
Evaluation of Aphid Resistance and Oleoresin Production in Indigenous Tropical Pine (Pinus merkusii Jungh. and de Vriese)

 ,
,  ,
,  , , ,
, , , 
 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
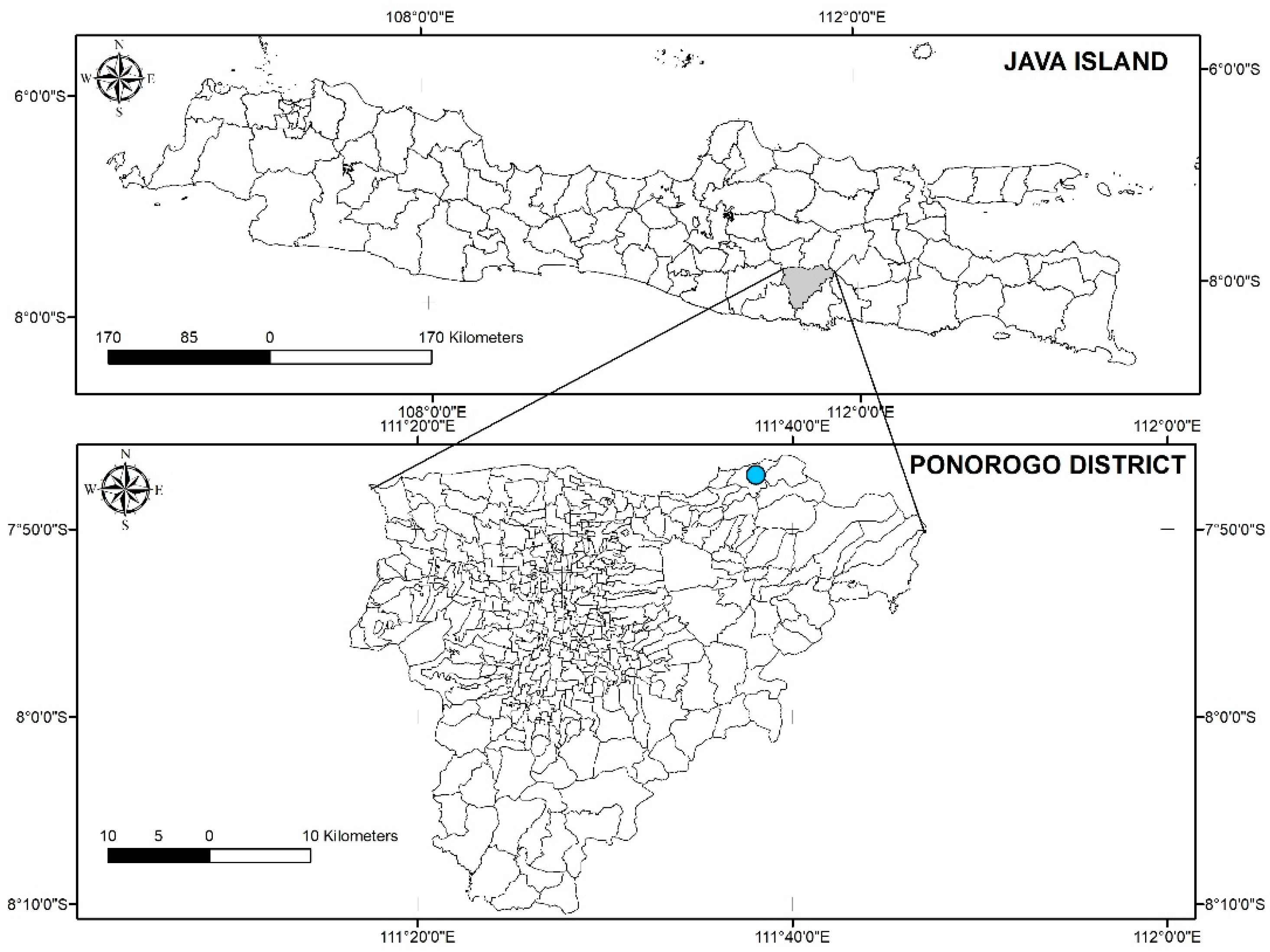
2.1. Trial Site
2.2. Experimental Design of Progeny Test
2.3. Data Collection
2.4. Data Analyses
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Imanuddin, R.; Hidayat, A.; Rachmat, H.H.; Turjaman, M.; Pratiwi; Nurfatriani, F.; Indrajaya, Y.; Susilowati, A. Reforestation and sustainable management of Pinus merkusii forest plantation in Indonesia: A review. Forests 2020, 11, 1235. [Google Scholar] [CrossRef]
- Hardiyanto, E.; Danarto, S. Ex situ Conservation of Pinus merkusii in Java, Indonesia. In In Situ and Ex Situ Conservation of Commercial Tropical Trees; Faculty of Forestry, Gadjah Mada University: Sleman, Indonesia; International Tropical Timber Organization: Yokohama, Japan, 2001; pp. 263–269. [Google Scholar]
- Susilowati, A.; Siregar, I.Z.; Supriyanto, S.; Wahyudi, I.; Corryanti, C. Genetic variation, heritability and correlation between resin production character of Pinus merkusii high resin yielder (Hry). Biotropia 2013, 20, 122–133. [Google Scholar] [CrossRef]
- Muslimin, I. Korelasi genetik pertumbuhan dan produksi getah pada uji keturunan Pinus merkusii di KPH Banyumas Barat (Genetic correlation of growth and resin yield in progeny test Pinus merkusii Jungh. et de Vriese at KPH Banyumas Barat). J. Penelit. Kehutan. Sumatrana 2017, 1, 22–34. [Google Scholar] [CrossRef] [Green Version]
- Susilowati, A.; Rachmat, H.H.; Siregar, I.Z.; Supriyanto. Genetic diversity of resin yielder Pinus merkusii from West Java—Indonesia revealed by microsatellites marker. IOP Conf. Ser. Earth Environ. Sci. 2018, 122, 012060. [Google Scholar] [CrossRef] [Green Version]
- Leksono, B. Heritabilitas dan perolehan genetik produksi getah, diameter batang, bentuk batang dan tipe percabangan Pinus merkusii jungh et de vriese (Heritability and genetic gain of resin production, stem diameter, stem and branching types of Pinus merkusii jungh et de vriese). Bul. Penelit. Kehutan. BPK Pematang. Siantar 1996, 11, 223–236. [Google Scholar]
- Chilima, C.Z.; Leather, S.R. Within-tree and seasonal distribution of the pine woolly aphid Pineus boerneri on Pinus kesiya trees. Agric. For. Entomol. 2001, 44, 139–145. [Google Scholar] [CrossRef]
- Rodas, C.A.; Serna, R.; Bolaños, M.D.; Granados, G.M.; Michael, J.; Hurley, B.P. Incidence and host susceptibility of Pineus boerneri (Hemiptera: Adelgidae) in Colombian pine plantations biology, incidence and host susceptibility of Pineus boerneri (Hemiptera: Adelgidae). South. For. 2015, 77, 165–171. [Google Scholar] [CrossRef]
- Siregar, U.; Diputra, I. Keragaman genetik Pinus merkusii Jungh. et de Vriese strain Tapanuli berdasarkan penanda mikrosatelit (Diversity of Pinus merkusii Jungh. et de Vriese of Tapanuli strain based on microsatellite markers). J. Silvikultur Trop. 2013, 4, 88–99. [Google Scholar] [CrossRef]
- Supriyanto; Iskandar, T. Penilaian kesehatan kebun benih semai Pinus merkusii dengan metode Fhm (Forest Health Monitoring) Di KPH Sumedang (Health assessment for seedling seed orchard of Pinus merkusii using forest health monitoring (FHM) Method in KPH Sumedang). J. Silvikultur Trop. 2018, 9, 99–108. [Google Scholar] [CrossRef]
- Sumantoro, P. Serangan Hama Kutu Lilin (Pineus Boerneri Annand) pada Tanaman Uji Keturunan Pinus Merkusii Generasi II Umur 9 Tahun di Tampomas Sumedang (Pineus Boerneri Annand Pest Attack at the Second Generation of Pinus Merkusii Seedling Seed Orchards in Tampomas Sumedang); Gadjah Mada University: Sleman, Indonesia, 2012. [Google Scholar]
- Negara, H.K.; Rachmawati, N.; Payung, D. Identifikasi Kerusakan Pohon Pinus Di Hutan Kota BanjarBaru (Pine tree damage identification at Banjarbaru city forest). J. Sylva Sci. 2019, 2, 635–644. [Google Scholar]
- Latumahina, F. Diagnosis of the Type of Pests of Tusam (Pinus merkusii Jung et de Vriese). IOP Conf. Ser. Earth Environ. Sci. 2020, 519, 012001. [Google Scholar] [CrossRef]
- McClure, M.S. Distribution and damage of two pineus species (Homoptera: Adelgidae) on red pine in New England. Ann. Entomol. Soc. Am. 1982, 75, 150–157. [Google Scholar] [CrossRef]
- Nageleisen, L.-M.; Bouget, C. Forest Insect Studies: Methods and Techniques. Key Considerations for Standardisation. An Overview of the Reflections of the “Entomological Forest In-Ventories” Working Group (Inv. Ent. For.); Bouget, C., Nageleisen, L.-M., Bonneil, P., Eds.; ONF: Paris, France, 2009; Volume 19, ISBN 9782842073435. [Google Scholar]
- Lazzari, S.M.N.; Cardoso, J.T. Pineus boerneri annand, 1928 (Hemiptera: Adelgidae) A new species to brazil: Morphology of eggs, nymphs and adults. Rev. Bras. Entomol. 2011, 55, 459–466. [Google Scholar] [CrossRef] [Green Version]
- Day, R.K.; Kairo, M.T.K.; Abraham, Y.J.; Kfir, R.; Murphy, S.T.; Mutitu, K.E.; Chilima, C.Z. Biological control of homopteran pests of conifers in Africa. In Biological Control in IPM Systems in Africa; Neuenschwander, C., Borgemeister, C., Langewald, J., Eds.; CAB International: Wallingford, UK, 2009; pp. 101–112. [Google Scholar]
- Perhutani. Laporan Tahunan Perhutani 2018 (Perhutani Annual Report of 2018); Perhutani: Jakarta, Indonesia, 2018. [Google Scholar]
- Sukadaryati, S. Pemanenan getah pinus menggunakan tiga cara penyadapan (Harvesting pine sap using three methods of tapping). J. Penelit. Has. Hutan 2014, 32, 62–70. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Ghasemi, A.; Zahediasi, S. Normality tests for statistical analysis: A guide for non-statisticians. Int. J. Endocrinol. Metab. 2012, 10, 486–489. [Google Scholar] [CrossRef] [Green Version]
- Beyene, K.M.; Bekele, S.A. Assessing univariate and multivariate homogeneity of variance: A guide for practitioners. Math. Theory Model. 2016, 6, 13–17. [Google Scholar]
- Pagnuco, I.A.; Pastore, J.I.; Abras, G.; Brun, M.; Ballarin, V.L. Analysis of genetic association using hierarchical clustering and cluster validation indices. Genomics 2017, 109, 438–445. [Google Scholar] [CrossRef]
- Steel, R.G.D.; Torrie, J.H. Principles and Procedures of Statistics: A Biometrical Approach, 2nd ed.; Mc Graw-Hill Book Company: Singapore, 1981. [Google Scholar]
- Woolaston, R.; Kanowski, P.; Nikles, D. Genetic parameter estimates for Pinus caribaea var. hondurensis in Coastal Quensland, Australia. Silvae Genet. 1990, 39, 21–28. [Google Scholar]
- Pswarayi; Barnes, R.D.; Birks, J.S.; Kanowski, P. Genetic parameter estimates for production and quality traits of Pinus elliottii Engelm. var. elliottii in Zimbabwe. AGRIS 1998, 45, 216–222. [Google Scholar]
- Liu, Q.; Zhou, Z.; Fan, H.; Liu, Y. Genetic variation and correlation among resin yield, growth, and morphologic traits of Pinus massoniana. Silvae Genet. 2013, 62, 38–44. [Google Scholar] [CrossRef]
- Westbrook, J.W.; Resende, M.F.R., Jr.; Munoz, P.; Walker, A.R.; Wegrzyn, J.L.; Nelson, C.D.; Neale, D.B.; Kirst, M.; Huber, D.A.; Gezan, S.A.; et al. Association genetics of oleoresin flow in loblolly pine: Discovering genes and predicting phenotype for improved resistance to bark beetles and bioenergy potential. New Phytol. 2013, 199, 89–100. [Google Scholar] [CrossRef] [PubMed]
- Paramanik, T.; Bhattacharyya, S. Gum production and its sustainable harvest from forest: A Review. Ambient Sci. 2021, 8, 1–5. [Google Scholar] [CrossRef]
- Daniawati, E. Pengaruh Berbagai Penutupan Tumbuhan Bawah dan arah Sadap Terhadap Produktivitas Getah Pinus Merkusii (Effect of Understorey Closure and Tapping Direction to the Pinus Merkusii Oleoresin Production); Institute Pertanian Bogor: Bogor, Indonesia, 2016. [Google Scholar]
- Rodrigues, K.C.S.; Fett-Neto, A.G. Oleoresin yield of Pinus elliottii in a subtropical climate: Seasonal variation and effect of auxin and salicylic acid-based stimulant paste. Ind. Crops Prod. 2009, 30, 316–320. [Google Scholar] [CrossRef]
- Indu Bhusban, D.; Pratibha, K.; Abhishek, R. Effects of temperature and relative humidity on Ethephon induced gum exudation in Acacia nilotica. Asian J. Multidiscip. Stud. 2014, 2, 115–116. [Google Scholar]
- Adam, l.O.M.; Ballal, M.E.M.; Fadl, K.E.M. Effect of tapping direction in relation to sun light on gum arabic Acacia senegal (L.) willd. yields in north kordofan state, sudan. For. Trees Livelihoods 2009, 19, 185–191. [Google Scholar] [CrossRef]
- Purwanto; Handarto; Cahyono, L.R. Mulyono Evaluasi uji keturunan Pinus merkusii tahan hama kutu lilin (Evaluation of Pinus merkusii seedling seed orchards to Aphid resistance). Bul. Penelit. Hutan Lestari Prod. 2017, 20, 8–12. [Google Scholar]
- Roberds, J.H.; Strom, B.L.; Hain, F.P.; Gwaze, D.P.; Mckeand, S.E.; Lott, L.H. Estimates of genetic parameters for oleoresin and growth traits in juvenile loblolly pine. Can. J. For. Res. 2009, 33, 2469–2476. [Google Scholar] [CrossRef]
- Fries, A. Genetic parameters, genetic gain and correlated responses in growth, fibre dimensions and wood density in a Scots pine breeding population. Ann. For. Sci. 2012, 69, 783–794. [Google Scholar] [CrossRef] [Green Version]
- Callister, A.N.; England, N.; Collins, S. Predicted genetic gain and realised gain in stand volume of Eucalyptus globulus. Tree Genet. Genomes 2013, 9, 361–375. [Google Scholar] [CrossRef]
- Liang, D.; Ding, C.; Zhao, G.; Leng, W.; Zhang, M.; Zhao, X.; Qu, G. Variation and selection analysis of Pinus koraiensis clones in northeast China. J. For. Res. 2018, 29, 611–622. [Google Scholar] [CrossRef]
- Strom, B.L.; Goyer, R.A.; Ingram, L.L., Jr.; Boyd, G.D.L.; Lott, L.H. Oleoresin characteristics of progeny of loblolly pines that escaped attack by the southern pine beetle. For. Ecol. Manag. 2002, 158, 169–178. [Google Scholar] [CrossRef]
- Liu, J.J.; Schoettle, A.W.; Sniezko, R.A.; Sturrock, R.N.; Zamany, A.; Williams, H.; Ha, A.; Chan, D.; Danchok, B.; Savin, D.P.; et al. Genetic mapping of Pinus flexilis major gene (Cr4) for resistance to white pine blister rust using transcriptome-based SNP genotyping. BMC Genom. 2016, 17, 753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nurtjahjaningsih, I.L.G.; Saito, Y.; Tsuda, Y.; Ide, Y. Genetic diversity of parental and offspring populations in a Pinus merkusii seedling seed orchard detected by microsatellite markers. Bul. Tokyo Univ. For. 2007, 118, 1–14. [Google Scholar]
- Siregar, I.Z.; Hattemer, H.H. Patterns of Genetic Structure and Variation of Merkus Pine (Pinus merkusii) in Indonesia. J. Trop. For. Sci. 2004, 16, 160–172. [Google Scholar]



{kind=link}
{kind=link}
| Source of Variation | df | Sum of Squares | Mean Square | F | p-Value |
|---|---|---|---|---|---|
| Diameter at breast height | |||||
| Block | 9 | 186 | 20.67 | 4.88 | <0.001 ** |
| Family | 33 | 105.74 | 3.20 | 1.50 | 0.048 * |
| Block × family | 217 | 880.24 | 4.06 | 1.90 | <0.001 ** |
| Error | 213 | 455.78 | 2.14 | ||
| West oleoresin production | |||||
| Block | 9 | 1678.90 | 186.55 | 8.12 | <0.001 ** |
| Family | 33 | 957.60 | 29.02 | 1.26 | 0.170 ns |
| Block × family | 217 | 6887.10 | 31.74 | 1.38 | 0.003 * |
| Error | 213 | 4018.30 | 22.96 | ||
| East oleoresin production | |||||
| Block | 9 | 958.9 | 106.55 | 5.39 | <0.001 ** |
| Family | 33 | 1098.1 | 33.28 | 1.68 | 0.017 * |
| Block × family | 217 | 4477.8 | 20.64 | 1.04 | 0.217 ns |
| Error | 213 | 3459 | 19.77 | ||
| Average oleoresin production | |||||
| Block | 9 | 1646.20 | 182.91 | 11.47 | <0.001 ** |
| Family | 33 | 971.70 | 29.44 | 1.85 | 0.005 ns |
| Block × family | 217 | 5077.80 | 23.40 | 1.47 | 0.002 ** |
| Error | 213 | 3397.30 | 15.95 | ||
| Plant resistance to aphids | |||||
| Block | 9 | 38.36 | 4.26 | 15.28 | <0.001 ** |
| Family | 33 | 17.86 | 0.54 | 1.93 | 0.046 * |
| Block × family | 217 | 102.88 | 0.47 | 1.69 | <0.001 ** |
| Error | 213 | 56.67 | 0.28 | ||
| Family | DBH (cm) | WOP (g) | EOP (g) | AOP (g) | PR Score |
|---|---|---|---|---|---|
| Mean ± sd | Mean ± sd | Mean ± sd | Mean ± sd | Mean ± sd | |
| F1 | 17.6 ± 2.0 ab | 7.8 ± 5.4 bc | 9.5 ± 7.6 abc | 7.2 ± 5.3 bcde | 3.8 ± 0.8 abcd |
| F2 | 17.6 ± 1.7 ab | 9.9 ± 6.4 bc | 9.7 ± 3.2 abc | 8.7 ± 3.8 bcd | 3.6 ± 0.8 abcd |
| F3 | 16.7 ± 1.7 ab | 9.0 ± 7.3 bc | 8.3 ± 4.1 abc | 8.6 ± 5.2 bcde | 3.7 ± 1 abcd |
| F4 | 16.9 ± 1.9 ab | 9.4 ± 5.7 bc | 7.2 ± 3.5 abc | 7.5 ± 4 bcde | 3.8 ± 0.4 abcd |
| F5 | 15.9 ± 1.6 b | 9.8 ± 4.3 bc | 6 ± 4.2 bc | 6.3 ± 3.9 cde | 3.8 ± 0.5 abcd |
| F6 | 16.9 ± 2.1 ab | 7.5 ± 5.1 bc | 5.9 ± 5.4 bc | 6.1 ± 4.9 cde | 3.4 ± 0.6 cd |
| F7 | 16.8 ± 1.8 ab | 5.4 ± 4.3 c | 5.6 ± 3.7 c | 5.0 ± 3.6 de | 3.7 ± 0.7 abcd |
| F8 | 17.3 ± 2.2 ab | 7.7 ± 7.4 bc | 5.7 ± 2.9 c | 6.1 ± 4.6 cde | 3.6 ± 0.6 bcd |
| F9 | 16.8 ± 1.7 ab | 6.0 ± 3.6 c | 6.1 ± 3.1 bc | 5.7 ± 2.8 cde | 3.9 ± 0.6 abc |
| F10 | 18.0 ± 2.0 a | 9.3 ± 7.5 bc | 7.6 ± 4 abc | 8.0 ± 5.0 bcde | 3.8 ± 0.8 abcd |
| F11 | 17.2 ± 1.3 ab | 9.3 ± 6.5 bc | 10.7 ± 5.9 ab | 8.6 ± 4.9 bcde | 4.1 ± 0.8 a |
| F12 | 15.9 ± 0.5 ab | 14.5 ± 6.0 a | 11.3 ± 3.7 a | 12.9 ± 3.3 a | 4 ± 0.8 ab |
| F13 | 16.2 ± 2.3 ab | 8.7 ± 5.0 bc | 7.4 ± 4.3 abc | 8.0 ± 4.1 bcde | 3.5 ± 0.7 bcd |
| F14 | 17.0 ± 2.1 ab | 7.0 ± 3.2 bc | 6.9 ± 4.9 abc | 6.9 ± 3.3 cde | 3.9 ± 0.6 abcd |
| F15 | 17.2 ± 2.3 ab | 9.7 ± 7.1 bc | 5.9 ± 2.7 bc | 7.8 ± 4.5 bcde | 3.6 ± 0.7 bcd |
| F16 | 17.2 ± 2.3 ab | 6.8 ± 3.6 bc | 5.4 ± 3.5 c | 6.1 ± 3.3 cde | 3.6 ± 0.7 bcd |
| F17 | 16.9 ± 1.4 ab | 8.8 ± 9.2 bc | 9.2 ± 6.7 abc | 7.5 ± 7.3 bcde | 3.5 ± 0.5 bcd |
| F18 | 16.9 ± 2.1 ab | 9.1 ± 6.3 bc | 10 ± 7.4 abc | 9.3 ± 6.4 bc | 3.6 ± 0.7 abcd |
| F19 | 16.5 ± 1.5 ab | 6.8 ± 4.3 bc | 8.8 ± 7.5 abc | 7.9 ± 5.6 bcde | 3.5 ± 0.7 bcd |
| F20 | 16.5 ± 1.6 ab | 7.9 ± 5.4 bc | 6 ± 2.7 bc | 6.7 ± 4.1 cde | 3.5 ± 1 bcd |
| F21 | 16.0 ± 2.7 ab | 7.3 ± 5.5 bc | 6.3 ± 3 bc | 4.5 ± 3.9 e | 3.8 ± 0.9 abcd |
| F22 | 17.3 ± 2.3 ab | 6.3 ± 4.5 bc | 5.4 ± 5.7 c | 4.7 ± 4.3 de | 3.7 ± 0.8 abcd |
| F23 | 16.9 ± 1.8 ab | 6.2 ± 4.9 bc | 7.1 ± 3 abc | 5.9 ± 3.6 cde | 3.4 ± 0.5 cd |
| F24 | 17.2 ± 1.9 ab | 8.6 ± 4.9 bc | 7 ± 2.4 abc | 7.4 ± 3.5 bcde | 3.6 ± 0.8 bcd |
| F25 | 17.2 ± 1.8 ab | 8.1 ± 7.0 bc | 6.1 ± 3.5 bc | 6.7 ± 5.1 cde | 3.8 ± 0.4 abcd |
| F26 | 17.3 ± 2.3 ab | 7.6 ± 5 bc | 5.6 ± 2 c | 6.6 ± 3.3 cde | 3.5 ± 0.5 bcd |
| F27 | 16.3 ± 1.7 ab | 6.6 ± 4.9 bc | 7.8 ± 3.9 abc | 6.1 ± 4.3 cde | 3.6 ± 0.7 bcd |
| F28 | 17.6 ± 2.6 ab | 10.6 ± 8.2 abc | 9.3 ± 6 abc | 9.5 ± 6.7 abc | 3.5 ± 0.9 bcd |
| F29 | 16.1 ± 1.4 ab | 8.3 ± 5.0 bc | 5.3 ± 2.9 c | 6.5 ± 3.4 cde | 3.5 ± 0.5 bcd |
| F30 | 17.0 ± 1.8 ab | 5.5 ± 2.7 c | 5.2 ± 3.4 c | 4.6 ± 2.9 e | 3.8 ± 0.6 abcd |
| F31 | 15.9 ± 1.1 b | 11.4 ± 9.9 ab | 10.6 ± 7.9 ab | 11 ± 7.7 ab | 3.7 ± 0.8 abcd |
| F32 | 16.8 ± 2.6 ab | 7.4 ± 4.4 bc | 9.3 ± 4.1 abc | 7.9 ± 3.3 bcde | 3.6 ± 0.6 bcd |
| F33 | 16.9 ± 1.7 ab | 8 ± 4.6 bc | 8.5 ± 6.3 abc | 6.8 ± 5.3 cde | 3.5 ± 0.7 bcd |
| F34 | 16.6 ± 1.4 ab | 8.4 ± 5.5 bc | 8.3 ± 5.8 abc | 8.8 ± 5.4 bcd | 3.3 ± 0.5 d |
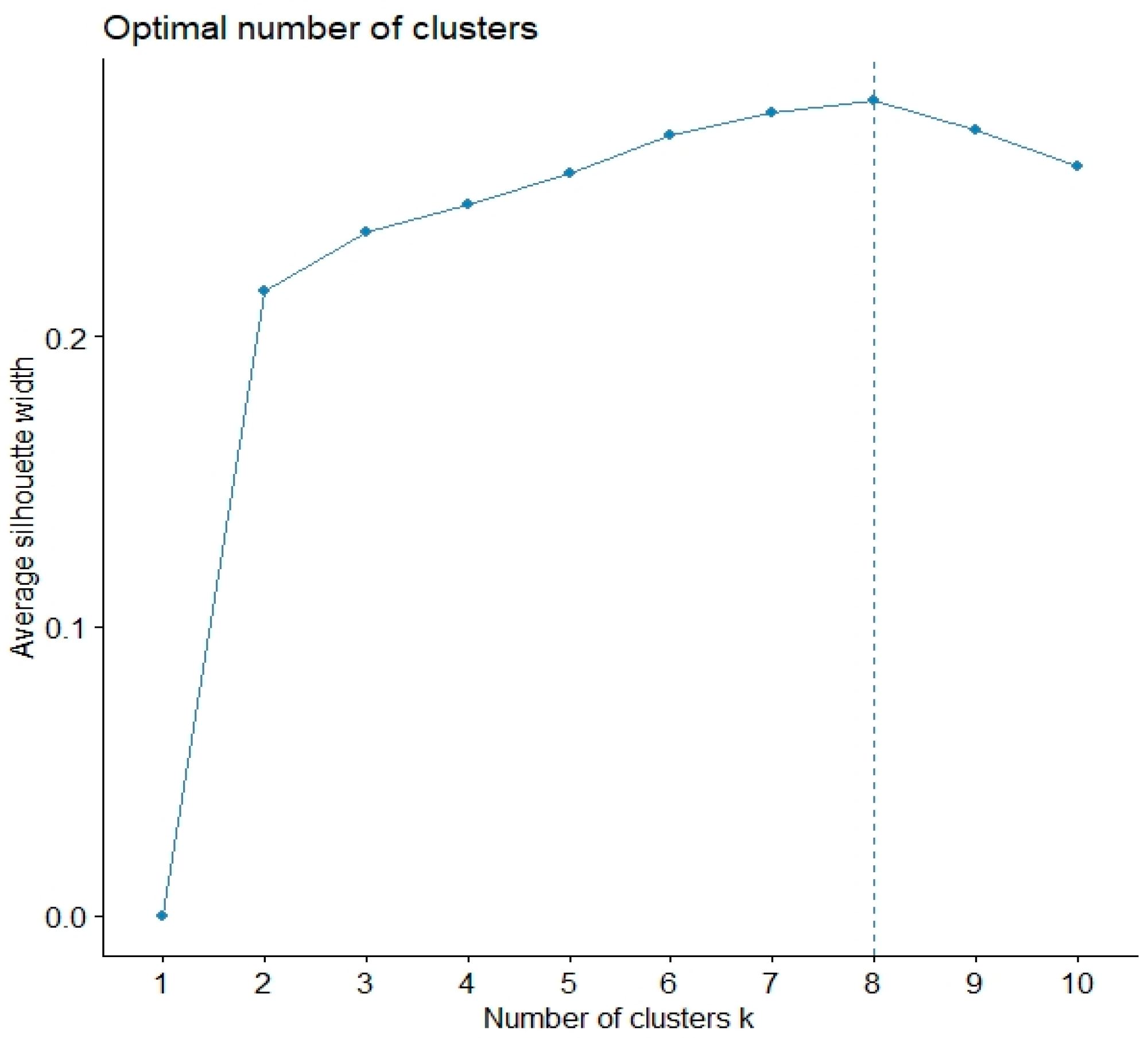
| Cluster | Mean ± sd of Trait Evaluation | Family Membership | |||
|---|---|---|---|---|---|
| DBH (cm) | WOP (g) | EOP (g) | PR score | ||
| C1 | 14.52 ± 0.68 b | 7.42 ± 0.90 cd | 8.32 ± 0.75 b | 3.68 ± 0.08 a | f11, f19, f10, f1, f32 |
| C2 | 12.45 ± 0.78 c | 11.6 ± 0.28 a | 10.3 ± 0.42 a | 3.40 ± 0.00 cd | f12, f31 |
| C3 | 14.30 ± 0.56 b | 8.37 ± 0.79 bc | 6.93 ± 1.00 c | 3.44 ± 0.09 bc | f20, f26, f24. f4, f17, f13, f34, f15, f3 |
| C4 | 12.45 ± 0.49 c | 7.40 ± 0.57 cd | 6.45 ± 0.64 cd | 3.60 ± 0.14 a | f14, f5 |
| C5 | 14.06 ± 0.38 b | 6.50 ± 1.23 de | 5.5 ± 0.49 de | 3.70 ± 0.07 a | f16, f25, f29, f21, f9 |
| C6 | 15.57 ± 0.31 a | 9.20 ± 0.95 b | 9.13 ± 0.68 ab | 3.57 ± 0.06 a | f28, f18, f2 |
| C7 | 15.50 ± 0.55 a | 5.66 ± 1.15 e | 5.00 ± 0.51 e | 3.64 ± 0.09 a | f22, f30, f7, f6, f8 |
| C8 | 14.37 ± 0.40 b | 5.70 ± 0.17 e | 6.83 ± 0.91 c | 3.30 ± 0.00 d | f23, f27, f33 |
| p-value | <0.001 ** | <0.001 ** | <0.001 ** | <0.001 ** | |
| Trait | Phenotypic Correlation | ||||
|---|---|---|---|---|---|
| DBH | WOP | EOP | AOP | PR | |
| DBH | - | - | - | ||
| WOP | 0.938 ** | - | - | ||
| EOP | 0.935 ** | 0.932 ** | - | ||
| AOP | 0.953 ** | 0.964 ** | 0.981 ** | ||
| PR | 0.992 ** | 0.926 ** | 0.927 ** | 0.942 ** | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Purwanto; Baskorowati, L.; Sumantoro, P.; Hendrati, R.L.; Susanto, M.; Mashudi; Setiadi, D.; Nurtjahjaningsih, I.L.G.; Pudjiono, S.; Kurniawan, A.; et al. Evaluation of Aphid Resistance and Oleoresin Production in Indigenous Tropical Pine (Pinus merkusii Jungh. and de Vriese). Forests 2022, 13, 977. https://doi.org/10.3390/f13070977
Purwanto, Baskorowati L, Sumantoro P, Hendrati RL, Susanto M, Mashudi, Setiadi D, Nurtjahjaningsih ILG, Pudjiono S, Kurniawan A, et al. Evaluation of Aphid Resistance and Oleoresin Production in Indigenous Tropical Pine (Pinus merkusii Jungh. and de Vriese). Forests. 2022; 13(7):977. https://doi.org/10.3390/f13070977
Chicago/Turabian StylePurwanto, Liliana Baskorowati, Pujo Sumantoro, Rina Laksmi Hendrati, Mudji Susanto, Mashudi, Dedi Setiadi, I.L.G. Nurtjahjaningsih, Sugeng Pudjiono, Agus Kurniawan, and et al. 2022. "Evaluation of Aphid Resistance and Oleoresin Production in Indigenous Tropical Pine (Pinus merkusii Jungh. and de Vriese)" Forests 13, no. 7: 977. https://doi.org/10.3390/f13070977