
Could Pine Wood Nematode (Bursaphelenchus xylophilus) Cause Pine Wilt Disease or Even Establish inside Healthy Trees in Finland Now—Or Ever?


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
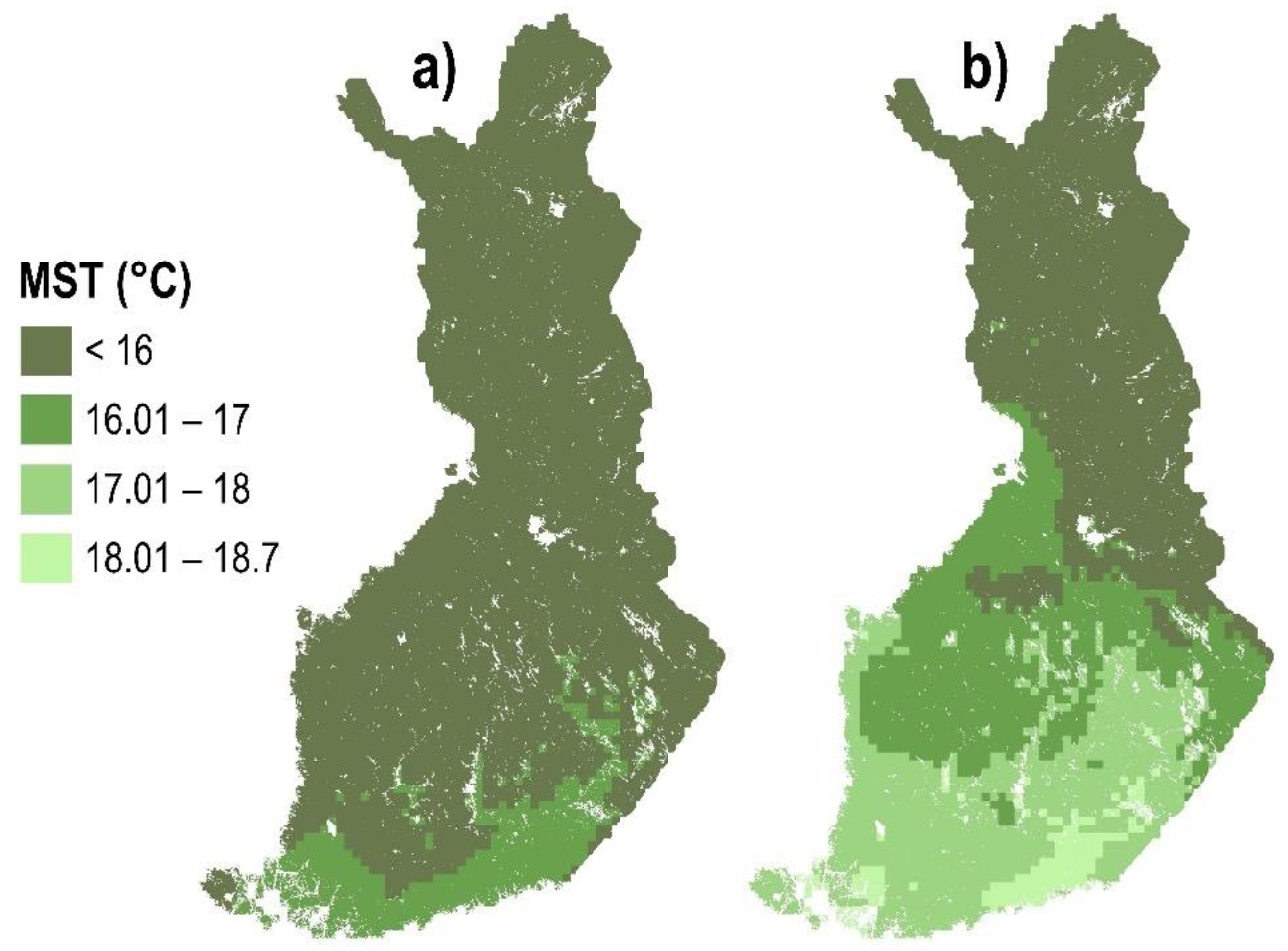
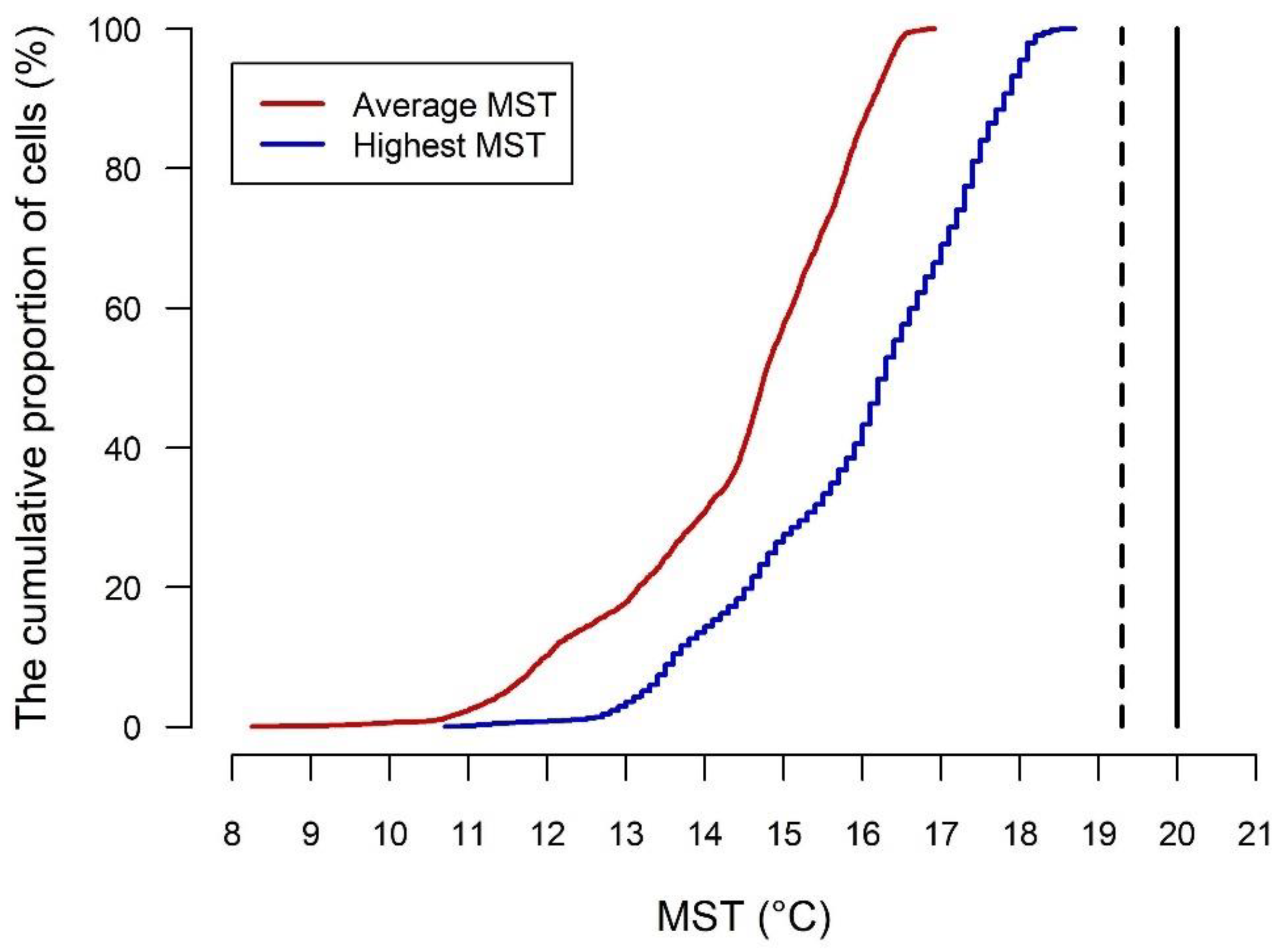
2.1. Method to Assess the Suitability of Climate for PWD in Healthy Trees
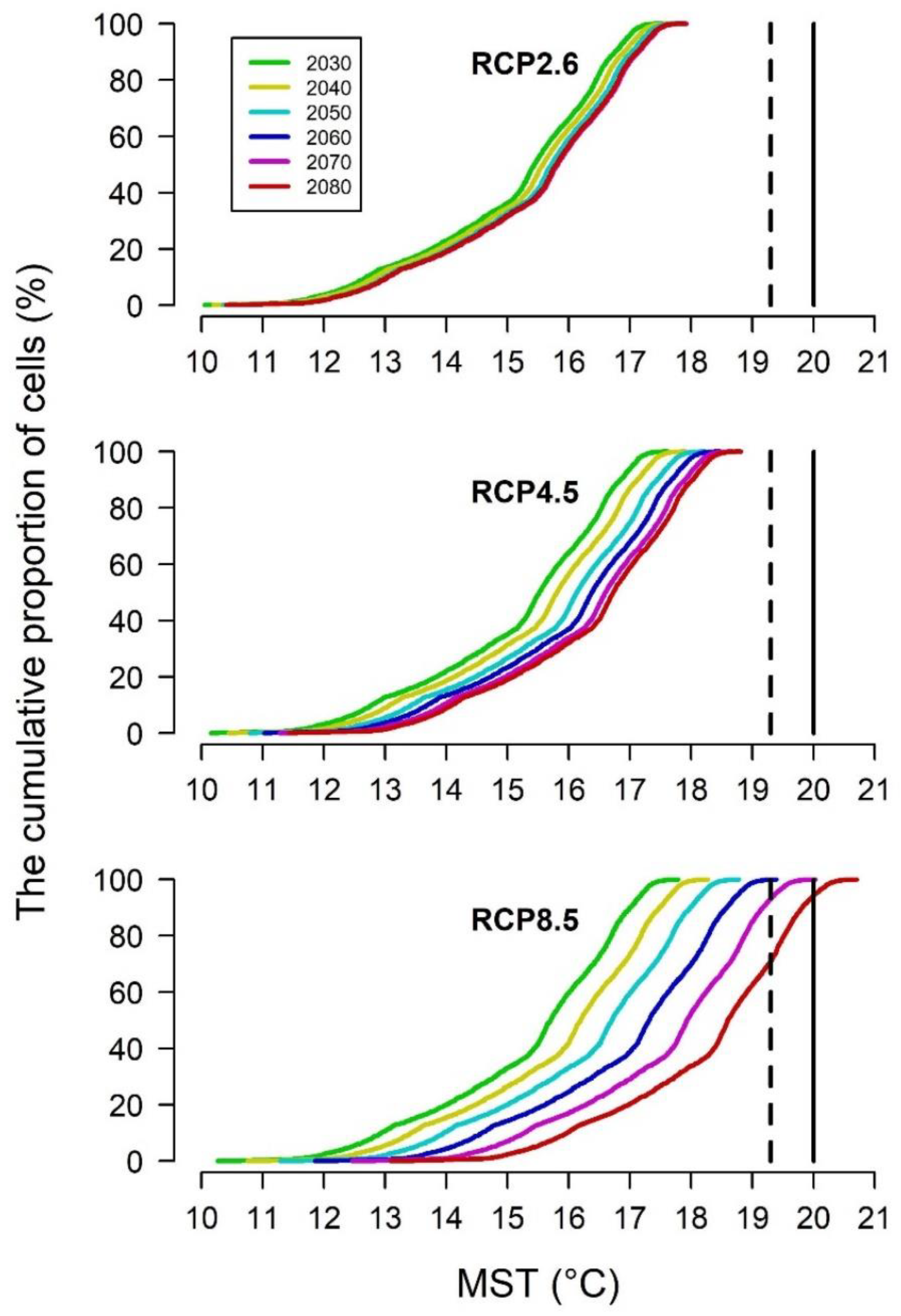
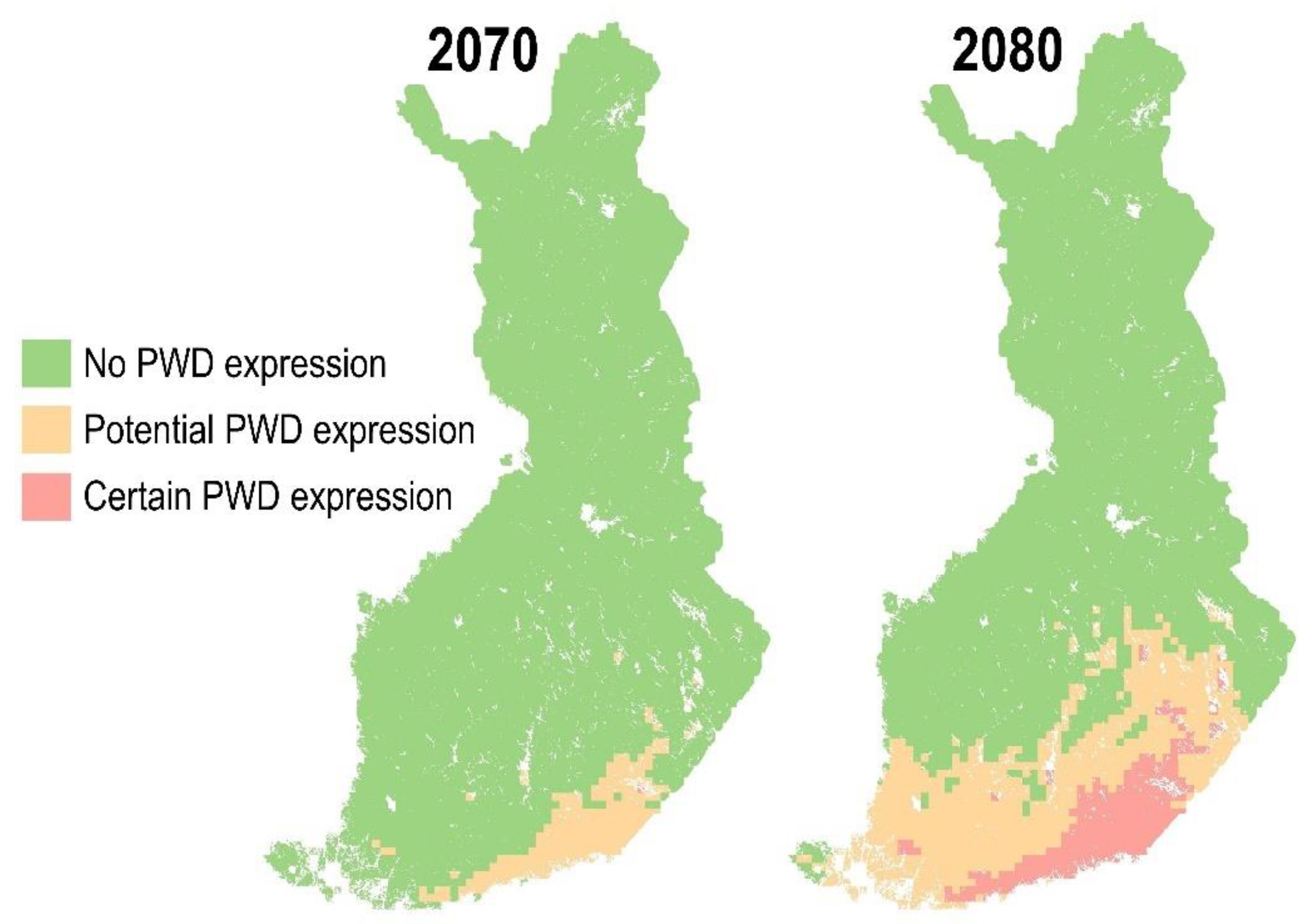
- MST 19.31 °C, threshold for potential PWD expression;
- MST 20 °C, threshold for certain PWD expression.
2.2. Method to Assess the Suitability of Climate for PWN Establishment inside Healthy Trees
2.2.1. Calculating Annual Growing Degree Days from Monthly Temperature Data
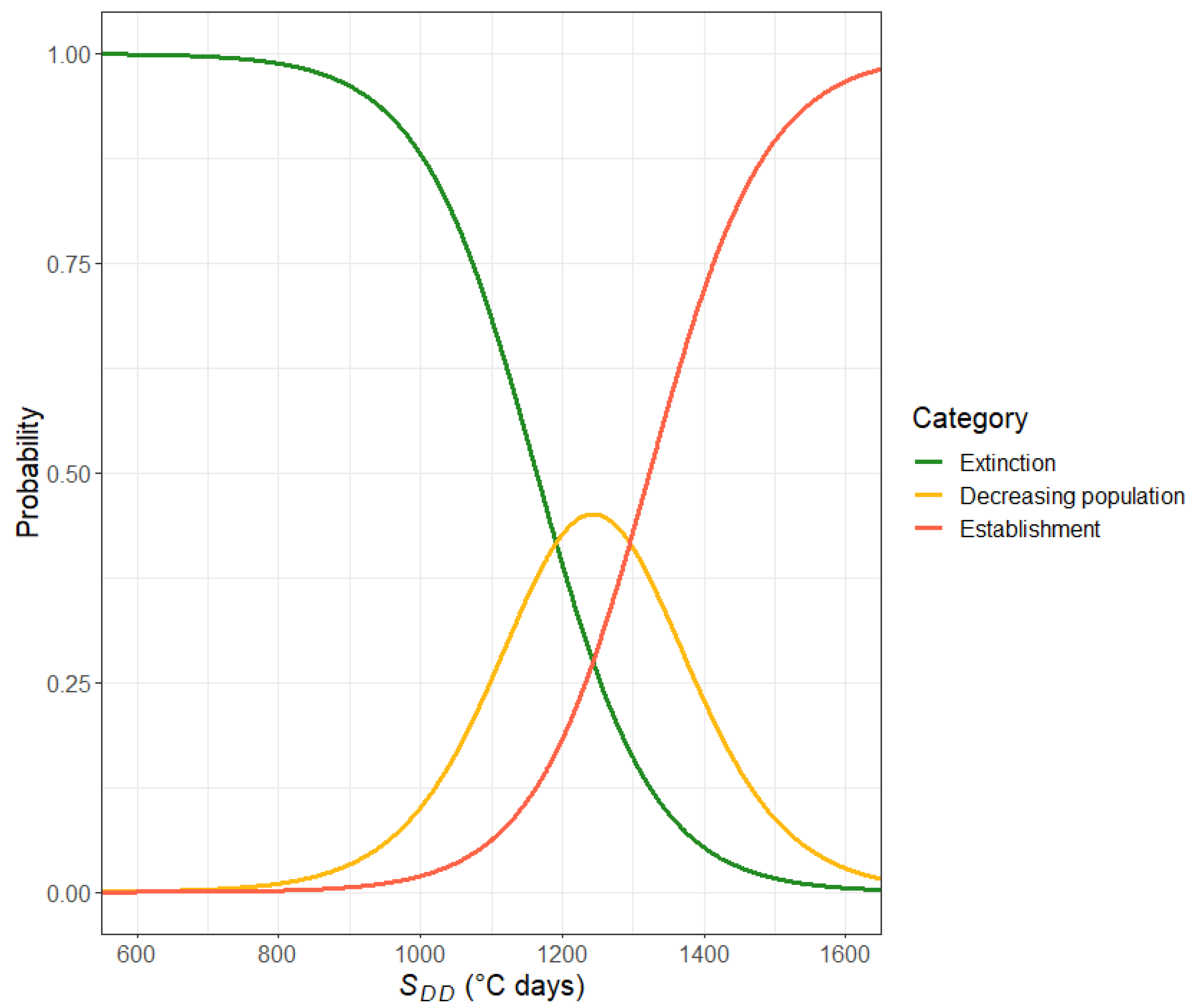
2.2.2. Relating Annual Growing Degree Days to the Probability of Extinction vs. Establishment
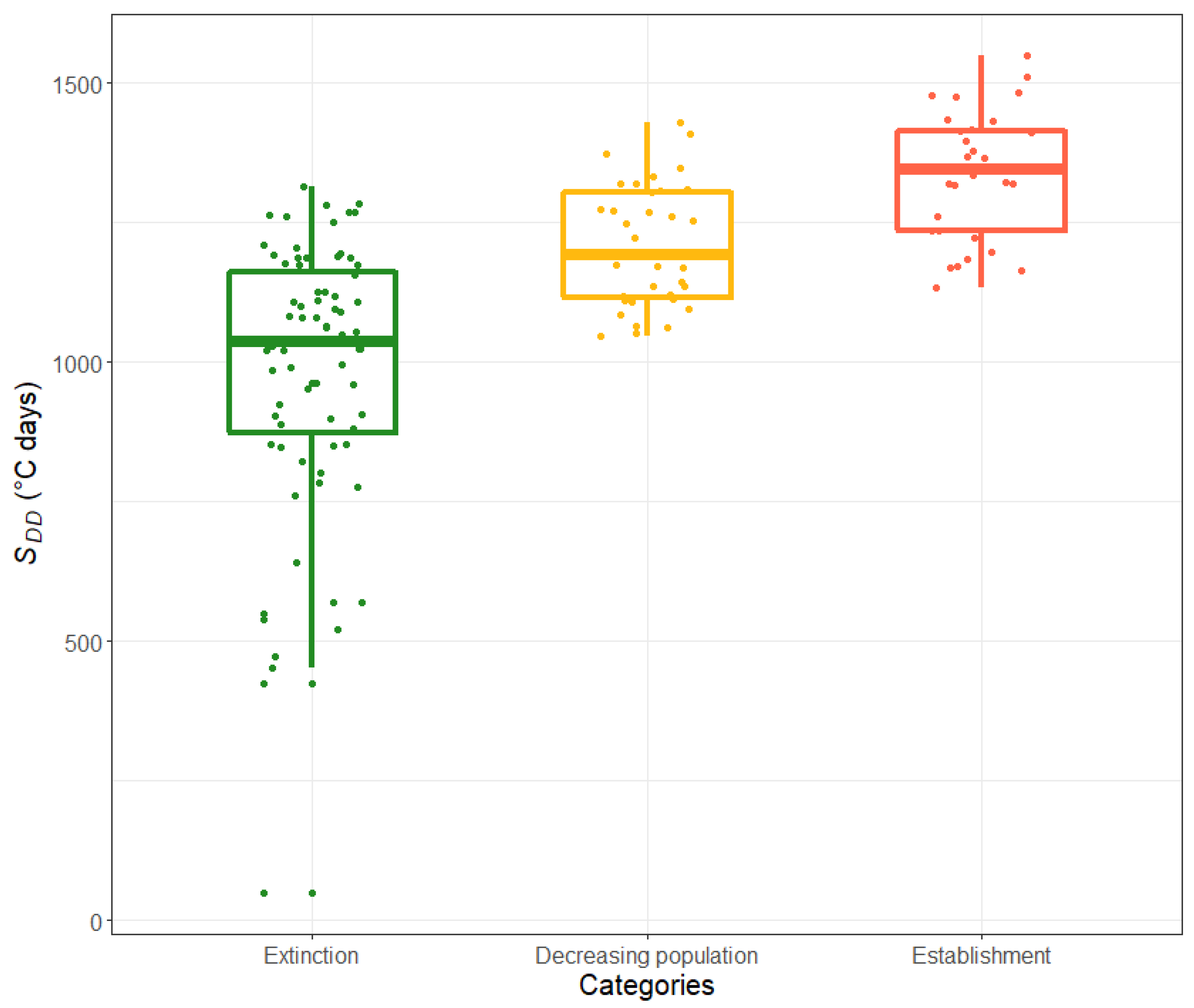
- Extinction. Following the infestation, the nematode numbers rapidly decrease. In most cases, the population collapses by the following summer, and in all cases, it completely disappears during the three years simulated.
- Decreasing population. Following the infestation, the nematode numbers decrease but do not become zero during the three years simulated. However, since the trend in nematode numbers is decreasing, in these conditions, the nematodes are likely to eventually disappear from the trees.
- Establishment. Following the infestation, the nematode numbers remain relatively stable throughout the three years simulated, suggesting that in such conditions, the PWN population can establish inside healthy trees.
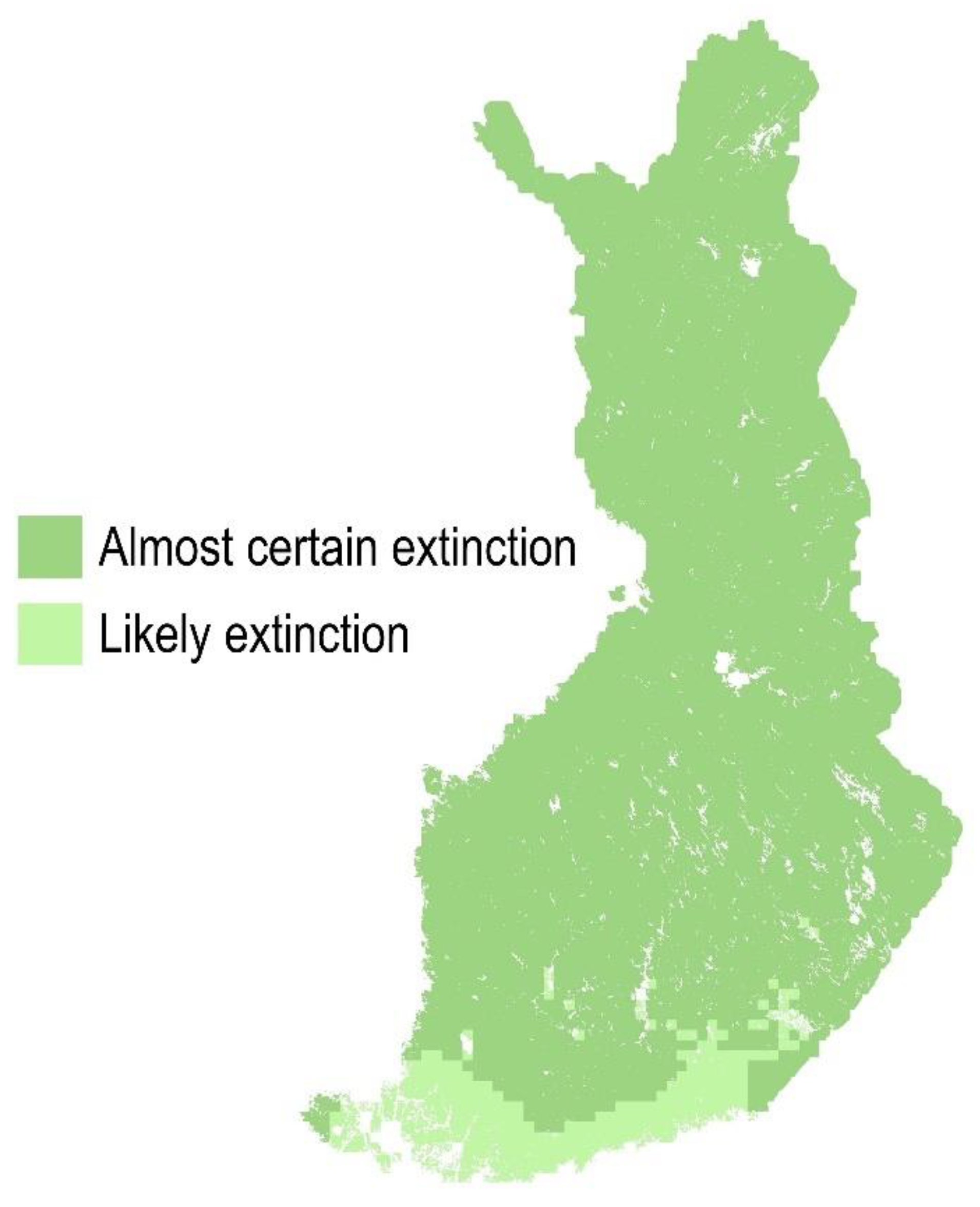
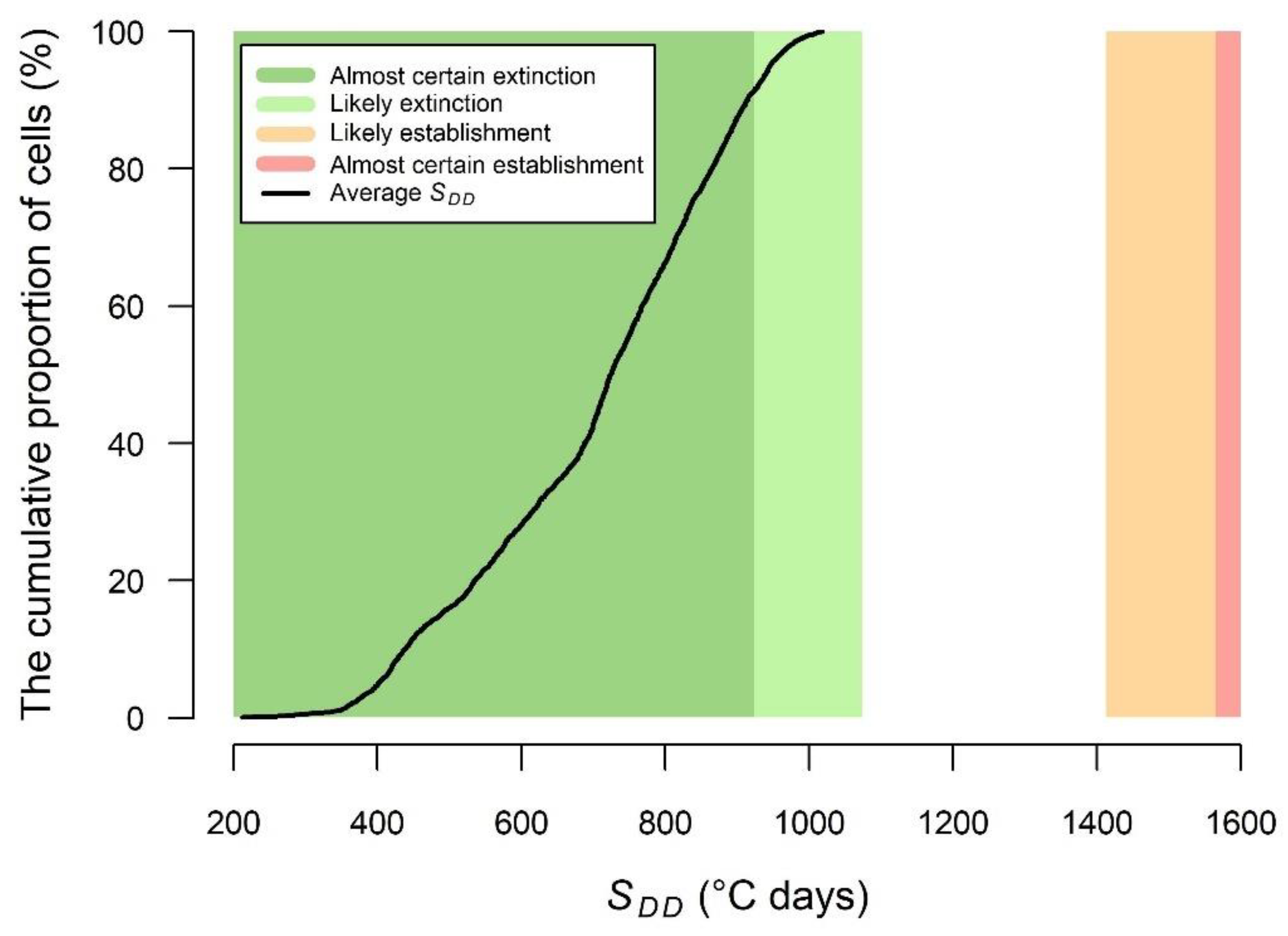
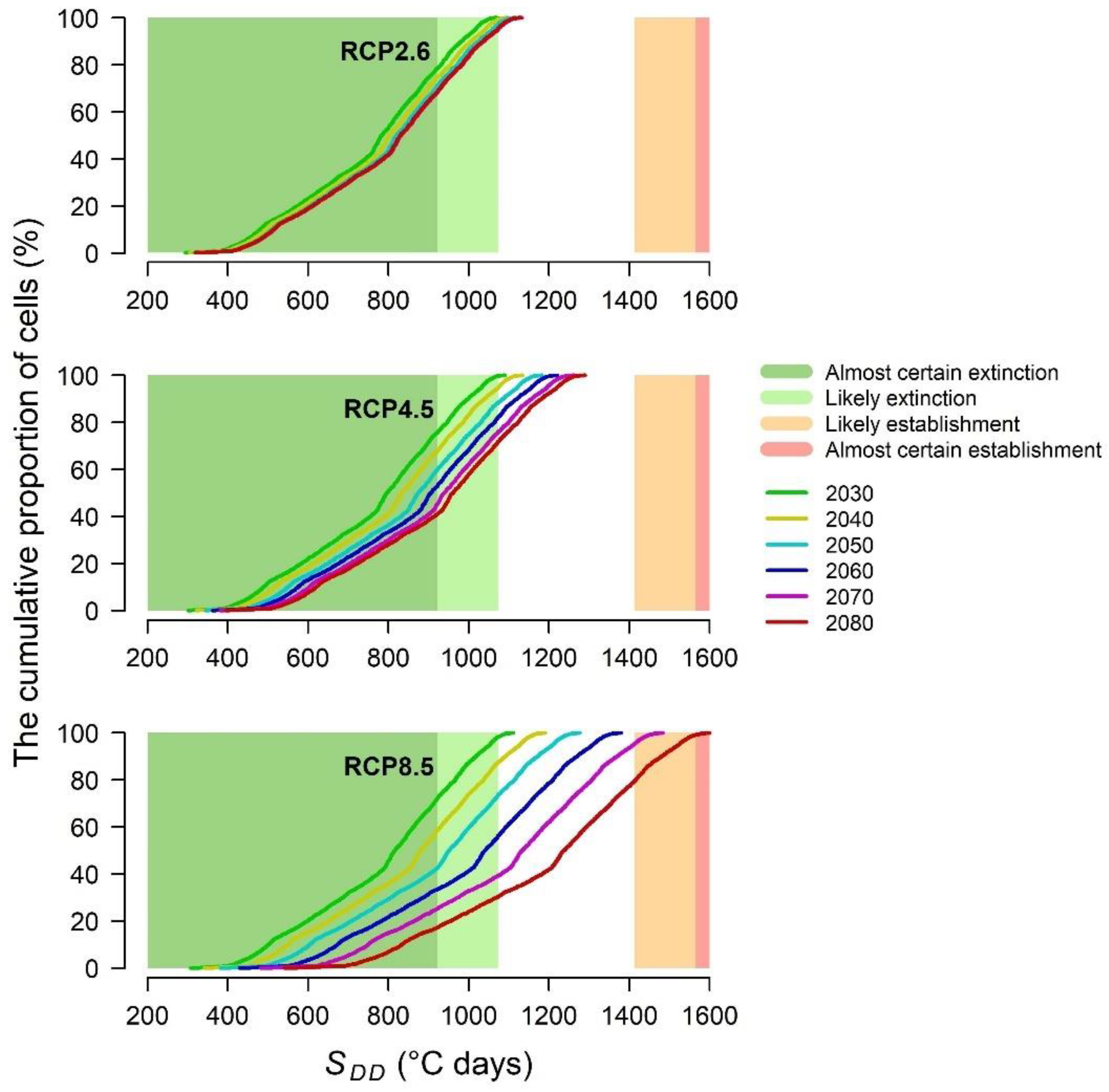
2.2.3. SDD Intervals for Estimating the Likelihood of PWN Extinction vs. Establishment inside Healthy Trees
- SDD < 923, almost certain extinction;
- 923 ≤ SDD < 1074, likely extinction;
- 1413 < SDD ≤ 1565, likely establishment;
- SDD > 1565, almost certain establishment.
2.3. Climate Data
2.4. Assessment of the Suitability of Climate for PWD and PWN Establishment
3. Results
3.1. PWD in Healthy Trees in the Present Climate
3.2. PWN Establishment inside Healthy Trees in the Present Climate
3.3. PWD in Healthy Trees in the Future Climate
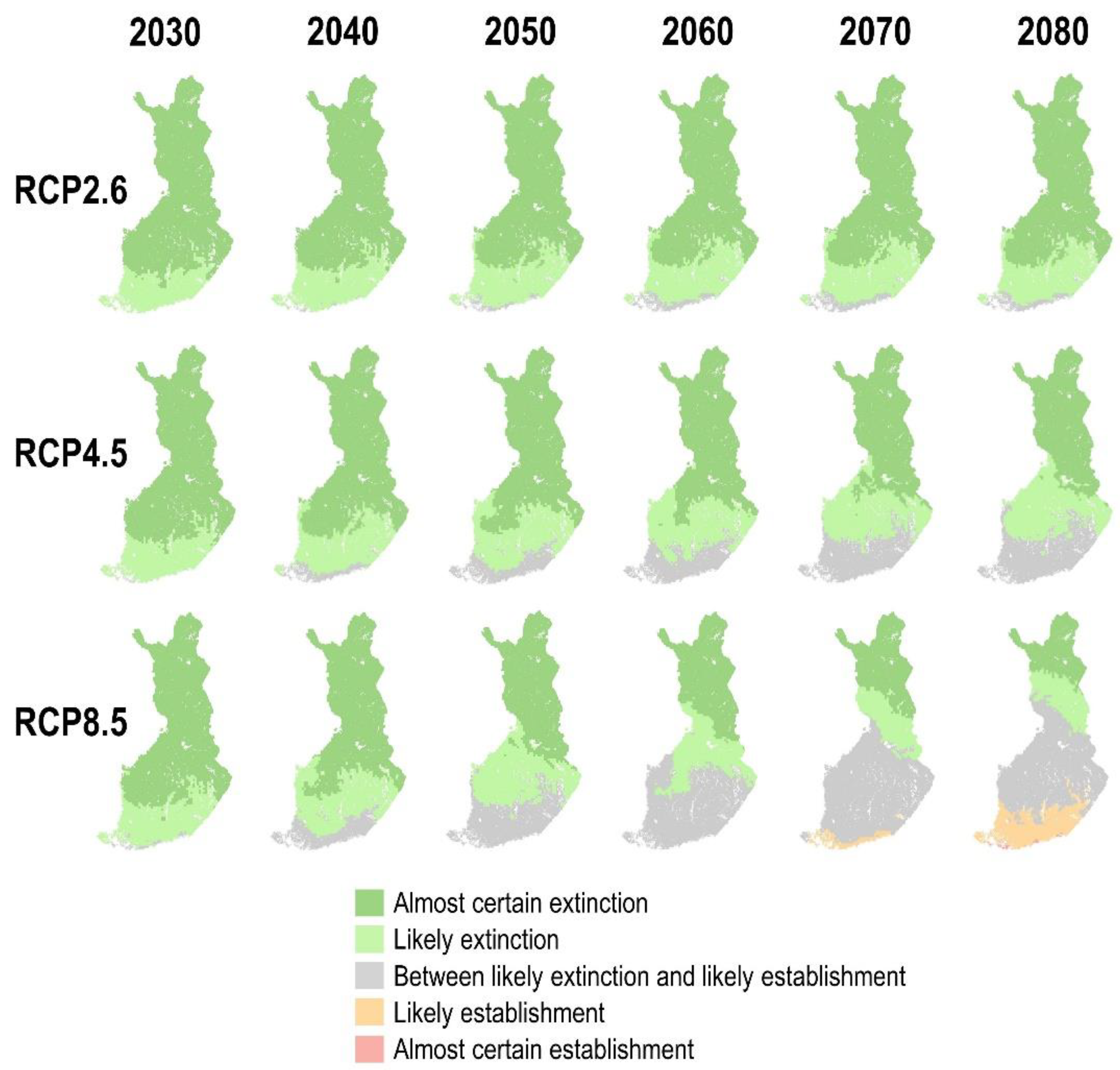
3.4. PWN Establishment inside Healthy Trees in the Future Climate
4. Discussion
4.1. The Suitability of the Finnish Climate for PWD and PWN Establishment
4.2. Comparison with Previous Studies
4.3. Strengths and Weaknesses of the Present Assessments
4.4. Implications for Risk Management
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Futai, K. Pine Wood Nematode, Bursaphelenchus xylophilus. Annu. Rev. Phytopathol. 2013, 51, 61–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Commission Implementing Decision of 26 September 2012 on Emergency Measures to Prevent the Spread within the Union of Bursaphelenchus xylophilus (Steiner et Buhrer) Nickle et al. (the Pine Wood Nematode) (Notified under Document C (2012) 6543) (2012/535/EU). Off. J. Eur. Union L 2012, 2, 42–52. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX:02012D0535-20170310 (accessed on 3 November 2021).
- Regulation (EU) 2016/2031 of the European Parliament of the Council of 26 October 2016 on protective measures against pests of plants, amending Regulations (EU) No 228/2013, (EU) No 652/2014 and (EU) No 1143/2014 of the European Parliament and of the Council and repealing Council Directives 69/464/EEC, 74/647/EEC, 93/85/EEC, 98/57/EC, 2000/29/EC, 2006/91/EC and 2007/33/EC. Off. J. Eur. Union L 2016, 317, 4–104. Available online: http://data.europa.eu/eli/reg/2016/2031/2019-12-14 (accessed on 3 November 2021).
- Evans, H.F.; McNamara, D.G.; Braasch, H.; Chadoeuf, J.; Magnusson, C. Pest Risk Analysis (PRA) for the Territories of the European Union (as PRA Area) on Bursaphelenchus xylophilus and Its Vectors in the Genus Monochamus. EPPO Bullet. 1996, 26, 199–249. [Google Scholar] [CrossRef]
- Evans, H.; Straw, N.; Watt, A. Climate Change: Implications for Insect Pests. In Climate Change: Impacts on UK Forests; Broadmeadow, M., Ed.; Technical report, Forestry Commission Bulletin 125; Forestry Commission: Edinburgh, UK, 2002; pp. 99–118. [Google Scholar]
- Gruffudd, H.R.; Jenkins, T.A.R.; Evans, H.F. Using an Evapo-Transpiration Model (ETpN) to Predict the Risk and Expression of Symptoms of Pine Wilt Disease (PWD) across Europe. Biol. Invasions 2016, 18, 2823–2840. [Google Scholar] [CrossRef]
- Mamiya, Y. History of Pine Wilt Disease in Japan. J. Nematol. 1988, 20, 219–226. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2618808/ (accessed on 3 November 2021).
- Tzean, S.S. The occurrence and biological control of Pine Wilt Disease Caused by Bursaphelenchus xylophilus. In Symposium of Forest Pathology and Entomology, Monograph No 971; Chang, T.C., Ed.; Chinese Forestry Society & Taiwan Provincial Forestry Experiment Station Press: Taiwan, 1997; pp. 7–16. (In Chinese) [Google Scholar]
- Shin, S.-C. Pine Wilt Disease in Korea. In Pine Wilt Disease; Zhao, B.G., Futai, K., Sutherland, J.R., Takeuchi, Y., Eds.; Springer: Tokyo, Japan, 2008; pp. 26–32. [Google Scholar] [CrossRef]
- Zhao, B.G. Pine Wilt Disease in China. In Pine Wilt Disease; Zhao, B.G., Futai, K., Sutherland, J.R., Takeuchi, Y., Eds.; Springer: Tokyo, Japan, 2008; pp. 18–25. [Google Scholar] [CrossRef]
- Mota, M.M.; Braasch, H.; Bravo, M.A.; Penas, A.C.; Burgermeister, W.; Metge, K.; Sousa, E. First Report of Bursaphelenchus xylophilus in Portugal and in Europe. Nematology 1999, 1, 727–734. [Google Scholar] [CrossRef]
- Robertson, L.; Arcos, S.C.; Escuer, M.; Merino, R.S.; Esparrago, G.; Abelleira, A.; Navas, A. Incidence of the Pinewood Nematode Bursaphelenchus xylophilus Steiner & Buhrer, 1934 (Nickle, 1970) in Spain. Nematology 2011, 13, 755–757. [Google Scholar] [CrossRef]
- Schröder, T.; McNamara, D.G.; Gaar, V. Guidance on Sampling to Detect Pine Wood Nematode Bursaphelenchus xylophilus in Trees, Wood and Insects. EPPO Bull. 2009, 39, 179–188. [Google Scholar] [CrossRef]
- Linit, M.J. Nematode-Vector Relationships in the Pine Wilt Disease System. J. Nematol. 1988, 20, 227–235. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2618795/ (accessed on 3 November 2021).
- Akbulut, S.; Stamps, W.T. Insect Vectors of the Pinewood Nematode: A Review of the Biology and Ecology of Monochamus Species: Insect Vectors of the Pinewood Nematode. For. Pathol. 2012, 42, 89–99. [Google Scholar] [CrossRef]
- Heliövaara, K.; Mannerkoski, I.; Siitonen, J. Suomen sarvijäärät Longhorn beetles of Finland (Coleoptera, Cerambycicae); Tremex Press: Helsinki, Finland, 2004; pp. 1–374. (In Finnish) [Google Scholar]
- Rassi, P.; Karjalainen, S.; Clayhills, T.; Helve, E.; Hyvärinen, E.; Laurinharju, E.; Malmberg, S.; Mannerkoski, I.; Martikainen, P.; Mattila, J.; et al. Provincial List of Finnish Coleoptera 2015. Sahlbergia 2015, 21 (Suppl. 1), 1–164. Available online: http://www.luomus.fi/sites/default/files/sahlbergia/sahlbergia_21_S1_2015.pdf (accessed on 3 November 2021).
- Sousa, E.; Bravo, M.A.; Pires, J.; Naves, P.; Penas, A.C.; Bonifácio, L.; Mota, M. Bursaphelenchus xylophilus (Nematoda; Aphelenchoididae) Associated with Monochamus galloprovincialis (Coleoptera; Cerambycidae) in Portugal. Nematology 2001, 3, 89–91. [Google Scholar] [CrossRef]
- Takeuchi, Y. Host Fate Following Infection by the Pine Wood Nematode. In Pine Wilt Disease; Zhao, B.G., Futai, K., Sutherland, J.R., Takeuchi, Y., Eds.; Springer: Tokyo, Japan, 2008; pp. 235–249. [Google Scholar] [CrossRef]
- Sikora, E.J.; Malek, R.B. Influence of Temperature on Development of Pine Wilt in Scots Pine. J. Nematol. 1991, 23, 188–193. [Google Scholar] [PubMed]
- Hopf-Biziks, A.; Schröder, T.; Schütz, S. The Pine Wood Nematode, Bursaphelenchus xylophilus (Steiner & Buhrer) Nickle, and Its Pathogenicity to German Pinus sylvestris Provenances. J. Plant Dis. Prot. 2016, 123, 43–49. [Google Scholar] [CrossRef]
- Mamiya, Y. Pathology of the Pine Wilt Disease Caused by Bursaphelenchus xylophilus. Annu. Rev. Phytopathol. 1983, 21, 201–220. [Google Scholar] [CrossRef]
- Malek, R.B.; Appleby, J.E. Epidemiology of Pine Wilt in Illinois. Plant Dis. 1994, 68, 180–186. [Google Scholar] [CrossRef]
- Rutherford, T.A.; Webster, J.M. Distribution of Pine Wilt Disease with Respect to Temperature in North America, Japan, and Europe. Can. J. For. Res. 1987, 17, 1050–1059. [Google Scholar] [CrossRef]
- Halik, S.; Bergdahl, D.R. Long-Term Survival of Bursaphelenchus xylophilus in Living Pinus sylvestris in an Established Plantation. For. Pathol. 1994, 24, 357–363. [Google Scholar] [CrossRef]
- Magnusson, C. Potential for Establishment of Bursaphelenchus xylophilus and the Pine Wilt Disease under Nordic Conditions. EPPO Bull. 1986, 16, 465–471. [Google Scholar] [CrossRef]
- Gruffudd, H.R.; Schröder, T.; Jenkins, T.A.R.; Evans, H.F. Modelling Pine Wilt Disease (PWD) for Current and Future Climate Scenarios as Part of a Pest Risk Analysis for Pine Wood Nematode Bursaphelenchus xylophilus (Steiner and Buhrer) Nickle in Germany. J. Plant Dis. Prot. 2019, 126, 129–144. [Google Scholar] [CrossRef]
- Pérez, G.; Díez, J.J.; Ibeas, F.; Pajares, J.A. Assessing Pine Wilt Disease Risk Under a Climate Change Scenario in Northwestern Spain. In Managing Forest Ecosystems: The Challenge of Climate Change; Bravo, F., LeMay, V., Jandl, R., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 351–365. [Google Scholar] [CrossRef]
- Hirata, A.; Nakamura, K.; Nakao, K.; Kominami, Y.; Tanaka, N.; Ohashi, H.; Takano, K.T.; Takeuchi, W.; Matsui, T. Potential Distribution of Pine Wilt Disease under Future Climate Change Scenarios. PLoS ONE 2017, 12, e0182837. [Google Scholar] [CrossRef] [Green Version]
- Ikegami, M.; Jenkins, T.A.R. Estimate Global Risks of a Forest Disease under Current and Future Climates Using Species Distribution Model and Simple Thermal Model—Pine Wilt Disease as a Model Case. For. Ecol. Manag. 2018, 409, 343–352. [Google Scholar] [CrossRef]
- van Vuuren, D.P.; Edmonds, J.; Kainuma, M.; Riahi, K.; Thomson, A.; Hibbard, K.; Hurtt, G.C.; Kram, T.; Krey, V.; Lamarque, J.-F.; et al. The representative concentration pathways: An overview. Clim. Chang. 2011, 109, 5–31. [Google Scholar] [CrossRef]
- Riahi, K.; Rao, S.; Krey, V.; Cho, C.; Chirkov, V.; Fischer, G.; Kindermann, G.; Nakicenovic, N.; Rafaj, P. RCP 8.5—A Scenario of Comparatively High Greenhouse Gas Emissions. Clim. Chang. 2011, 109, 33. [Google Scholar] [CrossRef] [Green Version]
- EU Project 265483 REPHRAME. Periodic Core Report: Project Objectives, Work Progress and Achievements, Project Management; REPHRAME Research; 2012; pp. 1–184. Available online: www.rephrame.eu (accessed on 3 November 2021).
- EU Project 265483 REPHRAME. Third Periodic Report; REPHRAME Research; 2016; pp. 1–187. Available online: www.rephrame.eu (accessed on 3 November 2021).
- Stokes, D.E. Pinewood Nematode, Bursaphelenchus lignicolus, a Nematode Pest of Some Pine Species; Technical report; Nematology Circular No. 53; Fla. Dept. Agric & Consumer Services, Division of Plant Industry: Gainesville, FL, USA, 1979; pp. 1–2. [Google Scholar]
- Sathyapala, S. Pest Risk Analysis, Biosecurity Risk to New Zealand of Pinewood Nematode (Bursaphelenchus xylophilus); Technical report; Forest Biosecurity Authority, Ministry of Agriculture and Forestry: Auckland, New Zealand, 2004. [Google Scholar]
- Wang, F.; Wang, Z.; Li, D.; Chen, Q. Identification and Characterization of a Bursaphelenchus xylophilus (Aphelenchida: Aphelenchoididae) Thermotolerance-Related Gene: Bx-HSP90. IJMS 2012, 13, 8819–8833. [Google Scholar] [CrossRef]
- Rutherford, T.A.; Riga, E.; Webster, J.M. Temperature-mediated behavioral relationships in Bursaphelenchus xylophilus, B. mucronatus and their hybrids. J. Nematol. 1992, 24, 4–44. [Google Scholar]
- Hitchin, E.R. Estimating Monthly Degree-Days. Build. Serv. Eng. Res. Technol. 1983, 4, 159–162. [Google Scholar] [CrossRef]
- Futai, K. Developmental Rate and Population Growth of Bursaphelenchus lignicolus (Nematoda: Aphelenchoididae) and B. mucronatus. Appl. Entomol. Zool. 1980, 15, 115–122. [Google Scholar] [CrossRef] [Green Version]
- Nowosad, J. Pollen: Analysis of Aerobiological Data. R Package Version 0.71. 2019. Available online: https://CRAN.R-project.org/package=pollen (accessed on 3 November 2021).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: https://www.r-project.org/ (accessed on 3 November 2021).
- Venables, W.N.; Ripley, B.D.; Venables, W.N. Modern Applied Statistics with S, 4th ed.; Statistics and computing; Springer: New York, NY, USA, 2002; pp. 1–497. ISBN 0-387-95457-0. [Google Scholar]
- Schlegel, B.; Steenbergen, M. Brant: Test for Parallel Regression Assumption, R package version 0.3-0; 2020. Available online: https://CRAN.R-project.org/package=brant (accessed on 3 November 2021).
- Aalto, J.; Pirinen, P.; Jylhä, K. New Gridded Daily Climatology of Finland: Permutation-based Uncertainty Estimates and Temporal Trends in Climate. J. Geophys. Res. Atmos. 2016, 121, 3807–3823. [Google Scholar] [CrossRef] [Green Version]
- Ruosteenoja, K.; Jylhä, K.; Kämäräinen, M. Climate Projections for Finland Under the RCP Forcing Scenarios. Geophysica 2016, 51, 17–50. [Google Scholar]
- Härmä, P.; Autio, I.; Teiniranta, R.; Hatunen, S.; Törmä, M.; Kallio, M.; Kaartinen, M. Copernicus Land Monitoring 2014–2020 in the Framework of Regulation (EU) No 377/2014 of the European Parliament and of the Council of 3 April 2014; Final Report; Finnish Environment Institute (SYKE), Geoinformatics and Land Use Division (GEO): Helsinki, Finland, 2019; pp. 1–27. [Google Scholar]
- Dropkin, V.H.; Foudin, A.; Kondo, E.; Linit, M.; Smith, M.; Robbins, K. Pinewood Nematode: A Threat. Plant Dis. 1981, 65, 12. [Google Scholar] [CrossRef]
- EPPO. Bursaphelenchus xylophilus. EPPO Datasheets on Pests Recommended for Regulation. Available online: https://gd.eppo.int (accessed on 3 November 2021).
- Donald, P.A.; Stamps, W.T.; Linit, M.J.; Todd, T.C. Pine Wilt Disease. The Plant Health Instructor; The American Phytopathological Society: New York, NY, USA, 2016. [Google Scholar] [CrossRef]
- Oh, W.-S.; Jeong, P.-Y.; Joo, H.-J.; Lee, J.-E.; Moon, Y.-S.; Cheon, H.-M.; Kim, J.-H.; Lee, Y.-U.; Shim, Y.-H.; Paik, Y.-K. Identification and Characterization of a Dual-Acting Antinematodal Agent against the Pinewood Nematode, Bursaphelenchus xylophilus. PLoS ONE 2009, 4, e7593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuroda, K.; Yamada, T.; Mineo, K.; Tamura, H. Effects of Cavitation on the Development of Pine Wilt Disease Caused by Bursaphelenchus xylophilus. Jpn. J. Phytopathol. 1988, 54, 606–615. [Google Scholar] [CrossRef] [Green Version]
- Zhao, B.G.; Futai, K.; Sutherland, J.R.; Takeuchi, Y. Pine Wilt Disease; Springer: Tokyo, Japan, 2008; pp. 1–459. [Google Scholar]
- Commission Delegated Regulation (EU) 2019/1702 of 1 August 2019 Supplementing Regulation (EU) 2016/2031 of the European Parliament and of the Council by Establishing the List of Priority Pests C/2019/5637. Off. J. the European Union. 2019, 260, pp. 8–10. Available online: http://data.europa.eu/eli/reg_del/2019/1702/oj (accessed on 3 November 2021).
- Kukkonen, H.; Pouttu, A.; Tomminen, J.; Lilja, P.; Ronni, P.; Niemi, K.; Sahin, T.; Soukainen, J.; Kainulainen, A.; Islander, A.; et al. Contingency plan for pine wood nematode. Evira Publ. 2011, 7, 1–68. (In Finnish) [Google Scholar]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tuomola, J.; Gruffudd, H.; Ruosteenoja, K.; Hannunen, S. Could Pine Wood Nematode (Bursaphelenchus xylophilus) Cause Pine Wilt Disease or Even Establish inside Healthy Trees in Finland Now—Or Ever? Forests 2021, 12, 1679. https://doi.org/10.3390/f12121679
Tuomola J, Gruffudd H, Ruosteenoja K, Hannunen S. Could Pine Wood Nematode (Bursaphelenchus xylophilus) Cause Pine Wilt Disease or Even Establish inside Healthy Trees in Finland Now—Or Ever? Forests. 2021; 12(12):1679. https://doi.org/10.3390/f12121679
Chicago/Turabian StyleTuomola, Juha, Hannah Gruffudd, Kimmo Ruosteenoja, and Salla Hannunen. 2021. "Could Pine Wood Nematode (Bursaphelenchus xylophilus) Cause Pine Wilt Disease or Even Establish inside Healthy Trees in Finland Now—Or Ever?" Forests 12, no. 12: 1679. https://doi.org/10.3390/f12121679