

High Growing Temperature Changes Nutritional Value of Broccoli (Brassica oleracea L. convar. botrytis (L.) Alef. var. cymosa Duch.) Seedlings

Abstract
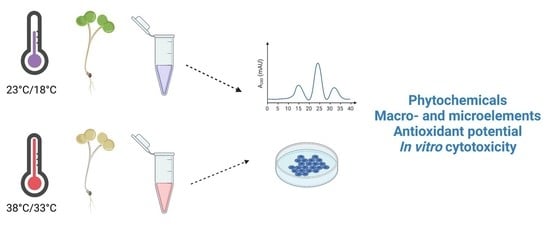
:
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of Extracts
2.3. The Amount of Phenolics, Soluble Sugars, Proteins, Glucosinolates, and Pigments
2.4. RP-HPLC Analysis of Individual Phenolics and L-Ascorbic Acid
2.5. Inductively Coupled Plasma Mass Spectrometry Analysis of Macro- and Microelements
2.6. Antioxidant Capacity
2.7. In Vitro Cytotoxicity
2.7.1. Dimethylthiazol Diphenyltetrazolium Bromide (MTT) Assay
2.7.2. Neutral Red Uptake Assay
2.8. Statistical Analysis
3. Results and Discussion
3.1. Effect of High Growing Temperature on the Amount of Different Groups of Phenolics in Broccoli Seedlings
3.2. Effect of High Growing Temperature on the Amount of Total Glucosinolates, Proteins, and Soluble Sugars
3.3. Effect of High Growing Temperature on Photosynthetic Pigments in Broccoli Seedlings
3.4. Effect of High Growing Temperature on Phenolics and Vitamin C in Control and Stressed Broccoli Seedlings
3.5. Effect of High Growing Temperature on Macro- and Microelements in Broccoli Seedlings
3.6. Effect of High Growing Temperature on Antioxidant Potential of Broccoli Seedlings
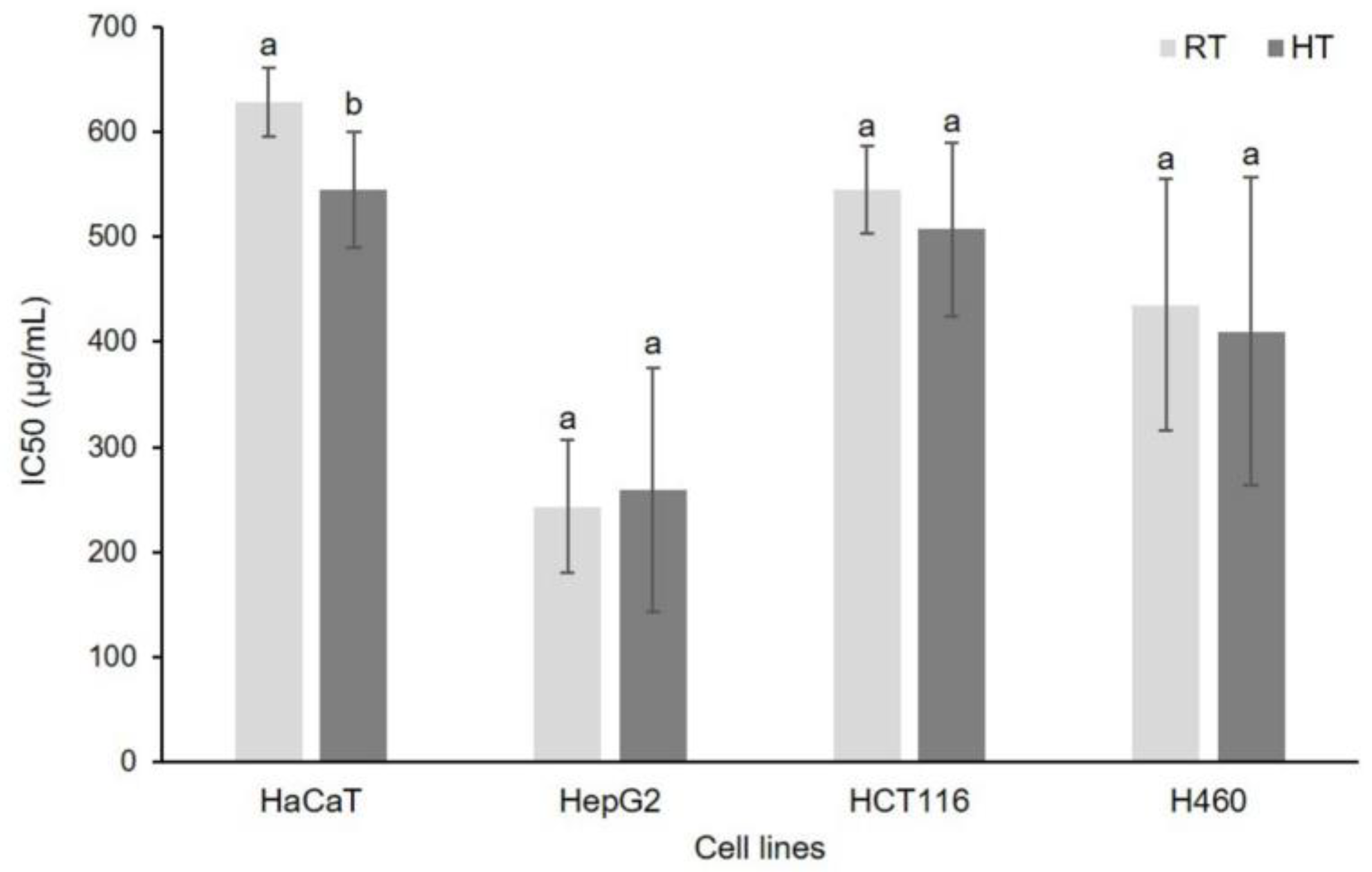
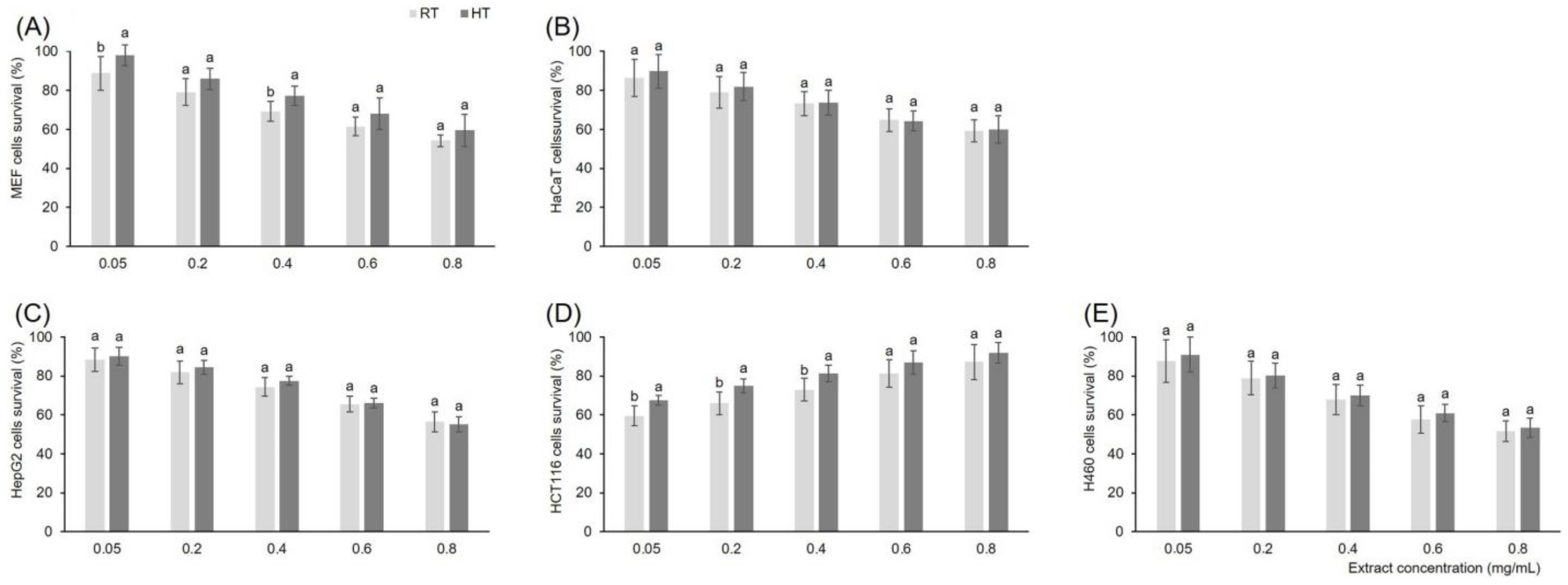
3.7. Effect of High Growing Temperature on In Vitro Cytotoxicity of Broccoli Seedling Extract
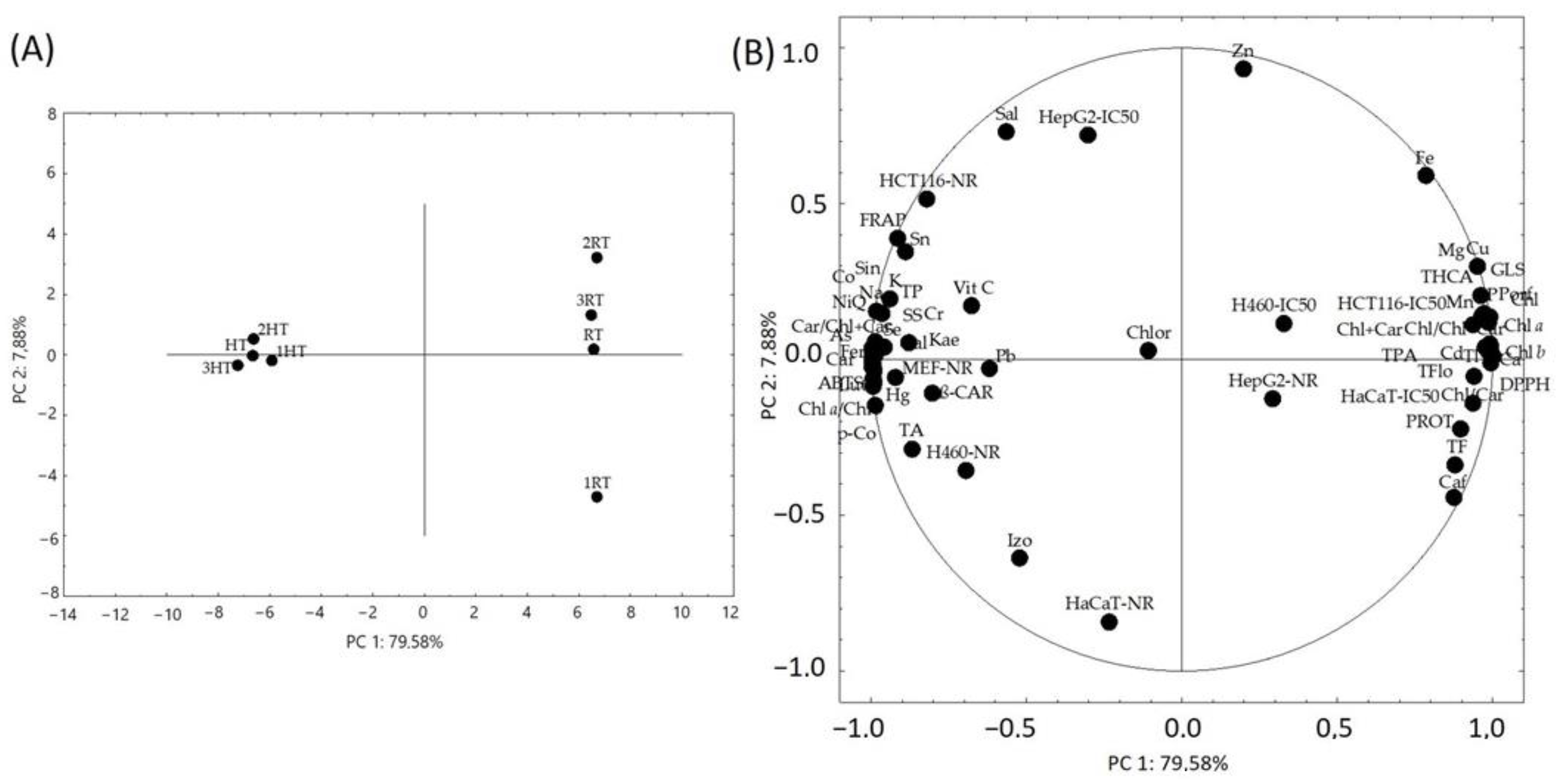
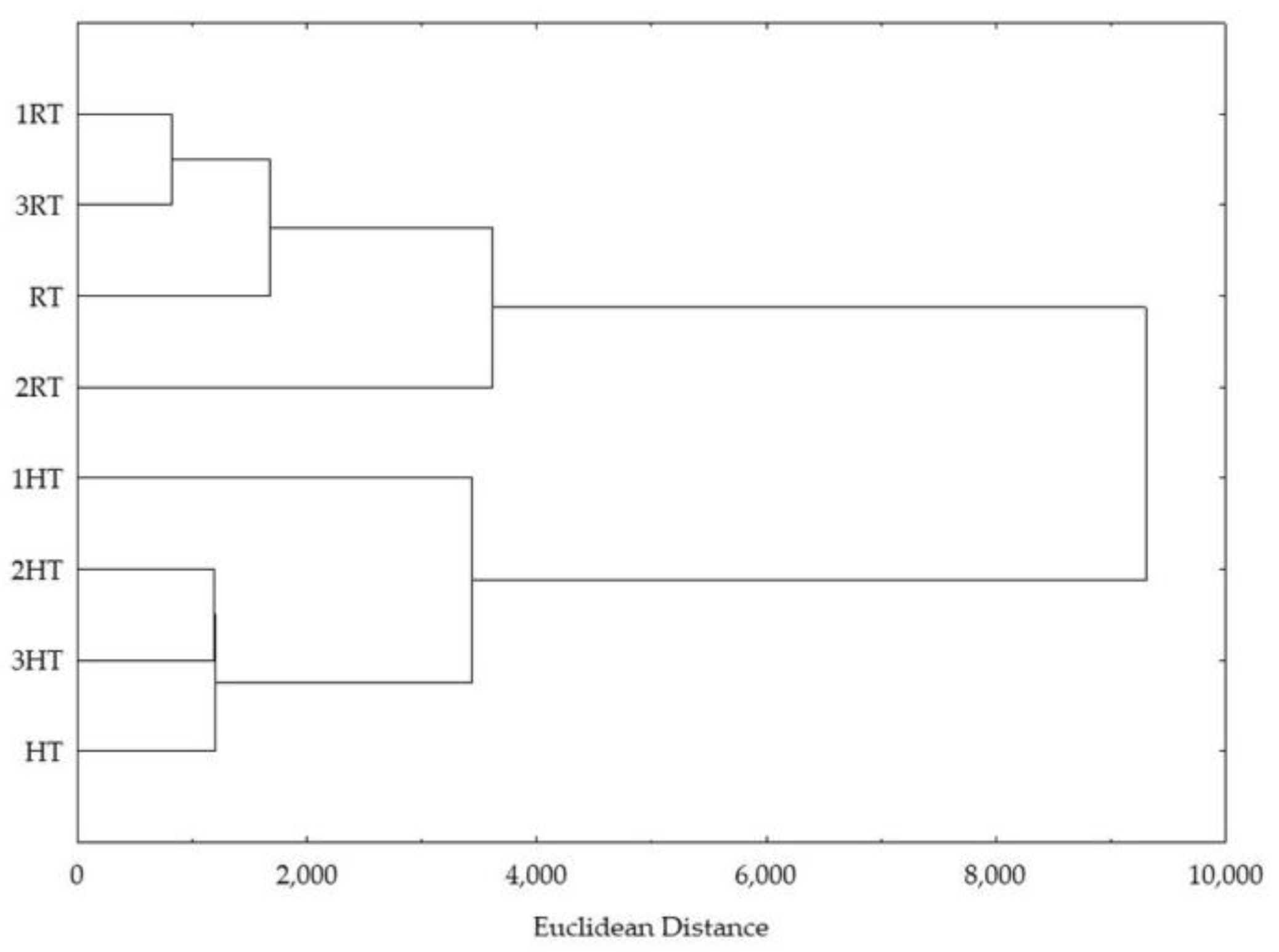
3.8. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zhang, L.; Dai, Y.; Yue, L.; Chen, G.; Yuan, L.; Zhang, S.; Li, F.; Zhang, H.; Li, G.; Zhu, S.; et al. Heat Stress Response in Chinese Cabbage (Brassica rapa L.) Revealed by Transcriptome and Physiological Analysis. PeerJ 2022, 10, e13427. [Google Scholar] [CrossRef]
- Parthasarathi, T.; Firdous, S.; David, E.M.; Lesharadevi, K.; Djanaguiraman, M. Effects of High Temperature on Crops. In Advances in Plant Defense Mechanisms; Kimatu, J.N., Ed.; IntechOpen: London, UK, 2022. [Google Scholar]
- Lazarević, B.; Carović-Stanko, K.; Safner, T.; Poljak, M. Study of High-Temperature-Induced Morphological and Physiological Changes in Potato Using Nondestructive Plant Phenotyping. Plants 2022, 11, 3534. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Xu, H.; Chu, C.; He, F.; Fang, S. High temperature can change root system architecture and intensify root interactions of plant seedlings. Front. Plant. Sci. 2020, 11, 160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jamloki, A.; Bhattacharyya, M.; Nautiyal, M.C.; Patni, B. Elucidating the Relevance of High Temperature and Elevated CO2 in Plant Secondary Metabolites (PSMs) Production. Heliyon 2021, 7, e07709. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Euring, D.; Cha, J.Y.; Lin, Z.; Lu, M.; Huang, L.J.; Kim, W.Y. Plant Hormone-Mediated Regulation of Heat Tolerance in Response to Global Climate Change. Front. Plant. Sci. 2021, 11, 627969. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.; Xiang, Y. An Overview of Biomembrane Functions in Plant Responses to High-Temperature Stress. Front. Plant. Sci. 2018, 9, 915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, S.; Ding, Y.; Zhu, C. Sensitivity and Responses of Chloroplasts to Heat Stress in Plants. Front. Plant. Sci. 2020, 11, 375. [Google Scholar] [CrossRef] [Green Version]
- Rurek, M. Plant Mitochondria under a Variety of Temperature Stress Conditions. Mitochondrion 2014, 19, 289–294. [Google Scholar] [CrossRef]
- Onyebuchi, C.; Kavaz, D. Effect of Extraction Temperature and Solvent Type on the Bioactive Potential of Ocimum gratissimum L. Extracts. Sci. Rep. 2020, 10, 21760. [Google Scholar] [CrossRef]
- Madrau, M.A.; Piscopo, A.; Sanguinetti, A.M.; Del Caro, A.; Poiana, M.; Romeo, F.V.; Piga, A. Effect of Drying Temperature on Polyphenolic Content and Antioxidant Activity of Apricots. Eur. Food Res. Technol. 2009, 228, 441–448. [Google Scholar] [CrossRef]
- Arfaoui, L. Dietary Plant Polyphenols: Effects of Food Processing on Their Content and Bioavailability. Molecules 2021, 26, 2959. [Google Scholar] [CrossRef] [PubMed]
- Rothwell, J.A.; Medina-Remón, A.; Pérez-Jiménez, J.; Neveu, V.; Knaze, V.; Slimani, N.; Scalbert, A. Effects of Food Processing on Polyphenol Contents: A Systematic Analysis Using Phenol-Explorer Data. Mol. Nutr. Food Res. 2015, 59, 160–170. [Google Scholar] [CrossRef] [PubMed]
- İlter, I.; Altay, Ö.; Köprüalan, Ö.; Kaymak-Ertekin, F.; Jafari, S.M. An Overview of High-Temperature Food Processes. In Unit Operations and Processing Equipment in the Food Industry, High-Temperature Processing of Food Product; Jafari, S.M., Ed.; Woodhead Publishing: Sawstone, UK, 2023; pp. 1–43. [Google Scholar]
- Aubert, C.; Bony, P.; Chalot, G.; Landry, P.; Lurol, S. Effects of Storage Temperature, Storage Duration, and Subsequent Ripening on the Physicochemical Characteristics, Volatile Compounds, and Phytochemicals of Western Red Nectarine (Prunus persica L. Batsch). J. Agric. Food Chem. 2014, 62, 4707–4724. [Google Scholar] [CrossRef] [PubMed]
- Mahn, A.; Rubio, M.P. Evolution of Total Polyphenols Content and Antioxidant Activity in Broccoli Florets During Storage at Different Temperatures. J. Food Qual. 2017, 3742183. [Google Scholar] [CrossRef] [Green Version]
- Kohler, I.H.; Huber, S.C.; Bernacchi1, C.J.; Baxter, I.R. Increased Temperatures May Safeguard the Nutritional Quality of Crops under Future Elevated CO2 Concentrations. Plant J. 2019, 97, 872–886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchioni, I.; Martinelli, M.; Ascrizzi, R.; Gabbrielli, C.; Flamini, G.; Pistelli, L.; Pistelli, L. Small Functional Foods: Comparative Phytochemical and Nutritional Analyses of Five Microgreens of the Brassicaceae Family. Foods 2021, 10, 427. [Google Scholar] [CrossRef] [PubMed]
- Cevallos-Casals, B.A.; Cisneros-Zevallos, L. Impact of Germination on Phenolic Content and Antioxidant Activity of 13 Edible Seed Species. Food Chem. 2010, 119, 1485–1490. [Google Scholar] [CrossRef]
- Baenas, N.; Moreno, D.A.; García-Viguera, C. Selecting Sprouts of Brassicaceae for Optimum Phytochemical Composition. J. Agric. Food Chem. 2012, 60, 11409–11420. [Google Scholar] [CrossRef]
- Šola, I.; Vujčić Bok, V.; Dujmović, M.; Rusak, G. Developmentally-Related Changes in Phenolic and L-ascorbic Acid Content and Antioxidant Capacity of Chinese Cabbage Sprouts. J. Food Sci. Technol. 2020, 57, 702–712. [Google Scholar] [CrossRef]
- Ayeni, A. Nutrient Content of Micro/Baby-Green and Field-Grown Mature Foliage of Tropical Spinach (Amaranthus sp.) and Roselle (Hibiscus sabdariffa L.). Foods 2021, 10, 2546. [Google Scholar] [CrossRef]
- Pająk, P.; Socha, R.; Gałkowska, D.; Rożnowski, J.; Fortuna, T. Phenolic Profile and Antioxidant Activity in Selected Seeds and Sprouts. Food Chem. 2014, 143, 300–306. [Google Scholar] [CrossRef] [PubMed]
- Riggio, G.; Jones, S.; Gibson, K. Risk of Human Pathogen Internalization in Leafy Vegetables During Lab-Scale Hydroponic Cultivation. Horticulturae 2019, 5, 25. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Xiao, Z.; Ager, E.; Kong, L.; Tan, L. Nutritional quality and health benefits of microgreens, a crop of modern agriculture. J. Future Foods 2021, 1, 58–66. [Google Scholar] [CrossRef]
- Turner, E.R.; Luo, Y.; Buchanan, R.L. Microgreen nutrition, food safety, and shelf life: A review. J. Food Sci. 2020, 85, 870–882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paradiso, V.M.; Castellino, M.; Renna, M.; Gattullo, C.E.; Calasso, M.; Terzano, R.; Allegretta, I.; Leoni, B.; Caponio, F.; Santamaria, P. Nutritional characterization and shelf-life of packaged microgreens. Food Funct. 2018, 9, 5629. [Google Scholar] [CrossRef] [Green Version]
- Šola, I.; Vujčić Bok, V.; Pinterić, M.; Auer, S.; Ludwig-Müller, J.; Rusak, G. Improving the phytochemical profile and bioactivity of Chinese cabbage sprouts by interspecific transfer of metabolites. Food Res. Int. 2020, 137, 109726. [Google Scholar] [CrossRef]
- Puccinelli, M.; Malorgio, F.; Rosellini, I.; Pezzarossa, B. Production of selenium-biofortified microgreens from selenium-enriched seeds of basil. J. Sci. Food Agric. 2019, 99, 5601–5605. [Google Scholar] [CrossRef]
- Przybysz, A.; Wrochna, M.; Małecka-Przybysz, M.; Gawrońska, H.; Gawroński, S.W. The effects of Mg enrichment of vegetable sprouts on Mg concentration, yield and ROS generation. J. Sci. Food Agric. 2015, 96, 3469–3476. [Google Scholar] [CrossRef]
- Przybysz, A.; Wrochna, M.; Małecka-Przybysz, M.; Gawrońska, H.; Gawroński, S.W. Vegetable sprouts enriched with iron: Effects on yield, ROS generation and antioxidative system. Sci. Hortic. 2016, 203, 110–117. [Google Scholar] [CrossRef]
- Ebert, A.W. Sprouts and Microgreens—Novel Food Sources for Healthy Diets. Plants 2022, 11, 571. [Google Scholar] [CrossRef]
- Choe, U.; Yu, L.L.; Wang, T.T.Y. The science behind microgreens as an exciting new food for the 21st century. J. Agric. Food Chem. 2018, 66, 11519–11530. [Google Scholar] [CrossRef]
- Kyriacou, M.C.; De Pascale, S.; Kyratzis, A.; Rouphael, Y. Microgreens as a component of space life support systems: A cornucopia of functional food. Front. Plant. Sci. 2017, 8, 1–4. [Google Scholar] [CrossRef]
- Sharma, S.; Singh, V.; Tanwar, H.; Mor, V.S.; Kumar, M.; Punia, R.C.; Dalal, M.S.; Khan, M.; Sangwan, S.; Bhuker, A.; et al. Impact of High Temperature on Germination, Seedling Growth and Enzymatic Activity of Wheat. Agriculture 2022, 12, 1500. [Google Scholar] [CrossRef]
- Podsędek, A. Natural Antioxidants and Antioxidant Capacity of Brassica Vegetables: A review. LWT 2007, 40, 1–11. [Google Scholar] [CrossRef]
- Fernández-León, M.F.; Fernández-León, A.M.; Lozano, M.; Ayuso, M.C.; González-Gómez, D. Identification, Quantification and Comparison of the Principal Bioactive Compounds and External Quality Parameters of Two Broccoli Cultivars. J. Funct. Foods 2012, 4, 465–473. [Google Scholar] [CrossRef]
- James, D.; Devaraj, S.; Bellur, P.; Lakkanna, S.; Vicini, J.; Boddupalli, S. Novel Concepts of Broccoli Sulforaphanes and Disease: Induction of Phase II Antioxidant and Detoxification Enzymes by Enhanced-Glucoraphanin Broccoli. Nutr. Rev. 2012, 70, 654–665. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R. Analysis of Total Phenols and Other Oxidation Substrates and Antioxidants by Means of Folin-Ciocalteu Reagent. Methods Enzymol. 1999, 299, 152–178. [Google Scholar]
- Zhishen, J.; Mengcheng, T.; Jianming, W. The Determination of Flavonoid Contents in Mulberry and Their Scavenging Effects on Superoxide Radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Howard, L.R.; Clear, J.R.; Brownmiller, C. Antioxidant Capacity and Phenolic Content in Blueberries as Affected by Genotype and Growing Season. J. Sci. Food Agric. 2003, 83, 1238–1247. [Google Scholar] [CrossRef]
- Council of Europe. European Pharmacopoeia, 4th ed.; Council of Europe: Strasbourg, France, 2004; pp. 2377–2378. [Google Scholar]
- Lee, J.; Durst, R.W.; Wrolstad, E. Determination of Total Monomeric Anthocyanin Pigment Content of Fruit Juices, Beverages, Natural Colorants, and Wines by the Ph Differential Method: Collaborative Study. J. AOAC Int. 2005, 88, 1269–1278. [Google Scholar] [CrossRef] [Green Version]
- Aghajanzadeh, T.; Hawkesford, M.; De Kok, L.J. The Significance of Glucosinolates for Sulfur Storage in Brassicaceae Seedlings. Front. Plant. Sci. 2014, 5, 1–10. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric Method for Determination of Sugars and Related Substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Sumanta, N.; Imranul Haque, C.; Nishika, J.; Suprakash, R. Spectrophotometric Analysis of Chlorophylls and Carotenoids from Commonly Grown Fern Species by Using Various Extracting Solvents. Res. J. Chem. Sci. 2014, 4, 63–69. [Google Scholar]
- Tariba Lovaković, B.; Lazarus, M.; Brčić Karačonji, I.; Jurica, K.; Živković Semren, T.; Lušić, D.; Brajenović, N.; Pelaić, Z.; Pizent, A. Multi-Elemental Composition and Antioxidant Properties of Strawberry Tree (Arbutus unedo L.) Honey from the Coastal Region of Croatia: Risk-Benefit Analysis. J. Trace Elem. Med. Biol. 2018, 45, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Chaillou, L.L.; Nazareno, M.A. New Method to Determine Antioxidant Activity of Polyphenols. J. Agric. Food Chem. 2006, 52, 8397–8402. [Google Scholar] [CrossRef]
- Bita, C.E.; Gerats, T. Plant Tolerance to High Temperature in a Changing Environment: Scientific Fundamentals and Production of Heat Stress-Tolerant Crops. Front. Plant. Sci. 2013, 4. [Google Scholar] [CrossRef] [Green Version]
- Cartea, M.E.; Francisco, M.; Soengas, P.; Velasco, P. Phenolic Compounds in Brassica Vegetables. Molecules 2011, 16, 251–280. [Google Scholar] [CrossRef] [PubMed]
- Šamec, D.; Karalija, E.; Šola, I.; Vujčić Bok, V.; Salopek-Sondi, B. The Role of Polyphenols in Abiotic Stress Response: The Influence of Molecular Structure. Plants 2021, 10, 118. [Google Scholar] [CrossRef] [PubMed]
- Soengas, P.; Rodríguez, V.M.; Velasco, P.; Cartea, M.E. Effect of Temperature Stress on Antioxidant Defenses in Brassica oleracea. ACS Omega 2018, 3, 5237–5243. [Google Scholar] [CrossRef]
- Ragusa, L.; Picchi, V.; Tribulato, A.; Cavallaro, C.; Branca, F. The Effect of the Germination Remperature on the Phytochemical Content of Broccoli and Rocket Sprouts. Int. J. Food Sci. Nutr. 2017, 68, 411–420. [Google Scholar] [CrossRef]
- Alhaithloul, H.A.S.; Galal, F.H.; Seufi, A.M. Effect of Extreme Temperature Changes on Phenolic, Flavonoid Contents and Antioxidant Activity of Tomato Seedlings (Solanum lycopersicum L.). PeerJ 2021, 9, e11193. [Google Scholar] [CrossRef]
- Avato, P.; Argentieri, M.P. Brassicaceae: A Rich Source of Health Improving Phytochemicals. Phytochem. Rev. 2015, 14, 1019–1033. [Google Scholar] [CrossRef]
- Baenas, N.; Gómez-Jodar, I.; Moreno, D.A.; García-Viguera, C.; Periago, P.M. Broccoli and Radish Sprouts are Safe and Rich in Bioactive Phytochemicals. Postharvest Biol. Technol. 2017, 127, 60–67. [Google Scholar] [CrossRef]
- Rodríguez, V.M.; Soengas, P.; Cartea, E.; Sotelo, T.; Velasco, P. Suitability of A European Nuclear Collection of Brassica oleracea L. Landraces to Grow at High Temperatures. J. Agron. Crop. Sci. 2013, 200, 183–190. [Google Scholar] [CrossRef] [Green Version]
- Valente Pereira, F.M.; Rosa, E.; Fahey, J.W.; Stephenson, K.K.; Carvalho, R.; Aires, A. Influence of Temperature and Ontogeny on the Levels of Glucosinolates in Broccoli (Brassica oleracea var. italica) Sprouts and Their Effect on the Induction of Mammalian Phase 2 Enzymes. J. Agric. Food Chem. 2002, 50, 6239–6244. [Google Scholar] [CrossRef]
- Lin, H.; Sun, J.; Hu, Z.; Cheng, C.; Lin, S.; Zou, H.; Yan, X. Variation in Glucosinolate Accumulation among Different Sprout and Seedling Stages of Broccoli (Brassica oleracea var. italica). Plants 2022, 11, 1563. [Google Scholar] [CrossRef]
- Kosová, K.; Vítámvás, P.; Urban, M.O.; Prášil, I.T.; Renaut, J. Plant Abiotic Stress Proteomics: The Major Factors Determining Alterations in Cellular Proteome. Front. Plant. Sci. 2018, 9, 122. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.Y.; Yang, S.L.; Li, J.Y.; Ma, J.H.; Pang, T.; Zou, C.M.; He, B.; Gong, M. Effects of Different Growth Temperatures on Growth, Development, and Plastid Pigments Metabolism of Tobacco (Nicotiana tabacum L.) Plants. Bot. Stud. 2018, 59, 5. [Google Scholar] [CrossRef] [Green Version]
- Choukri, H.; El Haddad, N.; Aloui, K.; Hejjaoui, K.; El-Baouchi, A.; Smouni, A.; Thavarajah, D.; Maalouf, F.; Kumar, S. Effect of High Temperature Stress During the Reproductive Stage on Grain Yield and Nutritional Quality of Lentil (Lens culinaris Medikus). Front. Nutr. 2022, 9, 857469. [Google Scholar] [CrossRef]
- Gulen, H.; Eris, A. Effect of Heat Stress on Peroxidase Activity and Total Protein Content in Strawberry Plants. Plant. Sci. 2004, 166, 739–744. [Google Scholar] [CrossRef]
- He, Y.; Liu, X.; Huang, B. Changes in Protein Content, Protease Activity, and Amino Acid Content Associated with Heat Injury in Creeping Bentgrass. J. Am. Soc. Hort. Sci. 2005, 130, 842–847. [Google Scholar] [CrossRef]
- Chauhan, J.; Srivastava, J.P.; Singhal, R.K.; Soufan., W.; Dadarwal, B.K.; Mishra, U.N.; Anuragi, H.; Rahman, M.A.; Sakran, M.I.; Brestic, M.; et al. Alterations of Oxidative Stress Indicators, Antioxidant Enzymes, Soluble Sugars, and Amino Acids in Mustard [Brassica juncea (L.) Czern and Coss.] in Response to Varying Sowing Time, and Field Temperature. Front. Plant. Sci. 2022, 13, 875009. [Google Scholar] [CrossRef] [PubMed]
- Sung, D.Y.; Kaplan, F.; Lee, K.J.; Guy, C.L. Acquired Tolerance to Temperature Extremes. Trends Plant. Sci. 2003, 8, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Palta, J.P. Leaf chlorophyll content. Remote Sens. Rev. 1990, 5, 207–213. [Google Scholar] [CrossRef]
- Lefsrud, M.G.; Kopsell, D.A.; Kopsell, D.E.; Curran-Celentano, J. Air Temperature Affects Biomass and Carotenoid Pigment Accumulation in Kale and Spinach Grown in a Controlled Environment. Hort Sci. 2005, 40, 2026–2030. [Google Scholar] [CrossRef] [Green Version]
- Cui, L.; Li, J.; Fan, Y.; Xu, S.; Zhang, Z. High temperature effects on photosynthesis, PSII functionality and antioxidant activity of two Festuca arundinacea cultivars with different heat susceptibility. Bot. Stud. 2006, 47, 61–69. [Google Scholar]
- Di Bella, M.C.; Niklas, A.; Toscano, S.; Picchi, V.; Romano, D.; Lo Scalzo, R.; Branca, F. Morphometric Characteristics, Polyphenols and Ascorbic Acid Variation in Brassica oleracea L. Novel Foods: Sprouts, Microgreens and Baby Leaves. Agronomy 2020, 10, 782. [Google Scholar] [CrossRef]
- Mahla, R.; Madan, S.; Munjal, R.; Chawla, S.; Dua, Y.; Arora, V. Changes in Protein Profile, Ascorbic Acid and Chlorophyll Stability Index of Wheat (Triticum aestivum L.) Seedlings under Heat Stress and Revival Conditions. J. Wheat Res. 2011, 3, 18–21. [Google Scholar]
- Jan, R.; Kim, N.; Lee, S.H.; Khan, M.A.; Asaf, S.; Park, J.R.; Asif, S.; Lee, I.J.; Kim, K.M. Enhanced Flavonoid Accumulation Reduces Combined Salt and Heat Stress Through Regulation of Transcriptional and Hormonal Mechanisms. Front. Plant. Sci. 2021, 12, 796956. [Google Scholar] [CrossRef]
- Tian, C.; Liu, X.; Chang, Y.; Wang, R.; Lv, T.; Cui, C.; Liu, M. Investigation of the Anti-Inflammatory and Antioxidant Activities of Luteolin, Kaempferol, Apigenin and Quercetin. S. Afr. J. Bot. 2021, 137, 257–264. [Google Scholar] [CrossRef]
- Akin, D.E.; Fales, S.L.; Rigsby, L.L.; Snook, M.E. Temperature Effects on Leaf Anatomy, Phenolic Acids, and Tissue Digestibility in Tall Fescue. Agron. J. 1987, 79, 271. [Google Scholar] [CrossRef]
- Laddomada, B.; Blanco, A.; Mita, G.; D’Amico, L.; Singh, R.P.; Ammar, K.; Crossa, J.; Guzmán, C. Drought and Heat Stress Impacts on Phenolic Acids Accumulation in Durum Wheat Cultivars. Foods 2021, 10, 2142. [Google Scholar] [CrossRef]
- Tovar, J.C.; Berry, J.C.; Quillatupa, C.; Castillo, S.E.; Acosta-Gamboa, L.; Fahlgren, N.; Gehan, M.A. Heat Stress Changes Mineral Nutrient Concentrations in Chenopodium quinoa Seed. Plant. Direct 2022, 6, e384. [Google Scholar] [CrossRef]
- Collado-González, J.; Piñero, M.C.; Otálora, G.; López-Marín, J.; del Amor, F.M. Merging Heat Stress Tolerance and Health-Promoting Properties: The Effects of Exogenous Arginine in Cauliflower (Brassica oleracea var. botrytis L.). Foods 2021, 10, 30. [Google Scholar] [CrossRef]
- Floegel, A.; Kim, D.O.; Chung, S.J.; Koo, S.I.; Chun, O.K. Comparison of ABTS/DPPH Assays to Measure Antioxidant Capacity in Popular Antioxidant-Rich US Foods. J. Food Compost. Anal. 2011, 24, 1043–1048. [Google Scholar] [CrossRef]
- Shalaby, E.; Shanab, S.M.M. Comparison of DPPH and ABTS Assays for Determining Antioxidant Potential of Water and Methanol Extracts of Spirulina platensis. Indian J. Mar. Sci. 2013, 42, 556–564. [Google Scholar]
- Chaudhary, A.; Rampal, G.; Sharma, U.; Thind, T.S.; Singh, B.; Vig, A.P.; Arora, S. Anticancer, Antioxidant Activities and GC-MS Analysis of Glucosinolates in Two Cultivars of Broccoli. Med. Chem. Drug Discov. 2012, 2, 30–37. [Google Scholar]
- Atjanasuppat, K.; Wongkham, W.; Meepowpan, P.; Kittakoop, P.; Sobhon, P.; Bartlett, A.; Whitfield, P.J. In vitro Screening for Anthelmintic and Antitumour Activity of Ethnomedicinal Plants from Thailand. J. Ethnopharmacol. 2009, 123, 475–482. [Google Scholar] [CrossRef] [PubMed]
- Paśko, P.; Zagrodzki, P.; Okoń, K.; Prochownik, E.; Krośniak, M.; Galanty, A. Broccoli Sprouts and Their Influence on Thyroid Function in Different In Vitro and In Vivo Models. Plants 2022, 11, 2750. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Room Temperature | High Temperature | Δ (%) | |
|---|---|---|---|
| TP (mg GAE/g DW) | 17.53 ± 2.04 b | 25.49 ± 1.51 a | +45 |
| TF (mg QE/g DW) | 20.91 ± 0.98 a | 19.12 ± 3.63 b | −9 |
| TFlo (mg QE/g DW) | 10.65 ± 3.22 a | 8.15 ± 1.61 b | −23 |
| THCA (mg CAE/g DW) | 11.86 ± 2.11 a | 10.21 ± 1.67 b | −14 |
| TPA (mg CAE/g DW) | 7.59 ± 0.57 a | 5.90 ± 0.95 b | −22 |
| TA (mg C-gluE/g DW) | 11.60 ± 5.90 a | 16.40 ± 5.92 a | +41 |
| GLS (mg SINE/g DW) | 88.91 ± 6.24 a | 53.72 ± 7.78 b | −40 |
| PROT (mg BSAE/g DW) | 57.56 ± 7.19 a | 51.86 ± 5.98 b | −10 |
| SS (mg SUCE/g DW) | 9.41 ± 7.69 b | 29.75 ± 6.05 a | +216 |
| mg/g DW | Room Temperature | High Temperature | Δ (%) |
|---|---|---|---|
| Chl a | 5.07 ± 0.34 a | 4.38 ± 0.55 b | −14 |
| Chl b | 5.88 ± 0.45 a | 2.36 ± 0.76 b | −60 |
| Chl a + Chl b | 10.94 ± 0.46 a | 6.74 ± 1.30 b | −38 |
| Chl a + Chl b + Car | 11.12 ± 0.41 a | 7.69 ± 1.27 b | −31 |
| Chl a/b | 0.86 ± 0.09 b | 2.01 ± 0.41 a | +134 |
| Chl/(Chl + Car) | 0.98 ± 0.02 a | 0.87 ± 0.03 b | −11 |
| Car/(Chl + Car) | 0.02 ± 0.01 b | 0.13 ± 0.03 a | +539 |
| Chl/Car | 43.32 ± 12.39 a | 7.11 ± 1.61 b | −84 |
| Car | 0.23 ± 0.10 b | 0.96 ± 0.05 a | +307 |
| Por | 21.95 ± 0.96 a | 13.00 ± 2.56 b | −41 |
| mg/g DW | Room Temperature | High Temperature | Δ (%) |
|---|---|---|---|
| L-ascorbic acid | 1.32 ± 0.03 a | 1.41 ± 0.12 a | +6 |
| Kaempferol | 2.13 ± 0.60 a | 2.48 ± 1.10 a | +16 |
| Luteolin | 0.39 ± 0.01 b | 0.06 ± 0.00 a | +60 |
| Isorhamnetin | 0.06 ± 0.03 a | 0.06 ± 0.01 a | +6 |
| Quercetin | 0.18 ± 0.11 b | 0.82 ± 0.25 a | +390 |
| Total identified flavonoids | 2.40 ± 0.68 a | 3.43 ± 1.35 a | +43 |
| Sinapic acid | 3.57 ± 0.34 b | 4.25 ± 0.14 a | +19 |
| Ferulic acid | 0.55 ± 0.10 b | 1.46 ± 0.15 a | +168 |
| Chlorogenic acid | 0.10 ± 0.02 a | 0.10 ± 0.02 a | 0 |
| p-Coumaric acid | 0.25 ± 0.05 b | 0.35 ± 0.10 a | +42 |
| Gallic acid | 2.61 ± 0.93 b | 5.03 ± 0.41 a | +93 |
| Salicylic acid | 0.03 ± 0.00 a | 0.03 ± 0.00 a | +8 |
| Caffeic acid | 0.15 ± 0.05 a | 0.12 ± 0.04 a | −20 |
| Total identified phenolic acids | 7.14 ± 0.84 b | 11.34 ± 0.34 a | +57 |
| Total identified phenolics | 9.64 ± 1.52 b | 14.77 ± 1.57 a | +53 |
| Room Temperature | High Temperature | Δ (%) | |
|---|---|---|---|
| As (µg/kg DW) | 201.31 ± 4.42 b | 786.70 ± 6.56 a | +291 |
| Ca (g/kg DW) | 16.59 ± 0.26 a | 10.65 ± 0.47 b | −36 |
| Cd (µg/kg DW) | 122.63 ± 4.57 a | 86.39 ± 4.71 b | −30 |
| Co (µg/kg DW) | 59.29 ± 4.72 b | 197.76 ± 3.12 a | +234 |
| Cr (µg/kg DW) | 97.25 ± 13.99 b | 199.01 ± 47.42 a | +105 |
| Cu (mg/kg DW) | 7.07 ± 0.32 a | 5.86 ± 0.01 b | −17 |
| Fe (mg/kg DW) | 145.78 ± 13.61 a | 124.46 ± 1.66 a | −15 |
| Hg (µg/kg DW) | 14.41 ± 0.87 b | 24.31 ± 0.92 a | +69 |
| K (g/kg DW) | 67.10 ± 2.51 b | 78.22 ± 2.41 a | +17 |
| Mg (g/kg DW) | 5.45 ± 0.18 a | 4.52 ± 0.11 b | −17 |
| Mn (mg/kg DW) | 49.30 ± 1.22 a | 40.59 ± 0.98 b | −18 |
| Na (g/kg DW) | 10.81 ± 0.35 b | 14.52 ± 0.30 a | +34 |
| Ni (µg/kg DW) | 249.95 ± 14.24 b | 818.15 ± 50.39 a | +227 |
| P (g/kg DW) | 6.19 ± 0.36 a | 3.13 ± 0.07 b | −49 |
| Pb (µg/kg DW) | 51.08 ± 7.42 b | 138.08 ± 97.95 a | +170 |
| Se (µg/kg DW) | 50.39 ± 2.88 b | 99.21 ± 7.31 a | +97 |
| Sn (µg/kg DW) | 94.58 ± 11.47 b | 123.43 ± 5.67 a | +31 |
| Tl (µg/kg DW) | 766.41 ± 42.20 a | 326.03 ± 4.54 b | −57 |
| Zn (mg/kg DW) | 54.70 ± 2.48 a | 54.01 ± 0.73 a | −1 |
| Inhibition % | Room Temperature | High Temperature | Δ (%) |
|---|---|---|---|
| ABTS | 57.18 ± 5.18 b | 93.64 ± 2.24 a | +64 |
| DPPH | 88.23 ± 0.87 a | 58.41 ± 3.19 b | −34 |
| FRAP | 97.74 ± 0.56 b | 99.00 ± 0.14 a | +1 |
| β-carotene | 7.55 ± 3.34 b | 13.07 ± 4.85 a | +73 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gmižić, D.; Pinterić, M.; Lazarus, M.; Šola, I. High Growing Temperature Changes Nutritional Value of Broccoli (Brassica oleracea L. convar. botrytis (L.) Alef. var. cymosa Duch.) Seedlings. Foods 2023, 12, 582. https://doi.org/10.3390/foods12030582
Gmižić D, Pinterić M, Lazarus M, Šola I. High Growing Temperature Changes Nutritional Value of Broccoli (Brassica oleracea L. convar. botrytis (L.) Alef. var. cymosa Duch.) Seedlings. Foods. 2023; 12(3):582. https://doi.org/10.3390/foods12030582
Chicago/Turabian StyleGmižić, Daria, Marija Pinterić, Maja Lazarus, and Ivana Šola. 2023. "High Growing Temperature Changes Nutritional Value of Broccoli (Brassica oleracea L. convar. botrytis (L.) Alef. var. cymosa Duch.) Seedlings" Foods 12, no. 3: 582. https://doi.org/10.3390/foods12030582