± 0.00 1 | 0.15 ± 0.03 b | 0.10 ± 0.00 1 | 0.20 ± 0.01 c | 0.16 ± 0.01 2 |
| Palmitic (C16:0) | 0.50 ± 0.02 a | 0.20 ± 0.00 1 | 0.77 ± 0.02 b | 0.24 ± 0.01 2 | 0.71 ± 0.03 c | 0.19 ± 0.02 3 | 0.45 ± 0.03 d | 0.15 ± 0.01 4 |
| Margaric (C17:0) | 0.15 ± 0.01 a | 0.08 ± 0.00 1 | 0.27 ± 0.01 b | 0.08 ±0.00 1 | 0.25 ± 0.01 b | 0.10 ± 0.02 2 | 0.15 ± 0.01 a | 0.07 ± 0.01 3 |
| Stearic (C18:0) | 0.67 ± 0.01 a | 0.39 ± 0.00 1 | 0.81 ± 0.02 b | 0.48 ± 0.02 2 | 0.75 ± 0.06 c | 0.43 ± 0.01 3 | 0.32 ± 0.02 d | 0.28 ± 0.01 4 |
| Oleic cis (C18:1) | 1.04 ± 0.04 a | 0.15 ± 0.00 1 | 1.46 ± 0.01 b | 0.20 ± 0.01 2 | 1.34 ± 0.04 c | 0.10 ± 0.01 3 | 0.65 ± 0.01 d | 0.20 ± 0.01 2 |
| Linoleic cis (C18:2) | 0.21 ± 0.02 a | 0.07 ± 0.00 1 | 0.30 ± 0.01 b | 0.11 ± 0.02 2 | 0.37 ± 0.01 c | 0.14 ± 0.01 3 | 0.17 ± 0.01 d | 0.08 ± 0.00 4 |
| α-Linolenic (C18:3 n-3) | 0.36 ± 0.04 a | 0.05 ± 0.00 1 | 0.55 ± 0.01 b | 0.07 ± 0.01 2 | 0.45 ± 0.00 c | 0.05 ± 0.00 1 | 0.15 ± 0.01 d | 0.03 ± 0.01 3 |
| γ-Linolenic (C18:3 n-6) | 0.07 ± 0.00 a | 0.03 ± 0.00 1 | 0.19 ± 0.01 b | 0.06 ± 0.01 2 | 0.16 ± 0.00 c | 0.03 ± 0.00 1 | 0.04 ± 0.00 d | 0.02 ± 0.01 3 |
| Arachidic (C20:0) | 0.09 ± 0.00 a | 0.05 ± 0.00 1 | 0.13 ± 0.02 b | 0.09 ± 0.01 2 | 0.12 ± 0.00 c | 0.08 ± 0.01 2 | 0.06 ± 0.00 f | 0.03 ± 0.02 3 |
| Arachidonic (C20:4) | 0.20 ± 0.03 a | 0.17 ± 0.00 1 | 0.20 ± 0.03 a | 0.15 ± 0.02 2 | 0.22 ± 0.02 a | 0.18 ± 0.00 3 | 0.09 ± 0.00 f | 0.07 ± 0.01 4 |
| Total (mg/L) | 4.40 ± 0.02 a | 2.04 ± 0.01 1 | 5.86 ± 0.01 b | 2.46 ± 0.02 2 | 5.51 ± 0.02 c | 2.23 ± 0.01 3 | 2.93 ± 0.01 d | 1.64 ± 0.01 4 |
| Type of Wort and Beer | FAN (mg/L) | Assimilation (%) | |
|---|---|---|---|
| Wort | Beer | ||
| 100% Barley malt | 240.7 ± 2.7 | 78.2 ± 3.2 a | 68 |
| 30% Amaranth flakes | 208.4 ± 6.2 1 | 56.5 ± 5.2 a,1 | 73 |
| 30% Quinoa flakes | 198.9 ± 4.8 1 | 46.2 ± 2.6 a,1 | 77 |
| 30% Maltose syrup | 134.1 ± 3.9 1 | 72.2 ± 5.3 a,2 | 46 |
| Amino Acid (mg/L) | 100% Barley Malt | 30% Amaranth Flakes | 30% Quinoa Flakes | 30% Maltose Syrup | |||||
|---|---|---|---|---|---|---|---|---|---|
| Wort | Beer | Wort | Beer | Wort | Beer | Wort | Beer | ||
| Group A | Lysine | 32.9 ± 2.4 a | 0.00 1 | 40.8 ± 2.7 b | 0.00 1 | 44.6 ± 1.5 c | 0.00 1 | 19.7 ± 2.1 d | 0.00 1 |
| Asparagine | 35.3 ± 2.2 a | 3.2 ± 0.2 1 | 38.9 ± 6.1 b | 4.8 ± 0.4 2 | 36.4 ± 2.3 a | 3.1 ± 0.2 1 | 21.2 ± 2.3 d | 0.00 3 | |
| Glutamine | 39.7 ± 2.3 a | 4.5 ± 0.1 1 | 42.6 ± 1.1 b | 3.4 ± 0.6 2 | 46.5 ± 3.4 c | 5.1 ± 0.9 3 | 25.8 ± 1.5 d | 0.00 3 | |
| Serine | 25.0 ± 2.1 a | 0.00 1 | 37.8 ± 2.2 b | 3.1 ± 0.7 2 | 36.6 ± 3.1 b | 3.0 ± 0.6 2 | 16.2 ± 2.2 d | 0.00 3 | |
| Arginine | 61.8 ± 7.2 a | 0.00 1 | 62.1 ± 6.9 a | 4.32 ± 0.9 2 | 62.2 ± 6.7 a | 0.00 1 | 46.4 ± 5.6 d | 3.2 ± 0.3 3 | |
| Threonine | 39.1 ± 4.2 a | 4.2 ± 0.4 1 | 20.9 ± 1.5 b | 3.0 ± 0.6 2 | 45.2 ± 1.4 c | 4.4 ± 0.2 1 | 19.6 ± 2.5 d | 0.00 3 | |
| Group B | Histidine | 15.8 ± 1.5 a | 8.2 ± 0.6 1 | 19.1 ± 2.0 b | 4.0 ± 0.4 2 | 18.3 ± 2.6 b | 10.2 ± 2.3 3 | 9.5 ± 2.4 c | 4.3 ± 0.3 2 |
| Valine | 47.0 ± 3.2 a | 15.2 ± 2.2 1 | 49.4 ± 3.4 a | 14.2 ± 2.5 1 | 34.2 ± 2.9 b | 16.5 ± 2.7 1 | 32.9 ± 3.2 b | 25.5 ± 3.5 2 | |
| Methionine | 29.0 ± 1.4 a | 0.00 1 | 32.2 ± 4.1 b | 0.00 1 | 35.3 ± 2.0 c | 0.00 1 | 17.4 ± 2.7 d | 0.00 1 | |
| Isoleucine | 40.1 ± 1.1 a | 10.3 ± 0.9 1 | 35.7 ± 3.2 b | 12.9 ± 1.8 2 | 41.3 ± 0.5 a | 5.5 ± 0.8 3 | 26.0 ± 4.3 c | 6.2 ± 0.6 4 | |
| Leucine | 90.7 ± 2.3 a | 22.7 ± 2.2 1 | 60.8 ± 4.5 b | 17.0 ± 2.1 2 | 100.5 ± 4.8 c | 10.4 ± 0.9 3 | 63.5 ± 3.6 b | 35.2 ± 3.7 4 | |
| Group C | Alanine | 91.0 ± 8.7 a | 65.2 ± 6.7 1 | 95.9 ± 1.2 b | 65.0 ± 5.4 1 | 50.4 ±4.1 c | 42.1 ± 6.9 2 | 72.8 ± 6.5 d | 30.2 ± 6.2 3 |
| Glycine | 17.6 ± 2.3 a | 12.2 ± 2.7 1 | 5.9 ± 1.1 b | 5.0 ± 1.4 2 | 17.5 ± 2.3 a | 16.2 ± 1.7 3 | 12.3 ± 2.2 d | 5.2 ± 1.1 2 | |
| Tyrosine | 76.3 ± 6.7 a | 50.1 ± 8.2 1 | 28.1 ± 2.9 b | 26.0 ± 3.2 2 | 55.2 ± 5.2 c | 50.0 ± 6.2 1 | 57.2 ± 4.7 c | 42.4 ± 3.6 3 | |
| Phenylalanine | 71.7 ± 3.3 a | 22.2 ± 3.1 1 | 39.9 ± 3.2 b | 28.1 ± 2.7 2 | 20.1 ± 1.9 c | 14.2 ± 2.5 3 | 50.2 ± 2.6 d | 30.3 ± 2.8 2 | |
| Group D | Proline | 290.2 ± 11.3 a | 285.7 ± 2.7 1 | 205.4 ± 3.4 b | 203.2 ± 3.9 2 | 190.1 ± 0.9 c | 189.8 ± 1.1 3 | 226.3 ± 2.5 d | 218.4 ± 3.4 4 |
| Total (mg/L) | 1003.2 ± 3.8 a | 504 ± 1.9 1 | 816 ± 3.2 b | 394 ± 1.7 2 | 834 ± 5.1 c | 370 ± 1.6 3 | 717 ± 3.2 d | 400 ± 1.8 4 | |
| Higher Alcohol (mg/L) | Type of Beer | |
|---|---|---|
| 30% Amaranth Flakes | 30% Maltose Syrup | |
| 1-proponaol | 10.3 ± 0.2 | 12.6 ± 0.3 |
| isobutanol | 17.4 ± 0.5 | 11.3 ± 0.3 |
| 2- and 3- methylbutanol | 19.1 ± 0.2 | 18.2 ± 0.2 |
| 2-phenylethanol | 13.3 ± 0.9 | 11.8 ± 1.1 |
| Total higher alcohols (mg/L) | 60.1 ± 1.8 | 53.9 ± 1.9 |
| Total esters (mg/L) | 6.93 ± 0.14 | 6.16 ± 0.10 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bogdan, P.; Kordialik-Bogacka, E.; Czyżowska, A.; Oracz, J.; Żyżelewicz, D. The Profiles of Low Molecular Nitrogen Compounds and Fatty Acids in Wort and Beer Obtained with the Addition of Quinoa (Chenopodium quinoa Willd.), Amaranth (Amaranthus cruentus L.) or Maltose Syrup. Foods 2020, 9, 1626. https://doi.org/10.3390/foods9111626
Bogdan P, Kordialik-Bogacka E, Czyżowska A, Oracz J, Żyżelewicz D. The Profiles of Low Molecular Nitrogen Compounds and Fatty Acids in Wort and Beer Obtained with the Addition of Quinoa (Chenopodium quinoa Willd.), Amaranth (Amaranthus cruentus L.) or Maltose Syrup. Foods. 2020; 9(11):1626. https://doi.org/10.3390/foods9111626
Chicago/Turabian StyleBogdan, Paulina, Edyta Kordialik-Bogacka, Agata Czyżowska, Joanna Oracz, and Dorota Żyżelewicz. 2020. "The Profiles of Low Molecular Nitrogen Compounds and Fatty Acids in Wort and Beer Obtained with the Addition of Quinoa (Chenopodium quinoa Willd.), Amaranth (Amaranthus cruentus L.) or Maltose Syrup" Foods 9, no. 11: 1626. https://doi.org/10.3390/foods9111626