Biochemical Properties of Polyphenol Oxidases from Ready-to-Eat Lentil (Lens culinaris Medik.) Sprouts and Factors Affecting Their Activities: A Search for Potent Tools Limiting Enzymatic Browning


 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Materials and Sprouting Conditions
2.3. Enzyme Assay
2.4. Protein Determination
2.5. Enzyme Extraction and Partial Purification
2.6. Characterization of PPO
2.6.1. Kinetic Data Analysis and Substrate Specificity
2.6.2. Effect of Temperature on Enzyme Activity
2.6.3. Effect of pH on Enzyme Activity
2.6.4. Effect of Antibrowning Agents on PPOs
2.6.5. Effect of Ions on Enzyme Activity
2.7. Statistical Analysis
3. Results and Discussion
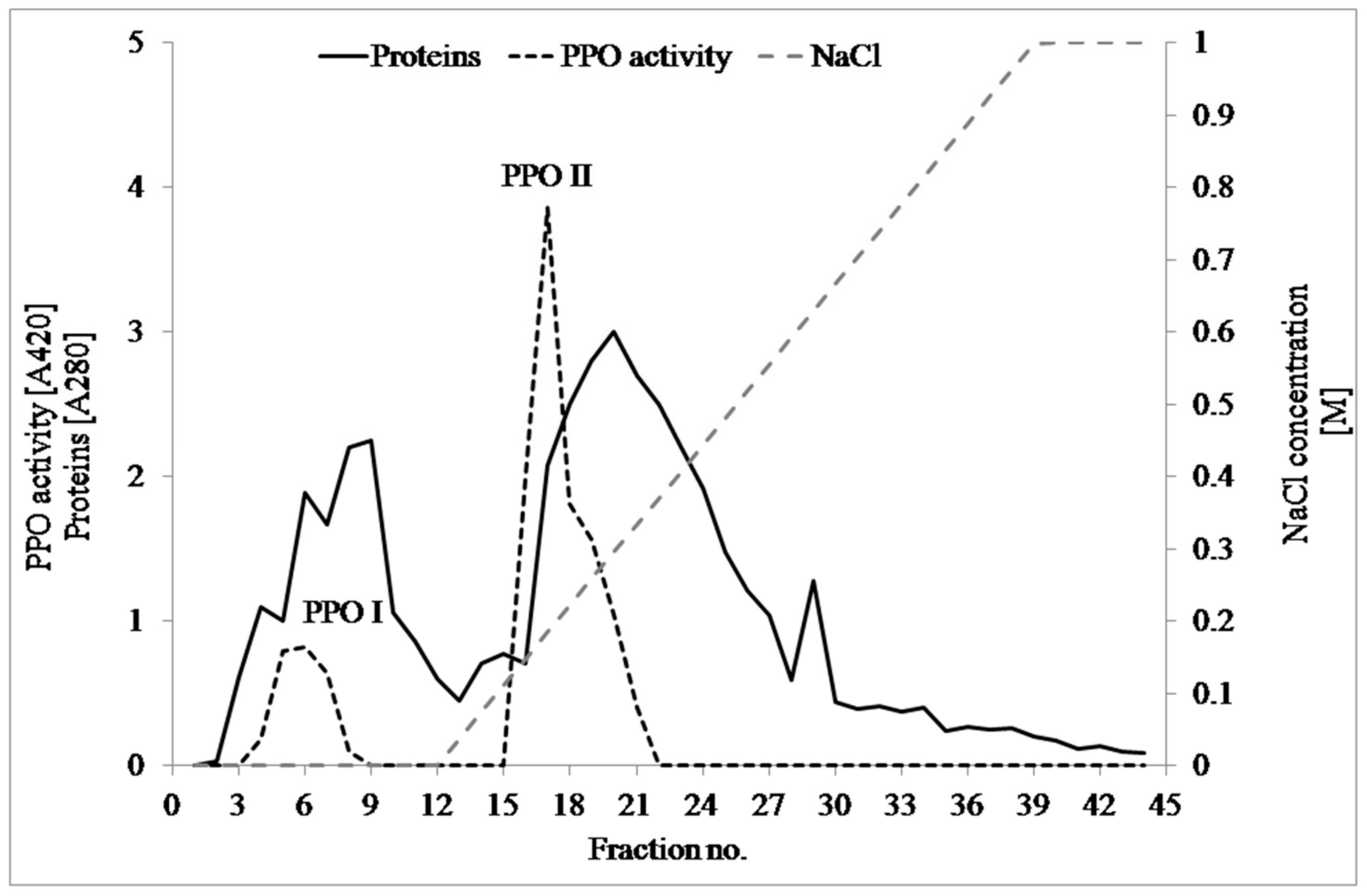
3.1. PPO Isolation and Partial Purification
3.2. Substrate Specificity and Some Kinetic Parameters of Lentil Sprout PPOs
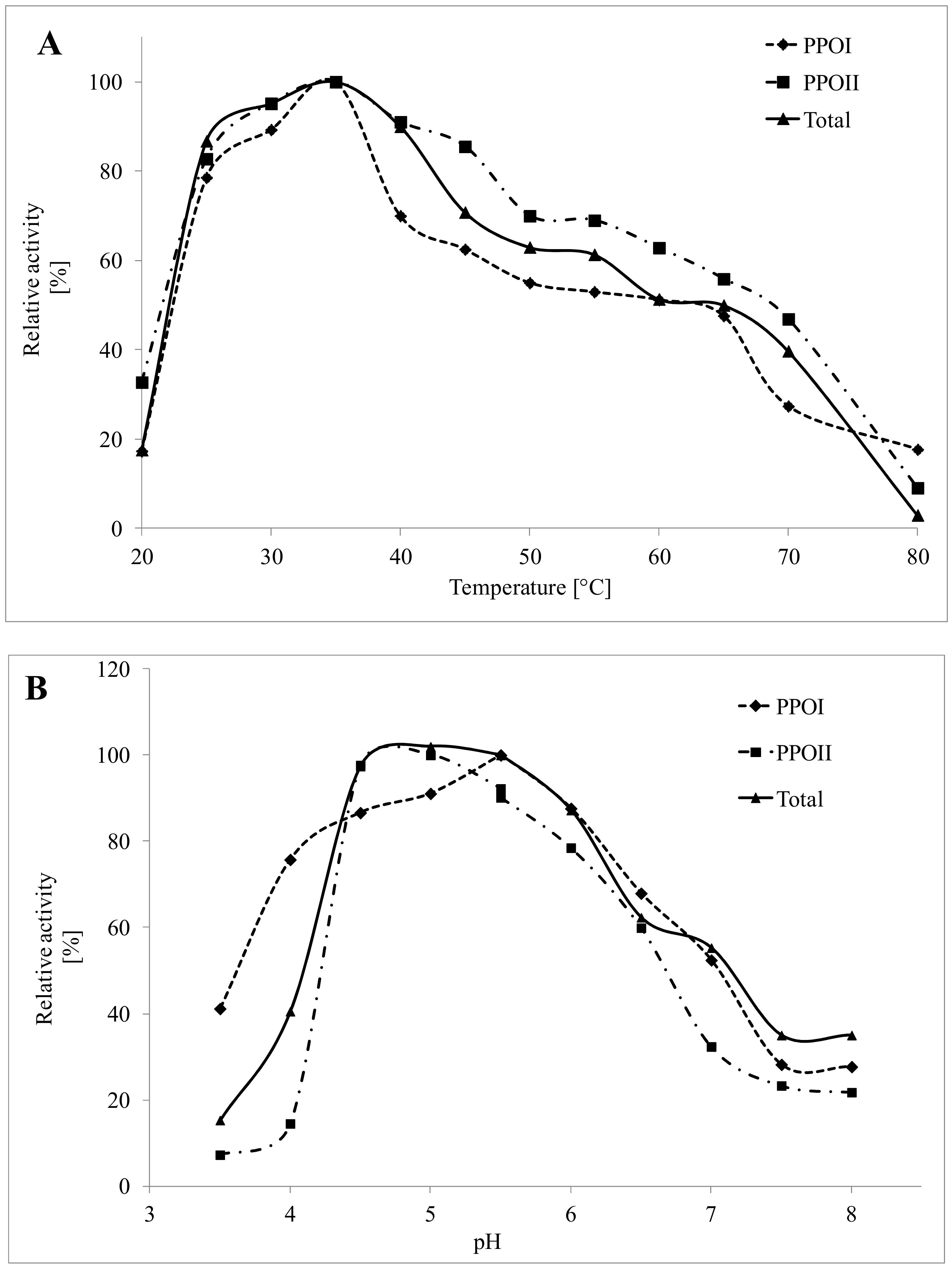
3.3. Effect of Temperature and pH on PPO Activity
3.4. Effect of Various Inhibitors and Metal Ions on PPO Activity
4. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Yoruk, R.; Marshall, M.R. Physicochemical properties and function of plant polyphenol oxidase: A review. J. Food Biochem. 2003, 27, 361–422. [Google Scholar] [CrossRef]
- Mayer, A.M. Polyphenol oxidases in plants and fungi: Going places? A review. Phytochemistry 2006, 67, 2318–2331. [Google Scholar] [CrossRef]
- Vaughn, K.C.; Duke, S.O. Function of polyphenol oxidase in higher plants. Physiol. Plant. 1984, 60, 106–112. [Google Scholar] [CrossRef]
- Chabanet, A.; Goldberg, R.; Catesson, A.M.; Quinet-Szely, M.; Delaunay, A.M.; Faye, L. Characterization and Localization of a Phenoloxidase in Mung Bean Hypocotyl Cell Walls. Plant Physiol. 1994, 106, 1095–1102. [Google Scholar] [CrossRef]
- McDougall, G.J.; Stewart, D.; Morrison, I.M. Cell-wall-bound oxidases from tobacco (Nicotiana tabacum) xylem participate in lignin formation. Planta 1994, 194, 9–14. [Google Scholar] [CrossRef]
- Zhang, Q.; Liu, Y.; He, C.; Zhu, S. Postharvest Exogenous Application of Abscisic Acid Reduces Internal Browning in Pineapple. J. Agric. Food Chem. 2015, 63, 5313–5320. [Google Scholar] [CrossRef]
- Sun, Y.; Zhang, W.; Zeng, T.; Nie, Q.; Zhang, F.; Zhu, L. Hydrogen sulfide inhibits enzymatic browning of fresh-cut lotus root slices by regulating phenolic metabolism. Food Chem. 2015, 177, 376–381. [Google Scholar] [CrossRef]
- Yi, J.H.; Dong, X.L.; Zhu, Z.B. Effect of polyphenol oxidase (PPO) enzymatic inducing factors on non-enzymatic browning of apple polyphenols. Mod. Food Sci. Technol. 2015, 31, 119–127. [Google Scholar]
- Sikora, M.; Świeca, M. Effect of ascorbic acid postharvest treatment on enzymatic browning, phenolics and antioxidant capacity of stored mung bean sprouts. Food Chem. 2018, 239, 1160–1166. [Google Scholar] [CrossRef]
- Sullivan, M.L. Beyond brown: Polyphenol oxidases as enzymes of plant specialized metabolism. Front. Plant Sci. 2014, 5, 783. [Google Scholar] [CrossRef] [PubMed]
- Swieca, M.; Sęczyk, L.; Gawlik-Dziki, U. Elicitation and precursor feeding as tools for the improvement of the phenolic content and antioxidant activity of lentil sprouts. Food Chem. 2014, 161, 288–295. [Google Scholar] [CrossRef]
- Ali, H.M.; El-Gizawy, A.M.; El-Bassiouny, R.E.I.; Saleh, M.A. Browning inhibition mechanisms by cysteine, ascorbic acid and citric acid, and identifying PPO-catechol-cysteine reaction products. J. Food Sci. Technol. 2015, 52, 3651–3659. [Google Scholar] [CrossRef]
- Arias, E.; González, J.; Peiró, J.M.; Oria, R.; Lopez-Buesa, P. Browning prevention by ascorbic acid and 4-hexylresorcinol: Different mechanisms of action on polyphenol oxidase in the presence and in the absence of substrates. J. Food Sci. 2007, 72, 464–470. [Google Scholar] [CrossRef]
- Mishra, B.B.; Gautam, S.; Sharma, A. Purification and characterisation of polyphenol oxidase (PPO) from eggplant (Solanum melongena). Food Chem. 2012, 134, 1855–1861. [Google Scholar] [CrossRef]
- Navarro, J.L.; Tárrega, A.; Sentandreu, M.A.; Sentandreu, E. Partial purification and characterization of polyphenol oxidase from persimmon. Food Chem. 2014, 157, 283–289. [Google Scholar] [CrossRef]
- Gawlik-Dziki, U.; Szymanowska, U.; Baraniak, B. Characterization of polyphenol oxidase from broccoli (Brassica oleracea var. botrytis italica) florets. Food Chem. 2007, 105, 1047–1053. [Google Scholar] [CrossRef]
- Aydemïr, T.; Akkanlı, G. Partial purification and characterisation of polyphenol oxidase from celery root (Apium graveolens L.) and the investigation of the effects on the enzyme activity of some inhibitors. Int. J. Food Sci. Technol. 2006, 41, 1090–1098. [Google Scholar] [CrossRef]
- Gawlik-Dziki, U.; Złotek, U.; Świeca, M. Characterization of polyphenol oxidase from butter lettuce (Lactuca sativa var. capitata L.). Food Chem. 2008, 107, 129–135. [Google Scholar] [CrossRef]
- Świeca, M.; Gawlik-Dziki, U.; Kowalczyk, D.; Złotek, U. Impact of germination time and type of illumination on the antioxidant compounds and antioxidant capacity of Lens culinaris sprouts. Sci. Hortic. (Amsterdam). 2012, 140, 87–95. [Google Scholar] [CrossRef]
- Cho, Y.K.; Ahn, H.K. Purification and characterization of polyphenol oxidase from potato: II. Inhibition and catalytic mechanism. J. Food Biochem. 1999, 23, 593–605. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Guo, L.; Ma, Y.; Shi, J.; Xue, S. The purification and characterisation of polyphenol oxidase from green bean (Phaseolus vulgaris L.). Food Chem. 2009, 117, 143–151. [Google Scholar] [CrossRef]
- Cheema, S.; Sommerhalter, M. Characterization of polyphenol oxidase activity in Ataulfo mango. Food Chem. 2015, 171, 382–387. [Google Scholar] [CrossRef]
- Doǧru, Y.Z.; Erat, M. Investigation of some kinetic properties of polyphenol oxidase from parsley (Petroselinum crispum, Apiaceae). Food Res. Int. 2012, 49, 411–415. [Google Scholar] [CrossRef]
- Nagai, T.; Suzuki, N. Partial purification of polyphenol oxidase from Chinese cabbage Brassica rapa L. J. Agric. Food Chem. 2001, 49, 3922–3926. [Google Scholar] [CrossRef]
- Nagai, T.; Suzuki, N. Polyphenol oxidase from bean sprouts (Glycine max L.). J. Food Sci. 2003, 68, 16–20. [Google Scholar] [CrossRef]
- Ünal, M.Ü. Properties of polyphenol oxidase from Anamur banana (Musa cavendishii). Food Chem. 2007, 100, 909–913. [Google Scholar] [CrossRef]
- Serradell, M.D.L.A.; Rozenfeld, P.A.; Martínez, G.A.; Civello, P.M.; Chaves, A.R.; Añón, M.C. Polyphenoloxidase activity from strawberry fruit (Fragaria x ananassa, Duch., cv Selva): Characterisation and partial purification. J. Sci. Food Agric. 2000, 80, 1421–1427. [Google Scholar] [CrossRef]
- Gómez-López, V.M. Some biochemical properties of polyphenol oxidase from two varieties of avocado. Food Chem. 2002, 77, 163–169. [Google Scholar] [CrossRef]
- Rocha, A.M.C.; Morais, A.M.M. Characterization of polyphenoloxidase (PPO) extracted from ‘Jonagored’ apple. Food Control 2001, 12, 85–90. [Google Scholar] [CrossRef]
- Lee, M.-K. Inhibitory effect of banana polyphenol oxidase during ripening of banana by onion extract and Maillard reaction products. Food Chem. 2007, 102, 146–149. [Google Scholar] [CrossRef]
- Yaǧar, H.; Saǧiroǧlu, A. Partially purification and characterization of polyphenol oxidase of quince. Turkish J. Chem. 2002, 26, 97–103. [Google Scholar]
- Złotek, U.; Gawlik-Dziki, U. Selected biochemical properties of polyphenol oxidase in butter lettuce leaves (Lactuca sativa L. var. capitata) elicited with dl-β-amino-n-butyric acid. Food Chem. 2015, 168, 423–429. [Google Scholar] [CrossRef] [PubMed]
- Gandía-Herrero, F.; García-Carmona, F.; Escribano, J. Purification and characterization of a latent polyphenol oxidase from beet root (Beta vulgaris L.). J. Agric. Food Chem. 2004, 52, 609–615. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Liu, W.; Wang, D.; Zhou, Y.; Lin, X.; Wang, X.; Li, S. Purification and partial characterization of polyphenol oxidase from the flower buds of Lonicera japonica Thunb. Food Chem. 2013, 138, 478–483. [Google Scholar] [CrossRef] [PubMed]
- Aydemir, T. Partial purification and characterization of polyphenol oxidase from artichoke (Cynara scolymus L.) heads. Food Chem. 2004, 87, 59–67. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Total Volume (mL) | Activity (U/mL) | Protein (mg/mL) | Total Activity (U) | Specific Activity (U/mg) | Yield (%) | Purification Fold | ||
|---|---|---|---|---|---|---|---|---|
| Crude extract | 250 | 550 | 732.00 | 137500 | 0.75 | 100.0 | 1.00 | |
| Salting out and dialysis | 55 | 2265 | 644.97 | 124575 | 3.51 | 90.60 | 4.67 | |
| Ion exchange chromatography | PPO I | 13 | 1815 | 92.56 | 23595 | 19.61 | 17.16 | 26.10 |
| PPO II | 19 | 4475 | 237.12 | 85016 | 18.87 | 61.83 | 25.11 | |
| Vmax (U·mL−1·min−1) | Km (mM) | Vmax/Km (U·mL−1·min−1·mM−1) | ||
|---|---|---|---|---|
| 4-methylcatechol | PPO I | 4878 ± 244 | 3.00 ± 0.14 | 1626 |
| PPO II | 3846 ± 192 | 3.40 ± 0.15 | 1131 | |
| Total | 5410 ± 270 | 1.50 ± 0.07 | 3607 | |
| Catechol | PPO I | 952 ± 48 | 1.76 ± 0.08 | 541 |
| PPO II | 1111 ± 56 | 0.94 ± 0.04 | 1176 | |
| Total | 1737 ± 87 | 1.32 ± 0.06 | 1320 | |
| Gallic acid | PPO I | 2817 ± 141 | 2.25 ± 0.10 | 1250 |
| PPO II | 3742 ± 152 | 5.00 ± 0.23 | 769 | |
| Total | 8250 ± 413 | 7.25 ± 0.33 | 1138 | |
| Caffeic acid | PPO I | 769±38 | 3.81 ± 0.17 | 202 |
| PPO II | 0 | 0 | 0 | |
| Total | 0 | 0 | 0 |
| Concentration of Compounds (mM) | % of Inhibition | |||
|---|---|---|---|---|
| PPO I | PPO II | Total | ||
| Ascorbic acid | 20 | 79.66 ± 3.03 | 79.42 ± 1.80 | 62.57 ± 2.38 |
| 2 | 50.85 ± 1.93 | 59.42 ± 2.58 | 43.86 ± 1.67 | |
| 0.2 | 46.61 ± 1.77 | 62.32 ± 0.71 | 24.56 ± 0.93 | |
| l-cysteine | 20 | 71.67 ± 2.72 | 72.09 ± 3.14 | 26.97 ± 1.02 |
| 2 | 68.33 ± 2.60 | 55.81 ± 2.43 | 21.35 ± 0.81 | |
| 0.2 | 56.67 ± 2.15 | 34.88 ± 1.52 | 10.11 ± 0.38 | |
| Na2S2O5 | 20 | 76.03 ± 2.89 | 71.58 ± 3.11 | 25.88 ± 0.98 |
| 2 | 56.20 ± 2.14 | 64.21 ± 2.79 | 14.12 ± 0.54 | |
| 0.2 | 18.18 ± 0.69 | 24.21 ± 1.05 | 1.18 ± 0.04 | |
| EDTA | 20 | 26.67 ± 1.01 | 39.77 ± 1.73 | 14.63 ± 0.56 |
| 2 | 24.17 ± 0.92 | 36.36 ± 1.58 | 3.66 ± 0.14 | |
| 0.2 | 20.83 ± 0.79 | 34.09 ± 1.48 | 2.44 ± 0.09 | |
| Citric acid | 20 | 30.13 ± 1.14 | 60.82 ± 2.65 | 43.18 ± 1.64 |
| 2 | 22.44 ± 0.85 | 54.39 ± 2.37 | 37.88 ± 1.44 | |
| 0.2 | 7.69 ± 0.29 | 57.89 ± 2.52 | 8.33 ± 0.32 | |
| Sodium azide | 20 | 22.50 ± 0.85 | 22.89 ± 1.00 | 9.09 ± 0.35 |
| 2 | 17.50 ± 0.67 | 15.66 ± 0.68 | 8.08 ± 0.31 | |
| 0.2 | 5.83 ± 0.22 | 8.43 ± 0.37 | 2.02 ± 0.08 | |
| Dithiothreitol | 20 | 17.50 ± 0.67 | 37.89 ± 1.65 | 30.26 ± 1.15 |
| 2 | 22.50 ± 0.86 | 31.58 ± 1.37 | 13.16 ± 0.50 | |
| 0.2 | 22.50 ± 0.86 | 15.58 ± 1.72 | 1.32 ± 0.05 | |
| Ion Concentration (mM) | Residual Activity (%) | ||
|---|---|---|---|
| 10 | 2 | ||
| Na+ | PPO I | 75.82 ± 3.26 | 82.42 ± 3.54 |
| PPO II | 90.38 ± 3.89 | 142.31 ± 6.12 | |
| Total | 89.81 ± 3.86 | 106.48 ± 4.58 | |
| K+ | PPO I | 76.99 ± 3.31 | 118.58 ± 5.10 |
| PPO II | 101.35 ± 4.36 | 159.46 ± 6.86 | |
| Total | 99.23 ± 4.27 | 109.46 ± 4.71 | |
| Mg2+ | PPO I | 46.90 ± 2.02 | 50.44 ± 2.17 |
| PPO II | 51.35 ± 2.21 | 52.70 ± 2.27 | |
| Total | 61.54 ± 2.65 | 67.69 ± 2.91 | |
| Zn2+ | PPO I | Nd | 42.24 ± 1.82 |
| PPO II | Nd | 38.42 ± 1.65 | |
| Total | Nd | 48.78 ± 2.10 | |
| Ba2+ | PPO I | Nd | 54.23 ± 2.33 |
| PPO II | Nd | 54.43 ± 2.34 | |
| Total | Nd | 86.05 ± 3.70 | |
| Fe3+ | PPO I | Nd | 53.49 ± 2.30 |
| PPO II | Nd | 56.96 ± 2.45 | |
| Total | Nd | 58.28 ± 2.51 | |
| Mn2+ | PPO I | Nd | 48.98 ± 2.11 |
| PPO II | Nd | 32.92 ± 1.42 | |
| Total | Nd | 27.30 ± 1.17 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sikora, M.; Świeca, M.; Franczyk, M.; Jakubczyk, A.; Bochnak, J.; Złotek, U. Biochemical Properties of Polyphenol Oxidases from Ready-to-Eat Lentil (Lens culinaris Medik.) Sprouts and Factors Affecting Their Activities: A Search for Potent Tools Limiting Enzymatic Browning. Foods 2019, 8, 154. https://doi.org/10.3390/foods8050154
Sikora M, Świeca M, Franczyk M, Jakubczyk A, Bochnak J, Złotek U. Biochemical Properties of Polyphenol Oxidases from Ready-to-Eat Lentil (Lens culinaris Medik.) Sprouts and Factors Affecting Their Activities: A Search for Potent Tools Limiting Enzymatic Browning. Foods. 2019; 8(5):154. https://doi.org/10.3390/foods8050154
Chicago/Turabian StyleSikora, Małgorzata, Michał Świeca, Monika Franczyk, Anna Jakubczyk, Justyna Bochnak, and Urszula Złotek. 2019. "Biochemical Properties of Polyphenol Oxidases from Ready-to-Eat Lentil (Lens culinaris Medik.) Sprouts and Factors Affecting Their Activities: A Search for Potent Tools Limiting Enzymatic Browning" Foods 8, no. 5: 154. https://doi.org/10.3390/foods8050154