In Vitro Characterisation of the Antioxidative Properties of Whey Protein Hydrolysates Generated under pH- and Non pH-Controlled Conditions

 , , and
, , and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Enzymatic Hydrolysis of WPC80
2.3. Determination of Degree of Hydrolysis (DH)
2.4. Sodium Dodecyl Sulfate Polyacrylamide Gel Electrophoresis (SDS-PAGE)
2.5. Liquid Chromatography (LC)
2.6. In Vitro Antioxidant Analysis
2.6.1. ORAC Assay
2.6.2. TEAC Assay
2.7. Cellular Antioxidant Assay
2.7.1. Tissue Culture
2.7.2. Cell Viability
2.7.3. Assay of Intracellular ROS Generation
2.8. Peptide Identification by LC-MS/MS
2.9. Statistical Analysis
3. Results and Discussion
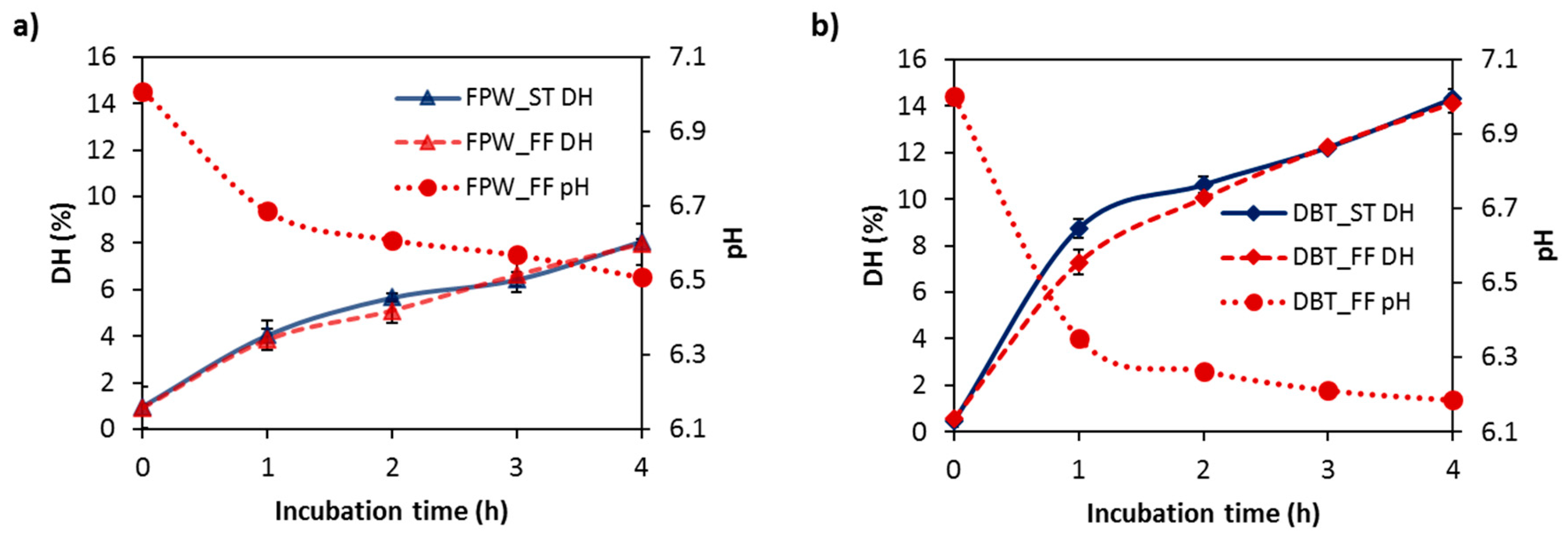
3.1. Degree of Hydrolysis (DH) of WPHs
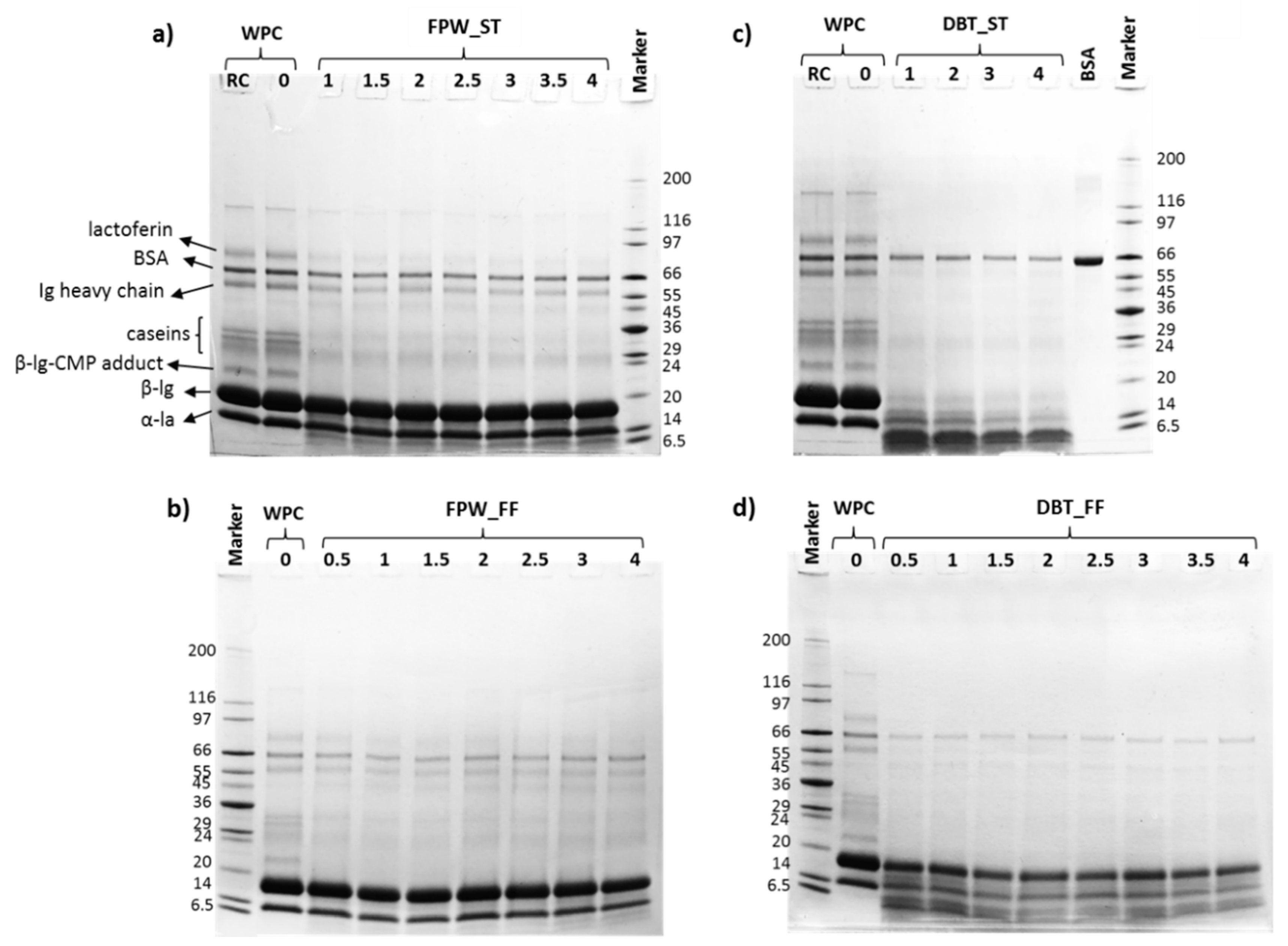
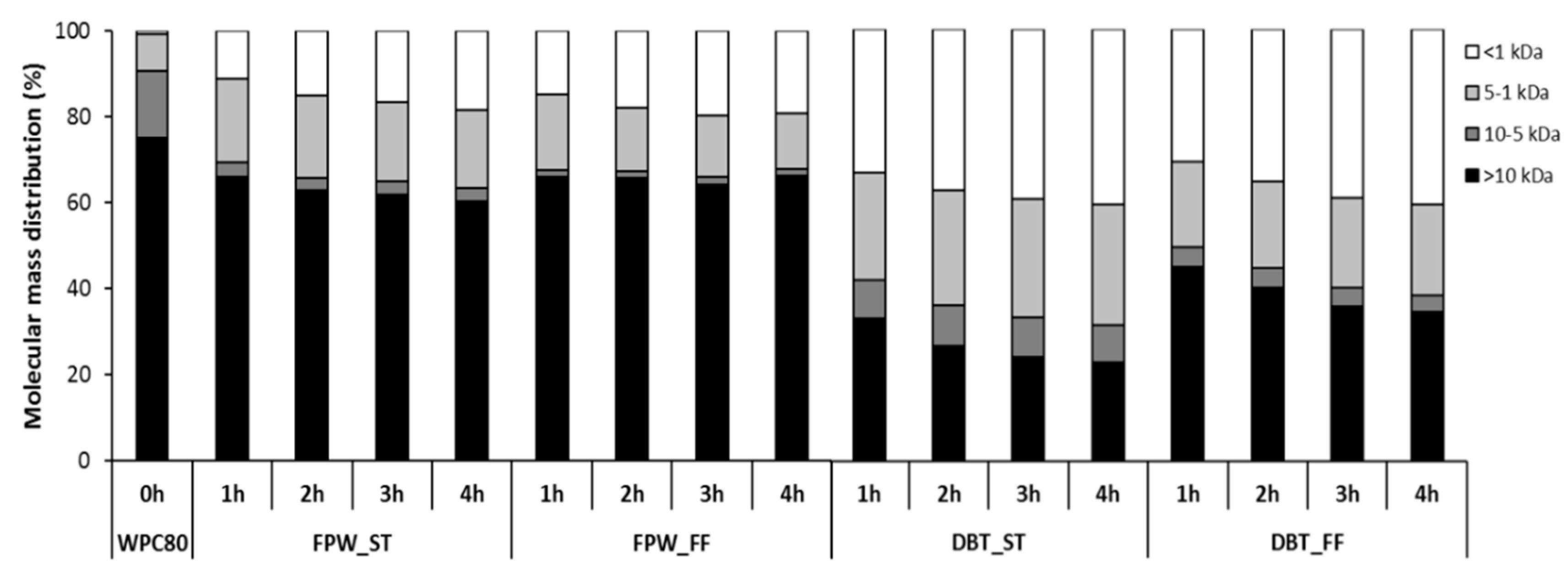
3.2. Electrophoresis and Molecular Mass Distribution Profiles
3.3. Reverse-Phase (RP) Peptide Profiles
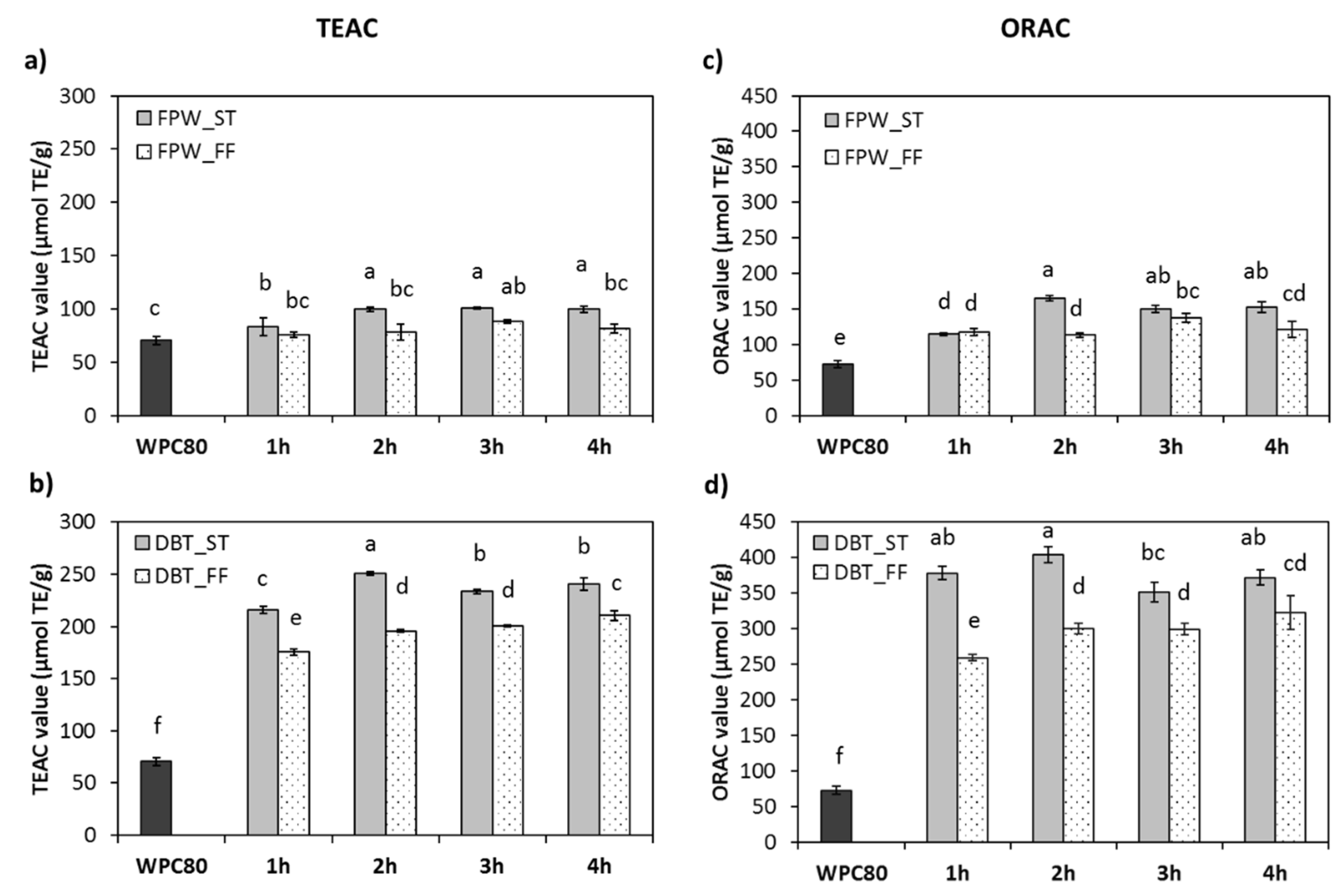
3.4. In Vitro Antioxidant Properties
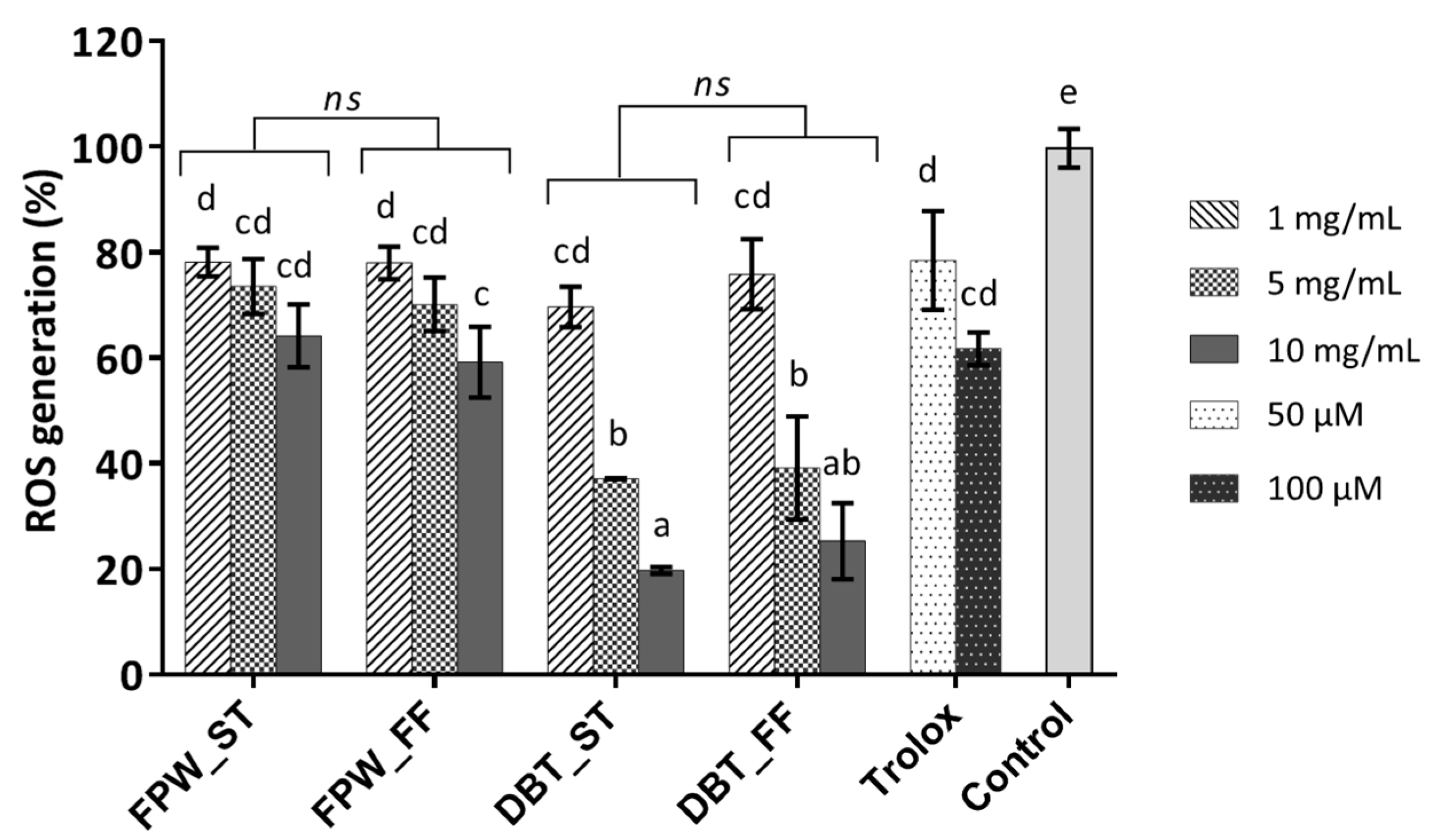
3.5. Cellular Antioxidant Activity
3.6. Peptide Identification by LC-MS/MS
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Brandelli, A.; Daroit, D.J.; Corrêa, A.P.F. Whey as a source of peptides with remarkable biological activities. Food Res. Int. 2015, 73, 149–161. [Google Scholar] [CrossRef] [Green Version]
- Nongonierma, A.B.; O’Keeffe, M.B.; FitzGerald, R.J. Milk protein hydrolysates and bioactive peptides. In Advanced Dairy Chemistry: Volume 1b: Proteins: Applied Aspects; McSweeney, P.L.H., O’Mahony, J.A., Eds.; Springer: New York, NY, USA, 2016; pp. 417–482. [Google Scholar]
- Corrochano, A.R.; Buckin, V.; Kelly, P.M.; Giblin, L. Invited review: Whey proteins as antioxidants and promoters of cellular antioxidant pathways. J. Dairy Sci. 2018, 101, 4747–4761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, I.T.; Nadeem, M.; Imran, M.; Ullah, R.; Ajmal, M.; Jaspal, M.H. Antioxidant properties of milk and dairy products: A comprehensive review of the current knowledge. Lipids Health Dis. 2019, 18, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mangano, K.M.; Bao, Y.; Zhao, C. Nutritional properties of whey proteins. In Whey Protein Production, Chemistry, Functionality, and Applications; Guo, M., Ed.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2019; pp. 103–140. [Google Scholar]
- Poprac, P.; Jomova, K.; Simunkova, M.; Kollar, V.; Rhodes, C.J.; Valko, M. Targeting free radicals in oxidative stress-related human diseases. Trends Pharm. Sci. 2017, 38, 592–607. [Google Scholar] [CrossRef]
- Niki, E. Assessment of antioxidant capacity in vitro and in vivo. Free Radic. Biol. Med. 2010, 49, 503–515. [Google Scholar] [CrossRef]
- Udenigwe, C.C.; Aluko, R.E. Food protein-derived bioactive peptides: Production, processing, and potential health benefits. J. Food Sci. 2012, 77, R11–R24. [Google Scholar] [CrossRef]
- Bamdad, F.; Bark, S.; Kwon, C.H.; Suh, J.-W.; Sunwoo, H. Anti-inflammatory and antioxidant properties of peptides released from β-lactoglobulin by high hydrostatic pressure-assisted enzymatic hydrolysis. Molecules 2017, 22, 949. [Google Scholar] [CrossRef] [Green Version]
- Contreras, M.d.M.; Hernández-Ledesma, B.; Amigo, L.; Martín-Álvarez, P.J.; Recio, I. Production of antioxidant hydrolyzates from a whey protein concentrate with thermolysin: Optimization by response surface methodology. LWT Food Sci. Technol. 2011, 44, 9–15. [Google Scholar] [CrossRef]
- Jiang, Z.; Yao, K.; Yuan, X.; Mu, Z.; Gao, Z.; Hou, J.; Jiang, L. Effects of ultrasound treatment on physicochemical, functional properties and antioxidant activity of whey protein isolate in the presence of calcium lactate. J. Sci. Food Agric. 2018, 98, 1522–1529. [Google Scholar] [CrossRef]
- Le Maux, S.; Nongonierma, A.B.; Barre, C.; FitzGerald, R.J. Enzymatic generation of whey protein hydrolysates under pH-controlled and non pH-controlled conditions: Impact on physicochemical and bioactive properties. Food Chem. 2016, 199, 246–251. [Google Scholar] [CrossRef] [Green Version]
- Madadlou, A.; Abbaspourrad, A. Bioactive whey peptide particles: An emerging class of nutraceutical carriers. Crit. Rev. Food Sci. Nutr. 2018, 58, 1468–1477. [Google Scholar] [CrossRef] [PubMed]
- Neto, Y.A.A.H.; Rosa, J.C.; Cabral, H. Peptides with antioxidant properties identified from casein, whey, and egg albumin hydrolysates generated by two novel fungal proteases. Prep. Biochem. Biotechnol. 2019, 49, 639–648. [Google Scholar] [CrossRef] [PubMed]
- O’Keeffe, M.B.; FitzGerald, R.J. Antioxidant effects of enzymatic hydrolysates of whey protein concentrate on cultured human endothelial cells. Int. Dairy J. 2014, 36, 128–135. [Google Scholar] [CrossRef]
- Power, O.; Jakeman, P.; FitzGerald, R.J. Antioxidative peptides: Enzymatic production, in vitro and in vivo antioxidant activity and potential applications of milk-derived antioxidative peptides. Amino Acids 2013, 44, 797–820. [Google Scholar] [CrossRef] [PubMed]
- Salami, M.; Moosavi-Movahedi, A.A.; Ehsani, M.R.; Yousefi, R.; Haertlé, T.; Chobert, J.-M.; Razavi, S.H.; Henrich, R.; Balalaie, S.; Ebadi, S.A.; et al. Improvement of the antimicrobial and antioxidant activities of camel and bovine whey proteins by limited proteolysis. J. Agric. Food Chem. 2010, 58, 3297–3302. [Google Scholar] [CrossRef]
- Korhonen, H.; Pihlanto, A. Bioactive peptides: Production and functionality. Int. Dairy J. 2006, 16, 945–960. [Google Scholar] [CrossRef]
- Tagliazucchi, D.; Helal, A.; Verzelloni, E.; Conte, A. Bovine milk antioxidant properties: Effect of in vitro digestion and identification of antioxidant compounds. Dairy Sci. Technol. 2016, 96, 657–676. [Google Scholar] [CrossRef] [Green Version]
- De Carvalho, N.C.; Pessato, T.B.; Fernandes, L.G.R.; De Lima Zollner, R.; Netto, F.M. Physicochemical characteristics and antigenicity of whey protein hydrolysates obtained with and without pH control. Int. Dairy J. 2017, 71, 24–34. [Google Scholar] [CrossRef]
- Cheison, S.C.; Kulozik, U. Impact of the environmental conditions and substrate pre-treatment on whey protein hydrolysis: A review. Crit. Rev. Food Sci. Nutr. 2017, 57, 418–453. [Google Scholar] [CrossRef]
- Cheison, S.C.; Leeb, E.; Toro-Sierra, J.; Kulozik, U. Influence of hydrolysis temperature and pH on the selective hydrolysis of whey proteins by trypsin and potential recovery of native alpha-lactalbumin. Int. Dairy J. 2011, 21, 166–171. [Google Scholar] [CrossRef]
- Fernández, A.; Kelly, P. pH-stat vs. Free-fall pH techniques in the enzymatic hydrolysis of whey proteins. Food Chem. 2016, 199, 409–415. [Google Scholar] [CrossRef] [PubMed]
- Butré, C.I.; Sforza, S.; Wierenga, P.A.; Gruppen, H. Determination of the influence of the pH of hydrolysis on enzyme selectivity of Bacillus licheniformis protease towards whey protein isolate. Int. Dairy J. 2015, 44, 44–53. [Google Scholar] [CrossRef]
- Butré, C.I.; Wierenga, P.A.; Gruppen, H. Effects of ionic strength on the enzymatic hydrolysis of diluted and concentrated whey protein isolate. J. Agric. Food Chem. 2012, 60, 5644–5651. [Google Scholar] [CrossRef] [PubMed]
- Butré, C.I.; Sforza, S.; Gruppen, H.; Wierenga, P.A. Introducing enzyme selectivity: A quantitative parameter to describe enzymatic protein hydrolysis. Anal. Bioanal. Chem. 2014, 406, 5827–5841. [Google Scholar] [CrossRef] [PubMed]
- Apak, R.; Özyürek, M.; Güçlü, K.; Çapanoğlu, E. Antioxidant activity/capacity measurement. 2. Hydrogen atom transfer (HAT)-based, mixed-mode (electron transfer (ET)/HAT), and lipid peroxidation assays. J. Agric. Food Chem. 2016, 64, 1028–1045. [Google Scholar] [CrossRef] [PubMed]
- Spellman, D.; McEvoy, E.; O’Cuinn, G.; FitzGerald, R.J. Proteinase and exopeptidase hydrolysis of whey protein: Comparison of the TNBS, OPA and pH stat methods for quantification of degree of hydrolysis. Int. Dairy. J. 2003, 13, 447–453. [Google Scholar] [CrossRef]
- O’Loughlin, I.B.; Murray, B.A.; Kelly, P.M.; FitzGerald, R.J.; Brodkorb, A. Enzymatic hydrolysis of heat-induced aggregates of whey protein isolate. J. Agric. Food Chem. 2012, 60, 4895–4904. [Google Scholar] [CrossRef] [Green Version]
- Spellman, D.; O’Cuinn, G.; FitzGerald, R.J. Bitterness in Bacillus proteinase hydrolysates of whey proteins. Food Chem. 2009, 114, 440–446. [Google Scholar] [CrossRef]
- Le Maux, S.; Nongonierma, A.B.; Lardeux, C.; FitzGerald, R.J. Impact of enzyme inactivation conditions during the generation of whey protein hydrolysates on their physicochemical and bioactive properties. Int. J. Food Sci. Technol. 2018, 53, 219–227. [Google Scholar] [CrossRef] [Green Version]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Yarnpakdee, S.; Benjakul, S.; Kristinsson, H.G.; Bakken, H.E. Preventive effect of nile tilapia hydrolysate against oxidative damage of HepG2 cells and DNA mediated by H2O2 and AAPH. J. Food Sci. Technol. 2015, 52, 6194–6205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nongonierma, A.B.; Cadamuro, C.; Le Gouic, A.; Mudgil, P.; Maqsood, S.; FitzGerald, R.J. Dipeptidyl peptidase IV (DPP-IV) inhibitory properties of a camel whey protein enriched hydrolysate preparation. Food Chem. 2019, 279, 70–79. [Google Scholar] [CrossRef] [PubMed]
- O’Keeffe, M.B.; FitzGerald, R.J. Identification of short peptide sequences in complex milk protein hydrolysates. Food Chem. 2015, 184, 140–146. [Google Scholar] [CrossRef] [PubMed]
- Heberle, H.; Meirelles, G.V.; Da Silva, F.R.; Telles, G.P.; Minghim, R. InteractiVenn: A web-based tool for the analysis of sets through Venn diagrams. BMC Bioinform. 2015, 16, 169. [Google Scholar] [CrossRef]
- Haileselassie, S.S.; Lee, B.H.; Gibbs, B.F. Purification and identification of potentially bioactive peptides from enzyme-modified cheese. J. Dairy Sci. 1999, 82, 1612–1617. [Google Scholar] [CrossRef]
- Nongonierma, A.B.; FitzGerald, R.J. Enzymes exogenous to milk in dairy technology | Proteinases. In Encyclopedia of Dairy Sciences, 2nd ed.; Fuquay, J.W., Ed.; Academic Press: San Diego, CA, USA, 2011; pp. 289–296. [Google Scholar]
- Spellman, D.; O’Cuinn, G.; FitzGerald, R.J. Physicochemical and sensory characteristics of whey protein hydrolysates generated at different total solids levels. J. Dairy Res. 2005, 72, 138–143. [Google Scholar] [CrossRef] [Green Version]
- O’Sullivan, D.; Nongonierma, A.B.; FitzGerald, R.J. Bitterness in sodium caseinate hydrolysates: Role of enzyme preparation and degree of hydrolysis. J. Sci. Food Agric. 2017, 97, 4652–4655. [Google Scholar] [CrossRef] [Green Version]
- Kalyankar, P.; Zhu, Y.; O’Cuinn, G.; FitzGerald, R.J. Investigation of the substrate specificity of glutamyl endopeptidase using purified bovine β-casein and synthetic peptides. J. Agric. Food Chem. 2013, 61, 3193–3204. [Google Scholar] [CrossRef]
- Power, O.; Fernández, A.; Norris, R.; Riera, F.A.; FitzGerald, R.J. Selective enrichment of bioactive properties during ultrafiltration of a tryptic digest of β-lactoglobulin. J. Funct. Foods 2014, 9, 38–47. [Google Scholar] [CrossRef]
- Giblin, L.; Yalçın, A.S.; Biçim, G.; Krämer, A.C.; Chen, Z.; Callanan, M.J.; Arranz, E.; Davies, M.J. Whey proteins: Targets of oxidation, or mediators of redox protection. Free Radic. Res. 2019, 53, 1136–1152. [Google Scholar] [CrossRef]
- Xu, R.; Liu, N.; Xu, X.; Kong, B. Antioxidative effects of whey protein on peroxide-induced cytotoxicity. J Dairy Sci. 2011, 94, 3739–3746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corrochano, A.R.; Ferraretto, A.; Arranz, E.; Stuknytė, M.; Bottani, M.; O’Connor, P.M.; Kelly, P.M.; De Noni, I.; Buckin, V.; Giblin, L. Bovine whey peptides transit the intestinal barrier to reduce oxidative stress in muscle cells. Food Chem. 2019, 288, 306–314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, B.; Peng, X.; Xiong, Y.L.; Zhao, X. Protection of lung fibroblast MRC-5 cells against hydrogen peroxide-induced oxidative damage by 0.1–2.8 kDa antioxidative peptides isolated from whey protein hydrolysate. Food Chem. 2012, 135, 540–547. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.-X.; Ling, Y.-F.; Sun, Z.; Zhang, L.; Yu, H.-X.; Kamau, S.M.; Lu, R.-R. Protective effect of whey protein hydrolysates against hydrogen peroxide-induced oxidative stress on PC12 cells. Biotechnol. Lett. 2012, 34, 2001–2006. [Google Scholar] [CrossRef] [PubMed]
- Piccolomini, A.; Iskandar, M.; Lands, L.; Kubow, S. High hydrostatic pressure pre-treatment of whey proteins enhances whey protein hydrolysate inhibition of oxidative stress and IL-8 secretion in intestinal epithelial cells. Food Nutr. Res. 2012, 56, 17549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iskandar, M.; Lands, L.; Sabally, K.; Azadi, B.; Meehan, B.; Mawji, N.; Skinner, C.; Kubow, S. High hydrostatic pressure pretreatment of whey protein isolates improves their digestibility and antioxidant capacity. Foods 2015, 4, 184–207. [Google Scholar] [CrossRef] [Green Version]
- Pyo, M.C.; Yang, S.-Y.; Chun, S.-H.; Oh, N.S.; Lee, K.-W. Protective effects of maillard reaction products of whey protein concentrate against oxidative stress through an Nrf2-dependent pathway in HepG2 cells. Biol. Pharm. Bull. 2016, 39, 1437–1447. [Google Scholar] [CrossRef] [Green Version]
- Kim, G.-N.; Kwon, Y.-I.; Jang, H.-D. Protective mechanism of quercetin and rutin on 2,2′-azobis(2-amidinopropane)dihydrochloride or Cu2+-induced oxidative stress in HepG2 cells. Toxicol. In Vitro 2011, 25, 138–144. [Google Scholar] [CrossRef]
- Honzel, D.; Carter, S.G.; Redman, K.A.; Schauss, A.G.; Endres, J.R.; Jensen, G.S. Comparison of chemical and cell-based antioxidant methods for evaluation of foods and natural products: Generating multifaceted data by parallel testing using erythrocytes and polymorphonuclear cells. J. Agric. Food Chem. 2008, 56, 8319–8325. [Google Scholar] [CrossRef] [Green Version]
- Lima, C.F.; Fernandes-Ferreira, M.; Pereira-Wilson, C. Phenolic compounds protect HepG2 cells from oxidative damage: Relevance of glutathione levels. Life Sci. 2006, 79, 2056–2068. [Google Scholar] [CrossRef] [Green Version]
- Athira, S.; Mann, B.; Sharma, R.; Kumar, R. Ameliorative potential of whey protein hydrolysate against paracetamol-induced oxidative stress. J. Dairy Sci. 2013, 96, 1431–1437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashok, N.R.; Vivek, K.H.; Aparna, H.S. Antioxidative role of buffalo (Bubalus bubalis) colostrum whey derived peptides during oxidative damage. Int. J. Pept. Res. 2019, 25, 1501–1508. [Google Scholar] [CrossRef]
- Athira, S.; Mann, B.; Saini, P.; Sharma, R.; Kumar, R.; Singh, A.K. Production and characterisation of whey protein hydrolysate having antioxidant activity from cheese whey. J. Sci. Food Agric. 2015, 95, 2908–2915. [Google Scholar] [CrossRef] [PubMed]
- Mann, B.; Kumari, A.; Kumar, R.; Sharma, R.; Prajapati, K.; Mahboob, S.; Athira, S. Antioxidant activity of whey protein hydrolysates in milk beverage system. J. Food Sci. Technol. 2015, 52, 3235–3241. [Google Scholar] [CrossRef]
- Picariello, G.; Iacomino, G.; Mamone, G.; Ferranti, P.; Fierro, O.; Gianfrani, C.; Di Luccia, A.; Addeo, F. Transport across Caco-2 monolayers of peptides arising from in vitro digestion of bovine milk proteins. Food Chem. 2013, 139, 203–212. [Google Scholar] [CrossRef]
- Mullally, M.M.; Meisel, H.; FitzGerald, R.J. Identification of a novel angiotensin-I-converting enzyme inhibitory peptide corresponding to a tryptic fragment of bovine β-lactoglobulin. FEBS Lett. 1997, 402, 99–101. [Google Scholar] [CrossRef] [Green Version]
- O’Keeffe, M.B.; Conesa, C.; FitzGerald, R.J. Identification of angiotensin converting enzyme inhibitory and antioxidant peptides in a whey protein concentrate hydrolysate produced at semi-pilot scale. Int. J. Food Sci. Technol. 2017, 52, 1751–1759. [Google Scholar] [CrossRef]
- Hernández-Ledesma, B.; Miguel, M.; Amigo, L.; Aleixandre, M.A.; Recio, I. Effect of simulated gastrointestinal digestion on the antihypertensive properties of synthetic β-lactoglobulin peptide sequences. J. Dairy Res. 2007, 74, 336–339. [Google Scholar] [CrossRef]
- Pihlanto-LeppÄLÄ, A.; Koskinen, P.; Piilola, K.; Tupasela, T.; Korhonen, H. Angiotensin I-converting enzyme inhibitory properties of whey protein digests: Concentration and characterization of active peptides. J. Dairy Res. 2000, 67, 53–64. [Google Scholar] [CrossRef]
- Pellegrini, A.; Dettling, C.; Thomas, U.; Hunziker, P. Isolation and characterization of four bactericidal domains in the bovine β-lactoglobulin. Biochim. Biophys. Acta (BBA) Gen. Subj. 2001, 1526, 131–140. [Google Scholar] [CrossRef]
- Tavares, T.; Contreras, M.d.M.; Amorim, M.; Pintado, M.; Recio, I.; Malcata, F.X. Novel whey-derived peptides with inhibitory effect against angiotensin-converting enzyme: In vitro effect and stability to gastrointestinal enzymes. Peptides 2011, 32, 1013–1019. [Google Scholar] [CrossRef] [PubMed]
- Lacroix, I.M.E.; Li-Chan, E.C.Y. Overview of food products and dietary constituents with antidiabetic properties and their putative mechanisms of action: A natural approach to complement pharmacotherapy in the management of diabetes. Mol. Nutr. Food Res. 2014, 58, 61–78. [Google Scholar] [CrossRef] [PubMed]
- Silveira, H.; Moraes, H.; Oliveira, N.; Coutinho, E.S.F.; Laks, J.; Deslandes, A. Physical exercise and clinically depressed patients: A systematic review and meta-analysis. Neuropsychobiology 2013, 67, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Bertucci, J.I.; Liggieri, C.S.; Colombo, M.L.; Vairo Cavalli, S.E.; Bruno, M.A. Application of peptidases from Maclura pomifera fruit for the production of active biopeptides from whey protein. LWT Food Sci. Technol. 2015, 64, 157–163. [Google Scholar] [CrossRef]
- Tulipano, G.; Sibilia, V.; Caroli, A.M.; Cocchi, D. Whey proteins as source of dipeptidyl dipeptidase IV (dipeptidyl peptidase-4) inhibitors. Peptides 2011, 32, 835–838. [Google Scholar] [CrossRef]
- Tulipano, G.; Faggi, L.; Nardone, A.; Cocchi, D.; Caroli, A.M. Characterisation of the potential of β-lactoglobulin and α-lactalbumin as sources of bioactive peptides affecting incretin function: In silico and in vitro comparative studies. Int. Dairy J. 2015, 48, 66–72. [Google Scholar] [CrossRef]
- FitzGerald, R.; Meisel, H. Lactokinins: Whey protein-derived ACE inhibitory peptides. Food/Nahrung 1999, 43, 165–167. [Google Scholar] [CrossRef]
- Maes, W.; Van Camp, J.; Vermeirssen, V.; Hemeryck, M.; Ketelslegers, J.M.; Schrezenmeir, J.; Van Oostveldt, P.; Huyghebaert, A. Influence of the lactokinin Ala-Leu-Pro-Met-His-Ile-Arg (ALPMHIR) on the release of endothelin-1 by endothelial cells. Regul. Pept. 2004, 118, 105–109. [Google Scholar] [CrossRef]







 ), non pH-controlled (
), non pH-controlled (  ) conditions and Debitrase (DBT) under pH-controlled (
) conditions and Debitrase (DBT) under pH-controlled (  ) and non pH-controlled (
) and non pH-controlled (  ) conditions. Peptide sequences underlined in different colours indicate the peptides identified in the samples described above.
), non pH-controlled ( ) conditions and Debitrase (DBT) under pH-controlled ( ) and non pH-controlled ( ) conditions. Peptide sequences underlined in different colours indicate the peptides identified in the samples described above.
) conditions. Peptide sequences underlined in different colours indicate the peptides identified in the samples described above.
), non pH-controlled ( ) conditions and Debitrase (DBT) under pH-controlled ( ) and non pH-controlled ( ) conditions. Peptide sequences underlined in different colours indicate the peptides identified in the samples described above.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide Sequence 1 | Region in Mature β-lg | Identified In | Reported Activity 2 | IC50/EC50 Values 3 | Reference | References to Related Peptide Sequence 1,2,3,4 | |||
|---|---|---|---|---|---|---|---|---|---|
| FPW_ FF | FPW_ ST | DBT_ FF | DBT_ ST | ||||||
| KVAGTWYSL | f(14−22) | ✓ | ✓ | - | - | - | VAGTWY (ORAC 5.63 µmol TE/µmol peptide, DPP-IVi IC50 74.9 µM [42], ACEi IC50 1,682 µM [59]), VAGT (ACEi IC50 610.3 µM; ORAC 1.66 µmol TE/mmol peptide [60]), WYSL (DPPH & superoxide scavenging activity with EC50 273.63 & 558.42 µM, respectively [61]) | ||
| VAGTW | f(15−19) | ✓ | ACEi | 534 | Pihlanto-LeppÄLÄ, et al. [62] | ||||
| GTWYSL | f(17−22) | ✓ | FRAP | na | Bamdad, et al. [9] | ||||
| AMAASDISLL | f(23−32) | ✓ | FRAP | na | Bamdad, et al. [9] | AASDISLLDAQSAPLR (antibacterial [63]) MAA (ORAC EC50 0.33 µmol TE/mmol peptide, ACEi IC50 515.5 µM [60]) | |||
| MAASDISLL | f(24−32) | ✓ | FRAP | na | Bamdad, et al. [9] | ||||
| LDAQSAPLR | f(32−40) | ✓ | ✓ | ACEi | 635 | Pihlanto-LeppÄLÄ, et al. [62] | |||
| DAQSAPLRVY | f(33−42) | ✓ | ✓ | ACEi | 13 | Tavares, et al. [64] | |||
| DAQSAPLR | f(33−40) | ✓ | ✓ | - | - | - | |||
| SAPLR | f(36−40) | ✓ | - | - | - | ||||
| ELKPTPEGDLEIL | f(45−57) | ✓ | ✓ | FRAP | na | Bamdad, et al. [9] | - | ||
| LKPTPEGDLEIL | f(46−57) | ✓ | DPP-IVi | 57 | Lacroix and Li-Chan [65] | - | |||
| IIAEKTKIPAVF | f(71−82) | ✓ | ✓ | FRAP | na | Bamdad, et al. [9] | IIAEK (ORAC 0.016 µmol TE/µmol peptide, ACEi IC50 63.7 µM [42]), IAEKTKIP (ORAC [10]), IPAVFK (ACEi IC50 144.8 µM, DPP-IVi IC50 149.1 µM, ORAC EC50 0.002 µmol TE/µmol peptide [42]) | ||
| AEKTKIPAVF | f(73−82) | ✓ | ✓ | ✓ | - | - | - | ||
| AEKTKIPA | f(73−80) | ✓ | ✓ | ✓ | - | - | - | ||
| KIPAVF | f(77−82) | ✓ | - | - | - | ||||
| IPAVF | f(78–82) | ✓ | DPP-IV | 44.7 | Silveira, et al. [66] | ||||
| VLDTDYKKY | f(94−102) | ✓ | ✓ | - | - | - | LDTDYKKYLLFCMENS (ABTS [67]), DTDYK (ABTS [56]), VLVLDTDYK (DPP-IVi IC50 424.4 µM [66]) | ||
| VLDTDYK | f(94−100) | ✓ | ABTS | na | Athira, et al. [56] | ||||
| ACEi | 946 | Pihlanto-LeppÄLÄ, et al. [62] | |||||||
| LDTDYKKY | f(95−102) | ✓ | ✓ | - | - | - | |||
| LDTDYKK | f(95−101) | ✓ | ORAC, ACEi | na | Contreras, et al. [10] | ||||
| DTDYKKYLLF | f(96−105) | ✓ | ✓ | - | - | - | |||
| DTDYKK | f(96−101) | ✓ | ✓ | - | - | - | |||
| LVRTPEVDDEALEKF | f(123−135) | ✓ | ✓ | - | - | - | VRTPEVDDEALE, LVRTPEVDDEALE, RTPEVDDEALE (ABTS [56]) | ||
| VRTPEVDDE | f(123−131) | ✓ | ✓ | ABTS | na | Athira, et al. [56] | |||
| VRTPEVDDEALEK | f(123−134) | ✓ | ✓ | - | - | - | |||
| RTPEVDDEALEK | f(124−134) | ✓ | ✓ | - | - | - | |||
| TPEVDDEALEK | f(125−135) | ✓ | ✓ | ✓ | ✓ | DPP-IVi | 319.5 | Silveira, et al. [66] | |
| ORAC | 0.004 | Power, et al. [42] | |||||||
| ABTS | na | Mann, et al. [57] | |||||||
| ALKALPM | f(139−145) | ✓ | - | - | - | ALPMHIR (ACEi IC50 43 µM [59], ORAC EC50 0.035 µmol TE/µmol peptide [42]) | |||
| KALPM | f(141−145) | ✓ | - | - | - | ||||
| ALPMH | f(142−146) | ✓ | ✓ | ✓ | ACEi | 521 | Mullally, et al. [59] | ||
| DPP-IVi | >100 | Tulipano, et al. [68] | |||||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kleekayai, T.; Le Gouic, A.V.; Deracinois, B.; Cudennec, B.; FitzGerald, R.J. In Vitro Characterisation of the Antioxidative Properties of Whey Protein Hydrolysates Generated under pH- and Non pH-Controlled Conditions. Foods 2020, 9, 582. https://doi.org/10.3390/foods9050582
Kleekayai T, Le Gouic AV, Deracinois B, Cudennec B, FitzGerald RJ. In Vitro Characterisation of the Antioxidative Properties of Whey Protein Hydrolysates Generated under pH- and Non pH-Controlled Conditions. Foods. 2020; 9(5):582. https://doi.org/10.3390/foods9050582
Chicago/Turabian StyleKleekayai, Thanyaporn, Aurélien V. Le Gouic, Barbara Deracinois, Benoit Cudennec, and Richard J. FitzGerald. 2020. "In Vitro Characterisation of the Antioxidative Properties of Whey Protein Hydrolysates Generated under pH- and Non pH-Controlled Conditions" Foods 9, no. 5: 582. https://doi.org/10.3390/foods9050582