A Comprehensive Understanding of Electro-Fermentation
by
 , , ,
, , ,
Drishti Dinesh Bhagchandanii
1,†, 
Rishi Pramod Babu
1,†,
Jayesh M. Sonawane
2 ,
,
Namita Khanna
3,
Soumya Pandit
4,* ,
,
Dipak A. Jadhav
5 ,
,
Santimoy Khilari
6 and
Ram Prasad
7,* 1
Amity Institute of Biotechnology, Amity University, Mumbai-Pune Expressway, Panvel, Mumbai, Maharashtra 410206, India
2
Departments of Chemical Engineering and Applied Chemistry, University of Toronto, Toronto, ON M5T 3A1, Canada
3
Department of Biotechnology, Birla Institute of Technology and Science, Dubai Campus, Dubai 345055, UAE
4
Department of Life Sciences, School of Basic Sciences and Research, Sharda University, Greater Noida 201306, India
5
Department of Agricultural Engineering, Maharashtra Institute of Technology, Aurangabad 431010, India
6
Department of Chemistry, Guru Ghasidas University, Bilaspur 495009, India
7
Department of Botany, Mahatma Gandhi Central University, Motihari 845401, India
*
Authors to whom correspondence should be addressed.
†
Both the authors have contributed equally to the work.
Fermentation 2020, 6(3), 92; https://doi.org/10.3390/fermentation6030092
Submission received: 17 July 2020
/
Revised: 13 September 2020
/
Accepted: 14 September 2020
/
Published: 21 September 2020
(This article belongs to the Special Issue Biorefineries)
Abstract
:Electro-fermentation (EF) is an upcoming technology that can control the metabolism of exoelectrogenic bacteria (i.e., bacteria that transfer electrons using an extracellular mechanism). The fermenter consists of electrodes that act as sink and source for the production and movement of electrons and protons, thus generating electricity and producing valuable products. The conventional process of fermentation has several drawbacks that restrict their application and economic viability. Additionally, metabolic reactions taking place in traditional fermenters are often redox imbalanced. Almost all metabolic pathways and microbial strains have been studied, and EF can electrochemically control this. The process of EF can be used to optimize metabolic processes taking place in the fermenter by controlling the redox and pH imbalances and by stimulating carbon chain elongation or breakdown to improve the overall biomass yield and support the production of a specific product. This review briefly discusses microbe-electrode interactions, electro-fermenter designs, mixed-culture EF, and pure culture EF in industrial applications, electro methanogenesis, and the various products that could be hence generated using this process.
1. Introduction
Fermentation has been the backbone of the food transformation industry to increase conservation time and alter production, limiting components to have a higher yield like the texture properties of foods and the organoleptic properties for a long time. However, in recent times fermentation is not subjected to only food transformation; it has become the leading biotechnological platform for generating of a variety of products starting from foods which have been fermented (yogurt, soy, bread), aromas, whole microbial cells (probiotics and starter cultures), solvents, enzymes, vitamins (B2, B12, C) as well as biopolymers like PHA, PHB. In industrial fermentations, several restrictions affect their application as well as increase their operational cost. Most of all, the upstream operations, including the procurement of purified substances by meticulous agro-industrial chains and maintenance of sterile conditions, be it for the inoculum culture or the fermenter, is both time consuming and a capital-intensive task. This can sum up to 50% of the total cost including downstream processing to make the conventional fermentation transition from expensive and time-consuming to more economically viable options. One approach regulates metabolic pathways in microorganisms, most metabolic pathways of significant microbes, and their strains have been studied [1]. In most cases, the culture media that is prepared is highly selective of the pathway and involves a highly optimized strain. By making some adjustments in the nitrogen source, minerals, buffers, chelation factors, and antifoaming microbial metabolism can be controlled. However, these components are the reason for the notable total production cost if not optimized [2].
It is to be noted that a significant feature of microorganisms is the metabolic activities undertaken by them. Apart from their other physicochemical attributes such as the ability to tolerate high and low temperatures, electroactive microorganisms have well established functional capabilities in utilizing electrodes as electron mediators for their metabolic processes. It is frequently observed that most of the anaerobic fermentation (AF) pathways are redox imbalanced along with specific setbacks in selecting products and the efficiency to convert a substrate of interest to product [3]. The process is heavily influenced by many factors that affect the reaction rate and efficiency of conversion from a given substrate. To solve the problems that limit the production of higher yield generation, discussion of diverse ideas that focus on innovative strategies that facilitate the stoichiometric equilibration inside the bioreactor using electrodes and electroactive bacteria [4]. Apart from the necessity to regulate the energy input and ionic utilization throughout the fermentation process, there is an additional need for neutralizing energetics for product generation while minimizing the losses. Air is sparged inside the inoculum culture to act as an electron acceptor to neutralize the redox equivalents of the microbe’s metabolism [5]. In anaerobic fermentation, nitrogen is utilized to reduce the collection of gaseous metabolites.
On the other hand, when extra electron donors are required, H2 gas is bubbled to increase reductive metabolism [6]. Limitations in mass transfer efficiency because of less solubility in the gas phase are major drawbacks of micro-bubbling [7]. To increase the gas phase’s solubility, the fermenters are subjected to high pressure, contributing to an increased total cost [8]. Along with these constraints, the idea of using chemicals to manipulate the medium conditions by using acids, bases, chelating agents, buffers, and antifoam agents keeps adding to the total operational cost. A medium’s pH can be altered by adding acids or bases to the culture media [9]. These modifications result in another concern of monitoring the osmotic conditions and salinity of the solutions, so the pH changes should not influence these conditions due to accumulations of ions. Isolation of soluble metabolites, impurities, and other secondary compounds from culture media is difficult. There is a possibility that the product itself can inhibit the growth of bacteria and its metabolism [10]. The natural evolution of microbes to alter enzymes and their pathways are used to convert carbon-based biomass into useful products. Without being concerned about the purity of the substrate (waste), the total capacity of the yield can be exploited effectively through engineering systems and various strategies to manage the nature of the culture. High temperature and pH salinity or physiochemical aspect of the fermenter can ensure culture purity, such as using extremophiles like hyperthermophile Thermotoga neapolitana which can guarantee culture purity even though the substrate is not sterile. Additionally, when it comes to the separation of metabolic channels and to promote the possibility of controlling the metabolic mechanisms, the use of extremophiles becomes the key [11]. All bioconversions of substrates that are not pure are infected by a huge uncontrollable population of microorganisms and redox conditions, resulting in low product selectivity and process stability [12]. As an alternative, electrostimulation can be another way to replace the conventional techniques in the optimization of the pure cultures for industrial processes and the biorefinery [6].
1.1. The Origin of Bioelectrochemical Systems and Electrofermentaion
In bioelectrochemical systems (BES), there is a bi-directional electron transfer between the microorganisms or biomacromolecules that catalyze exchange processes [13]. BESs have been studied since the late 20th century, and considerable advancements have been made so far (Figure 1). The introduction of microbial fuel cells for the synthesis of electric power production propelled further research in optimization and developing current microbial electrosynthesis technology (MET) [14]. The generation of value-added chemical compounds like hydrogen is triggered by electric currents in different systems called microbial electrolysis cells (MECs). In comparing microbial fuel cells MFCs and MECs, both systems involve exoelectrogenic bacteria like the Geobacter and Shewanella that create an anodic biofilm that can transfer electrons generated during the metabolic processes to an external anode, which is known as the bioanode [15]. An electric circuit connected to the cathode transfers these electrons from the anode to the cathode where the reaction occurs. A new profound focus came into studying the microbial biofilm formation in the cathode surfaces (biocathode) on examining the microbial electrosynthesis. It was found to reduce CO2 to compounds of interest where the electrochemical system is the only electron donor [16]. Two processes were unique, which was discovered in a research study about the microbe-electrode interaction with the primary focus on the electrochemical interaction between the cathodic terminal and the microbes [17]. The electrons are transferred into the organism through catalysis by the outer membrane cytochromes in one process [18]. In the other approach, enzymes such as hydrogenases catalyze the diffusion into the cytoplasm, which acts as a mediator and transfers the electrons and protons devoid of the ATP generation processes get directly into the nicotinamide adenine dinucleotide (NAD) [19].
One of the most widely studied models is the microbial fuel cell, wherein the microorganism facilitates electron mobility to the MFC anode post oxidation with electron donors, creating an electron flow towards the cathode (Figure 2) [20]. Additionally, a few microorganisms are known for their reducing electron acceptors like nitrate and metal ions at the cathode [21]. On the other hand, a bio-electrical system (BES) can initiate microbial electrosynthesis becoming a microbial electrosynthesis cell. This promising electron transport integrates fundamental research in microbiology, electrochemistry, and material science with applications in sustainable energy production and bio-inspired materials development [22]. Enzymatic electrocatalysis was the first BES model to be widely recognized, contributing to advancing amperometric biosensors in the late 20th century. This model discussed the electron extraction from enzymatic reactions by either direct tunneling or indirect via redox mediator transport to a solid-state electrode at which a current is detected [23]. When the redox mediators are absent, electron tunneling in enzymes occurs only across minute distances of a few nanometers. Generally, the redox centers of these enzymes, and the relay centers such as sulfur-iron clusters, are firmly embedded in an insulating protein matrix where the electron transport to a solid-state electrode is restricted [24]. Immobilization of enzymes on electrodes was needed to facilitate direct electron transfer for practical applications. Additionally, the 3D structure of an enzyme is necessary to study its catalytic activity. Many techniques for immobilizing enzymes are prevalent, including the direct immobilization and utilization of conductive support or enzyme–cofactor–mediator complexes to facilitate attachment [25]. Although the attachment increases the durability and efficacy of the enzymatic electron transport, they last for only a short period of time. Unlike isolated enzymes, a few microorganisms called electrochemically active bacteria (EAB) can undergo self-renewal to overcome incompatibility between biological and synthetic surfaces. Extracellular electron transport is the key process that links solid-state electron donors or acceptors to the microorganisms. These extracellular electron transfer systems are discussed briefly later in this paper.
1.2. Electro-Fermentation and its Principles
Electro-fermentation (EF) is a fairly new field that can manipulate microbe metabolism by utilizing solid-state electrodes as electron mediators wherein the anodic terminal is used as an electron acceptor and the cathodic terminal as an electron donor (Figure 2) [10]. All the electrochemical redox reactions can be catalyzed by incorporating a microbial environment with electrochemistry, which the EF approach is known for [27]. The relationship between electrochemical and biochemical reactions together helps in inhibiting the thermochemical limitations of conventional AF. Thereby driving it to a redox-balancing and a simultaneous regulation of electro-metabolic routes resulting in specific biobased products [28]. It is called an electro fermentation because electrodes in a bioreactor, which acts as an electron source or a sink. This helps in the control of electron transfer kinetics. The development of specific strains and drastically increasing the range of metabolic pathways and influence many microbial processes can be done using electrochemical interferences, which is not the same in the traditional fermentation methods such as stirred type bioreactors, photobioreactors, and airlift bioreactors [29]. A significant increase in the synthesis of fatty acids, biogas, and platform chemicals was observed due to the regulated usage of reducing ions at the cathode that is aided by the electron metabolism. Electro fermentation can help stabilizing and optimize fermentation metabolisms through controlled redox and pH irregularities, for instance, the substrate purity and the by-product formation. Additionally, it upregulates the breakdown of the carbon chain or the elongation of various oxidative and reductive conditions [30]. Electro-fermentation also aids in a higher adenosine triphosphate generation, which results in a higher yield of biomass. EF also directs the fermentation mainly to enhance the production of one specific product, as ethanol converted from glycerol [6].
In a basic bioelectrochemical system, the electrodes synergize with microorganisms by exchanging electrons in different configurations, as shown in (Figure 3), which can then be used for electricity generation, treatment of waste effluents in wastewater, bioremediation, and generation of high-value products [3]. Almost all microbes need an electron mediator. Organic or inorganic substances found in the atmosphere usually function as a donor and the roles of the acceptor [6]. Microorganisms can utilize inorganic compounds as lithotrophic, lithoheterotrophic, or lithoautotrophic, where the electron donors and acceptors are used from organic compounds or carbon dioxide from a carbon source [31]. Through direct electrochemical stimulation via the solid electroconductive material, electrons exchange with microorganisms in traditional microbial electrochemical technology [32]. The lithotrophic organisms have shown potential utilize solid conductors to function as electron donors or acceptors to some extent. Electron transfer is facilitated by nanowires, cytochromes, nicotinamide adenine dinucleotide (NAD), and other membrane-bound enzymes or the most important extracellular electron transfer (EET) and the transmembrane transport [33]. Soluble redox mediators like neutral red, riboflavin, play an important role in mediating the EET and transmembrane transport to the cytoplasm apart from using electroconductive materials for exchange electrons with direct electrochemical stimulations [14]. Redox mediators like reduced neutral red, which replaced the use of menaquinone in the fumarate reductase complex were utilized in a pure and mixed culture of hydrogen-utilizing microorganisms for the metabolic activates. Moreover, neutral red was oxidized by Actinobacillus succiongenes to synthesize succinate by fermenting hydrogen and fumarate using carbon dioxide. In different organisms, the electron transfer from the cathode to the cathodic biofilm can facilitate the straight uptake of electrons from the electrode; some of those microorganisms are Sporomusa sp., Clostridium sp. and Moorella thermoacetica to reduce CO2 to acetate. By reacting with the cathode, these acetogenic microorganisms can thrive in autotropic conditions [34]. The acetyl-CoA pathway is turned on since the metabolic pathway needs a terminal electron acceptor other than O2 like carbon dioxide. Other bacteria like sulfur-reducing bacteria and other types of the archaebacteria, which are non-acetogenic bacteria, utilize the acetyl-CoA pathway for neutralizing CO2. The transfer of electrons was evident in the Archaea domain as well. To synthesize methane, the methanogens from the Archaea domain require CO2 and H2. Electro-methanogenesis is done by transporting the electron straight from the cathode to the Archaea biofilm, primarily in Methanobacterium palustre. It was found that methanogen synthesis in the absence of supplied hydrogen was possible when thermophilic bacteria were used, and in the case, of Methanococcus maripaludis. It was shown that the electrons from the cathode were utilized in the absence of hydrogenase [35].
1.3. Anodic Fermentation
Electro-fermentation technology ferments energy-rich substrates like alcohol and carbohydrates by utilizing polarized electrodes as additional electron sources or sink. In the process of anodic electro-fermentation (AEF), the working electrode functions as an anode and dissipates surplus electrons. This phenomenon takes place when the final products are more oxidized than the substrates. To summarize the above statement, when the polarized anodic electrode is the working electrode and is utilized as a driving tool in electro-fermentation, the process is known as anodic electro-fermentation (AEF) and is represented in (Figure 4). Commonly used microorganisms for this process are engineered Shewanella oneidensis, Clostridium cellobioparum, Geobacter sulfurreducens, engineered Escherichia coli [36]. Nevertheless, further research is needed to throw light on the evidence of the effectiveness and feasibility of this process [37].
2. Microbe–Electrode Interaction
The electrodes act as an extra electron source and a sink, which, results in the complete control of the metabolism of the microorganism [38]. Based on the level of reduction of the desired product or the metabolite couple, the microbes can utilize the anode and cathode as electron mediators to their advantage in EF, as shown in (Figure 5). The fermentative pathway could either profit from electron supply or removal, and this knowledge is important as it plays a major role in driving an optimized product-driven EF [16]. A study of a rudimentary working model of an EF for glucose and glycerol predicted the yields of various metabolites. The chain elongation can be done efficiently in organic metabolites if there is an abundant number of redox cofactors such as NADH and NADPH, stimulated by the cathode, which can drive certain reduction reactions [30]. On the other hand, the side of the anode that acts as an electron sink can facilitate a higher biomass yield and assisted power reduction utilization compared to normal anaerobic catabolism. It leads to additional ATP synthesis and the potential increase in the biomass output [39].
The electron transfer mechanism using exoelectrogenic bacteria to the anodic chamber has been studied and examined thoroughly in the past years (Figure 1). However, in recent times the phenomenon of endoelectrogenis is and how the electron is flowing from the cathode can play a significant role in the microorganism’s metabolic activities [40]. Direct electron transfer uses cytochrome dehydrogenase partnerships, using periplasmic hydrogenase-mediated electron transfer and the direct electron transfer because of c-type cytochrome, as shown for exoelectrogen bacteria such as Geobacter and Shewanella are a few of the different electron-transfer mechanisms (Figure 5) [41]. These mechanisms have shown that the electron transfer can conserve energy by either coupled or uncoupled reactions. Electron carriers like cytochromes, ferredoxin, quinones, flavins, i.e., main components in the electrode–microbe interactions, were examined to see their potential role in catalysis by key components [40]. The electrons attained from electrodes aid in the catalytic generation of NADH or ATP, which are energy carriers [42].
In electro fermentation, there is an applied potential on the electrode surface, which facilitates the interaction within the biocatalysts and electrodes [43]. By doubling the electrocatalytic rate and controlling the bacterial metabolic routes while performing EF processes play a significant role in delivering higher product generation [44]. The controlled transfer of hydrogen ion and electrons between the electrode interface and the microbe is directly influenced by the potential that is given to the electrode [45]. The existence of a material that supports conduction in a microbiome together with external stimulus affects the rate of extracellular electron transfer (EET), which provides process specificity and flexibility throughout the bioelectrochemical process [46]. Fabricating electrodes to allow oxygen reduction reaction (ORR) kinetics at the cathodic end with higher efficiency contributes to a higher reduction equivalent than product generation. Graphite flakes, carbon cloth, carbon mesh, and graphite are the carbon-based electrodes currently being used because of their economic viability [47]. In comparison with native electrodes, these carbon-based electrodes can have higher sensitivity, selectivity, and economic value, if the electrodes are subjected to electrode surface modifications [48]. The two dimensional and the three-dimensional structures of the electrode are known to allow macroporous structures with a larger surface area for the bacteria to adhere to and start the electrostatic interactions that are not the same as older electrodes. Better conductivity, larger surface area, and other favorable surface properties that directly contribute to better attachment of bacteria to the electrode and a higher electron transfer rate are present in the 3D electrodes [49]. The 3D electrodes also allow the EET between the microorganisms and the interface of the electrode. In comparison with the regular 2D electrodes, it is much better to modify and coat a macroporous electrode with regards to the EF domain [50].
2.1. Electron Transfer Mechanism
Microbial electrolysis cells generate electricity using microorganism’s metabolic activities, and these microbes are capable of moving electrons across the membrane. In the anodic chamber, the microorganisms oxidize the substrate, which leads to the release of carbon dioxide, protons, and electrons. This process of transferring electrons in and out of the cell is known as extracellular electron transfer (EET). The movement of electrons happens in three steps, i.e., the oxidation of the substrate, the transfer of electrons to the corresponding proteins, and the final step of the extracellular electron transport [51]. The flow of electrons is from an electron donor with relatively lower redox potential towards an electron acceptor, which has a relatively high redox potential. The main component for the transfer of electrons is the multiheme c-type cytochrome present in the Gram-negative bacteria. Other than the cytochrome, the filamentous conductive pill found in bacteria also facilitates the movement of electrons. Certain other essential proteins that facilitate the transfer of electrons include cytochrome, ferredoxin, rubredoxin, hydrogenase, and formate dehydrogenase. Diverse bioelectrochemical reactions can be catalyzed simultaneously during the electron transfer cycle. One such reaction occurs on transfer for electrons from the cathode to the corresponding microorganisms. The electrons are transferred to proteins present in the outer membrane at the initial stages by diffusible intracellular electron carriers such as NAD+/ NADH [52]. Electrons can be transferred to the cathode and vice versa using specific pathways; three such pathways have been identified, i.e., indirect transfer, direct transfer, and transfer through biological projections. The direct transfer of electrons is considered one of the ideal pathways for electron transport. It occurs from the cathode to the corresponding microbes and has been extensively observed in biocathodes for microbial communities such as betaproteobacteria [53]. This mechanism is facilitated without the assistance of any mediator. Although the accuracy of this mechanism has not yet been confirmed, redox enzymes are thought to be involved in electrochemical reduction reactions. Proteins such as ferrodoxin, and other redox proteins, are also considered crucial candidates for the extracellular transfer of electrons. The two most studied microbes for the process of extracellular electron transfer include Geobacter and Shewanella species. These Gram-negative bacteria are metal-reducing in nature. The electron transfer process helps aid the metal reduction reaction by assigning the metal as an electron acceptor to carry out the reduction reaction. However, when the metal oxides or hydroxides used as electron acceptors are poorly soluble in water, an alternative protein such as MHC cytochrome is used for this phenomenon [54]. Geobacter and Shewanella species that have been extensively studied as iron-reducing bacteria [55] have been observed to reduce highly toxic hexavalent chromium using the cathodic terminal. This study indicated that electron transfer could be possible in both directions (i.e., microbe to anode and cathode to microbe) in the current- generating bacteria. The transfer of electrons mediated by soluble electron mediators is referred to as the indirect transfer of electrons. This pathway is highly capable of increasing electron transfer efficiency. For example, Shewanella oneidensis MR-1 [56]. In earlier studies, it was shown that EET could occur either via direct contact or via mediators (indirect pathway [57]. Although, in the latest study, it has been proposed that these two mechanisms cannot generate high current density due to certain limitations, such as the lack of proper surface area and diffusion in the mediators. The microbes instead use conduction based EET to facilitate transport while producing high current density. This conduction-based technology utilized biological projections such as pili. Few other pathways that were suggested under this concept include redox conduction, also known as electron hopping, and conduction through a biofilm matrix [58]. If the transfer of electrons takes place through nanowires or pili, the mechanism is referred to as the transfer via biological projections. This strategy is considered unique since different projections are used for the transfer of electrons. Microorganisms generate electrons, which then are transported by these biological projections to the electron carrier protein. Nanowires assist in generating high current density to support this mechanism. Often, the pili used in this mechanism contributes to the accumulation of cells on the biofilm, these pili display metal-like behavior that increases the temperature and decreases the conductivity, which can be proven harmful for the (MES). As mentioned above, the MHCs are crucial components that assist in transferring of electrons [57]. The MHC’s form covalent bonds with each other and hold them together to form the polypeptide chain. The distance between the two MHC is approx. 1–2 nm. This unique structure of MHC facilitates the EET. Based on these three mechanisms, microbes can support the extracellular transfer of electrons. Nevertheless, significant research is still underway to explore these conductive EET pathways.
2.1.1. Anode Electron Transfer
Ease of construction, high conduction rate, non-corrosiveness, large surface area, high porosity, the fouling resistance, scalability, and inexpensive construction costs are some important features that contribute to building a more efficient anodic chamber. These properties play an important role in mechanisms such as adhesion and delivery of electron given out by exo-electrogenic bacteria [59]. One of the most used materials to produce the anodic chamber is carbon because it is compatible with electrochemically active biofilm and an adaptable morphology. Further, it has a considerably low potential, is cheaper, and is chemically stable under anaerobic conditions [60]. Alternatively, carbon-based material such as graphite granules and graphite fiber brush are some possible candidates to produce of anode because of the presence of a high surface area [61]. Some other materials used for the production of the anodic assembly include carbon cloth coated with Pt/C catalyst [62], vitreous carbon [63], and carbon felt [64]. The materials utilized to produce of anodes in an MFC could be used in a MEC to produce of hydrogen and methane [65]. In a recent study, graphite fiber fabric, which is modified with a multiwall carbon nanotube with nickel support, was observed to be a potential alternative for producing the anodic chamber. It leads to an increase of 52% in methane production [66]. Other than the carbon-based material, transition metals like iron (Fe) [67] and alloys such as Ti/Ru and Ti/RuO2 [59] can be used as alternative materials to decrease operating costs during the production of methane.
2.1.2. Cathode Electron Transfer
Reactions taking place at the anodic chamber are used for the generation of current, while the reactions taking place at the cathodic chamber consume the current produced for generating valuable biochemical products. Cathodic modifications by adding charged species such as chitosan by increasing the surface area with the help of a three-dimensional scaffold electrode support increased current consumption. The EET taking place from the microbe to the cathodic chamber can catalyze diverse bioelectrochemical reactions. Methanogenesis at the anodic terminal occurs via a sequential pathway, starting with electron to proton transfer, followed by hydrogen formation and then reducing carbon dioxide; in this process, the cathode acts as a sink for the hydrogen produced [68]. A material to catalyze the reduction reaction to support the non-spontaneous hydrogen reaction is required at the cathodic end. Carbon stick and graphite felt are some of the most commonly opted materials for the production of the cathode as they are highly productive in terms of methane production and have been observed to contribute to a high carbon capture efficiency as well [69]. An alternative solution to enhance methanogenesis is to increase the porosity in carbon materials using carbon nanostructures such as (MWCNT), which are doped with catalytic material like cobalt tetramethyl phenyl porphyrin, manganese oxide, etc. The electrochemical pathways utilized for methane production by reducing carbon dioxide through the direct electron transfer method can be carried out at both electrode terminals.
Valuable metals such as platinum can be used to catalyze the reaction and to hence reduce the overpotential. However, this process is very expensive and has detrimental effects on the environment [70]. Thus, alternatives such as stain steel are a potential and can be utilized for fabricating anaerobic reactors coupled with single-chambered MEC to enhance the process of methanogenesis. Further alloy-based materials such as Ti/Ru and iron-graphite can boost the process of methanogenesis during sewage sludge digestion [68]. Carbon-based materials such as graphite [71] and reticular vitreous carbon [72], which is commonly used to produce the anode, can also be used for the production of the cathode. In a recent study, it was observed that when specific precious metals were compared to non-precious carbon-based materials, using plain graphite produced higher columbic efficiency (CE) of (100%), these were relatively comparable to platinum-based cathodes that produced an average (CE) of about 96% [73]. Therefore, plain graphite is considered a potentially efficient alternative for platinum-based cathodes. Various efforts, like increasing the interfacial area and interfacial interactions, are being made to enhance the process of electron transport from microbes to the cathodic terminal. Attaching nanoparticles on the cathode was tried using nano-nickel [74], carbon nanotubes [75], conjugated oligoelectrolytes (COEs) [76], and carbon nanotubes on reticulated vitreous carbon (NanoWeb-RVC) [77]. It has also been observed that graphene-modified biocathodes supported and enhanced the bioelectrochemical hydrogen production in an MES system [75]. Positively charged surface modifications were made in another attempt. Modifying the cathode with positively charged functional groups increased the electron transfer between the cathode and microbes [78]. Although, a bacteria Sporamusa ovata, which is negatively charged, preferred cathode attachment and enhanced the electron acceptance from the cathode to reduce it to carbon dioxide and acetate [78]. These negatively charged modifications are observed were due to the cell walls of the bacteria attached to it. An enriched biofilm was observed at the positively charged end of the anode. In contrast, a repulsive interaction was observed at the cathodic end due to the negatively charged modifications made to it. Hence attaching microbes on a cathode have a charge in the barrier due to which it has been observed that both the zeta-potential and the hydrophobicity of cells were increased in a current consuming biofilm [79]. According to the changes in the cell surface and its features in the cathode, different modifications can be tried in contrast to that of the anode. Regarding the mechanism, the two most extensively studied microorganisms for extracellular electron transfer are Geobacter and Shewanella species. Geobacter and Shewanella are both metal-reducing and gram-negative bacteria. Extracellular electron transfer in microorganisms is utilized in the metal reduction process by the microorganism and, the metal is used as an electron acceptor. When metal (hydr)oxides that are insoluble in water are present as electron acceptors, extracellular electron transfer occurs through the multiheme c-type cytochromes in Geobacter and Shewanella [80]. Due to this phenomenon, the microorganisms can extracellularly transfer electrons which can be applied for BES. The mode of extracellular electron transfer is broadly classified into the following: (1) direct electron transfer: nanowire [81] or (2) direct contact via cytochrome [82]; and (3) mediators-shuttled: endogenous, exogenous as a redox compound or a by-product [83]. The extracellular electron transfer mechanism of Shewanella spp is depicted in (Figure 6). Multiheme c-type cytochrome is a main component of the electron transfer channel in gram-negative bacteria [84]. Filamentous conductive pili or nanowires also actively participate in electron transfer in the in Shewanella [43] and Geobacter [81]. BES utilizes two directions, from the microbes to the electrode (anode) in MFC and from the electrodes (cathode) to the microbe in MES, with the same or different mode [85]. Electrons flow from an electron donor with a comparatively lower redox potential to an electron acceptor with higher redox potential.
2.1.3. Applied Voltage as a Power Source
The applied voltage is one of the key parameters that have been proven to contribute as well as highly influence the performance of BESs for the process of methanogenesis. However, it is very important to keep in mind the specific range of applied electrical potential differences that could be detrimental to microbes [86]. In a recent study, [87] the impact of the applied voltage on hydrogenotrophic methanogens was evaluated. The study showed that on applying a voltage of 0.3 V, the proliferation of hydrogenotrophic methanogens could be observed, which had a positive impact on the methane generated by anaerobic digestion (AD) of activated sludge. Compared to the conventional anaerobic digestion technique, an increase of about 22.4% was observed in the process of methanogenesis on applying a voltage of 0.3 V using electrodes fabricated by iron-graphite materials [88]. However, it was also observed that a slight shift of the applied voltage to 0.6 V could result in the inhibition of these microbes due to alkaline like pH conditions in the system. This sudden increase in the pH caused a significant decrease in the production of methane by 43%. In contrast to the above study, in another research [89], it was observed that when using a two-chamber cell with a short HRT for about 3 days, with an applied voltage of 0.6 V, a significant increase was observed in methane production. Nevertheless, voltage conditions of 1.0 V and 2.0 V decreased the production of methane by 37%. This was mainly caused due to the destruction of cell membranes, which then harmed the growth and metabolism of methanogens. Another reactor that utilized electrodes fabricated using Ti/RuO2 and used waterweed as its substrate showed that a greater number of total solids were discarded, and methane production on applying a high applied voltage corresponded to about 56.3% and 0.24 m3/m3/d d1 at 1.0 V, respectively [90]. The contradictory results of different studies mentioned above suggest the importance of applied voltage in an anaerobic BES and the different factors that influence it, such as substrate type, electrode material, and cell configuration. The dependence of the applied voltage on these factors demonstrates the importance of carrying out voltage optimizations for every BES to enhance and optimize the energy efficiency of the system.
3. Electro-Fermenter Design
Fermenter design plays a considerable role in the fermentation industry. Electro-fermenter design determines the operational cost, fabrication cost, the type of electrode materials, redox mediator, membranes separation membranes that are to be used as well as any new add on to an existing fermenter to aid its electrochemical needs all which adds up to the total cost of fabricating an electro-fermenter [91]. As shown in (Figure 7), there is a huge array of configurations of fermenters in the industry that employ anaerobic fermenters to produce various bioproducts on the one hand and the other; reactors that use electric current to facilitate the process of fermentation-electrochemical reactors [92]. These electrochemical reactors should be incorporated into conventional fermenters to meet their electrochemical needs and optimize their biological, hydraulic, mass transport requirements. Currently, various bioreactors are being utilized for bioprocessing and any novel solution in the research domain. Over the last 10 years, electrochemical cells have been reviewed and studied thoroughly to use microorganisms as catalysts. Various solutions could be used in various electrode materials, shapes, membrane properties, and cell dimensions are examined and utilized [10].
Economic feasibility, performance, and stability are the main factors that govern the design of any instrument for industrial or large-scale applications. In electro-fermentation applications, carbon and graphene are used extensively because of their high conductivity, low cost as and chemical stability. Manipulating an electrode’s properties increases the electronic conductivity and transfer of electrons, which is more desirable in an electrode [93]. The microorganism’s ability to form a microbial biofilm and whether it can adhere to the electrode or not and is determined by the porosity and the surface area, and the biocompatibility of the materials. The electrochemical reactions at the surface can be increased if the surface functional groups like oxygen, nitrogen are present [1]. In the event of a culture electrode being set, the medium will remain in contact with the biofilm for a significant time, until the designs reach the optimum conditions; there is no leakage or flow through the structure, even though the medium is disrupted to increase the surface area relative to the volume of the reactor. On the other hand, a mobile electrode, small solid conductors are suspended in the medium, which also acts as a matrix for the microorganisms to adhere to the substrate. This method is utilized in fluidized bed reactors wherein the electron outflow towards an external circuit takes place by a stochastic contact with the electrode and the conductive particles. The facilitated EET should be the preferred way of transferring electrons as well as a mediator must be chosen following the microorganisms; the main objective of an EF is to trigger suspended microbial populations [94].
Usually, the counter electrodes functioning, and their respective reactions could occur either in the reactor or in different chambers separated by one or more IEM [91]. As shown in Figure 8, the single-chamber setup is perfect only if the counterreaction products do not affect the purity of the working reaction products, conventional fermenters, along with simple incorporation of electrodes, can act as an EF system which is much more economically feasible than constructing a whole new electro-fermenter. The anodic and cathodic products interact with each other and affect the overall output of the EF system in terms of purity of the product, the EF process must take place in specially dedicated reactors which are separated by membranes or proper separators which is more expensive to construct in comparison with the modified fermenter system [92]. Suppose the extraction of the product and concentration are primary concerns of the EF then in that case, multiple separators or membranes are meant to be included in the design to facilitate the electro-osmotic drag of the cations and anions through selective membranes [95]. To convert specific complex substrates into products that are unique and pure is very uncommon. In the EF process, both the production and the extraction can be correlated to each other. In complex broths, the product of interest should be contained relatively higher than other compounds [10]. The electric field prevalent in the culture medium prevents the product’s inhibition in the microbiome population. The entire extraction process happens simultaneously along with the fermentation process wherein the extraction takes place in the reactor. For fermentation processes that look for microbial-biomass synthesis like the industrial production of probiotics and starters, the EF proves to be a valuable technique wherein the traditional methods consist of major setbacks like cell count as well as their activity inhibited by-product collection and various pH and redox controlling agents most of them being chemicals which results in increasing the osmotic pressure on the cell membrane and the overall salinity which further complicates the entire process.
4. Mixed Microbial Cultures Used in Electro Fermentation Processes in Industrial Biomass Refineries
The food and beverage industries that use the conventional methods of fermentation all employ enriched mixed cultures. Compared to pure-culture fermentation processes utilizing cocultures in the conventional fermentation, which show additional benefits based on the main goal of the fermentation, simplifying the process or its economics as less costly protocols are needed to prevent contamination as well as maintain an axenic environment [43]. By removing the inhibitors, when one microbe facilitates the growth of the second microbe by synthesizing growth factors, this process is called syntropy; the process ensures the functioning of optimal environmental conditions. A few cascade pathways need a minimum of two unique organisms to complete product formation in most cases [96]. The production of both primary and secondary metabolites in cocultivation conditions leads to new products and waste production. For instance, the inoculation of Aspergillus awamori as the second strain of the coculture along with Saccharomyces cerevisiae leads to an increase in ethanol synthesis from starch [44]. Complex substrates like mixed organic materials (lignocellulose or waste effluents) can be utilized for the synthesis of valuable chemicals or fuel, if the consortia of the two microbes can utilize mixed/impure/unrefined substrates like biomass waste only then can they be exploited [6]. Turning to the technique of EF can help in making the anabolic and catabolic processes of the mixed cultures better and manage various limitations as well. For example, the complexity in controlling the product formation stability because of the substrate impurities can be regulated by the EF processes. The technique of EF could be essential to refine the industrial processes [97]. Multiple studies have succeeded in using an electrochemical approach to influence the microbial consortia.
To regulate complex microbiome and link their structure, composition, and increase functioning in the reactor and stability and product formation, the electrochemical systems are considered better compared to than conventional fermentation strategies [98].
In a recent study, the electrode was introduced in an anaerobic digester, where the process was successfully achieved in enriching the Geobacter species in the suspended sludge and anodic biofilm. A raise in the production of methane was observed in the process. However, of all the methane synthesized, 50% of it was contributed from unknown metabolic pathways. It was said that a direct interspecies electron transfer within the Geobacter and Mehtanosaeta species was established [99]. Other parallel studies have shown possible ways to control metabolic pathways through the EET to and from the electrodes to achieve more precise microbiome fermentations and optimize the conversion of waste-derived substrates into products of interest. A GM strain of Shewanella oneidensis which focused on the conversion of glycerol to ethanol and CO2 to correct specific redox reactions that are imbalanced were carried out. This approach was observed to support any reaction as long as the substrate is more reduced than the compound of interest [100].
The electro-assisted dark fermentation, a patented technique, was examined in a single bioreactor containing a microbiome to synthesize hydrogen using different organic substrates. An identical procedure was then used to produce other chemicals such as acetone, butanol, ethanol, acetic acid, and butyric acid [101].
In different studies, several EF techniques have been used to boost the yield of glycerol conversions, wherein hydrogen synthesis from pure or glycerol by-products of biodiesel fuel production by electrochemical approach was substantially increased [6]. Another study in glycerol fermentation showed that the metabolite synthesis could be altered if an electric current is supplied. For example, the alcohol and medium-chain fatty acids (MCFAs) were generated but the production of the 1,3-propanediol (1,3-PDO), which included polymer production, solvent, and antifreeze production, was not changed. By imposing a cathodic potential of (20.9 V) or a cathodic current from 1 to 10A/m2, the conversion of glycerol can to 1–3 PDO could be stimulated. Glycerol biocathode fermentation could be achieved by polarizing the biocathode at −1.1V, which also resulted in the highest concentrations of 1,3-PDO in a glycerol EF study [1].
5. Industrial Fermentations of Pure Cultures Using an Electro Fermentation Approach
In addition to food processing and transformation utilizing traditional microbiomes, metabolic pathways engineering has also been used for industrial applications to synthesize metabolites in bioreactors, by using pure microbial cultures with specific strains [102]. Fermentations of pure cultures aim at maximum product formation. The selection of product and purity of culture can be achieved using a single, unique, and pure substrate. As mentioned earlier, the most important fraction of cost is represented by the production of unadulterated and sterile carbon sources such as glycerol, corn-starch, sugarcane, vanillin, etc. [103]. Sustainable production of substrate requires dedicated use of land and agro-industrial based transformations, specifically for chemicals produced in bulk quantities. Corn-based dextrose used to produce chemicals such as lactic acid and succinic acid accounts for about 50% of greenhouse gas emissions and cost in terms of the invested energy [104]. Despite the presence of air filtration and sterilization systems, when large quantities of air are sparged, the stability of pure cultures and sterility of reactors is of major concern [105]. Ultimately, sparging of gas increases the foam formation in broths; hence a continuous addition of antifoaming agents along with a foam monitoring device is required to keep this issue under control. All the electron equivalents are extracted in a single product to achieve an ideal and balanced fermentation, thus promoting isolation and purification of the downstream product [6]. Diverse metabolites are produced even while utilizing single microbial specie cultures because there are several pathways present where the cell usually can achieve homeostasis, hence producing diverse end products [14]. Integrating fermentation along with biochemistry can be profitable to (1) enhance the yield and productivity; (2) improve process kinetics [106]; (3) alter of decreasing the fermentation product spectrum [107]; (4) to provide O2, H2, and electrons in-situ, with an enhanced distribution [55]; (5) improve electron flux process and carry out thermodynamically unfavorable reactions [30]. About 10 years later, Kim and Kim [108] showed an incline towards more reduced products (like butanol) during glucose fermentation with C. acetobutylicum. By utilizing an electrode occupied using a modified strain of S. oneidensis, the stoichiometric conversion of glycerol to ethanol could be attained [99]. In a more recent experiment [109], a modified strain of Pseudomonas putida KT2440, due to its ability to generate its mediators such as phenazines, was used in fermentation, and through this, it could potentially produce biochemicals which would otherwise require oxygen sparging. Two scientists, Kracke and KrÖmer [38] established core connections of anaerobic carbon pathway in E. coli. It was observed that through this, there was a boost in anaerobic glycerol and sugar fermentation with advances in the product yield. In another experiment similar to this, Pandit and Mahadevan [110] exhibited a sequential and complex strategy to enhance productivity, optimizing the growth rate without enhancing the electrical aspect. A continuous and static potential of about 15 V was applied to S. cerevisiae culture, which induces a significant rise in both ethanol yield and productivity (two up to three times faster), without any resultant current [111]. A recent observation exhibited similar results using pure-culture fermentation. Species like Clostridia sp. and Brevibacterium sp. were used to produce butanol and glutamate respectively [91].
6. Microbial Electrosynthesis for Value-Added Chemicals-Reactors Design and Recent Development
6.1. Integrating MEC-Microbial Fuel Cell (MFC)
MFC’s are systems used to generate electricity by degrading waste effluents [112]. These systems have been proposed as renewable power sources to operate MEC. Basically, in a coupled system, a single-chambered MFC is integrated with a dual-chambered MEC and hence used to produce hydrogen without the support of any external power supply. Apart from hydrogen production, these systems can be used for power generation as well as to produce formic acid using carbon dioxide as the chief substrate.
6.2. Submersible Microbial Electrolysis Cell
This type of system is designed to eliminate the extrinsic power supply needed for the functioning of MECs. Such cells are typically designed to be fit into an existing anaerobic digester, where two joint chambers are added to function as two cathode chambers. There is no anodic chamber present, and one of the two cathodic chambers is only used to produce electricity, while the other half is used for hydrogen production [113].
6.3. Integration of Pyrolysis-MEC
Pyrolysis is a process through which organic matter is thermodynamically decomposed at high temperature in the absence of oxygen. This process is used for generating bio-hydrocarbons. The main drawback of this process is handled by supplying the final product of fuel with a hydrogen source in the final step. Although due to the present unavailability of a commercial and renewable form of hydrogen, fossil fuels are being used for this process, which creates a significant impact on the sustainability of the environment and hence for this process, hydrogen can be alternatively derived from a liquid or aqueous form of waste streams via the process of pyrolysis carried out in a MEC thus meeting both the needs [114]. The blend generated as a final product through this process is called a bio-oil.
6.4. Integrating MEC and Process of Lignocellulosic Ethanol Biorefinery
The production of lignocellulosic ethanol is considered fundamental second-generation biofuels. Lignocellulose is derived from material like wheat straw by hydrothermal processing, followed by enzymatic hydrolysis. This process is used to produce ethanol and other by-products such as VFA’s, phenolics, xylose, and other polysaccharides. The entire concept of biorefineries could be highly beneficial as it helps in the purification of polyphenols and target modification of fruit-based phenolics. The integration of the procedure used in an ethanol biorefinery could lead to the maximization of energy output, hence supplied and simultaneously support valorization of the ethanol waste stream [114].
7. Bio-Electro Products
7.1. Short-Chain Fatty Acids (SCFAs)
When performing AF, microbial cultures using the process of acidogenesis convert organic and inorganic material or inorganic carbon dioxide to an intermediate form of fatty acid [115,116]. Carbon dioxide and waste effluents can be utilized for producing short-chain fatty acids (SCFAs) or volatile fatty acids (VFAs; C2 to C5) with the help of an electro-fermentation [117]. The electrodes have the capability of controlling the rate of kinetics during EF. They have an added advantage of electrochemically influencing acidification of monosaccharides by employing anaerobic bacteria, hence increasing the productivity of VFAs [118]. Electrofermentation through the process of electro-metabolism can be utilized as a viable route for allowing electron movement to attain diverse fatty acids. Further, the produced SCFAs can be utilized in various other bioprocesses as an electron donor. SCFAs, when used as an electron donor, can be used for producing bioplastics using the process of anoxygenesis, to produce diesel using photosynthetic machinery, for the generation of bioelectricity, etc. [11]. Production of VFAs has considerable applications as an important chemical in the textile industry, food, beverage, polymer, and plastic industries [118]. Acetogenic and chain elongating microorganisms are utilized in metabolic pathways for converting organic and inorganic substrates or carbon dioxide to produce VFAs [119]. One possible channel for producing n-butyrate could be by reducing carbon dioxide to acetate using hydrogen as the electron donor via the acetyl-CoA pathway. In BES, this channel was primarily observed to have been driven by electro-derived electrons along with a high acetate production utilizing carbon dioxide [18]. A few acetogens utilize the process of aldol condensation of two acetyl CoA molecules to produce n-butyrate as the final product (Equation (1)). An alternative pathway to produce n-butyrate is through the elongation of acetate utilizing the reverse beta-oxidation process. Ethanol or lactic acid is used as an electron donor for this process (Equation (1)) [120].
7.2. Medium-Chain Fatty Acids (MCFA)
Medium-chain fatty acids (MCFAs) are naturally hydrophobic and have a low ratio of oxygen/carbon. Hydrophobic character facilitates higher energy density, low solubility, and easy separation from the fermentation broth [120]. Production of MCFAs through the biological route uses the reverse β-oxidation pathway. This process needs a carbon source, reducing equivalents and energy from different electrons donors such as ethanol, hydrogen, and lactic acid [121]. In BES, the regenerative source of acetate is utilized to produce MCFAs at the cathodic chamber. Acetate is considered a major intermediate to produce MCFAs as it has a comparatively higher commercial value and energy content compared to the generation of methane using acetate [122]. In BESs, EF with an applied or in-situ potential can use acetate, propionate, and butyrate present majorly in waste effluents for chain elongation to produce MCFAs. Various experiments have been carried out for producing n-caprylate using ethanol as an electron donor [123]. Clostridium kluyveri observed the formation of caproate (C6) and caprylate (C8) from acetate in a BES at −0.9V against a normal hydrogen electrode (NHE) on the cathode with hydrogen provided in-situ as an electron donor [121]. Compared to ethanol, the long hydrocarbon tail of the MCFA will provide more energy density and comparatively lower solubility. The heating value of 1 mole of caproic acid (3452 kJ) is greater than 2 moles of ethanol (2638 kJ) needed to produce it, and this indicates the importance of electrodes for commercially viable product recovery in the process of chain elongation [124]. When addressing waste management related issues, MCFA has diverse applications in aviation fuels, producing renewable diesel precursors, antimicrobial, flavor, and fragrance related intermediates [122].
7.3. Bio Alcohols
The intermediates formed in AF are called bio solvents [125]. BES’s final product formation depends on the decoupling of oxidation and reduction reaction [92]. In addition to hydrogen production, the electrons produced at the anodic chamber can also be utilized to reduce VFA intermediates to form bio-alcohols [17]. The rate of the kinetics of these reactions relies on the biocathode. The biocathode, in turn, has electro-metabolic control over the entire mechanism [115]. Experiments using the cathode as the working electrode with a poised potential of 0.6 V, against a standard hydrogen electrode were carried out to produce 1,3-propanediol (1,3-PDO) from glycerol utilizing electroactive Geobacter sulfurreducens. This method increased productivity by about 32% in contrast to AF [95]. When a cathode potential of −0.55 V has utilized the production of ethanol and hydrogen and n-butyrate is observed by biologically reducing acetate [115]. For this process bacterium, Clostridium ijungdahlii is added to a mixed microbiome [95]. Another genetically modified organism, Ralstonia eutropha H16, can also be utilized to produce long-chain alcohols such as iso-butanol and 3-methyl-1-butanol [96]. Bio-alcohol production with some specific bacterial strains helps enhance productivity with the EF process (Equations (2) and (3)).
7.4. Biohydrogen
Carbohydrates such as glucose are preferred as electron donors for promoting the growth of microbes used for biohydrogen production [126]. On the other hand, protons could be used as electron acceptors utilized to reduce hydrogen and hence balance the intracellular reducing power. To maintain electrical neutrality, electrons generated during biohydrogen production need to be discarded [117]. Microbial electrolysis cell (MEC) is an efficient source that could be used to produce biohydrogen [127]. Generated due to the in-situ electrode potential, exoelectrogenic bacteria present in MEC oxidize organic substrates to produce electrons and protons. The bacteria then transfer the produced electrons and protons to the anodic chamber and the solution, respectively [43]. The protons are further transferred to the cathode, where they are reduced for biohydrogen production [43]. Exoelectrogenic bacteria oxidizes acetic acid at anode simultaneously; hydrogen gas is produced from H+ molecules reducing electrolytic water at the cathodic end [128]. Consecutively these reactions can be carried out in a single as well as a double-chambered arrangement. In the latter case, both the electrodes have spatial separation with an ion-exchange membrane. The energy applied externally is mainly to transform waste into hydrogen in MEC (Equations (4)–(6)). This energy is generally lower than the energy needed for electrolysis of water (1.8–2.0 V). When present in the microenvironment, solid electrodes enhance the fermentation process for hydrogen production [129]. In single-chambered arrangements during glycerol fermentation, polarized electrodes exhibited an increase in hydrogen production [130]. Neutral red can also be used as an electron mediator to produce hydrogen [40].
7.5. Biomethane
An important requirement is upgrading biogas technology with electrode inventions to improve the content of biomethane present in the total biogas. Combining AF with MEC for upgrading biogas technology via electro methanogenesis is an upcoming area for research [131]. Electrons and protons are released utilizing the exoelectrogenic bacteria towards the anode and the solution, respectively. The electroactive microorganisms or electrotrophs at the cathodic terminal accept these transferred electrons with higher efficiency via the external circuit, which occurs when a low voltage is poised [132]. The use of hydrogen produced from the cathodic terminal can alternatively be utilized as an electron carrier to support high methane formation (Equations (7) and (8)) [133]. Methane can be produced via two mechanisms near the cathodic terminal during EF, either by using hydrogenotrophic methanogenesis where hydrogen is used as an electrochemical mediator or by using electrons directly as a source of power reduction [134].
The poising capacity of −0.8V on biocathode (vs. standard hydrogen electrode (SHE)) is an important external stimulation technique to push the reduction of the biogas fraction of organic substrates or carbon dioxide into methane [134]. Additionally, on supplying a potential, the endothermic barrier could be controlled on hydrogen formation towards methane production [135]. Hydrogenotrophic methanogens, such as species of methanobacteria are the fundamental players in supporting carbon dioxide fraction towards the efficient formation of methane [136]. With the rise of electrochemical interventions in traditional fermentation processed, the exoelectrogenic bacteria formed can utilize the end products of fermentation, such as acetate, to then release electrons [137]. High methane content in the total biogas during the EF process should be attributed to electrochemically drive carbon dioxide reduction at the cathodic terminal to form methane by employing hydrogenotrophic methanogens [138]. Hydrogen is instantly converted by hydrogenotrophic methanogens to produce methane [66]. Although, the key source of electrons for the reduction at the cathodic terminal is the oxidation of substrates by using anodic exoelectrogenic supplementation. This further helps in increasing the electrolysis mechanism towards higher production of the final product [139]. Some complex substrates can be directly oxidized by exoelectrogenic bacteria such as Rhodoferax ferrireducens, Klebsiella pneumonia, and Aeromonas hydrophila for generating power in pure cultures of MEC [30]. Hydrogen, VFAs, and alcohols can be consumed as a feedstock by these bacteria [140]. Anaerobic fermentative bacteria first oxidize glucose to organic acids followed by final consumption products of fermentation by exoelectrogenic bacteria present at the anodic terminal. Hence eliminating the process of feedback inhibition, which takes place during glucose fermentation [141]. The EF strategy to produce methane could be enhanced by understanding the mechanisms involved during the interactions between microbes and the electrode the understanding of the alteration shown by the microbiome towards the production of methane.
7.6. CO2 to Value-Added Products
An unprecedented increase in the levels of carbon dioxide has been observed over the years. This phenomenon can be managed by tackling the environment and by opting for sustainable options [142]. One such domain where scientists have shown immense interest and through which the carbon dioxide levels can be reduced includes microbial electrosynthesis technology (MET). MET application comprises the synthesis of chemicals and the production of biofuels by utilizing microorganisms. It utilizes carbon dioxide as its principal substrate [143]. Combining EF with electrochemistry and microbial fermentation helps improve bio-sequestration capabilities; this is mainly due to microbe-electrode interactions. The microbe-electrode interactions help to derive the production of a specific product while optimizing the thermodynamic energy needed to synthesize the product. The EF operates using the directional movement of charged ions with high precision and viability [144]. However, further research needs to be carried out to boost their practical applications other than the already present commercialized technologies. This would help in maximizing the performance of the same towards the environment and hence provide sustainable options. Operating parameters of MET for the production of various value-added products are given in Table 1.
8. Microbial Electrolysis for Hydrogen-Reactors Design and Recent Development
The first-ever study published on MEC was in the year 2005. Ever since there has been an exponential increase in the number of papers being submitted for the same. MECs use the anodic terminal as an electron sink to oxidize organic substrates to generate protons and electrons by exoelectrogenic bacteria (they transport electrons through extracellular mechanisms) [59]. The protons hence produced travel through the proton exchange membrane (PEM) towards the cathodic terminal, and these protons are then reduced by electrons transferred through the external electrical circuit to produce hydrogen finally [144]. It was observed in a study that the MEC system cannot function spontaneously and that an additional energy source is needed to drive the reaction [148]. Any electrons hence digested by the exoelectrogens can be utilized at the anodic terminal of the MEC. This has caused a considerable variation in organics substrates studied in MECs, right from defined pure chemicals to a mixture of real wastewaters, observed in the experiment [149]. The anode potential can range from 0.2 to 0.5 V with the different carbon sources present. According to the Nernst equation, the cathode potential can be affected by the amount of protons present (pH of electrolyte), temperature, and partial pressure of hydrogen. With a drop in the cathode potential, an increase in the temperature, pH, and partial pressure of hydrogen have been observed [150]. A study observed that the pH is the most important parameter to determine the cathode potential. Due to thermodynamic limitations, energy input is essential for MECs. Theoretically, according to a study, the reaction should be feasible spontaneously with glucose as a substrate. Still, glucose must be completely oxidized to carbon dioxide, which generally does not take place during anaerobic fermentation. An alternative approach to decrease or eliminate energy requirement in MEC is to raise the cathode’s potential by altering the pH or pH2 [150]. This situation appears to be feasible because the cathodic potential could be kept high if the pH of the electrolyte used is lower than 5 although to maintain a low pH would mean to obstruct the migration of proton through the PEM, therefore resulting in a low pH at the anodic chamber which then turns out to be harmful to the microbes. Alternatively, maintaining a low pH2 is extremely impractical while carrying out operations.
9. Setbacks in Lactic Acid Fermentation
Some of the industrial drawbacks of lactic acid fermentation include using products related to the human food chain. Despite the advancements in technology regarding this scientific discovery [151], the generation of lactic acid bacteria exhibited some limitations related to biotechnology industrially. Limitations in the biomass generation of fermenting microbes industrially are mainly due to metabolic inhibition, which is applied due to the decrease in intracellular and extracellular pH [36]. This phenomenon occurs due to the accumulation of organic acids in a non-buffered batch or fed-batch bioreactor [36]. Based on some previously present data [151], it has been established that EF can be successfully applied to lactic acid bacterial species that have been genetically modified with heme and menaquinones to carry out the process of respiration. An important prerequisite for applying electro-fermentation is the presence of a complete or even a partially complete transport system [6]. Electron mediators used to shuttle electrons between the electrodes and the microbes are considered an alternative solution for genetically less equipped bacteria for the process. These bacteria could be less equipped, but they have high relevance in the dairy and probiotic industry. Some examples are Streptococcus sp., Lactobacillus acidophilus, and Bifidobacterium animalis subsp. Other applications in lactic acid fermentation using secondary and waste products could use hyperthermophilic strains. Even in the presence of waste effluents, the purity of the culture can be maintained due to the presence of the extreme temperature conditions (80 to 90 °C) [27]. Thermotoga neapolitana, a Gram-negative bacterium, supports dark fermentation, which is an efficient way to produce lactic acid; this bacterium grows most efficiently over about 80 °C. The hence produced lactic acid, along with the other substrates, can be used as feed for cultures. In the latest experiment, it had been observed that the bacterium T. neapolitana when used with acetic acid and carbon dioxide, enhanced lactic acid production [27]. Capnophilic lactic fermentation is a patented process observed to be a potential advantage where microbial synthesis can be combined with carbon dioxide. In a study [7,9,104], it was shown that the respiratory mechanisms in T. Maritima, opened the possibility of making these strains as electroactive microorganisms in bioelectrochemical applications, such as EF. Therefore, various opportunities have been observed based on the principles of EF: (1) stabilizing/optimizing metabolic reactions carried out while the process of fermentation by controlling the possible disequilibrium due to purity of substrate, redox/pH conditions, by-product accumulation, etc.; (2) establishing the oxidative or reductive conditions to support breakdown or elongation of carbon chain; (3) increase ATP synthesis and thus help in the improvement in the yield of microbial biomass, which means a decrease in production costs by improving the cell count and yield of biomass in the initial and probiotics fermentation; and (4) security and safety-related to food and production of health-care additives, nutraceuticals, and food ingredients. Some other important targets are based on the agricultural and environmental impacts, the reduction of industry, and the development of brand new and highly competitive green chemistry products. This would lead to a reduction in the dependency on petroleum-based products that are non-renewable and meet the climate change targets and lead to a more sustainable and greener environment. The fundamental step to this is to invent new bio-refining technologies, transform waste effluents, and produce bio-based products and biofuels.
10. Current Challenges and Impediments to Scaling-Up the MEC
Though EF techniques have some of the most wanted attributes in industrial fermentations, it is still an uphill battle to set up a pilot or an industrial scale fermentation. The concept of EF is new, and the technique was developed only in the past ten years. It still has not been upscaled yet, making the possibilities of studying large scale reactors and its possible optimizations are non-existent as of now [36]. Only a few pilot studies are reported so far. The biggest challenge to the scale-up is the same as any other bioelectrochemical system, system efficiency. There is a lot of energy wastage and a lower specific production rate conventionally when the size of the reactor is increased. Primarily maintaining the electron transfer efficiency from the exoelectrogens to the anode is a huge task when scaling up the EF reactor. The relation between the current density and anode surface has not been systematically quantified; instead, it is just accepted based on the convention as the current density decreases as the anode size increases. This results in the reduction of energy efficiency as well as an increase in energy input cost for H2 synthesis. Creating an electrochemically active system that shows high activity and maintains the required mass/electron transfer within the biofilm is necessary. The system’s total energy loss plays a massive role in the overall EF efficiency. The main reason for such energy losses is the pH gradient, a common issue in most membrane-based bioelectrochemical systems regardless of the type of membrane used. This could potentially result in the shifting of major potentials at the cathode and causing interferences in the anode exoelectrogens metabolism site. The proton gradient that uses a cation exchange membrane loses 0.38 V for every 1 V spent on the system [36]. The ion selection of the membrane and the ions showed in the electrolyte, and its pH buffering potential can decide the pH gradient between the two compartments. The buffering of an electrolyte by chemical methods is not practical at the industrial scale. Hence, a more practical approach to rectify this issue is necessary [143]. Another type of energy loss is the ohmic energy loss; the name itself speaks about its origin: the ohmic resistance of the anode and the cathode and the conductivity of the electrolytes and the membrane. Due to their low relative conductivities and lesser ohmic resistance, carbon-based electrodes may serve more ideally at very large-scale industries. Additionally, current electrolytes conduction is also an issue, especially in using bioelectrochemical systems to treat wastewater [145].
Along with the EF reactors’ issues, combining the EF reactors with other industrial-scale techniques is challenging [143]. The incorporation with other methods can expand the potentiality of MEFs drastically and uplift their industrial competence. The biggest impediment to this approach is that all the stated above options are still in very early stages and need the required insights, which are scarce. Advances in system structure designs are necessary to meet EF’s requirements and other possible integration processes. This leads to high expenses in system construction, which then needs to be complemented with additional benefits. Additionally, comparability between the MECs and the integrated approach is unclear and needs to be consistently investigated. Because the two techniques are mutually dependent, one process’s performance could significantly influence that. If the system is not well designed, this influence could be detrimental. Dynamic configuration assessment is also important, in particular, because of the comparatively low MEC rate. Different factors required for MEC to scale up reactors and other strategies for achieving a dynamic balance status may be problematic and should be solved during scaling-up [142]. Characteristics of the microbial electrolysis cell and associated operating parameters are given in Table 2.
V is the total reactor volume, σ is the ionic conductivity of the electrolyte, U is the cell voltage, PBS phosphate buffer solution. Current densities and production rates are expressed concerning the cathode projected surface area. The anode Faradaic yield (An.) is the ratio of the electrons circulating in the electrical circuit to the electrons that could be extracted from the amount of substrate oxidized.
Faradaic yield is the ratio of the electrons recovered from the H2 produced to the electrons circulating in the electrical circuit [149]. The γG and γH are the energy and thermal yields defined.
11. Future Perspectives
EF is a forward-looking and inventive field and has a wide area of research. It is believed that more focus is needed on the multidisciplinary aspect of EF research. Much attention is also paid to the cathodic terminal, as it is a vital component in the production of hydrogen and methane. Advancements and modification of the cathode could lead to an efficient performance of the reactor. Modifications of the cathode should mainly focus on producing a porous electrode, making the process of electron transfer and formation of biofilms more efficient. Recent advancements related to the abiotic cathode are mainly focusing on the modification of the surface to support the conduction of active substances that have been loaded through electro-deposition and other chemical reactions. They help to improve the catalyst capacity and the electron transfer rate. Other than the efficiency of the electrode, the capital costs should also be kept in mind. The capital costs might be much higher than that of conventional wastewater treatment systems based on the materials used for the same [150]. Other challenges that need to be solved in the future include the process of scale-up fermentation, inventing different materials to enhance the catalytic activity, and support the adhesion of microorganisms and optimization of energy requirements [145]. A deeper study is also required for transport-related reactions that occur in the reactor, as these could lead to the formation of new cells and modifications in the electrode architecture. This would then solve various issues like limitations related to transport, low CE, etc. The future of this technology seems promising and has a great deal of scope due to its various applications and integrations with other technologies, which would, therefore, increase the growth of the industry and promote sustainable development [151].
12. Conclusions
EF is an evolving technology that still needs to be explored and could give rise to new frontiers in biorefinery with the aid of a broad range of substrates, electroactive biocatalysts, and specific bio-products. EF portrays a symbiotic relationship between two technologies, i.e., electrochemical fuel cell technology and fermentation. There has been an increase in the demand for different chemicals due to their use as precursor molecules in various applications, and, as a result of this demand for chemicals, there is an immediate need for an alternative and sustainable process that can be used for the development of these valuable chemicals. Biological routes are chosen for the generation of chemicals as they are comparatively sustainable and cause less harm to the environment compared to chemically synthesized routes. Applying the biological pathway helps in the significant decrease in production costs as well as a considerable decrease in the waste generated. Nevertheless, these processes do have certain thermodynamic and cellular metabolism related limitations such as low productivity and conversion efficiency. One way to overcome these limitations is by placing electrodes in a fermentation medium to drive the electrochemical reaction, which is the basic working principle of an EF. The electrochemical energy hence generated can externally generate a shift from balanced to unbalanced fermentation, increasing the possibility of attaining a theoretically high yield. However, such conversions need a constant flow of current to maintain the redox conditions and sure good productivity. Similar to MFCs and MECs, some limitations of EF are efficient electrode material and reactor configuration. Inserting electrodes into the medium allows microbial dynamics of the system; this allows new opportunities in observing certain electroactive microbes and their specific interactions. EF can hence give innovative solutions to a range of drawbacks that limit the process of fermentation at an industrial scale.
Author Contributions
Conceptualization, R.P., S.P. and J.M.S.; writing—original draft preparation, D.D.B.; R.P.B. and S.P.; writing—review and amp; editing, R.P.; D.A.J.; S.K. and N.K.; visualization, N.K., S.P. and R.P.; supervision, S.P.; J.M.S., R.P. funding acquisition, S.P., R.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Acknowledgments
The authors wish to thank the financial and facility support from Sharda University, Greater Noida, Delhi NCR, and Mahatma Gandhi Central University, Bihar, India.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| EF | Electro-fermentation |
| PHA | Polyhydroxyalkanoates |
| PHB | Polyhydroxybutyrate |
| BES | Bioelectrochemical systems |
| AF | Anaerobic fermentation |
| ATP | Adenosine triphosphate |
| Moxd | Medox |
| MRed | Med RED |
| MET | Microbial electron transfer |
| EET | Extracellular electron transfer |
| NAD | Nicotinamide adenine dinucleotide |
| MFC | Microbial fuel cells |
| MES | Microbial Electrosynthesis |
| MHC | Major histocompatibility complex |
| MWCNT | Multi walled carbon nanotubes |
| NADH | Nicotinamide adenine dinucleotide hydrogen |
| EAB | Electroactive Bacteria |
| MEC | Microbial electrolysis cell |
| IEM | Ion exchange membrane |
| PEM | Proton exchange membrane |
| ORR | Oxygen reduction reaction |
| VFA | Volatile fatty acid |
| SCFA | Short-chain fatty acids |
| MCFA | Medium-chain fatty acids |
| BES | Bioelectrochemical systems |
| SHE | Standard hydrogen electrode |
| MET | Microbial electrosynthesis technology |
| PBS | Phosphate buffer saline/solution |
| σ | Ionic conductor |
| U | Cell voltage |
| An. | Anodic faradic yield |
| γG | Energy yield |
| γH | Thermal yield |
References
- Flynn, J.M.; Ross, D.E.; Hunt, K.A.; Bond, D.R.; Gralnick, J.A. Enabling Unbalanced Fermentations by Using Engineered Electrode-Interfaced Bacteria. mBio 2010, 1. [Google Scholar] [CrossRef] [Green Version]
- Soccol, C.R.; Pandey, A.; Larroche, C.; Pandey, A.; Larroche, C. Fermentation Processes Engineering in the Food Industry; CRC Press: Boca Raton, FL, USA, 2013; ISBN 978-0-429-11200-3. [Google Scholar]
- Humphrey, A.E.; Lee, S.E. Industrial Fermentation: Principles, Processes, and Products. In Riegel’s Handbook of Industrial Chemistry; Springer: Dordrecht, The Netherlands, 1992; pp. 916–986. [Google Scholar]
- Albuquerque, M.G.E.; Eiroa, M.; Torres, C.; Nunes, B.R.; Reis, M.A.M. Strategies for the development of a side stream process for polyhydroxyalkanoate (PHA) production from sugar cane molasses. J. Biotechnol. 2007, 130, 411–421. [Google Scholar] [CrossRef] [PubMed]
- Pandey, A.; Kaushik, N.; Biswas, S. Bioprocess and Bioproducts: Technology Trends and Opportunities; Asiatech Publishers: New Delhi, India, 2009; ISBN 8187680202. [Google Scholar]
- Schievano, A.; PepéSciarria, T.; Vanbroekhoven, K.; De Wever, H.; Puig, S.; Andersen, S.J.; Rabaey, K.; Pant, D. Electro-Fermentation—Merging Electrochemistry with Fermentation in Industrial Applications. Trends Biotechnol. 2016, 34, 866–878. [Google Scholar] [CrossRef]
- Angenent, L.T.; Karim, K.; Al-Dahhan, M.H.; Wrenn, B.A.; Domíguez-Espinosa, R. Production of bioenergy and biochemicals from industrial and agricultural wastewater. Trends Biotechnol. 2004, 22, 477–485. [Google Scholar] [CrossRef] [PubMed]
- Bajracharya, S.; Vanbroekhoven, K.; Buisman, C.J.N.; Pant, D.; Strik, D.P.B.T.B. Application of gas diffusion biocathode in microbial electrosynthesis from carbon dioxide. Environ. Sci. Pollut. Res. 2016, 23, 22292–22308. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-P.; Zheng, P.; Sun, Z.-H.; Ni, Y.; Dong, J.-J.; Wei, P. Strategies of pH control and glucose-fed batch fermentation for production of succinic acid by Actinobacillus succinogenes CGMCC1593. J. Chem. Technol. Biotechnol. 2008, 83, 722–729. [Google Scholar] [CrossRef]
- Roy, S.; Schievano, A.; Pant, D. Electro-stimulated microbial factory for value added product synthesis. Bioresour. Technol. 2016, 213, 129–139. [Google Scholar] [CrossRef]
- Mohan, S.V.; Butti, S.K.; Amulya, K.; Dahiya, S.; Modestra, J.A. Waste Biorefinery: A New Paradigm for a Sustainable Bioelectro Economy. Trends Biotechnol. 2016, 34, 852–855. [Google Scholar] [CrossRef]
- Rabaey, K.; Angenent, L.; Schroder, U.; Keller, J. Bioelectrochemical Systems; IWA Publishing: London, UK, 2009; ISBN 978-1-84339-233-0. [Google Scholar]
- Thrash, J.C.; Coates, J.D. Review: Direct and Indirect Electrical Stimulation of Microbial Metabolism. Environ. Sci. Technol. 2008, 42, 3921–3931. [Google Scholar] [CrossRef]
- Lovley, D.R. Powering microbes with electricity: Direct electron transfer from electrodes to microbes. Environ. Microbiol. Rep. 2011, 3, 27–35. [Google Scholar] [CrossRef]
- Lee, H.-S.; Rittmann, B.E. Significance of Biological Hydrogen Oxidation in a Continuous Single-Chamber Microbial Electrolysis Cell. Environ. Sci. Technol. 2010, 44, 948–954. [Google Scholar] [CrossRef]
- Choi, O.; Sang, B.-I. Extracellular electron transfer from cathode to microbes: Application for biofuel production. Biotechnol. Biofuels 2016, 9, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nevin, K.P.; Hensley, S.A.; Franks, A.E.; Summers, Z.M.; Ou, J.; Woodard, T.L.; Snoeyenbos-West, O.L.; Lovley, D.R. Electrosynthesis of organic compounds from carbon dioxide is catalyzed by a diversity of acetogenic microorganisms. Appl. Environ. Microbiol. 2011, 77, 2882–2886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nikhil, G.N.; Venkata Subhash, G.; Yeruva, D.K.; Venkata Mohan, S. Synergistic yield of dual energy forms through biocatalyzedelectrofermentation of waste: Stoichiometric analysis of electron and carbon distribution. Energy 2015, 88, 281–291. [Google Scholar] [CrossRef]
- Logan, B.E. Scaling up microbial fuel cells and other bioelectrochemical systems. Appl. Microbiol. Biotechnol. 2010, 85, 1665–1671. [Google Scholar] [CrossRef]
- Rosenbaum, M.A.; Franks, A.E. Microbial catalysis in bioelectrochemical technologies: Status quo, challenges and perspectives. Appl. Microbiol. Biotechnol. 2014, 98, 509–518. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Ren, Z.J. A comprehensive review of microbial electrochemical systems as a platform technology. Biotechnol. Adv. 2013, 31, 1796–1807. [Google Scholar] [CrossRef]
- Leech, D.; Kavanagh, P.; Schuhmann, W. Enzymatic fuel cells: Recent progress. Electrochim. Acta 2012, 84, 223–234. [Google Scholar] [CrossRef]
- Habermüller, K.; Mosbach, M.; Schuhmann, W. Electron-transfer mechanisms in amperometric biosensors. Fresenius J. Anal. Chem. 2000, 366, 560–568. [Google Scholar] [CrossRef]
- Ghindilis, A.L.; Atanasov, P.; Wilkins, E. Enzyme-catalyzed direct electron transfer: Fundamentals and analytical applications. Electroanalysis 1997, 9, 661–674. [Google Scholar] [CrossRef]
- Kumar, A.; Hsu, L.H.-H.; Kavanagh, P.; Barrière, F.; Lens, P.N.L.; Lapinsonnière, L.; Lienhard V, J.H.; Schröder, U.; Jiang, X.; Leech, D. The ins and outs of microorganism–electrode electron transfer reactions. Nat. Rev. Chem. 2017, 1, 1–13. [Google Scholar] [CrossRef]
- d’Ippolito, G.; Dipasquale, L.; Vella, F.M.; Romano, I.; Gambacorta, A.; Cutignano, A.; Fontana, A. Hydrogen metabolism in the extreme thermophile Thermotoganeapolitana. Int. J. Hydrog. Energy 2010, 35, 2290–2295. [Google Scholar] [CrossRef]
- Manzini, E.; Scaglia, B.; Schievano, A.; Adani, F. Dark fermentation effectiveness as a key step for waste biomass refineries: Influence of organic matter macromolecular composition and bioavailability. Int. J. Energy Res. 2015, 39, 1519–1527. [Google Scholar] [CrossRef]
- Agler, M.T.; Wrenn, B.A.; Zinder, S.H.; Angenent, L.T. Waste to bioproduct conversion with undefined mixed cultures: The carboxylate platform. Trends Biotechnol. 2011, 29, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Rabaey, K.; Rozendal, R.A. Microbial electrosynthesis—Revisiting the electrical route for microbial production. Nat. Rev. Microbiol. 2010, 8, 706–716. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.; Aryal, N.; Sarma, P.M.; Vanbroekhoven, K.; Lal, B.; Benetton, X.D.; Pant, D. Bioelectrocatalyzed reduction of acetic and butyric acids via direct electron transfer using a mixed culture of sulfate-reducers drives electrosynthesis of alcohols and acetone. Chem. Commun. Camb. Engl. 2013, 49, 6495–6497. [Google Scholar] [CrossRef]
- Metabolic and Practical Considerations on Microbial Electrosynthesis.—Abstract—Europe PMC. Available online: https://europepmc.org/article/med/21353525 (accessed on 20 June 2020).
- Kracke, F.; Vassilev, I.; Krömer, J.O. Microbial electron transport and energy conservation—The foundation for optimizing bioelectrochemical systems. Front. Microbiol. 2015, 6. [Google Scholar] [CrossRef] [Green Version]
- Effect of Applied Voltage, Initial Concentration, and Natural Organic Matter on Sequential Reduction/Oxidation of Nitrobenzene by Graphite Electrodes—PubMed. Available online: https://pubmed.ncbi.nlm.nih.gov/22571797/ (accessed on 19 June 2020).
- Lovley, D.R. Syntrophy Goes Electric: Direct Interspecies Electron Transfer. Annu. Rev. Microbiol. 2017, 71, 643–664. [Google Scholar] [CrossRef]
- Speers, A.M.; Young, J.M.; Reguera, G. Fermentation of Glycerol into Ethanol in a Microbial Electrolysis Cell Driven by a Customized Consortium. Environ. Sci. Technol. 2014, 48, 6350–6358. [Google Scholar] [CrossRef]
- Moscoviz, R.; Toledo-Alarcón, J.; Trably, E.; Bernet, N. Electro-Fermentation: How to Drive Fermentation Using Electrochemical Systems. Trends Biotechnol. 2016, 34, 856–865. [Google Scholar] [CrossRef]
- Logan, B.E.; Call, D.; Cheng, S.; Hamelers, H.V.M.; Sleutels, T.H.J.A.; Jeremiasse, A.W.; Rozendal, R.A. Microbial Electrolysis Cells for High Yield Hydrogen Gas Production from Organic Matter. Environ. Sci. Technol. 2008, 42, 8630–8640. [Google Scholar] [CrossRef] [PubMed]
- Rago, L.; Ruiz, Y.; Baeza, J.A.; Guisasola, A.; Cortés, P. Microbial community analysis in a long-term membrane-less microbial electrolysis cell with hydrogen and methane production. Bioelectrochemistry 2015, 106, 359–368. [Google Scholar] [CrossRef] [PubMed]
- Park, D.H.; Laivenieks, M.; Guettler, M.V.; Jain, M.K.; Zeikus, J.G. Microbial Utilization of Electrically Reduced Neutral Red as the Sole Electron Donor for Growth and Metabolite Production. Appl. Environ. Microbiol. 1999, 65, 2912–2917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hara, M.; Onaka, Y.; Kobayashi, H.; Fu, Q.; Kawaguchi, H.; Vilcaez, J.; Sato, K. Mechanism of Electromethanogenic Reduction of CO2 by a Thermophilic Methanogen. Energy Procedia 2013, 37, 7021–7028. [Google Scholar] [CrossRef] [Green Version]
- Bader, J.; Mast-Gerlach, E.; Popović, M.K.; Bajpai, R.; Stahl, U. Relevance of microbial coculture fermentations in biotechnology. J. Appl. Microbiol. 2010, 109, 371–387. [Google Scholar] [CrossRef]
- Lenin Babu, M.; Venkata Subhash, G.; Sarma, P.N.; Venkata Mohan, S. Bio-electrolytic conversion of acidogenic effluents to biohydrogen: An integration strategy for higher substrate conversion and product recovery. Bioresour. Technol. 2013, 133, 322–331. [Google Scholar] [CrossRef]
- Gorby, Y.A.; Yanina, S.; McLean, J.S.; Rosso, K.M.; Moyles, D.; Dohnalkova, A.; Beveridge, T.J.; Chang, I.S.; Kim, B.H.; Kim, K.S.; et al. Electrically conductive bacterial nanowires produced by Shewanellaoneidensis strain MR-1 and other microorganisms. Proc. Natl. Acad. Sci. USA 2006, 103, 11358–11363. [Google Scholar] [CrossRef] [Green Version]
- Rahimnejad, M.; Adhami, A.; Darvari, S.; Zirepour, A.; Oh, S.-E. Microbial fuel cell as new technology for bioelectricity generation: A review. Alex. Eng. J. 2015, 54, 745–756. [Google Scholar] [CrossRef] [Green Version]
- Venkata Mohan, S.; Velvizhi, G.; Vamshi Krishna, K.; Lenin Babu, M. Microbial catalyzed electrochemical systems: A bio-factory with multi-facet applications. Bioresour. Technol. 2014, 165, 355–364. [Google Scholar] [CrossRef]
- Optimizing Treatment Performance of Microbial Fuel Cells by Reactor Staging Industrial & Engineering Chemistry Research. Available online: https://pubs.acs.org/doi/abs/10.1021/ie1004898 (accessed on 19 June 2020).
- Yu, J.; Song, X.; Wang, J.; Chen, J.; Liu, Q. BioelectrocatalyticDechlorination of Trichloroacetic Acid by Hemoglobin Modified Graphite Electrode in Aqueous Solutions of Different pH and Temperature. Int. J. Electrochem. Sci. 2012, 7, 11. [Google Scholar]
- Patil, S.A.; Hägerhäll, C.; Gorton, L. Electron transfer mechanisms between microorganisms and electrodes in bioelectrochemical systems. Bioanal. Rev. 2012, 4, 159–192. [Google Scholar] [CrossRef]
- Cao, Y.; Li, X.; Li, F.; Song, H. CRISPRi–sRNA: Transcriptional–Translational Regulation of Extracellular Electron Transfer in Shewanellaoneidensis. ACS Synth. Biol. 2017, 6, 1679–1690. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-S.; Dhar, B.R.; An, J.; Rittmann, B.E.; Ryu, H.; Santo Domingo, J.W.; Ren, H.; Chae, J. The Roles of Biofilm Conductivity and Donor Substrate Kinetics in a Mixed-Culture Biofilm Anode. Environ. Sci. Technol. 2016, 50, 12799–12807. [Google Scholar] [CrossRef] [PubMed]
- Croese, E.; Pereira, M.A.; Euverink, G.-J.W.; Stams, A.J.M.; Geelhoed, J.S. Analysis of the microbial community of the biocathode of a hydrogen-producing microbial electrolysis cell. Appl. Microbiol. Biotechnol. 2011, 92, 1083–1093. [Google Scholar] [CrossRef] [Green Version]
- Shi, L.; Squier, T.C.; Zachara, J.M.; Fredrickson, J.K. Respiration of metal (hydr)oxides by Shewanella and Geobacter: A key role for multihaem c-type cytochromes. Mol. Microbiol. 2007, 65, 12–20. [Google Scholar] [CrossRef] [Green Version]
- Gregory, K.B.; Lovley, D.R. Remediation and Recovery of Uranium from Contaminated Subsurface Environments with Electrodes. Environ. Sci. Technol. 2005, 39, 8943–8947. [Google Scholar] [CrossRef]
- Zuo, Y.; Xing, D.; Regan, J.M.; Logan, B.E. Isolation of the exoelectrogenic bacterium Ochrobactrumanthropi YZ-1 by using a U-tube microbial fuel cell. Appl. Environ. Microbiol. 2008, 74, 3130–3137. [Google Scholar] [CrossRef] [Green Version]
- Hernandez, M.E.; Newman, D.K. Extracellular electron transfer. Cell. Mol. Life Sci. CMLS 2001, 58, 1562–1571. [Google Scholar] [CrossRef]
- Rabaey, K.; Boon, N.; Höfte, M.; Verstraete, W. Microbial phenazine production enhances electron transfer in biofuel cells. Environ. Sci. Technol. 2005, 39, 3401–3408. [Google Scholar] [CrossRef]
- Kadier, A.; Simayi, Y.; Abdeshahian, P.; Azman, N.F.; Chandrasekhar, K.; Kalil, M.S. A comprehensive review of microbial electrolysis cells (MEC) reactor designs and configurations for sustainable hydrogen gas production. Alex. Eng. J. 2016, 55, 427–443. [Google Scholar] [CrossRef] [Green Version]
- Logan, B.E.; Hamelers, B.; Rozendal, R.; Schröder, U.; Keller, J.; Freguia, S.; Aelterman, P.; Verstraete, W.; Rabaey, K. Microbial fuel cells: Methodology and technology. Environ. Sci. Technol. 2006, 40, 5181–5192. [Google Scholar] [CrossRef] [PubMed]
- You, S.; Zhao, Q.; Zhang, J.; Jiang, J.; Wan, C.; Du, M.; Zhao, S. A graphite-granule membrane-less tubular air-cathode microbial fuel cell for power generation under continuously operational conditions. J. Power Sources 2007, 173, 172–177. [Google Scholar] [CrossRef]
- Sun, R.; Zhou, A.; Jia, J.; Liang, Q.; Liu, Q.; Xing, D.; Ren, N. Characterization of methane production and microbial community shifts during waste activated sludge degradation in microbial electrolysis cells. Bioresour. Technol. 2014, 175, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Sugnaux, M.; Happe, M.; Cachelin, C.P.; Gloriod, O.; Huguenin, G.; Blatter, M.; Fischer, F. Two stage bioethanol refining with multi litre stacked microbial fuel cell and microbial electrolysis cell. Bioresour. Technol. 2016, 221, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Gil-Carrera, L.; Escapa, A.; Carracedo, B.; Morán, A.; Gómez, X. Performance of a semi-pilot tubular microbial electrolysis cell (MEC) under several hydraulic retention times and applied voltages. Bioresour. Technol. 2013, 146, 63–69. [Google Scholar] [CrossRef]
- Clauwaert, P.; Verstraete, W. Methanogenesis in membraneless microbial electrolysis cells. Appl. Microbiol. Biotechnol. 2009, 82, 829–836. [Google Scholar] [CrossRef]
- Feng, Q.; Song, Y.-C. Surface Modification of a Graphite Fiber Fabric Anode for Enhanced Bioelectrochemical Methane Production. Energy Fuels 2016, 30, 6467–6474. [Google Scholar] [CrossRef]
- Feng, Y.; Zhang, Y.; Quan, X.; Chen, S. Enhanced anaerobic digestion of waste activated sludge digestion by the addition of zero valent iron. Water Res. 2013, 52, 242–250. [Google Scholar] [CrossRef]
- Asztalos, J.R.; Kim, Y. Enhanced digestion of waste activated sludge using microbial electrolysis cells at ambient temperature. Water Res. 2015, 87, 503–512. [Google Scholar] [CrossRef]
- Blasco-Gómez, R.; Batlle-Vilanova, P.; Villano, M.; Balaguer, M.D.; Colprim, J.; Puig, S. On the Edge of Research and Technological Application: A Critical Review of Electromethanogenesis. Int. J. Mol. Sci. 2017, 18, 874. [Google Scholar] [CrossRef]
- Kadier, A.; Simayi, Y.; Kalil, M.S.; Abdeshahian, P.; Hamid, A.A. A review of the substrates used in microbial electrolysis cells (MECs) for producing sustainable and clean hydrogen gas. Renew. Energy 2014, 71, 466–472. [Google Scholar] [CrossRef]
- Hu, K.; Chen, W.; Jia, S.; Wang, W.; Han, F. Enhanced Degradation of Waste Activated Sludge in Microbial Electrolysis Cell by Ultrasonic Treatment. Front. Microbiol. 2019, 10, 128. [Google Scholar] [CrossRef] [PubMed]
- Gajaraj, S.; Huang, Y.; Zheng, P.; Hu, Z. Methane production improvement and associated methanogenic assemblages in bioelectrochemically assisted anaerobic digestion. Biochem. Eng. J. 2017, 117, 105–112. [Google Scholar] [CrossRef]
- Siegert, M.; Yates, M.D.; Call, D.F.; Zhu, X.; Spormann, A.; Logan, B.E. Comparison of Nonprecious Metal Cathode Materials for Methane Production by Electromethanogenesis. ACS Sustain. Chem. Eng. 2014, 2, 910–917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.; Lee, B.; Tian, D.; Jun, H. Bioelectrochemical enhancement of methane production from highly concentrated food waste in a combined anaerobic digester and microbial electrolysis cell. Bioresour. Technol. 2018, 247, 226–233. [Google Scholar] [CrossRef]
- Su, M.; Wei, L.; Qiu, Z.; Jia, Q.; Shen, J. A graphene modified biocathode for enhancing hydrogen production. RSC Adv. 2015, 5, 32609–32614. [Google Scholar] [CrossRef]
- Yan, H.; Catania, C.; Bazan, G.C. Membrane-intercalating conjugated oligoelectrolytes: Impact on bioelectrochemical systems. Adv. Mater. 2015, 27, 2958–2973. [Google Scholar] [CrossRef]
- Jourdin, L.; Grieger, T.; Monetti, J.; Flexer, V.; Freguia, S.; Lu, Y.; Chen, J.; Romano, M.; Wallace, G.G.; Keller, J. High Acetic Acid Production Rate Obtained by Microbial Electrosynthesis from Carbon Dioxide. Environ. Sci. Technol. 2015, 49, 13566–13574. [Google Scholar] [CrossRef]
- Jourdin, L.; Freguia, S.; Donose, B.C.; Chen, J.; Wallace, G.G.; Keller, J.; Flexer, V. A novel carbon nanotube modified scaffold as an efficient biocathode material for improved microbial electrosynthesis. J. Mater. Chem. A 2014, 2, 13093–13102. [Google Scholar] [CrossRef] [Green Version]
- Choi, O.; Kim, T.; Woo, H.M.; Um, Y. Electricity-driven metabolic shift through direct electron uptake by electroactive heterotroph Clostridium pasteurianum. Sci. Rep. 2014, 4, 6961. [Google Scholar] [CrossRef]
- Reguera, G.; McCarthy, K.D.; Mehta, T.; Nicoll, J.S.; Tuominen, M.T.; Lovley, D.R. Extracellular electron transfer via microbial nanowires. Nature 2005, 435, 1098–1101. [Google Scholar] [CrossRef] [PubMed]
- Myers, C.R.; Myers, J.M. Localization of cytochromes to the outer membrane of anaerobically grown Shewanella putrefaciens MR-1. J. Bacteriol. 1992, 174, 3429–3438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marsili, E.; Baron, D.B.; Shikhare, I.D.; Coursolle, D.; Gralnick, J.A.; Bond, D.R. Shewanella secretes flavins that mediate extracellular electron transfer. Proc. Natl. Acad. Sci. USA 2008, 105, 3968–3973. [Google Scholar] [CrossRef] [Green Version]
- Cao, B.; Shi, L.; Brown, R.N.; Xiong, Y.; Fredrickson, J.K.; Romine, M.F.; Marshall, M.J.; Lipton, M.S.; Beyenal, H. Extracellular polymeric substances from Shewanella sp. HRCR-1 biofilms: Characterization by infrared spectroscopy and proteomics. Environ. Microbiol. 2011, 13, 1018–1031. [Google Scholar] [CrossRef] [PubMed]
- Ding, A.; Yang, Y.; Sun, G.; Wu, D. Impact of applied voltage on methane generation and microbial activities in an anaerobic microbial electrolysis cell (MEC). Chem. Eng. J. 2016, 283, 260–265. [Google Scholar] [CrossRef]
- Chen, Y.; Yu, B.; Yin, C.; Zhang, C.; Dai, X.; Yuan, H.; Zhu, N. Biostimulation by direct voltage to enhance anaerobic digestion of waste activated sludge. RSC Adv. 2015, 6, 1581–1588. [Google Scholar] [CrossRef]
- Zhen, G.; Kobayashi, T.; Lu, X.; Kumar, G.; Xu, K. Biomethane recovery from Egeria densa in a microbial electrolysis cell-assisted anaerobic system: Performance and stability assessment. Chemosphere 2016, 149, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Gildemyn, S.; Verbeeck, K.; Slabbinck, R.; Andersen, S.J.; Prévoteau, A.; Rabaey, K. Integrated Production, Extraction, and Concentration of Acetic Acid from CO2 through Microbial Electrosynthesis. Environ. Sci. Technol. Lett. 2015, 2, 325–328. [Google Scholar] [CrossRef]
- Lai, B.; Yu, S.; Bernhardt, P.V.; Rabaey, K.; Virdis, B.; Krömer, J.O. Anoxic metabolism and biochemical production in Pseudomonas putida F1 driven by a bioelectrochemical system. Biotechnol. Biofuels 2016, 9, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, M.; Freguia, S.; Dennis, P.G.; Keller, J.; Rabaey, K. Development of bioelectrocatalytic activity stimulates mixed-culture reduction of glycerol in a bioelectrochemical system. Microb. Biotechnol. 2015, 8, 483–489. [Google Scholar] [CrossRef]
- ElMekawy, A.; Hegab, H.M.; Losic, D.; Saint, C.P.; Pant, D. Applications of graphene in microbial fuel cells: The gap between promise and reality. Renew. Sustain. Energy Rev. 2017, 72, 1389–1403. [Google Scholar] [CrossRef]
- Moscoviz, R.; Trably, E.; Bernet, N. Electro-fermentation triggering population selection in mixed-culture glycerol fermentation. Microb. Biotechnol. 2017, 11, 74–83. [Google Scholar] [CrossRef]
- Farid, M.A.; El-Enshasy, H.A.; El-Deen, A.M.N. Alcohol production from starch by mixed cultures of Aspergillus awamori and immobilized Saccharomyces cerevisiae at different agitation speeds. J. Basic Microbiol. 2002, 42, 162–171. [Google Scholar] [CrossRef]
- Palanisamy, K. State of the art and future concept of food waste fermentation to bioenergy. Renew. Sustain. Energy Rev. 2016, 53, 547–557. [Google Scholar]
- Zhao, Z.; Zhang, Y.; Wang, L.; Quan, X. Potential for direct interspecies electron transfer in an electric-anaerobic system to increase methane production from sludge digestion. Sci. Rep. 2015, 5, 11094. [Google Scholar] [CrossRef] [PubMed]
- Hafez, H.M. Method and System for Electro-assisted Hydrogen Production from Organic Material. U.S. Patent 9,458,474, 4 October 2016. [Google Scholar]
- Selembo, P.A.; Perez, J.M.; Lloyd, W.A.; Logan, B.E. High hydrogen production from glycerol or glucose by electrohydrogenesis using microbial electrolysis cells. Int. J. Hydrogen Energy 2009, 34, 5373–5381. [Google Scholar] [CrossRef]
- Butanol Production from Food Waste: A Novel Process for Producing Sustainable Energy and Reducing Environmental Pollution Biotechnology for Biofuels Full Text. Available online: https://biotechnologyforbiofuels.biomedcentral.com/articles/10.1186/s13068-015-0332-x (accessed on 19 June 2020).
- Lee, J.W.; Kim, H.U.; Choi, S.; Yi, J.; Lee, S.Y. Microbial production of building block chemicals and polymers. Curr. Opin. Biotechnol. 2011, 22, 758–767. [Google Scholar] [CrossRef] [PubMed]
- Sindhu, R.; Ammu, B.; Binod, P.; Deepthi, S.K.; Ramachandran, K.B.; Soccol, C.R.; Pandey, A. Production and characterization of poly-3-hydroxybutyrate from crude glycerol by Bacillus sphaericus NII 0838 and improving its thermal properties by blending with other polymers. Braz. Arch. Biol. Technol. 2011, 54, 783–794. [Google Scholar] [CrossRef] [Green Version]
- Cok, B.; Tsiropoulos, I.; Roes, A.L.; Patel, M.K. Succinic acid production derived from carbohydrates: An energy and greenhouse gas assessment of a platform chemical toward a bio-based economy. Biofuels Bioprod. Biorefin. 2014, 8, 16–29. [Google Scholar] [CrossRef]
- Choi, O.; Um, Y.; Sang, B.-I. Butyrate production enhancement by Clostridium tyrobutyricum using electron mediators and a cathodic electron donor. Biotechnol. Bioeng. 2012, 109, 2494–2502. [Google Scholar] [CrossRef]
- Park, D.H.; Zeikus, J.G. Utilization of Electrically Reduced Neutral Red by Actinobacillus succinogenes: Physiological Function of Neutral Red in Membrane-Driven Fumarate Reduction and Energy Conservation. J. Bacteriol. 1999, 181, 2403–2410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, T.S.; Kim, B.H. Electron flow shift inClostridiumacetobutylicum fermentation by electrochemically introduced reducing equivalent. Biotechnol. Lett. 1988, 10, 123–128. [Google Scholar] [CrossRef]
- Schmitz, S.; Nies, S.; Wierckx, N.; Blank, L.M.; Rosenbaum, M.A. Engineering mediator-based electroactivity in the obligate aerobic bacterium Pseudomonas putida KT2440. Front. Microbiol. 2015, 6, 284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandit, A.V.; Mahadevan, R. In silico characterization of microbial electrosynthesis for metabolic engineering of biochemicals. Microb. Cell Factories 2011, 10, 76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathew, A.S.; Wang, J.; Luo, J.; Yau, S.-T. Enhanced ethanol production via electrostatically accelerated fermentation of glucose using Saccharomyces cerevisiae. Sci. Rep. 2015, 5, 15713. [Google Scholar] [CrossRef] [Green Version]
- Deval, A.S.; Parikh, H.A.; Kadier, A.; Chandrasekhar, K.; Bhagwat, A.M.; Dikshit, A.K. Sequential microbial activities mediated bioelectricity production from distillery wastewater using bio-electrochemical system with simultaneous waste remediation. Int. J. Hydrogen Energy 2017, 42, 1130–1141. [Google Scholar] [CrossRef]
- Thygesen, A.; Thomsen, A.B.; Possemiers, S.; Verstraete, W. Integration of microbial electrolysis cells (MECs) in the biorefinery for production of ethanol, H2 and phenolics. Waste Biomass Valoriz. 2010, 1, 9–20. [Google Scholar] [CrossRef]
- Steinbusch, K.J.J.; Hamelers, H.V.M.; Schaap, J.D.; Kampman, C.; Buisman, C.J.N. Bioelectrochemical ethanol production through mediated acetate reduction by mixed cultures. Environ. Sci. Technol. 2010, 44, 513–517. [Google Scholar] [CrossRef]
- Lee, W.S.; Chua, A.S.M.; Yeoh, H.K.; Ngoh, G.C. A review of the production and applications of waste-derived volatile fatty acids. Chem. Eng. J. 2013, 235, 83–99. [Google Scholar] [CrossRef]
- Venkata Mohan, S.; Mohanakrishna, G.; Goud, R.K.; Sarma, P.N. Acidogenic fermentation of vegetable based market waste to harness biohydrogen with simultaneous stabilization. Bioresour. Technol. 2009, 100, 3061–3068. [Google Scholar] [CrossRef]
- Sarkar, O.; Venkata Mohan, S. Deciphering acidogenic process towards biohydrogen, biohythane, and short chain fatty acids production: Multi-output optimization strategy. Biofuel Res. J. 2016, 3, 458–469. [Google Scholar] [CrossRef] [Green Version]
- Zhou, M.; Yan, B.; Wong, J.W.C.; Zhang, Y. Enhanced volatile fatty acids production from anaerobic fermentation of food waste: A mini-review focusing on acidogenic metabolic pathways. Bioresour. Technol. 2018, 248, 68–78. [Google Scholar] [CrossRef] [PubMed]
- Gildemyn, S.; Molitor, B.; Usack, J.G.; Nguyen, M.; Rabaey, K.; Angenent, L.T. Upgrading syngas fermentation effluent using Clostridium kluyveri in a continuous fermentation. Biotechnol. Biofuels 2017, 10, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Eerten-Jansen, M.C.A.A.; Ter Heijne, A.; Grootscholten, T.I.M.; Steinbusch, K.J.J.; Sleutels, T.H.J.A.; Hamelers, H.V.M.; Buisman, C.J.N. Bioelectrochemical Production of Caproate and Caprylate from Acetate by Mixed Cultures. ACS Sustain. Chem. Eng. 2013, 1, 513–518. [Google Scholar] [CrossRef]
- Steinbusch, K.J.J.; Hamelers, H.V.M.; Plugge, C.M.; Buisman, C.J.N. Biological formation of caproate and caprylate from acetate: Fuel and chemical production from low grade biomass. Energy Environ. Sci. 2010, 4, 216–224. [Google Scholar] [CrossRef]
- Kucek, L.A.; Xu, J.; Nguyen, M.; Angenent, L.T. Waste Conversion into n-Caprylate and n-Caproate: Resource Recovery from Wine Lees Using Anaerobic Reactor Microbiomes and In-line Extraction. Front. Microbiol. 2016, 7, 1892. [Google Scholar] [CrossRef] [Green Version]
- Grootscholten, T.I.M.; dal Borgo, F.K.; Hamelers, H.V.M.; Buisman, C.J.N. Promoting chain elongation in mixed culture acidification reactors by addition of ethanol. Biomass Bioenergy 2013, 48, 10–16. [Google Scholar] [CrossRef]
- Logan, B.E.; Rabaey, K. Conversion of wastes into bioelectricity and chemicals by using microbial electrochemical technologies. Science 2012, 337, 686–690. [Google Scholar] [CrossRef] [Green Version]
- Wong, J.W.-C.; Tyagi, R.D.; Pandey, A. Current Developments in Biotechnology and Bioengineering: Solid Waste Management; Elsevier Science: Amsterdam, The Netherlands, 2016; ISBN 978-0-444-63664-5. [Google Scholar]
- Montpart, N.; Rago, L.; Baeza, J.A.; Guisasola, A. Hydrogen production in single chamber microbial electrolysis cells with different complex substrates. Water Res. 2015, 68, 601–615. [Google Scholar] [CrossRef]
- Pasupuleti, S.B.; Sarkar, O.; Mohan, S.V. Upscaling of biohydrogen production process in semi-pilot scale biofilm reactor: Evaluation with food waste at variable organic loads. Int. J. Hydrogen Energy 2014, 39, 7587–7596. [Google Scholar] [CrossRef]
- Schröder, U.; Harnisch, F.; Angenent, L.T. Microbial electrochemistry and technology: Terminology and classification. Energy Environ. Sci. 2015, 8, 513–519. [Google Scholar] [CrossRef]
- Zhao, Z.; Sun, J.; Yu, Q.; Ma, W.; Quan, X.; Zhang, Y. Enhanced decomposition of waste activated sludge via anodic oxidation for methane production and bioenergy recovery. Int. Biodeterior. Biodegrad. 2016, 106, 161–169. [Google Scholar] [CrossRef]
- Van Steendam, C.; Smets, I.; Skerlos, S.; Raskin, L. Improving anaerobic digestion via direct interspecies electron transfer requires development of suitable characterization methods. Curr. Opin. Biotechnol. 2019, 57, 183–190. [Google Scholar] [CrossRef]
- Cheng, S.; Xing, D.; Call, D.F.; Logan, B.E. Direct biological conversion of electrical current into methane by electromethanogenesis. Environ. Sci. Technol. 2009, 43, 3953–3958. [Google Scholar] [CrossRef]
- Liu, H.; Grot, S.; Logan, B.E. Electrochemically Assisted Microbial Production of Hydrogen from Acetate. Environ. Sci. Technol. 2005, 39, 4317–4320. [Google Scholar] [CrossRef] [PubMed]
- Logan, B.E. Exoelectrogenic bacteria that power microbial fuel cells. Nat. Rev. Microbiol. 2009, 7, 375–381. [Google Scholar] [CrossRef]
- Lovley, D.R.; Ueki, T.; Zhang, T.; Malvankar, N.S.; Shrestha, P.M.; Flanagan, K.A.; Aklujkar, M.; Butler, J.E.; Giloteaux, L.; Rotaru, A.-E.; et al. Geobacter: The microbe electric’s physiology, ecology, and practical applications. Adv. Microb. Physiol. 2011, 59, 1–100. [Google Scholar] [CrossRef]
- Freguia, S.; Rabaey, K.; Yuan, Z.; Keller, J. Syntrophic Processes Drive the Conversion of Glucose in Microbial Fuel Cell Anodes. Environ. Sci. Technol. 2008, 42, 7937–7943. [Google Scholar] [CrossRef]
- Venkata Mohan, S.; Modestra, J.A.; Amulya, K.; Butti, S.K.; Velvizhi, G. A Circular Bioeconomy with Biobased Products from CO2 Sequestration. Trends Biotechnol. 2016, 34, 506–519. [Google Scholar] [CrossRef]
- Butti, S.K.; Mohan, S.V. Autotrophic biorefinery: Dawn of the gaseous carbon feedstock. FEMS Microbiol. Lett. 2017, 364. [Google Scholar] [CrossRef]
- Modestra, J.A.; Babu, M.C.L.; Mohan, S.V. Electro-fermentation of real-field acidogenic spent wash effluents for additional biohydrogen production with simultaneous treatment in a microbial electrolysis cell. Sep. Purif. Technol. 2015. [Google Scholar] [CrossRef]
- Villano, M.; Aulenta, F.; Ciucci, C.; Ferri, T.; Giuliano, A.; Majone, M. Bioelectrochemical reduction of CO2 to CH4 via direct and indirect extracellular electron transfer by a hydrogenophilic methanogenic culture. Bioresour. Technol. 2010, 101, 3085–3090. [Google Scholar] [CrossRef] [PubMed]
- Su, M. Production of Acetate from Carbon Dioxide in Bioelectrochemical Systems Based on Autotrophic Mixed Culture. J. Microbiol. Biotechnol. 2013, 23, 1140–1146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganigué, R.; Puig, S.; Batlle-Vilanova, P.; Balaguer, M.D.; Colprim, J. Microbial electrosynthesis of butyrate from carbon dioxide. Chem. Commun. Camb. Engl. 2015, 51, 3235–3238. [Google Scholar] [CrossRef] [PubMed]
- Kadier, A.; Simayi, Y.; Chandrasekhar, K.; Ismail, M.; Kalil, M.S. Hydrogen gas production with an electroformed Ni mesh cathode catalysts in a single-chamber microbial electrolysis cell (MEC). Int. J. Hydrogen Energy 2015, 40, 14095–14103. [Google Scholar] [CrossRef]
- Kumar, G.; Saratale, R.G.; Kadier, A.; Sivagurunathan, P.; Zhen, G.; Kim, S.H.; Saratale, G.D. A review on bio-electrochemical systems (BESs) for the syngas and value added biochemicals production. Chemosphere 2017, 177, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Rozendal, R.A.; Hamelers, H.V.M.; Euverink, G.J.W.; Metz, S.J.; Buisman, C.J.N. Principle and perspectives of hydrogen production through biocatalyzed electrolysis. Int. J. Hydrogen Energy 2006, 31, 1632–1640. [Google Scholar] [CrossRef]
- Pedersen, M.B.; Iversen, S.L.; Sørensen, K.I.; Johansen, E. The long and winding road from the research laboratory to industrial applications of lactic acid bacteria. FEMS Microbiol. Rev. 2005, 29, 611–624. [Google Scholar] [CrossRef]
- Lechardeur, D.; Cesselin, B.; Fernandez, A.; Lamberet, G.; Garrigues, C.; Pedersen, M.; Gaudu, P.; Gruss, A. Using heme as an energy boost for lactic acid bacteria. Curr. Opin. Biotechnol. 2011, 22, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Vargas, M.; Kashefi, K.; Blunt-Harris, E.L.; Lovley, D.R. Microbiological evidence for Fe(III) reduction on early Earth. Nature 1998, 395, 65–67. [Google Scholar] [CrossRef]
- Rozendal, R.A.; Jeremiasse, A.W.; Hamelers, H.V.M.; Buisman, C.J.N. Hydrogen Production with a Microbial Biocathode. Environ. Sci. Technol. 2008, 42, 629–634. [Google Scholar] [CrossRef] [PubMed]
- Kadier, A.; Jain, P.; Lai, B.; Kalil, M.S.; Kondaveeti, S.; Alabbosh, K.F.S.; Abu-Reesh, I.M.; Mohanakrishna, G. Biorefinery perspectives of microbial electrolysis cells (MECs) for hydrogen and valuable chemicals production through wastewater treatment. Biofuel Res. J. 2020, 7, 1128–1142. [Google Scholar] [CrossRef] [Green Version]
- Venkata Mohan, S.; Mohanakrishna, G.; Sarma, P.N. Integration of acidogenic and methanogenic processes for simultaneous production of biohydrogen and methane from wastewater treatment. Int. J. Hydrogen Energy 2008, 33, 2156–2166. [Google Scholar] [CrossRef]
- Rathinam, N.K.; Bibra, M.; Salem, D.R.; Sani, R.K. Thermophiles for biohydrogen production in microbial electrolytic cells. Bioresour. Technol. 2019, 277, 171–178. [Google Scholar] [CrossRef]
- Shemfe, M.; Gadkari, S.; Yu, E.; Rasul, S.; Scott, K.; Head, I.M.; Gu, S.; Sadhukhan, J. Life cycle, techno-economic and dynamic simulation assessment of bioelectrochemical systems: A case of formic acid synthesis. Bioresour. Technol. 2018, 255, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Gil-Carrera, L.; Escapa, A.; Mehta, P.; Santoyo, G.; Guiot, S.R.; Morán, A.; Tartakovsky, B. Microbial electrolysis cell scale-up for combined wastewater treatment and hydrogen production. Bioresour. Technol. 2013, 130, 584–591. [Google Scholar] [CrossRef]
- Heidrich, E.S.; Edwards, S.R.; Dolfing, J.; Cotterill, S.E.; Curtis, T.P. Performance of a pilot scale microbial electrolysis cell fed on domestic wastewater at ambient temperatures for a 12month period. Bioresour. Technol. 2014, 173, 87–95. [Google Scholar] [CrossRef] [Green Version]
- San-Martín, M.I.; Sotres, A.; Alonso, R.M.; Díaz-Marcos, J.; Morán, A.; Escapa, A. Assessing anodic microbial populations and membrane ageing in a pilot microbial electrolysis cell. Int. J. Hydrogen Energy 2019, 44, 17304–17315. [Google Scholar] [CrossRef]
- Redwood, M.D.; Orozco, R.L.; Majewski, A.J.; Macaskie, L.E. Electro-extractive fermentation for efficient biohydrogen production. Bioresour. Technol. 2012, 107, 166–174. [Google Scholar] [CrossRef] [Green Version]
- Rabaey, K.; Verstraete, W. Microbial fuel cells: Novel biotechnology for energy generation. Trends Biotechnol. 2005, 23, 291–298. [Google Scholar] [CrossRef]
- Sivagurunathan, P.; Kuppam, C.; Mudhoo, A.; Saratale, G.D.; Kadier, A.; Zhen, G.; Chatellard, L.; Trably, E.; Kumar, G. A comprehensive review on two-stage integrative schemes for the valorization of dark fermentative effluents. Crit. Rev. Biotechnol. 2018, 38, 868–882. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Timeline showing development in microbial electrosynthesis and electron transfer systems. (A) Chronological development of biocathode applied technologies and (B) advancements in anodic microbial electrocatalysis and extracellular electron transfer in microbial electrochemical cell [26].
Figure 1.
Timeline showing development in microbial electrosynthesis and electron transfer systems. (A) Chronological development of biocathode applied technologies and (B) advancements in anodic microbial electrocatalysis and extracellular electron transfer in microbial electrochemical cell [26].

Figure 2.
Diagrammatic representation of different electrofermentation process and its operation. (A) Microbial electrosynthesis; (B) Microbial electrolysis cell.
Figure 2.
Diagrammatic representation of different electrofermentation process and its operation. (A) Microbial electrosynthesis; (B) Microbial electrolysis cell.

Figure 3.
A diagram representing the principles of electro-fermentation, Adapted from ref. [16].
Figure 3.
A diagram representing the principles of electro-fermentation, Adapted from ref. [16].

Figure 4.
A diagrammatic representation of the possible mechanisms that can occur during anodic electro-fermentation (EF). (A) The substrate is directly converted into the product and the excess of electron is fully dissipated at the anode through mechanisms of extracellular electron transfer. (B) The electron excess generated during the formation of the oxidized products is not fully dissipated at the anode and a section of the substrate is utilized for this purpose. The electron elimination at the anode leads to the reducing NADH/NAD+ ratio resulting in the compensating cellular regulation favoring pathways to regulate NADH. (C) The fermentative microorganism (yellow) consumes the substrate but is not able to interact with the anode. The electroactive microorganism (red) acts as a mediator between the fermentative microorganism and the anode through interspecies electron transfer mechanisms [36].
Figure 4.
A diagrammatic representation of the possible mechanisms that can occur during anodic electro-fermentation (EF). (A) The substrate is directly converted into the product and the excess of electron is fully dissipated at the anode through mechanisms of extracellular electron transfer. (B) The electron excess generated during the formation of the oxidized products is not fully dissipated at the anode and a section of the substrate is utilized for this purpose. The electron elimination at the anode leads to the reducing NADH/NAD+ ratio resulting in the compensating cellular regulation favoring pathways to regulate NADH. (C) The fermentative microorganism (yellow) consumes the substrate but is not able to interact with the anode. The electroactive microorganism (red) acts as a mediator between the fermentative microorganism and the anode through interspecies electron transfer mechanisms [36].

Figure 5.
Interaction of microbe-electrode with electron mediators showing direct interspecies electron transfer depicting the electron transfer by cytochromes and enzymatic electron transfer [45].
Figure 5.
Interaction of microbe-electrode with electron mediators showing direct interspecies electron transfer depicting the electron transfer by cytochromes and enzymatic electron transfer [45].

Figure 6.
A diagram representing different extracellular electron transfer (EET) mechanisms in Shewanella spp., wherein electrons extracted at the inner membrane are transferred by the c-type cytochrome CymA to the outer membrane and can be transported to an electron acceptor by metal-reduction (MTR) proteins to complete EET and reduction reactions facilitated by the mediators at the cathode.
Figure 6.
A diagram representing different extracellular electron transfer (EET) mechanisms in Shewanella spp., wherein electrons extracted at the inner membrane are transferred by the c-type cytochrome CymA to the outer membrane and can be transported to an electron acceptor by metal-reduction (MTR) proteins to complete EET and reduction reactions facilitated by the mediators at the cathode.

Figure 7.
Diagrammatic representations of continuous stirred tank electro-fermenter [16].
Figure 7.
Diagrammatic representations of continuous stirred tank electro-fermenter [16].

Figure 8.
Diagrammatic representation of immobilized cells with different configurations: (A) fluidized bed, rotating, (B) bed packed bed, and biofilters (C,D). (D) the reactor contains in situ separation systems, along with three-chamber configurations. IEM, Ion-exchange membrane [16].
Figure 8.
Diagrammatic representation of immobilized cells with different configurations: (A) fluidized bed, rotating, (B) bed packed bed, and biofilters (C,D). (D) the reactor contains in situ separation systems, along with three-chamber configurations. IEM, Ion-exchange membrane [16].


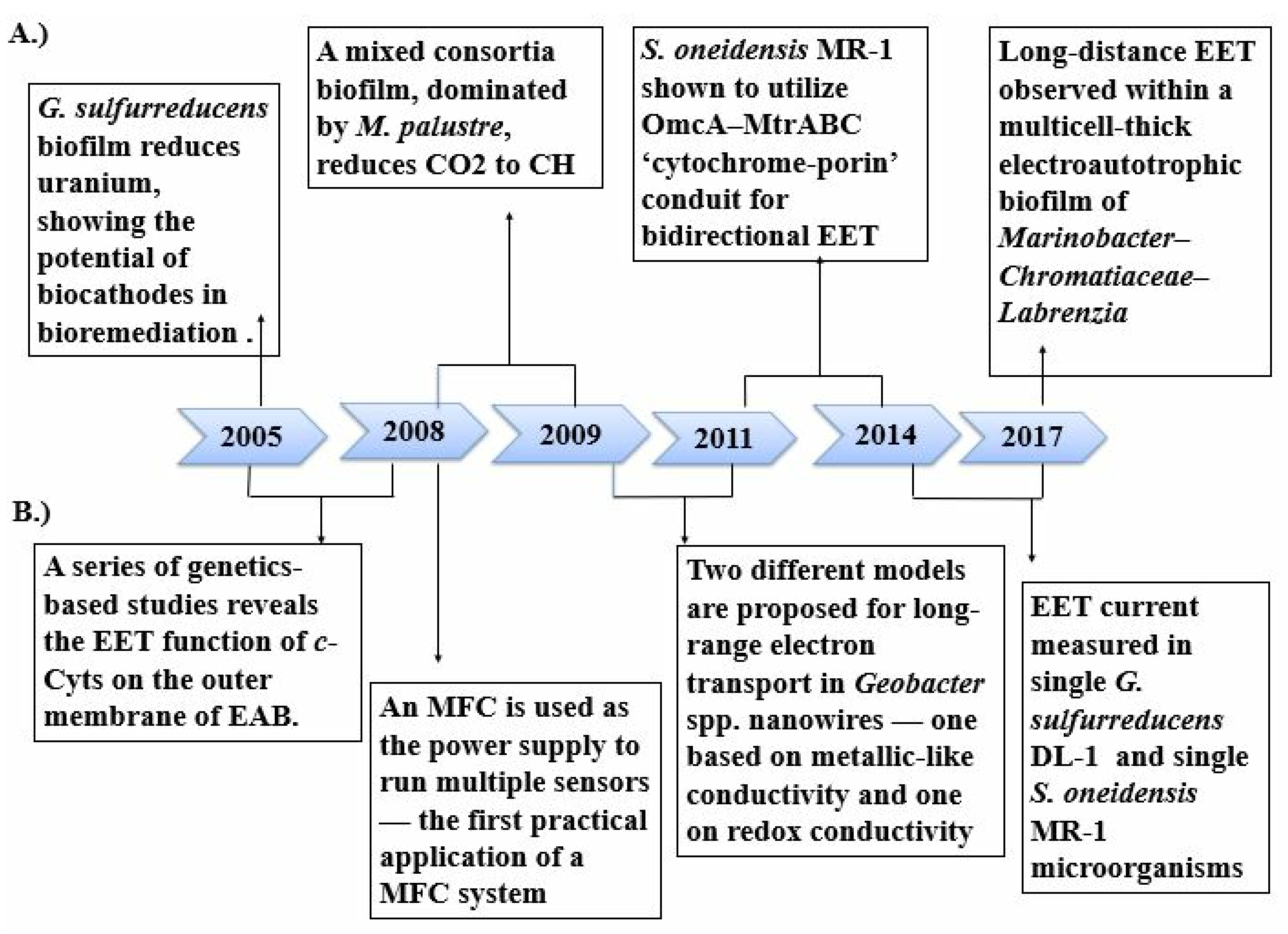{kind=link}
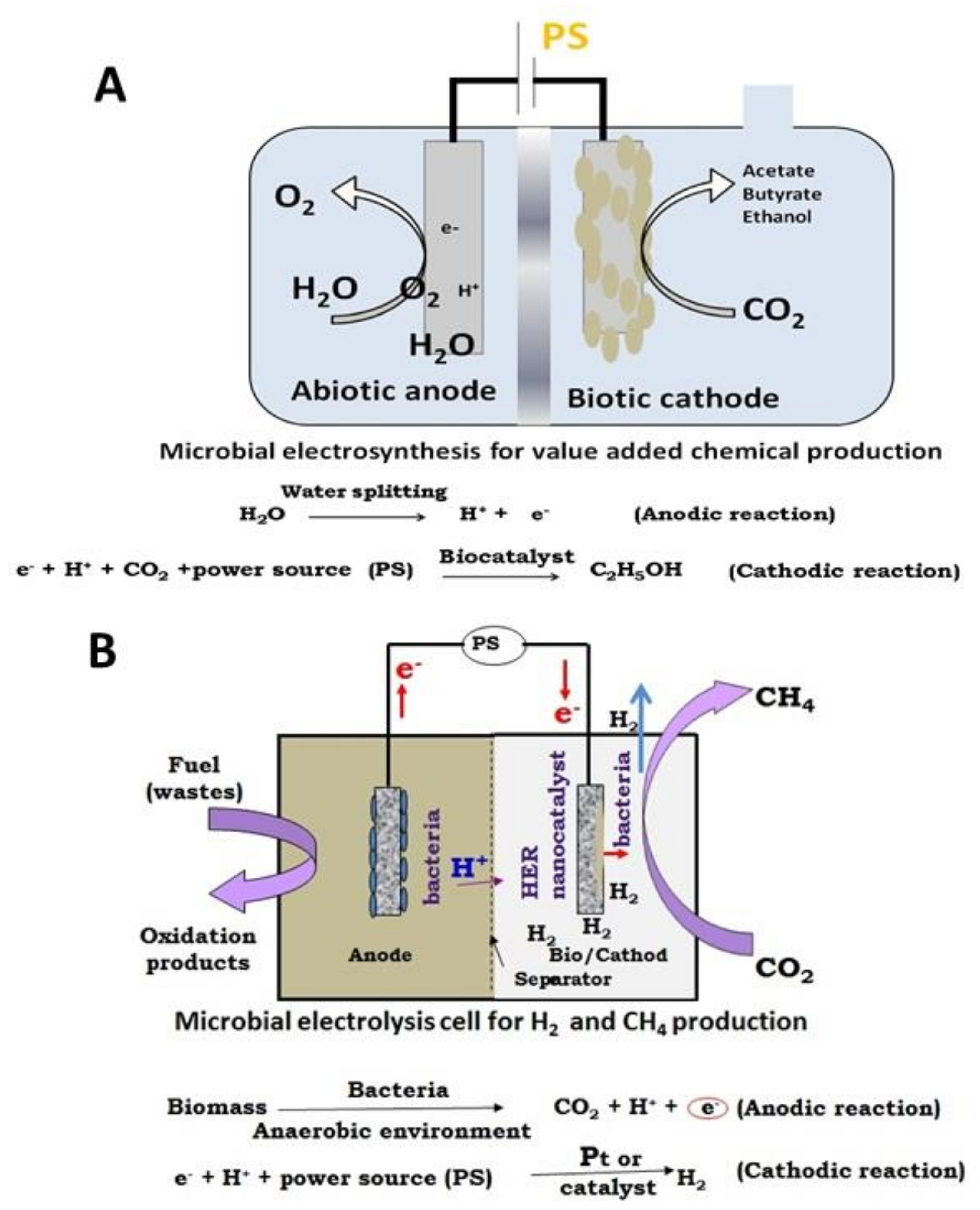{kind=link}
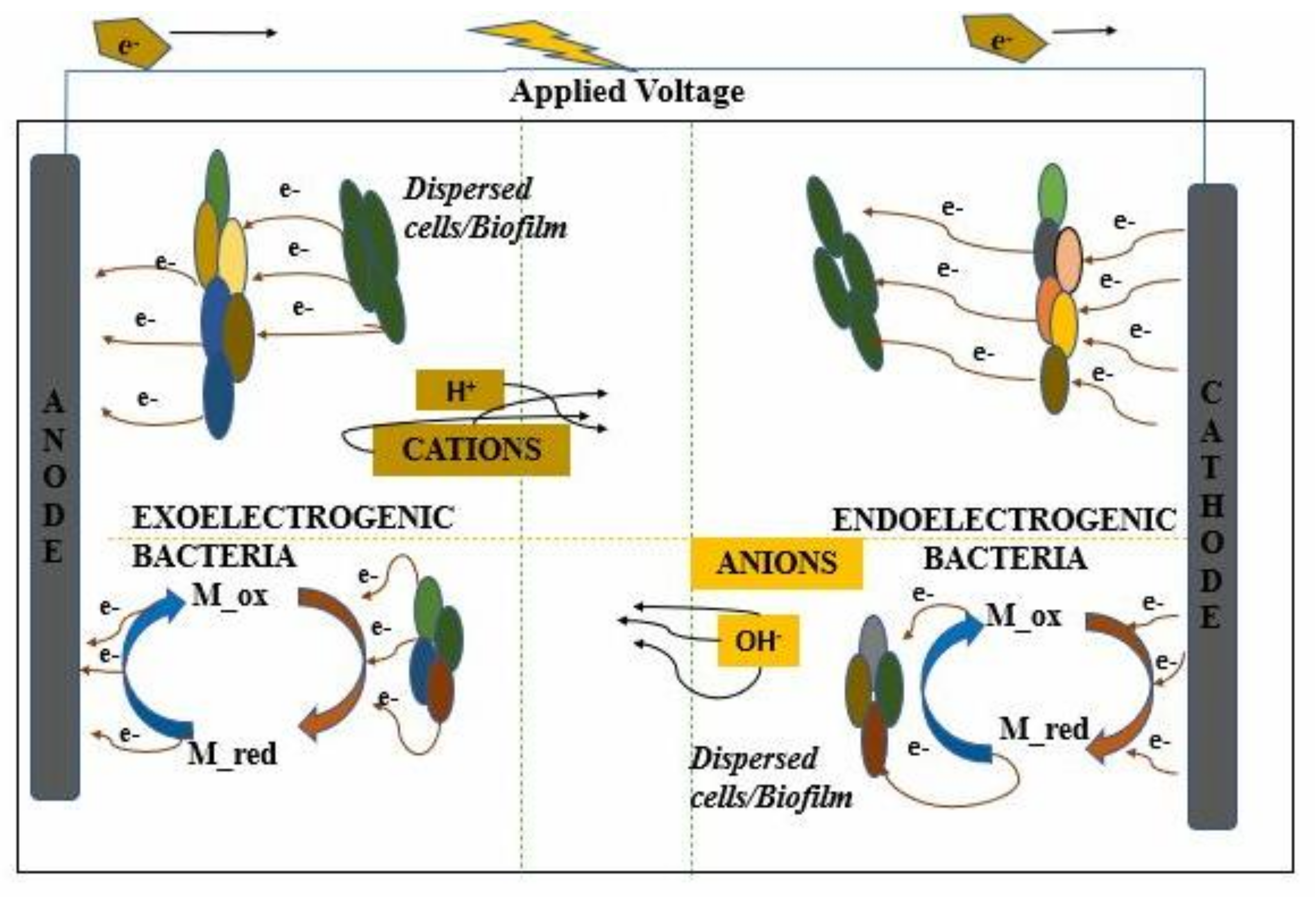{kind=link}
{kind=link}
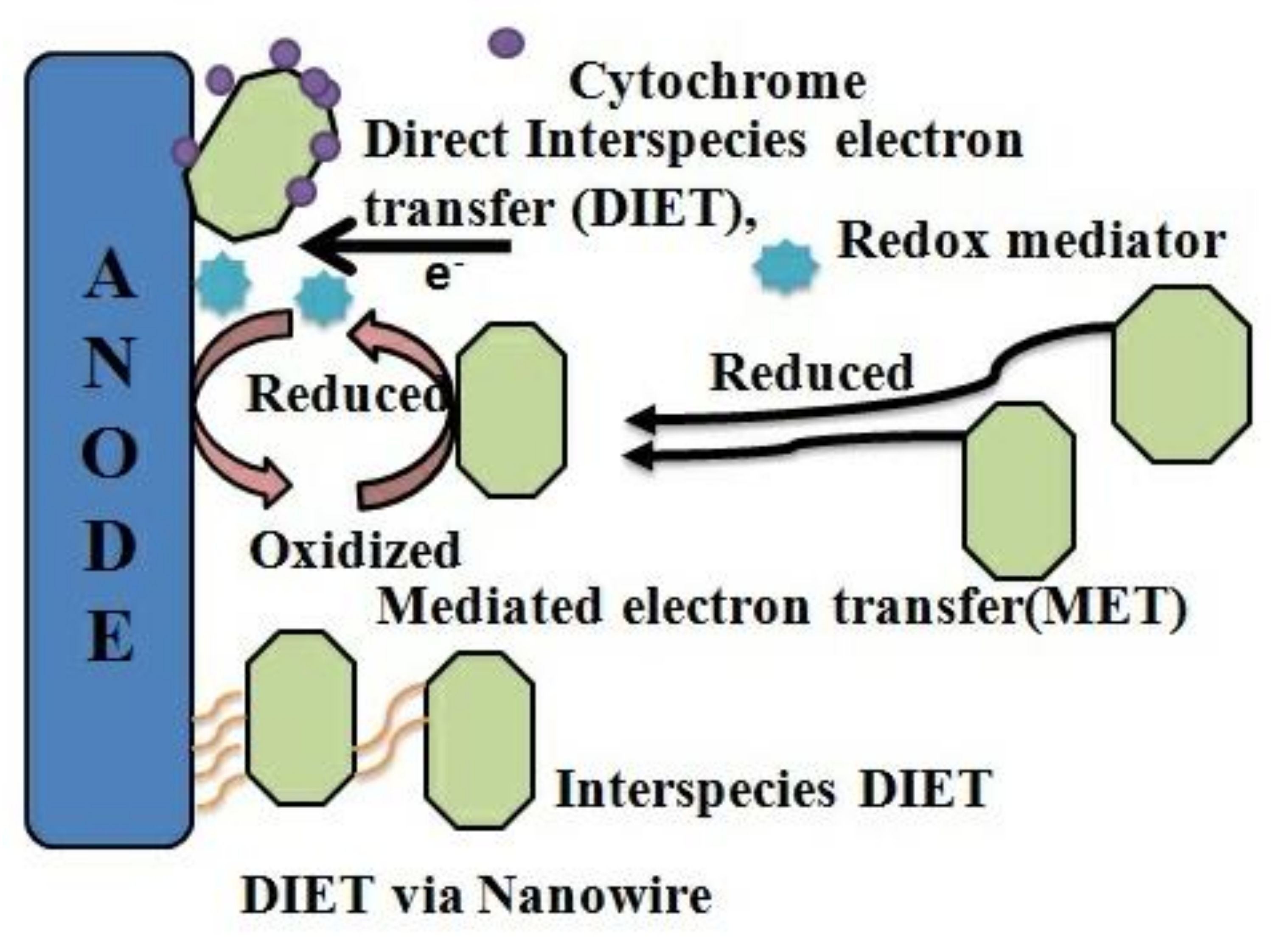{kind=link}
{kind=link}
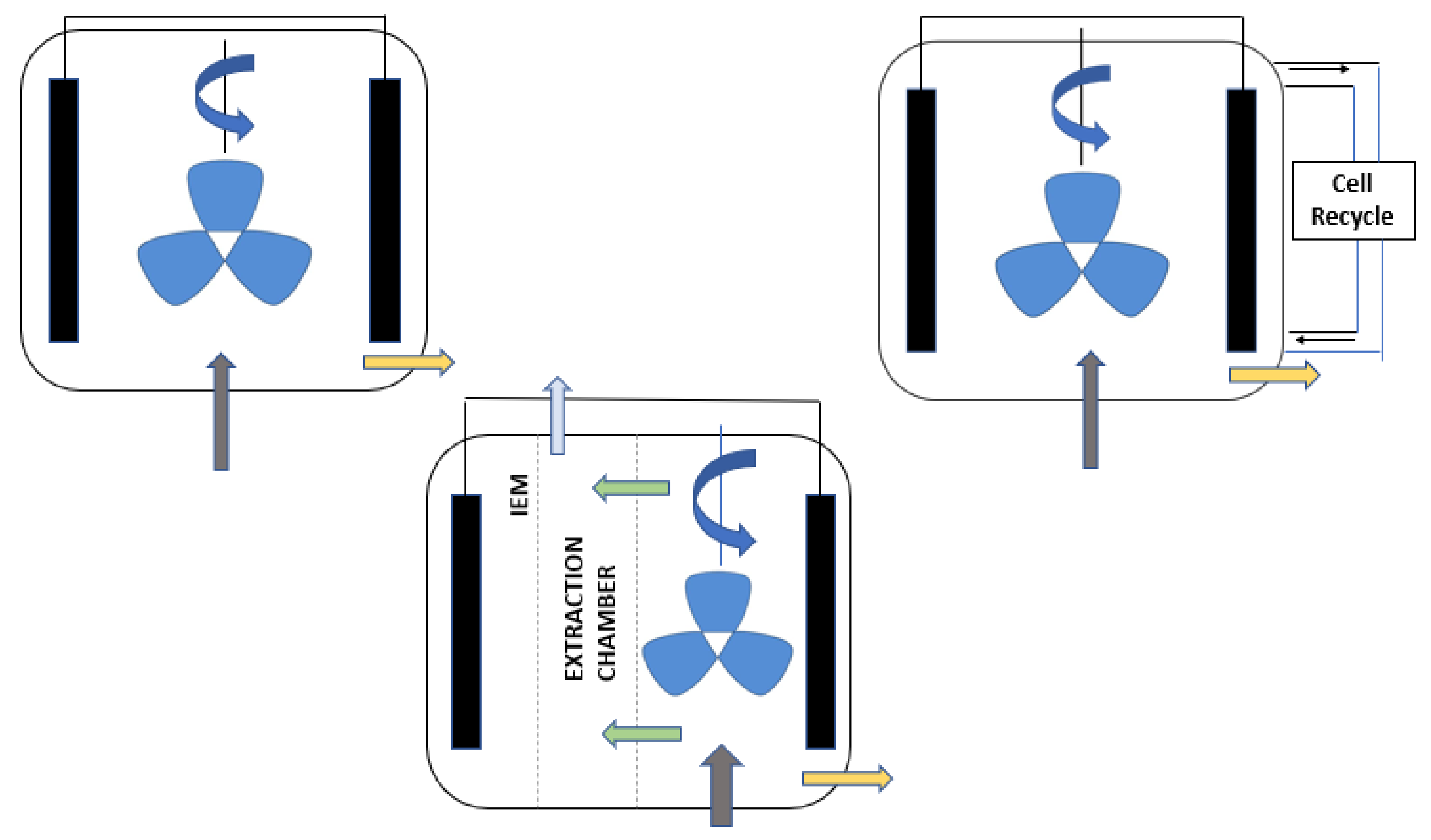{kind=link}
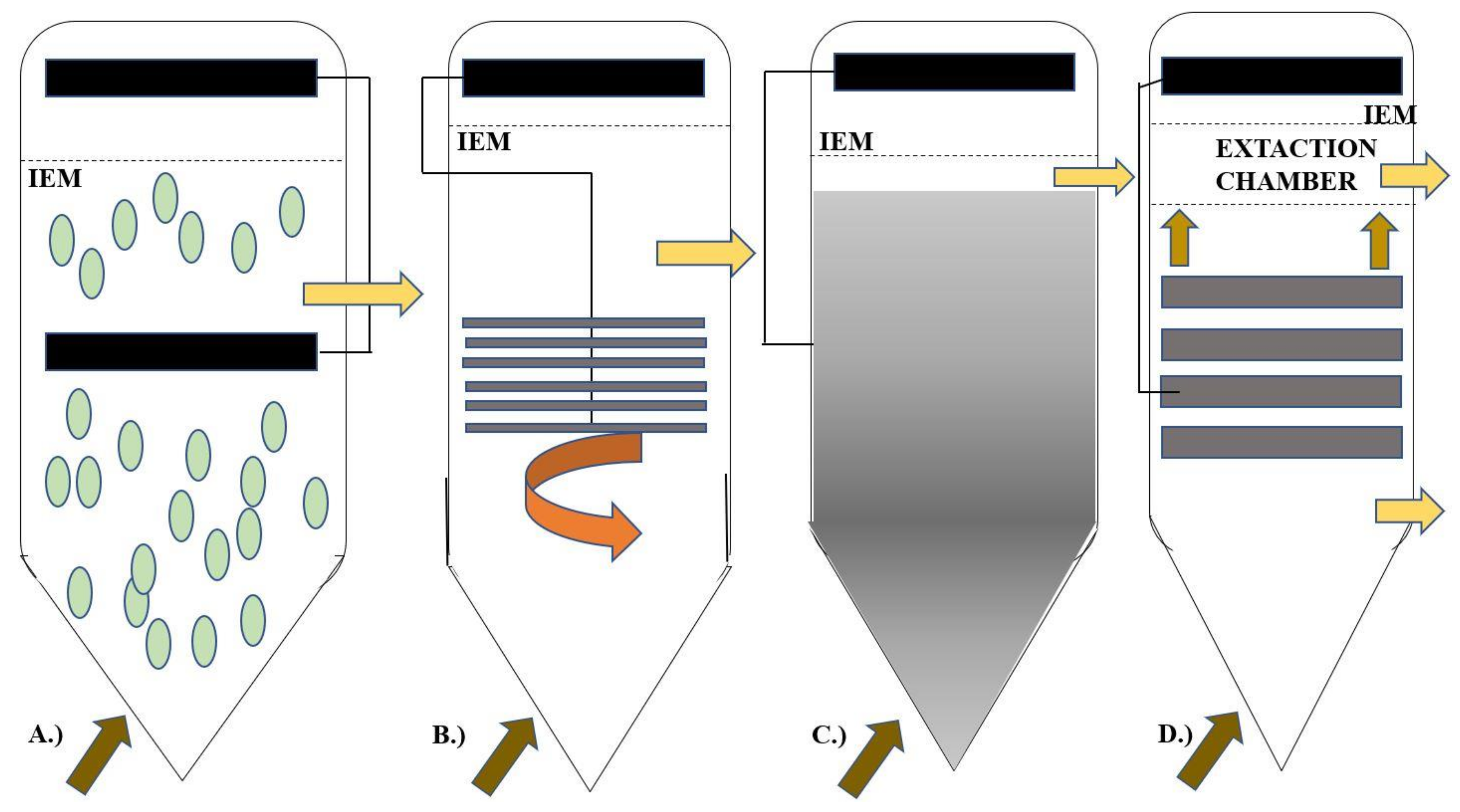{kind=link}
Table 1.
Operating parameters for specific product formation [43].
Table 1.
Operating parameters for specific product formation [43].
| Sr.No. | Substrate | Applied Voltage (V) | Product Synthesized | Production Rate | Electrofermentation | Microbial Catalyst | References |
|---|---|---|---|---|---|---|---|
| 1 | Food waste | −0.6 V | Acetate, butyrate, andpropionate | 4595 mg/L d | Anodic | Mixed microbial culture | [84] |
| 2 | CO2 | −0.9 V Ecat | CH4 | 9.2 L/m2 d | Cathodic | Mixed enriched culture biocatalyst | [145] |
| 3 | Acetate | 0.5 V | H2 | 0.1 L/m2d | Anodic | Mixed enriched culture biocatalyst | [127] |
| 4 | CO2 | −0.9 to −1.1 | H2 and acetate | 0.38 mM d | Cathodic | Mixed culture domestic wastewater treatment plant sludge | [146] |
| 5 | CO2 | þ0.197 V/SHE | Butyrate, ethanol, and butanol | 1.82 MMC/d | Cathodic | Clostridium | [147] |
Table 2.
Characteristics of microbial electrolysis cell (MEC) pilots with volumes of at least several liters.
Table 2.
Characteristics of microbial electrolysis cell (MEC) pilots with volumes of at least several liters.
| Cathode Surface Area (cm2) | V (L) | σ(S·m−1) | Anolyte | Current Density (A·m−2) | U (V) | Gas Production (L·d−1·m−2) | Energy Yields (–) | Faradaic Yields (%) | Ref. | |||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CH4 | H2 | γH | γG | An. | Cath H2 | |||||||
| 3024 | 5 × 2 | 0.7 | Wastewater | 0.61 | 1.08 | yes | 1.0 | 0.22 | 0.19 | 19 | 23 | [146] |
| 3360 | 120 | 0.17 | Wastewater acetate | 0.3 max | 1.1 | 0.06 | 3.6 | NA | 0.69 | 16 | 55 | [147] |
| 840 | 2 × 2 | 0.05 | Wastewater | 0.24 | 1.0 | 0.23 | 1.06 | 1.1 | 0.5 | NA | 44 | [146] |
| 4466 | 33 | NA | Sewage sludge | 0.01 | 2.0 | 0.28 | 0 | 6.45 | 0 | NA | NA | [88] |
| 1800 | 16 | 2.6 | Pig slurry, acetate | 1.75 | 1.0 | <2% | 17.8 | 1.5 | 1.25 | 7–9 | 78 | [148] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Bhagchandanii, D.D.; Babu, R.P.; Sonawane, J.M.; Khanna, N.; Pandit, S.; Jadhav, D.A.; Khilari, S.; Prasad, R. A Comprehensive Understanding of Electro-Fermentation. Fermentation 2020, 6, 92. https://doi.org/10.3390/fermentation6030092
AMA Style
Bhagchandanii DD, Babu RP, Sonawane JM, Khanna N, Pandit S, Jadhav DA, Khilari S, Prasad R. A Comprehensive Understanding of Electro-Fermentation. Fermentation. 2020; 6(3):92. https://doi.org/10.3390/fermentation6030092
Chicago/Turabian StyleBhagchandanii, Drishti Dinesh, Rishi Pramod Babu, Jayesh M. Sonawane, Namita Khanna, Soumya Pandit, Dipak A. Jadhav, Santimoy Khilari, and Ram Prasad. 2020. "A Comprehensive Understanding of Electro-Fermentation" Fermentation 6, no. 3: 92. https://doi.org/10.3390/fermentation6030092
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.