
Arbuscular Mycorrhizal Fungi Promote Gleditsia sinensis Lam. Root Growth under Salt Stress by Regulating Nutrient Uptake and Physiology

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Plant Seeds, AMF Inoculant, and Soil
2.3. Inoculation and Salt Stress Treatment
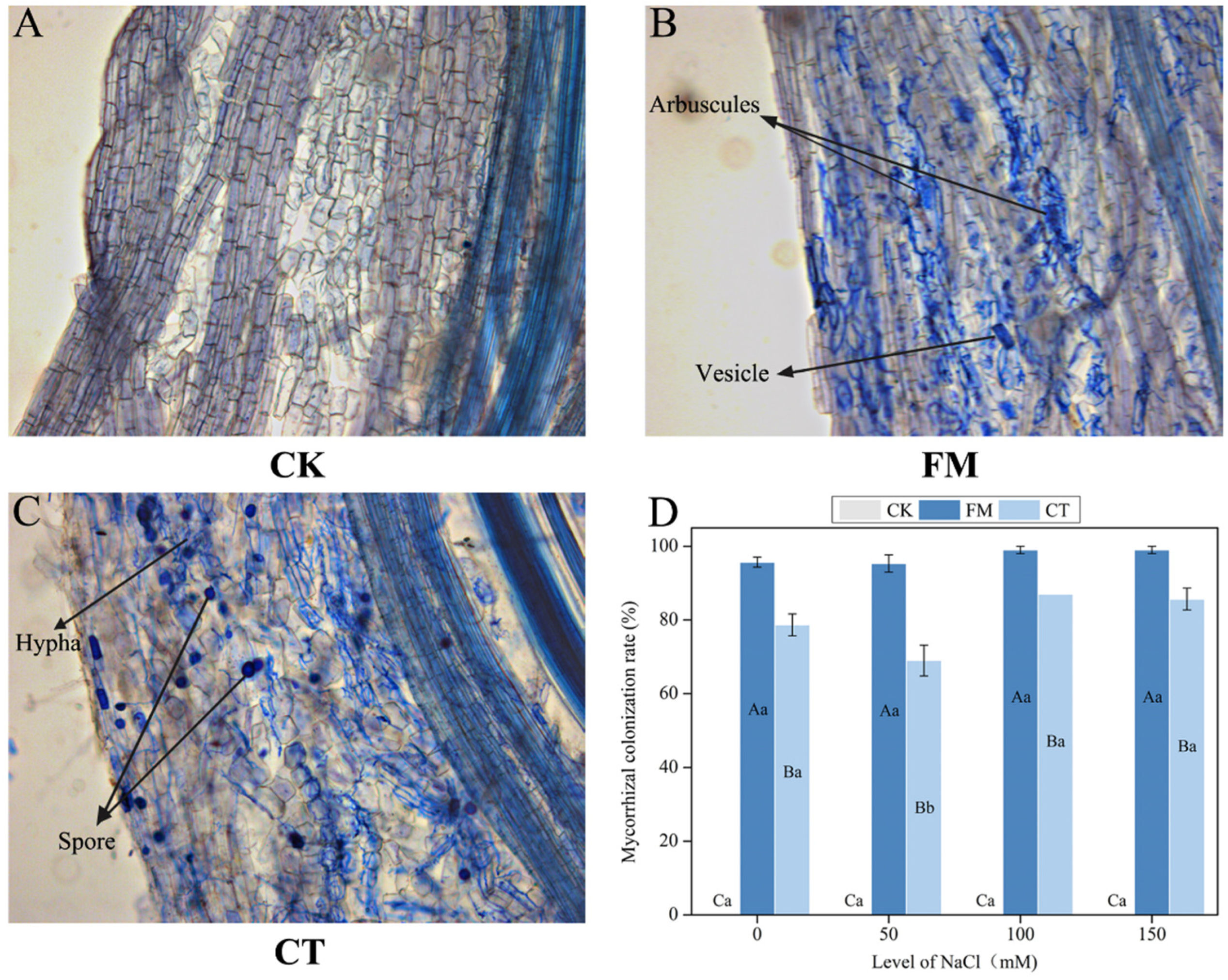
2.4. Determination of Mycorrhizal Colonization Status
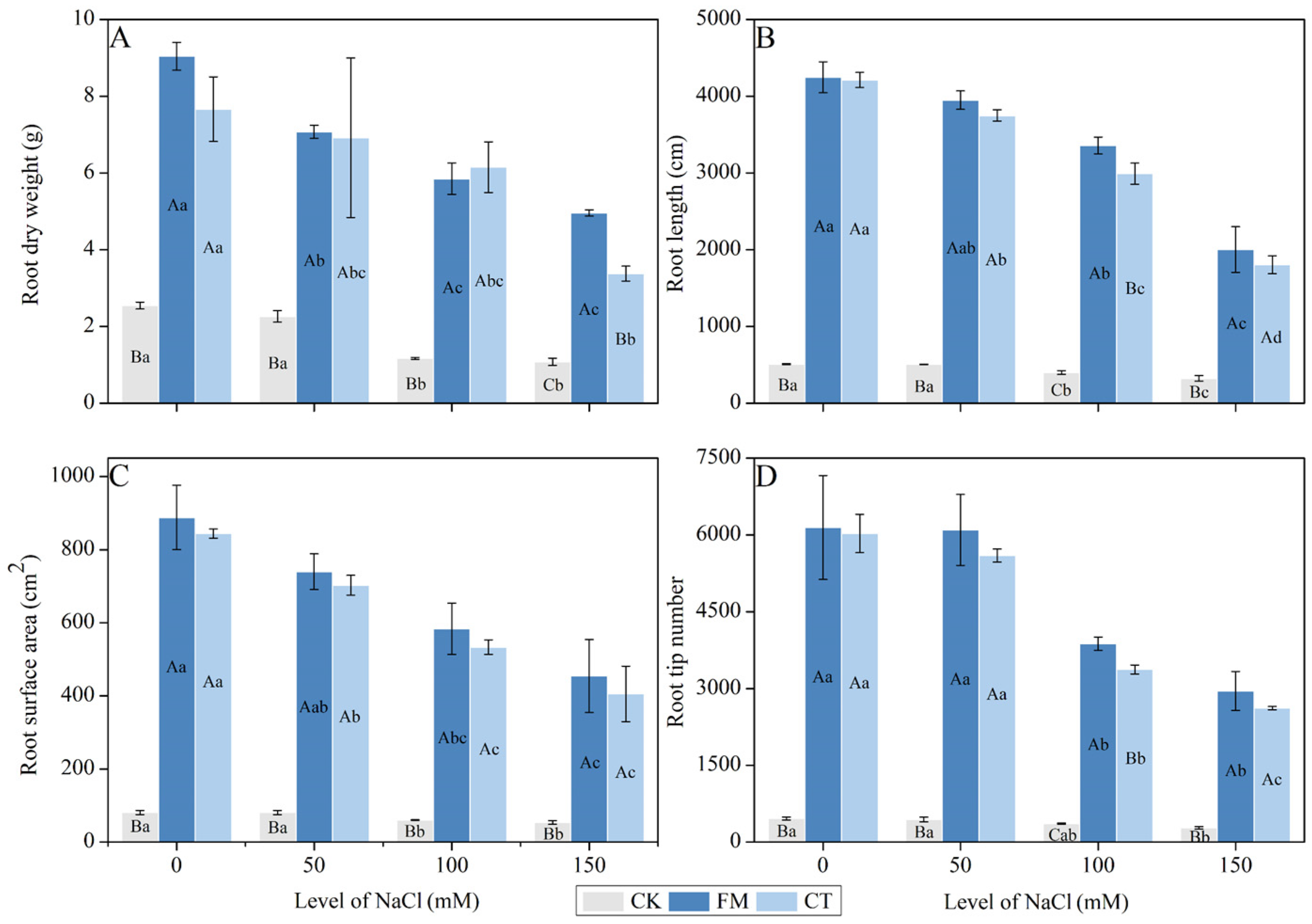
2.5. Determination of Root Dry Weight and Morphology
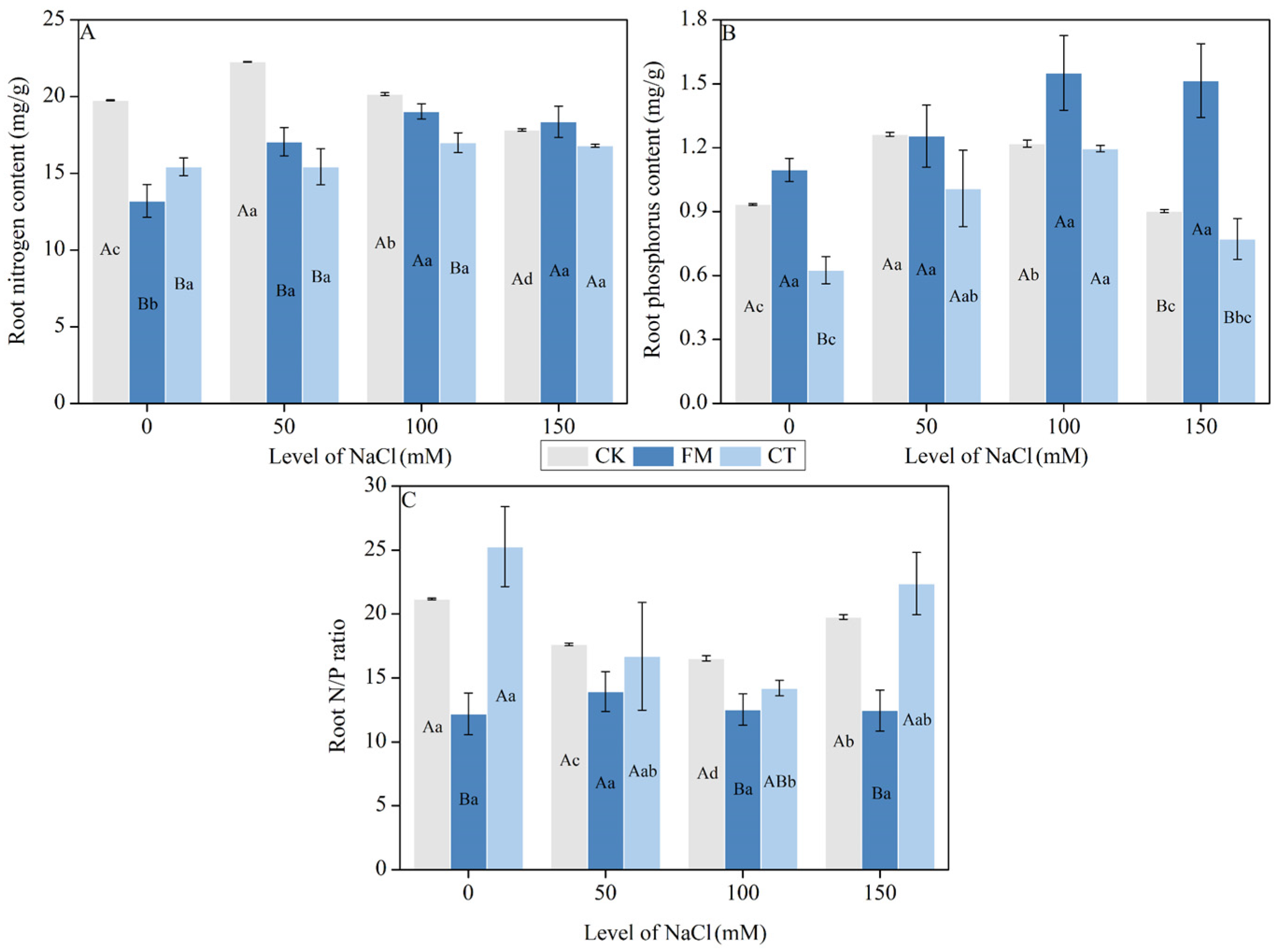
2.6. Determination of Root Nitrogen and Phosphorus
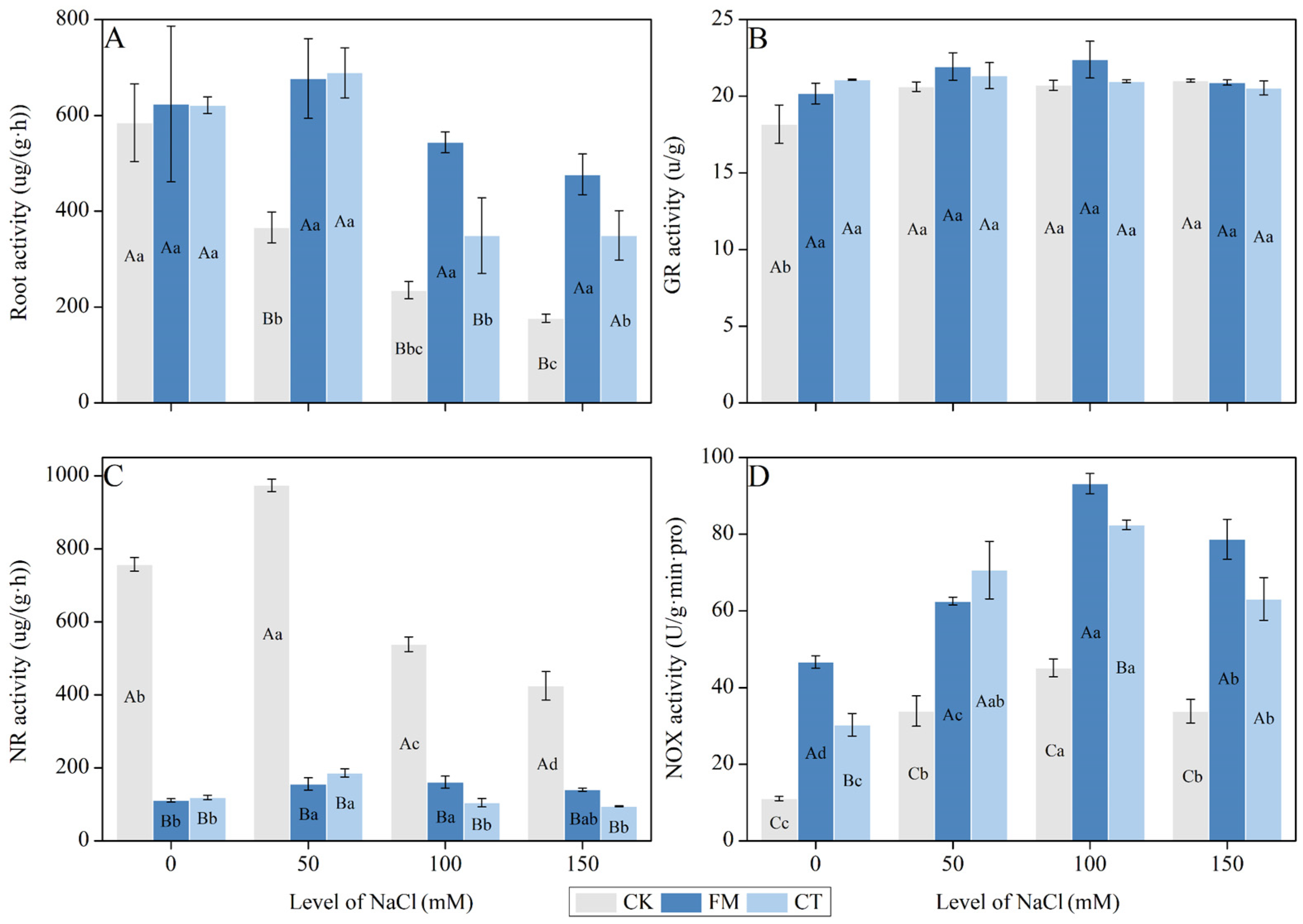
2.7. Determination of Root Enzyme Activities
2.8. Determination of Soil Nitrogen and Phosphorus
2.9. Statistical Analyses
3. Results
3.1. Mycorrhizal Colonization Status
3.2. Root Dry Weight and Morphology
3.3. Root Nitrogen and Phosphorus Content
3.4. Root Physiology
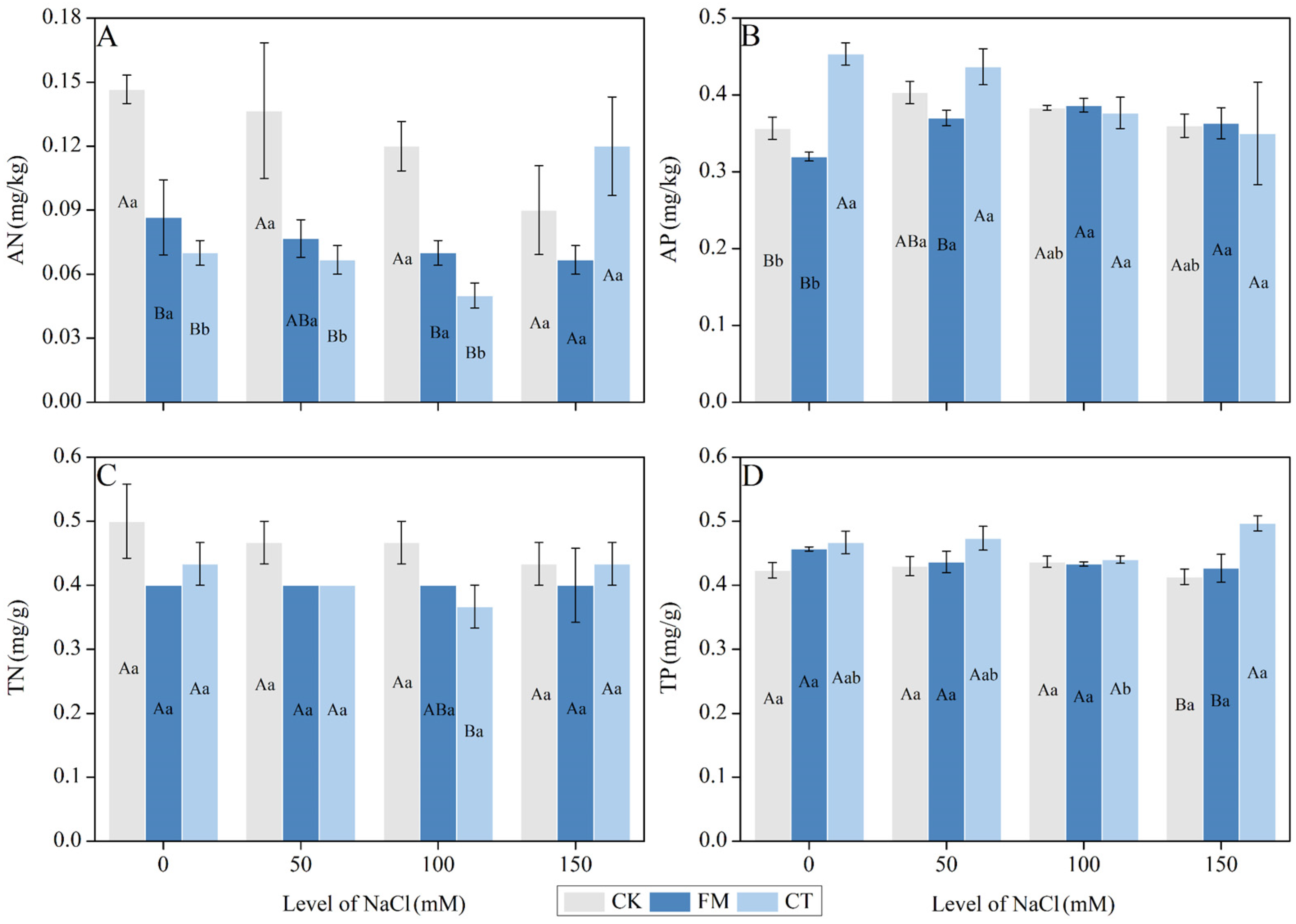
3.5. Soil Nitrogen and Phosphorus Content
3.6. Two-Way Analysis of Variance Results
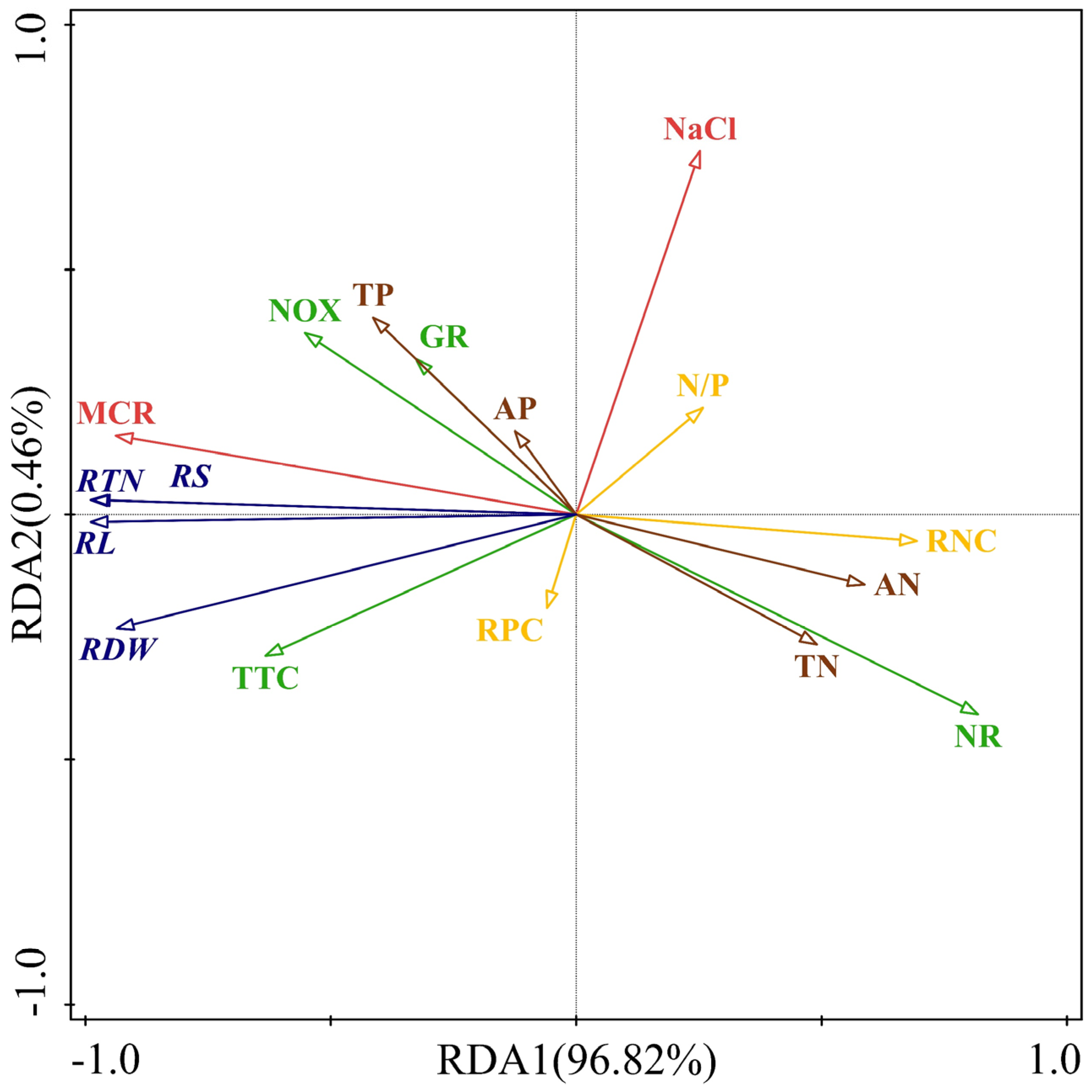
3.7. Redundant Analysis Results
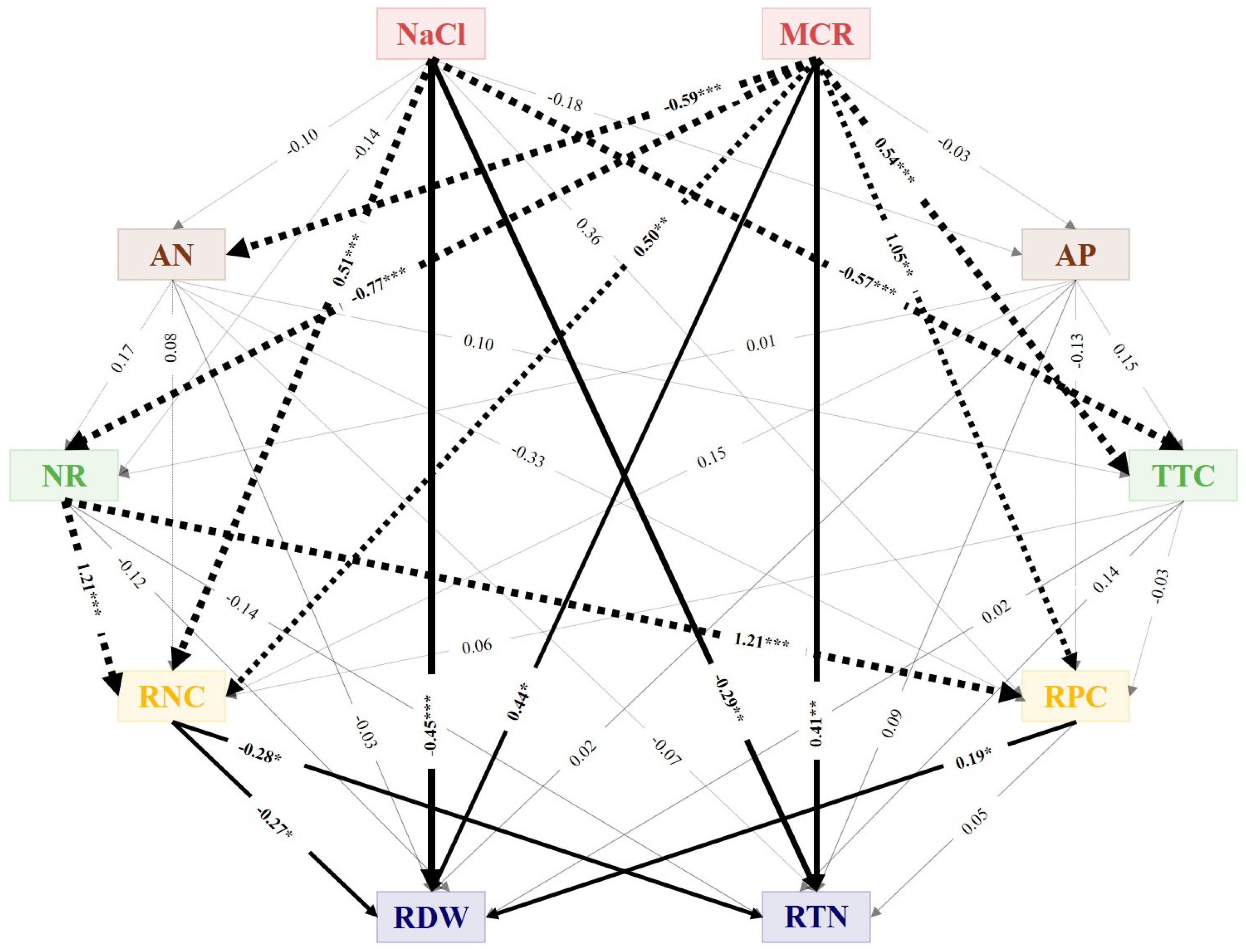
3.8. Structural Equation Model Results
4. Discussion
4.1. Responses of Root Growth to AMF Inoculation under Salt Stress
4.2. Effects of Nutrient Uptake on Root Growth under Salt Stresss
4.3. Effects of Physiology on Root Growth under Salt Stresss
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Mukhopadhyay, R.; Sarkar, B.; Jat, H.S.; Sharma, P.C.; Bolan, N.S. Soil salinity under climate change: Challenges for sustainable agriculture and food security. J. Environ. Manag. 2021, 280, 111736. [Google Scholar] [CrossRef] [PubMed]
- Kopittke, P.M.; Menzies, N.W.; Wang, P.; McKenna, B.A.; Lombi, E. Soil and the intensification of agriculture for global food security. Environ. Int. 2019, 132, 105078. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Lozano, J.M.; Porcel, R.; Azcon, C.; Aroca, R. Regulation by arbuscular mycorrhizae of the integrated physiological response to salinity in plants: New challenges in physiological and molecular studies. J. Exp. Bot. 2012, 63, 4033–4044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Zhai, L.; Ma, J.; Zhang, J.; Wang, G.G.; Liu, X.; Zhang, S.; Song, J.; Wu, Y. Comparative physiological mechanisms of arbuscular mycorrhizal fungi in mitigating salt-induced adverse effects on leaves and roots of Zelkova serrata. Mycorrhiza 2020, 30, 341–355. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Fu, Z.; Ren, Q.; Zhu, L.; Lin, J.; Zhang, J.; Cheng, X.; Ma, J.; Yue, J. Effects of Arbuscular Mycorrhizal Fungi on Growth, Photosynthesis, and Nutrient Uptake of Zelkova serrata (Thunb.) Makino Seedlings under Salt Stress. Forests 2019, 10, 186. [Google Scholar] [CrossRef] [Green Version]
- Bardgett, R.D.; Mommer, L.; De Vries, F.T. Going underground: Root traits as drivers of ecosystem processes. Trends Ecol. Evol. 2014, 29, 692–699. [Google Scholar] [CrossRef]
- Saleem, M.; Law, A.D.; Sahib, M.R.; Pervaiz, Z.H.; Zhang, Q. Impact of root system architecture on rhizosphere and root microbiome. Rhizosphere 2018, 6, 47–51. [Google Scholar] [CrossRef]
- Simpson, C.R.; Gonzales, J.; Enciso, J.; Nelson, S.D.; Sétamou, M. Root distribution and seasonal fluctuations under different grove floor management systems in citrus. Sci. Hortic. 2020, 272, 109364. [Google Scholar] [CrossRef]
- Krishnamoorthy, R.; Kim, K.; Kim, C.; Sa, T. Changes of arbuscular mycorrhizal traits and community structure with respect to soil salinity in a coastal reclamation land. Soil Biol. Biochem. 2014, 72, 1–10. [Google Scholar] [CrossRef]
- de Barros Silva Leite, M.C.; dos Santos Freire, M.B.G.; de Queiroz, J.V.J.; Maia, L.C.; Duda, G.P.; de Medeiros, E.V. Mycorrhizal Atriplex nummularia promote revegetation and shifts in microbial properties in saline Brazilian soil. Appl. Soil Ecol. 2020, 153, 103574. [Google Scholar] [CrossRef]
- Kumar Arora, N.; Fatima, T.; Mishra, J.; Mishra, I.; Verma, S.; Verma, R.; Verma, M.; Bhattacharya, A.; Verma, P.; Mishra, P.; et al. Halo-tolerant plant growth promoting rhizobacteria for improving productivity and remediation of saline soils. J. Adv. Res. 2020, 26, 69–82. [Google Scholar] [CrossRef] [PubMed]
- Etesami, H.; Glick, B.R. Halotolerant plant growth–promoting bacteria: Prospects for alleviating salinity stress in plants. Environ. Exp. Bot. 2020, 178, 104124. [Google Scholar] [CrossRef]
- Goswami, M.; Deka, S. Plant growth-promoting rhizobacteria—Alleviators of abiotic stresses in soil: A review. Pedosphere 2020, 30, 40–61. [Google Scholar] [CrossRef]
- Metwally, R.A.; Abdelhameed, R.E. Synergistic effect of arbuscular mycorrhizal fungi on growth and physiology of salt-stressed Trigonella foenum-graecum plants. Biocatal. Agric. Biotechnol. 2018, 16, 538–544. [Google Scholar] [CrossRef]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis. Q. Rev. Biol. 2008, 3, 273–281. [Google Scholar] [CrossRef]
- Hamdia, M.A.; Shaddad, M. Salt tolerance of crop plants. J. Stress Physiol. Biochem. 2010, 6, 64–90. [Google Scholar]
- Klinsukon, C.; Lumyong, S.; Kuyper, T.W.; Boonlue, S. Colonization by arbuscular mycorrhizal fungi improves salinity tolerance of eucalyptus (Eucalyptus camaldulensis) seedlings. Sci. Rep. 2021, 11, 4362. [Google Scholar] [CrossRef]
- Kaya, C.; Ashraf, M.; Sonmez, O.; Aydemir, S.; Tuna, A.L.; Cullu, M.A. The influence of arbuscular mycorrhizal colonisation on key growth parameters and fruit yield of pepper plants grown at high salinity. Sci. Hortic. 2009, 121, 1–6. [Google Scholar] [CrossRef]
- Hashem, A.; Alqarawi, A.A.; Radhakrishnan, R.; Al-Arjani, A.F.; Aldehaish, H.A.; Egamberdieva, D.; Abd Allah, E.F. Arbuscular mycorrhizal fungi regulate the oxidative system, hormones and ionic equilibrium to trigger salt stress tolerance in Cucumis sativus L. Saudi J. Biol. Sci. 2018, 25, 1102–1114. [Google Scholar] [CrossRef]
- Parihar, M.; Rakshit, A.; Rana, K.; Prasad Meena, R.; Chandra Joshi, D. A consortium of arbuscular mycorrizal fungi improves nutrient uptake, biochemical response, nodulation and growth of the pea (Pisum sativum L.) under salt stress. Rhizosphere 2020, 15, 100235. [Google Scholar] [CrossRef]
- Qiu, Y.; Zhang, N.; Zhang, L.; Zhang, X.; Wu, A.; Huang, J.; Yu, S.; Wang, Y. Mediation of arbuscular mycorrhizal fungi on growth and biochemical parameters of Ligustrum vicaryi in response to salinity. Physiol. Mol. Plant Pathol. 2020, 112, 101522. [Google Scholar] [CrossRef]
- Zhang, J.P.; Tian, X.H.; Yang, Y.X.; Liu, Q.X.; Zhang, W.D. Gleditsia species: An ethnomedical, phytochemical and pharmacological review. J. Ethnopharmacol. 2016, 178, 155–171. [Google Scholar] [CrossRef] [PubMed]
- Mei, X.; Ju, L.; Fang, C.; Li, J.; Bi, S.; Chen, Y.; Zou, C.; Zhou, C.; Ma, L.; Miao, L. Promoting the hydrolysis and acidification of rice straw by adding Gleditsia sinensis pod powder. Ind. Crops Prod. 2017, 100, 35–40. [Google Scholar] [CrossRef]
- Wang, J.; Yuan, J.; Ren, Q.; Zhang, B.; Zhang, J.; Huang, R.; Wang, G.G. Arbuscular mycorrhizal fungi enhanced salt tolerance of Gleditsia sinensis by modulating antioxidant activity, ion balance and P/N ratio. Plant Growth Regul. 2022, 97, 33–49. [Google Scholar] [CrossRef]
- Yang, H.; Li, W.; Yu, X.; Zhang, X.; Zhang, Z.; Liu, Y.; Wang, W.; Tian, X. Insights into molecular structure, genome evolution and phylogenetic implication through mitochondrial genome sequence of Gleditsia sinensis. Sci. Rep. 2021, 11, 14850. [Google Scholar] [CrossRef]
- Wang, J.; Zhong, H.; Zhu, L.; Yuan, Y.; Xu, L.; Wang, G.G.; Zhai, L.; Yang, L.; Zhang, J. Arbuscular Mycorrhizal Fungi Effectively Enhances the Growth of Gleditsia sinensis Lam. Seedlings under Greenhouse Conditions. Forests 2019, 10, 567. [Google Scholar] [CrossRef] [Green Version]
- Phillips, J.M. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans. Br. Mycol. Soc. 1970, 55, 158–161. [Google Scholar] [CrossRef]
- Giovannetti, M.; Mosse, B. An evaluation of techniques for measuring vesicular arbuscular mycorrhizal infection in roots. New Phytol. 1980, 84, 489–500. [Google Scholar] [CrossRef]
- Zhou, T.; Wang, L.; Sun, X.; Wang, X.; Chen, Y.; Rengel, Z.; Liu, W.; Yang, W. Light intensity influence maize adaptation to low P stress by altering root morphology. Plant Soil 2019, 447, 183–197. [Google Scholar] [CrossRef]
- Debouba, M.; Maaroufi-Dghimi, H.; Suzuki, A.; Ghorbel, M.H.; Gouia, H. Changes in growth and activity of enzymes involved in nitrate reduction and ammonium assimilation in tomato seedlings in response to NaCl stress. Ann. Bot. 2007, 99, 1143–1151. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Pu, Y.; Yang, D.; Yin, X.; He, Z.; Yang, Y.; Yang, Y. Molecular cloning and characterization of the glutathione reductase gene from Stipa purpurea. Biochem. Biophys. Res. Commun. 2018, 495, 1851–1857. [Google Scholar] [CrossRef] [PubMed]
- Smiri, M.; Chaoui, A.; Rouhier, N.; Gelhaye, E.; Jacquot, J.-P.; El Ferjani, E. NAD pattern and NADH oxidase activity in pea (Pisum sativum L.) under cadmium toxicity. Physiol. Mol. Biol. Plants 2010, 16, 305–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, B.; Yang, H.; Song, W.; Liu, C.; Xu, J.; Zhao, W.; Zhou, Z. Effect of N fertilization rate on soil alkali-hydrolyzable N, subtending leaf N concentration, fiber yield, and quality of cotton. Crop J. 2016, 4, 323–330. [Google Scholar] [CrossRef] [Green Version]
- Lu, R. Analysis Methods of Soil Agricultural Chemistry; China Agricultural Science and Technology Publishing House: Beijing, China, 2000. [Google Scholar]
- Garg, N.; Pandey, R. Effectiveness of native and exotic arbuscular mycorrhizal fungi on nutrient uptake and ion homeostasis in salt-stressed Cajanus cajan L. (Millsp.) genotypes. Mycorrhiza 2015, 25, 165–180. [Google Scholar] [CrossRef] [PubMed]
- Arab Yarahmadi, M.; Shahsavani, S.; Akhyani, A.; Dorostkar, V. Pomegranate growth affected by arbuscular mycorrhizae, phosphorus fertilizer, and irrigation Water Salinity. Commun. Soil Sci. Plant Anal. 2018, 49, 478–488. [Google Scholar] [CrossRef]
- Estrada, B.; Aroca, R.; Maathuis, F.J.; Barea, J.M.; Ruiz-Lozano, J.M. Arbuscular mycorrhizal fungi native from a Mediterranean saline area enhance maize tolerance to salinity through improved ion homeostasis. Plant Cell Environ. 2013, 36, 1771–1782. [Google Scholar] [CrossRef]
- Sawers, R.J.; Svane, S.F.; Quan, C.; Gronlund, M.; Wozniak, B.; Gebreselassie, M.N.; Gonzalez-Munoz, E.; Chavez Montes, R.A.; Baxter, I.; Goudet, J.; et al. Phosphorus acquisition efficiency in arbuscular mycorrhizal maize is correlated with the abundance of root-external hyphae and the accumulation of transcripts encoding PHT1 phosphate transporters. New Phytol. 2017, 214, 632–643. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Bu, J.; Liang, C. Root Morphology and Growth Regulated by Mineral Nutrient Absorption in Rice Roots Exposed to Simulated Acid Rain. Water Air Soil Pollut. 2016, 227, 457. [Google Scholar] [CrossRef]
- Fan, L.; Dalpé, Y.; Fang, C.; Dubé, C.; Khanizadeh, S. Influence of arbuscular mycorrhizae on biomass and root morphology of selected strawberry cultivars under salt stress. Botany 2011, 89, 397–403. [Google Scholar] [CrossRef]
- Kaur, H.; Sirhindi, G.; Bhardwaj, R.; Alyemeni, M.N.; Siddique, K.H.M.; Ahmad, P. 28-homobrassinolide regulates antioxidant enzyme activities and gene expression in response to salt- and temperature-induced oxidative stress in Brassica juncea. Sci. Rep. 2018, 8, 8735. [Google Scholar] [CrossRef]
- Evelin, H.; Giri, B.; Kapoor, R. Contribution of Glomus intraradices inoculation to nutrient acquisition and mitigation of ionic imbalance in NaCl-stressed Trigonella foenum-graecum. Mycorrhiza 2012, 22, 203–217. [Google Scholar] [CrossRef] [PubMed]
- Gao, T.; Liu, X.; Shan, L.; Wu, Q.; Liu, Y.; Zhang, Z.; Ma, F.; Li, C. Dopamine and arbuscular mycorrhizal fungi act synergistically to promote apple growth under salt stress. Environ. Exp. Bot. 2020, 178, 104159. [Google Scholar] [CrossRef]
- Elser, J.J.; Acharya, K.; Kyle, M.; Cotner, J.; Makino, W.; Markow, T.; Watts, T.; Hobbie, S.; Fagan, W.; Schade, J.; et al. Growth rate-stoichiometry couplings in diverse biota. Ecol. Lett. 2003, 6, 936–943. [Google Scholar] [CrossRef] [Green Version]
- Matzek, V.; Vitousek, P.M. N:P stoichiometry and protein:RNA ratios in vascular plants: An evaluation of the growth-rate hypothesis. Ecol. Lett. 2009, 12, 765–771. [Google Scholar] [CrossRef]
- Evelin, H.; Kapoor, R.; Giri, B. Arbuscular mycorrhizal fungi in alleviation of salt stress: A review. Ann. Bot. 2009, 104, 1263–1280. [Google Scholar] [CrossRef] [Green Version]
- Gusewell, S. N:P ratios in terrestrial plants: Variation and functional significance. New Phytol. 2004, 164, 243–266. [Google Scholar] [CrossRef]
- Hidri, R.; Metoui-Ben Mahmoud, O.; Debez, A.; Abdelly, C.; Barea, J.-M.; Azcon, R. Modulation of C:N:P stoichiometry is involved in the effectiveness of a PGPR and AM fungus in increasing salt stress tolerance of Sulla carnosa Tunisian provenances. Appl. Soil Ecol. 2019, 143, 161–172. [Google Scholar] [CrossRef]
- Ait-El-Mokhtar, M.; Laouane, R.B.; Anli, M.; Boutasknit, A.; Wahbi, S.; Meddich, A. Use of mycorrhizal fungi in improving tolerance of the date palm (Phoenix dactylifera L.) seedlings to salt stress. Sci. Hortic. 2019, 253, 429–438. [Google Scholar] [CrossRef]
- Hadian-Deljou, M.; Esna-Ashari, M.; Mirzaie-asl, A. Alleviation of salt stress and expression of stress-responsive gene through the symbiosis of arbuscular mycorrhizal fungi with sour orange seedlings. Sci. Hortic. 2020, 268, 109373. [Google Scholar] [CrossRef]
- Leshem, Y.; Seri, L.; Levine, A. Induction of phosphatidylinositol 3-kinase-mediated endocytosis by salt stress leads to intracellular production of reactive oxygen species and salt tolerance. Plant J. 2007, 51, 185–197. [Google Scholar] [CrossRef]
- Foreman, J.; Demidchik, V.; Bothwell, J.; Mylona, P.; Miedema, H.; Torres, M.A.; Linstead, P.; Costa, S.; Brownlee, C.; Jones, J. Reactive oxygen species produced by NADPH oxidase regulate plant cell growth. Nature 2003, 422, 442–446. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhang, H.; Lou, X.; Tang, M. Mycorrhizal and non-mycorrhizal Medicago truncatula roots exhibit differentially regulated NADPH oxidase and antioxidant response under Pb stress. Environ. Exp. Bot. 2019, 164, 10–19. [Google Scholar] [CrossRef]
- Ben Rejeb, K.; Benzarti, M.; Debez, A.; Bailly, C.; Savouré, A.; Abdelly, C. NADPH oxidase-dependent H2O2 production is required for salt-induced antioxidant defense in Arabidopsis thaliana. J. Plant Physiol. 2015, 174, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Islam, E.; Yang, X.; Li, T.; Liu, D.; Jin, X.; Meng, F. Effect of Pb toxicity on root morphology, physiology and ultrastructure in the two ecotypes of Elsholtzia argyi. J. Hazard. Mater. 2007, 147, 806–816. [Google Scholar] [CrossRef] [PubMed]
- Begum, N.; Ahanger, M.A.; Zhang, L. AMF inoculation and phosphorus supplementation alleviates drought induced growth and photosynthetic decline in Nicotiana tabacum by up-regulating antioxidant metabolism and osmolyte accumulation. Environ. Exp. Bot. 2020, 176, 104088. [Google Scholar] [CrossRef]
- Ashraf, M.; Shahzad, S.M.; Imtiaz, M.; Rizwan, M.S. Salinity effects on nitrogen metabolism in plants—focusing on the activities of nitrogen metabolizing enzymes: A review. J. Plant Nutr. 2018, 41, 1065–1081. [Google Scholar] [CrossRef]
- Vázquez, M.; Barea, J.M.; Azcón, R. Influence of arbuscular mycorrhizae and a genetically modified strain of Sinorhizobium on growth, nitrate reductase activity and protein content in shoots and roots of Medicago sativa as affected by nitrogen concentrations. Soil Biol. Biochem. 2002, 34, 899–905. [Google Scholar] [CrossRef]
- Sveen, T.R.; Netherway, T.; Juhanson, J.; Oja, J.; Borgström, P.; Viketoft, M.; Strengbom, J.; Bommarco, R.; Clemmensen, K.; Hallin, S.; et al. Plant-microbe interactions in response to grassland herbivory and nitrogen eutrophication. Soil Biol. Biochem. 2021, 156, 108208. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | NaCl | AMF | NaCl × AMF |
|---|---|---|---|
| MCR | 0.000 *** | 0.000 *** | 0.002 ** |
| RDW | 0.000 *** | 0.000 *** | 0.296NS |
| RL | 0.000 *** | 0.000 *** | 0.000 *** |
| RS | 0.000 *** | 0.000 *** | 0.007 ** |
| RTN | 0.000 *** | 0.000 *** | 0.001 ** |
| RNC | 0.001 ** | 0.000 *** | 0.000 *** |
| RPC | 0.000 *** | 0.000 *** | 0.100NS |
| N/P | 0.015 * | 0.000 *** | 0.085NS |
| TTC | 0.000 *** | 0.000 *** | 0.210NS |
| GR | 0.035 * | 0.050NS | 0.244NS |
| NR | 0.000 *** | 0.000 *** | 0.000 *** |
| NOX | 0.000 *** | 0.000 *** | 0.015 * |
| AN | 0.409NS | 0.000 *** | 0.024 * |
| AP | 0.167NS | 0.047NS | 0.070NS |
| TN | 0.667NS | 0.018 * | 0.782NS |
| TP | 0.707NS | 0.000 *** | 0.112NS |
| G. sinensis | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| AN | AP | NR | TTC | RNC | RPC | RDW | RTN | ||
| NaCl | SDE | −0.10 | −0.18 | −0.14 | −0.57 | 0.51 | 0.36 | −0.45 | −0.29 |
| SIE | 0.00 | 0.00 | −0.02 | −0.04 | −0.26 | −0.12 | −0.02 | −0.13 | |
| STE | −0.10 | −0.18 | −0.16 | −0.61 | 0.25 | 0.24 | −0.47 | −0.42 | |
| MCR | SDE | −0.59 | −0.03 | −0.77 | 0.54 | 0.50 | 1.05 | 0.44 | 0.41 |
| SIE | 0.00 | 0.00 | −0.10 | −0.06 | −1.09 | −0.88 | 0.33 | 0.40 | |
| STE | −0.59 | −0.03 | −0.88 | 0.47 | −0.59 | 0.16 | 0.76 | 0.81 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, S.; Zhu, L.; Wang, J.; Liu, X.; Jia, Z.; Li, C.; Liu, J.; Zeng, J.; Zhang, J. Arbuscular Mycorrhizal Fungi Promote Gleditsia sinensis Lam. Root Growth under Salt Stress by Regulating Nutrient Uptake and Physiology. Forests 2022, 13, 688. https://doi.org/10.3390/f13050688
Ma S, Zhu L, Wang J, Liu X, Jia Z, Li C, Liu J, Zeng J, Zhang J. Arbuscular Mycorrhizal Fungi Promote Gleditsia sinensis Lam. Root Growth under Salt Stress by Regulating Nutrient Uptake and Physiology. Forests. 2022; 13(5):688. https://doi.org/10.3390/f13050688
Chicago/Turabian StyleMa, Shilin, Lingjun Zhu, Jinping Wang, Xin Liu, Zhaohui Jia, Chong Li, Jing Liu, Jingyi Zeng, and Jinchi Zhang. 2022. "Arbuscular Mycorrhizal Fungi Promote Gleditsia sinensis Lam. Root Growth under Salt Stress by Regulating Nutrient Uptake and Physiology" Forests 13, no. 5: 688. https://doi.org/10.3390/f13050688