Diversity Patterns Associated with Varying Dispersal Capabilities as a Function of Spatial and Local Environmental Variables in Small Wetlands in Forested Ecosystems

 ,
, 
Abstract
:1. Introduction
2. Methods
2.1. Study Area
2.2. Environmental Predictors
2.2.1. Wetland Metrics
2.2.2. Water Quality
2.2.3. Phytoplankton and Benthic Algae Biomass
2.3. Response Variables
2.3.1. Amphibians
2.3.2. Aquatic Macroinvertebrates
2.3.3. Zooplankton
2.3.4. Statistical Analyses
3. Results
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ceballos, G.; Ehrlich, P.R.; Barnosky, A.D.; García, A.; Pringle, R.M.; Palmer, T.M. Accelerated modern human–induced species losses: Entering the sixth mass extinction. Sci. Adv. 2015, 1, e1400253. [Google Scholar] [CrossRef] [Green Version]
- Gibbs, J.P. Importance of small wetlands for the persistence of local populations of wetland-associated animals. Wetlands 1993, 13, 25–31. [Google Scholar] [CrossRef]
- Dudgeon, D.; Gao, B. Weak effects of plant diversity on leaf-litter breakdown in a tropical stream. Mar. Freshw. Res. 2010, 61, 1218–1225. [Google Scholar] [CrossRef]
- Margono, B.A.; Potapov, P.V.; Turubanova, S.; Stolle, F.; Hansen, M.C. Primary forest cover loss in Indonesia over 2000–2012. Nat. Clim. Chang. 2014, 4, 730–735. [Google Scholar] [CrossRef]
- Hairston, N.G., Jr. Zooplankton egg banks as biotic reservoirs in changing environments. Limnol. Oceanogr. 1996, 41, 1087–1092. [Google Scholar] [CrossRef]
- Poff, N.L. Landscape filters and species traits: Towards mechanistic understanding and prediction in stream ecology. J. N. Am. Benthol. Soc. 1997, 16, 391–409. [Google Scholar] [CrossRef]
- Beachy, C.K.; Ryan, T.J.; Bonett, R.M. How Metamorphosis Is Different in Plethodontids: Larval Life History Perspectives on Life-Cycle Evolution. Herpetologica 2017, 73, 252–258. [Google Scholar] [CrossRef] [Green Version]
- Socolar, J.B.; Gilroy, J.J.; Kunin, W.E.; Edwards, D.P. How should beta-diversity inform biodiversity conservation? Trends Ecol. Evol. 2016, 31, 67–80. [Google Scholar] [CrossRef] [Green Version]
- Qian, H. Beta diversity in relation to dispersal ability for vascular plants in North America. Glob. Ecol. Biogeogr. 2009, 18, 327–332. [Google Scholar] [CrossRef]
- Brown, B.; Swan, C. Dendritic network structure constrains metacommunity properties in riverine ecosystems. J. Anim. Ecol. 2010, 79, 571–580. [Google Scholar] [CrossRef]
- Mouquet, N.; Loreau, M. Community patterns in source-sink metacommunities. Am. Nat. 2003, 162, 544–557. [Google Scholar] [CrossRef] [Green Version]
- Zedler, P.H. Vernal pools and the concept of “isolated wetlands”. Wetlands 2003, 23, 597–607. [Google Scholar] [CrossRef]
- Allen, M.R. Measuring and modeling dispersal of adult zooplankton. Oecologia 2007, 153, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Brooks, R.T. Potential impacts of global climate change on the hydrology and ecology of ephemeral freshwater systems of the forests of the northeastern United States. Clim. Chang. 2009, 95, 469–483. [Google Scholar] [CrossRef]
- Skelly, D.K. Tadpole communities: Pond permanence and predation are powerful forces shaping the structure of tadpole communities. Am. Sci. 1997, 85, 36–45. [Google Scholar]
- Cornell, H.V.; Lawton, J.H. Species interactions, local and regional processes, and limits to the richness of ecological communities: A theoretical perspective. J. Anim. Ecol. 1992, 61, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Ricklefs, R.E. Community diversity: Relative roles of local and regional processes. Science 1987, 235, 167–171. [Google Scholar] [CrossRef] [PubMed]
- Oertli, B.; Joye, D.A.; Castella, E.; Juge, R.; Cambin, D.; Lachavanne, J.-B. Does size matter? The relationship between pond area and biodiversity. Biol. Conserv. 2002, 104, 59–70. [Google Scholar] [CrossRef] [Green Version]
- Thompson, R.; Townsend, C. A truce with neutral theory: Local deterministic factors, species traits and dispersal limitation together determine patterns of diversity in stream invertebrates. J. Anim. Ecol. 2006, 75, 476–484. [Google Scholar] [CrossRef]
- Schwartz, S.S.; Jenkins, D.G. Temporary aquatic habitats: Constraints and opportunities. Aquat. Ecol. 2000, 34, 3–8. [Google Scholar] [CrossRef]
- Sarremejane, R.; England, J.; Sefton, C.E.M.; Parry, S.; Eastman, M.; Stubbington, R. Local and regional drivers influence how aquatic community diversity, resistance and resilience vary in response to drying. Oikos 2020. [Google Scholar] [CrossRef]
- Welborn, G.A.; Skelly, D.K.; Werner, E.E. Mechanisms creating community structure across a freshwater habitat gradient. Annu. Rev. Ecol. Syst. 1996, 27, 337–363. [Google Scholar] [CrossRef] [Green Version]
- Nicolet, P.; Biggs, J.; Fox, G.; Hodson, M.J.; Reynolds, C.; Whitfield, M.; Williams, P. The wetland plant and macroinvertebrate assemblages of temporary ponds in England and Wales. Biol. Conserv. 2004, 120, 261–278. [Google Scholar] [CrossRef]
- Serrano, L.; Fahd, K. Zooplankton communities across a hydroperiod gradient of temporary ponds in the Doñana National Park (SW Spain). Wetlands 2005, 25, 101–111. [Google Scholar] [CrossRef]
- Vanschoenwinkel, B.; Gielen, S.; Seaman, M.; Brendonck, L. Any way the wind blows-frequent wind dispersal drives species sorting in ephemeral aquatic communities. Oikos 2008, 117, 125–134. [Google Scholar] [CrossRef]
- Paton, P.W.C.; Crouch, W.B. Using the phenology of pond-breeding amphibians to develop conservation strategies. Conserv. Biol. 2002, 16, 194–204. [Google Scholar] [CrossRef] [Green Version]
- Newman, R.A. Adaptive plasticity in amphibian metamorphosis. BioScience 1992, 42, 671–678. [Google Scholar] [CrossRef]
- Semlitsch, R.D. Differentiating migration and dispersal processes for pond-breeding amphibians. J. Wildl. Manag. 2008, 72, 260–267. [Google Scholar] [CrossRef]
- Semlitsch, R.D.; Bodie, J.R. Biological criteria for buffer zones around wetlands and riparian habitats for amphibians and reptiles. Conserv. Biol. 2003, 17, 1219–1228. [Google Scholar] [CrossRef] [Green Version]
- Kovar, R.; Brabec, M.; Vita, R.; Bocek, R. Spring migration distances of some Central European amphibian species. Amphibia Reptilia 2009, 30, 367–378. [Google Scholar] [CrossRef]
- Havel, J.E.; Shurin, J.B. Mechanisms, effects, and scales of dispersal in freshwater zooplankton. Limnol. Oceanogr. 2004, 49, 1229–1238. [Google Scholar] [CrossRef]
- Abell, R.A.; Olson, D.M.; Fund, W.W.; Dinerstein, E.; Eichbaum, W.; Diggs, J.T.; Hurley, P.; Walters, S.; Wettengel, W.; Allnutt, T.; et al. Freshwater Ecoregions of North America: A Conservation Assessment; Island Press: Washington, DC, USA, 2000; ISBN 978-1-55963-734-3. [Google Scholar]
- Milanovich, J.R.; Peterman, W.E.; Nibbelink, N.P.; Maerz, J.C. Projected Loss of a Salamander Diversity Hotspot as a Consequence of Projected Global; Loyola University Chicago: Chicago, IL, USA, 2010. [Google Scholar]
- Napton, D.E.; Auch, R.F.; Headley, R.; Taylor, J.L. Land changes and their driving forces in the Southeastern United States. Reg. Environ. Chang. 2010, 10, 37–53. [Google Scholar] [CrossRef]
- Pitt, A.L.; Baldwin, R.F.; Lipscomb, D.J.; Brown, B.L.; Hawley, J.E.; Allard-Keese, C.M.; Leonard, P.B. The missing wetlands: Using local ecological knowledge to find cryptic ecosystems. Biodivers. Conserv. 2012, 21, 51–63. [Google Scholar] [CrossRef]
- Griffith, J.A.; Stehman, S.V.; Loveland, T.R. Landscape trends in mid-Atlantic and southeastern United States ecoregions. Environ. Manag. 2003, 32, 572–588. [Google Scholar] [CrossRef]
- Brown, D.G.; Johnson, K.M.; Loveland, T.R.; Theobald, D.M. Rural land-use trends in the conterminous United States, 1950–2000. Ecol. Appl. 2005, 15, 1851–1863. [Google Scholar] [CrossRef] [Green Version]
- Pitt, A.L.; Howard, J.H.; Baldwin, R.F.; Baldwin, E.D.; Brown, B.L. Small Parks as Local Social–Ecological Systems Contributing to Conservation of Small Isolated and Ephemeral Wetlands. Nat. Areas J. 2018, 38, 237–249. [Google Scholar] [CrossRef]
- Stumpf, K.A. The estimation of forest vegetation cover descriptions using a vertical densitometer. In Proceedings of the Joint Inventory and Biometrics Working Groups Session at the SAF National Convention, Indianapolis, IN, USA, 8–10 November 1993. [Google Scholar]
- Johansson, T. Estimating canopy density by the vertical tube method. For. Ecol. Manag. 1985, 11, 139–144. [Google Scholar] [CrossRef]
- Schiesari, L. Pond canopy cover: A resource gradient for anuran larvae. Freshw. Biol. 2006, 51, 412–423. [Google Scholar] [CrossRef] [Green Version]
- Boes, M.W.; Benard, M.F. Carry-Over Effects in Nature: Effects of Canopy Cover and Individual Pond on Size, Shape, and Locomotor Performance of Metamorphosing Wood Frogs. Copeia 2013, 2013, 717–722. [Google Scholar] [CrossRef]
- Rowland, F.E.; Tuttle, S.K.; González, M.J.; Vanni, M.J. Canopy cover and anurans: Nutrients are the most important predictor of growth and development. Can. J. Zool. 2016, 94, 225–232. [Google Scholar] [CrossRef] [Green Version]
- Siewicki, T.C.; Pullaro, T.; Pan, W.; McDaniel, S.; Glenn, R.; Stewart, J. Models of total and presumed wildlife sources of fecal coliform bacteria in coastal ponds. J. Environ. Manag. 2007, 82, 120–132. [Google Scholar] [CrossRef]
- Stevenson, R.J.; McCormick, P.V.; Frydenborg, R. Methods for Evaluating Wetland Condition; FAO: Rome, Italy, 2002. [Google Scholar]
- Barbour, M.T.; Gerritsen, J.; Snyder, B.D.; Stribling, J.B. Rapid Bioassessment Protocols for Use in Streams and Wadeable Rivers: Periphyton, Benthic Macroinvertebrates and Fish; US Environmental Protection Agency, Office of Water: Washington, DC, USA, 1999; Volume 339.
- Danielson, T.J. Methods for Evaluating Wetland Condition 14 Wetland Biological Assessment Case Studies; Diane Publishing: Collingdale, PA, USA, 2003. [Google Scholar]
- Biggs, B.J.; Kilroy, C. Stream Periphyton Monitoring Manual; National Institute of Water and Atmospheric Research: Christchurch, New Zealand, 2000. [Google Scholar]
- Ward, H.B.; Whipple, G.C. Fresh-Water Biology; John Wiley & Sons, Inc.: New York, NY, USA, 1918. [Google Scholar]
- R Core Team. R: A Language Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2014. [Google Scholar]
- Charney, N.; Record, S. Vegetarian: Jost Diversity Measures for Community Data. 2012. Available online: https://rdrr.io/cran/vegetarian/ (accessed on 1 July 2017).
- Jost, L. Entropy and diversity. Oikos 2006, 113, 363–375. [Google Scholar] [CrossRef]
- Jost, L. Partitioning diversity into independent alpha and beta components. Ecology 2007, 88, 2427–2439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chao, A.; Jost, L.; Chiang, S.C.; Jiang, Y.-H.; Chazdon, R.L. A two-stage probabilistic approach to multiple-community similarity indices. Biometrics 2008, 64, 1178–1186. [Google Scholar] [CrossRef]
- Borcard, D.; Legendre, P.; Drapeau, P. Partialling out the spatial component of ecological variation. Ecology 1992, 73, 1045–1055. [Google Scholar] [CrossRef] [Green Version]
- Poos, M.S.; Jackson, D.A. Addressing the removal of rare species in multivariate bioassessments: The impact of methodological choices. Ecol. Indic. 2012, 18, 82–90. [Google Scholar] [CrossRef]
- Borcard, D.; Legendre, P. All-scale spatial analysis of ecological data by means of principal coordinates of neighbour matrices. Ecol. Model. 2002, 153, 51–68. [Google Scholar] [CrossRef]
- Oksanen, J.; Kindt, R.; Legendre, P.; O’Hara, B.; Stevens, M.H.H.; Oksanen, M.J.; Suggests, M. The Vegan Package; The R Foundation: Vienna, Austria, 2007. [Google Scholar]
- Sinsch, U. Mini-review: The orientation behaviour of amphibians. Herpetol. J. 1991, 1, 1–544. [Google Scholar]
- Gill, D.E. The metapopulation ecology of the red-spotted newt, Notophthalmus viridescens (Rafinesque). Ecol. Monogr. 1978, 48, 145–166. [Google Scholar] [CrossRef]
- Gamble, L.R.; McGarigal, K.; Compton, B.W. Fidelity and dispersal in the pond-breeding amphibian, Ambystoma opacum: Implications for spatio-temporal population dynamics and conservation. Biol. Conserv. 2007, 139, 247–257. [Google Scholar] [CrossRef]
- Berven, K.A.; Grudzien, T.A. Dispersal in the wood frog (Rana sylvatica): Implications for genetic population structure. Evolution 1990, 44, 2047–2056. [Google Scholar] [PubMed]
- Smith, A.M.; Green, D.M. Dispersal and the metapopulation paradigm in amphibian ecology and conservation: Are all amphibian populations metapopulations? Ecography 2005, 28, 110–128. [Google Scholar] [CrossRef]
- Drake, D.L.; Ousterhout, B.H.; Johnson, J.R.; Anderson, T.L.; Peterman, W.E.; Shulse, C.D.; Hocking, D.J.; Lohraff, K.L.; Harper, E.B.; Rittenhouse, T.A.; et al. Pond-breeding amphibian community composition in Missouri. Am. Midl. Nat. 2015, 174, 180–187. [Google Scholar] [CrossRef]
- Drayer, A.N.; Richter, S.C. Physical wetland characteristics influence amphibian community composition differently in constructed wetlands and natural wetlands. Ecol. Eng. 2016, 93, 166–174. [Google Scholar] [CrossRef]
- Ernst, R.; Rödel, M.-O. Patterns of community composition in two tropical tree frog assemblages: Separating spatial structure and environmental effects in disturbed and undisturbed forests. J. Trop. Ecol. 2008, 24, 111–120. [Google Scholar] [CrossRef]
- Petranka, J.W.; Smith, C.K.; Floyd Scott, A. Identifying the minimal demographic unit for monitoring pond-breeding amphibians. Ecol. Appl. 2004, 14, 1065–1078. [Google Scholar] [CrossRef]
- Hopey, M.E.; Petranka, J.W. Restriction of wood frogs to fish-free habitats: How important is adult choice? Copeia 1994, 1994, 1023–1025. [Google Scholar] [CrossRef]
- Horváth, Z.; Vad, C.F.; Tóth, A.; Zsuga, K.; Boros, E.; Vörös, L.; Ptacnik, R. Opposing patterns of zooplankton diversity and functioning along a natural stress gradient: When the going gets tough, the tough get going. Oikos 2014, 123, 461–471. [Google Scholar] [CrossRef] [Green Version]
- Sokol, E.R.; Brown, B.L.; Carey, C.C.; Tornwall, B.M.; Swan, C.M.; Barrett, J.E. Linking management to biodiversity in built ponds using metacommunity simulations. Ecol. Model. 2015, 296, 36–45. [Google Scholar] [CrossRef]
- Astorga, A.; Oksanen, J.; Luoto, M.; Soininen, J.; Virtanen, R.; Muotka, T. Distance decay of similarity in freshwater communities: Do macro- and microorganisms follow the same rules? Glob. Ecol. Biogeogr. 2012, 21, 365–375. [Google Scholar] [CrossRef]
- Williams, D.D.; Hynes, H.B.N. Stream habitat selection by aerially colonizing invertebrates. Can. J. Zool. 1976, 54, 685–693. [Google Scholar] [CrossRef]
- Schwind, R. Spectral regions in which aquatic insects see reflected polarized light. J. Comp. Physiol. A 1995, 177, 439–448. [Google Scholar] [CrossRef]
- Skelly, D.K. Distributions of pond-breeding anurans: An overview of mechanisms. Isr. J. Zool. 2001, 47, 313–332. [Google Scholar] [CrossRef]
- Church, D.R. Role of current versus historical hydrology in amphibian species turnover within local pond communities. Copeia 2008, 2008, 115–125. [Google Scholar] [CrossRef]
- Urban, M.C.; Tewksbury, J.J.; Sheldon, K.S. On a collision course: Competition and dispersal differences create no-analogue communities and cause extinctions during climate change. Proc. R. Soc. Lond. B Biol. Sci. 2012, 279, 2072–2080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cadotte, M.W. Dispersal and species diversity: A meta-analysis. Am. Nat. 2006, 167, 913–924. [Google Scholar] [CrossRef]
- Leibold, M.; Holyoak, M.; Mouquet, N.; Amarasekare, P.; Chase, J.; Hoopes, M.; Holt, R.; Shurin, J.; Law, R.; Tilman, D.; et al. The metacommunity concept: A framework for multi-scale community ecology. Ecol. Lett. 2004, 7, 601–613. [Google Scholar] [CrossRef]
- Marsh, D.M.; Trenham, P.C. Metapopulation dynamics and amphibian conservation. Conserv. Biol. 2001, 15, 40–49. [Google Scholar] [CrossRef]
- Semlitsch, R.D. Critical elements for biologically based recovery plans of aquatic-breeding amphibians. Conserv. Biol. 2002, 16, 619–629. [Google Scholar] [CrossRef] [Green Version]
- Cushman, S.A. Effects of habitat loss and fragmentation on amphibians: A review and prospectus. Biol. Conserv. 2006, 128, 231–240. [Google Scholar] [CrossRef]
- Wilson, M.A.; Carpenter, S.R. Economic valuation of freshwater ecosystem services in the United States: 1971–1997. Ecol. Appl. 1999, 9, 772–783. [Google Scholar]
- Zedler, J.B.; Kercher, S. Wetland resources: Status, trends, ecosystem services, and restorability. Annu. Rev. Environ. Resour. 2005, 30, 39–74. [Google Scholar] [CrossRef] [Green Version]
- Dudgeon, D.; Arthington, A.H.; Gessner, M.O.; Kawabata, Z.-I.; Knowler, D.J.; Lévêque, C.; Naiman, R.J.; Prieur-Richard, A.-H.; Soto, D.; Stiassny, M.L.J.; et al. Freshwater biodiversity: Importance, threats, status and conservation challenges. Biol. Rev. 2006, 81, 163–182. [Google Scholar] [CrossRef] [PubMed]
- Amezaga, J.M.; Santamaría, L.; Green, A.J. Biotic wetland connectivity—Supporting a new approach for wetland policy. Acta Oecol. 2002, 23, 213–222. [Google Scholar] [CrossRef] [Green Version]
- Johnson, P.T.; Hoverman, J.T.; McKenzie, V.J.; Blaustein, A.R.; Richgels, K.L. Urbanization and wetland communities: Applying metacommunity theory to understand the local and landscape effects. J. Appl. Ecol. 2013, 50, 34–42. [Google Scholar] [CrossRef]
- The Navigable Waters Protection Rule: Definition of “Waters of the United States”. Available online: https://www.federalregister.gov/documents/2020/04/21/2020-02500/the-navigable-waters-protection-rule-definition-of-waters-of-the-united-states (accessed on 20 September 2020).




{kind=link}
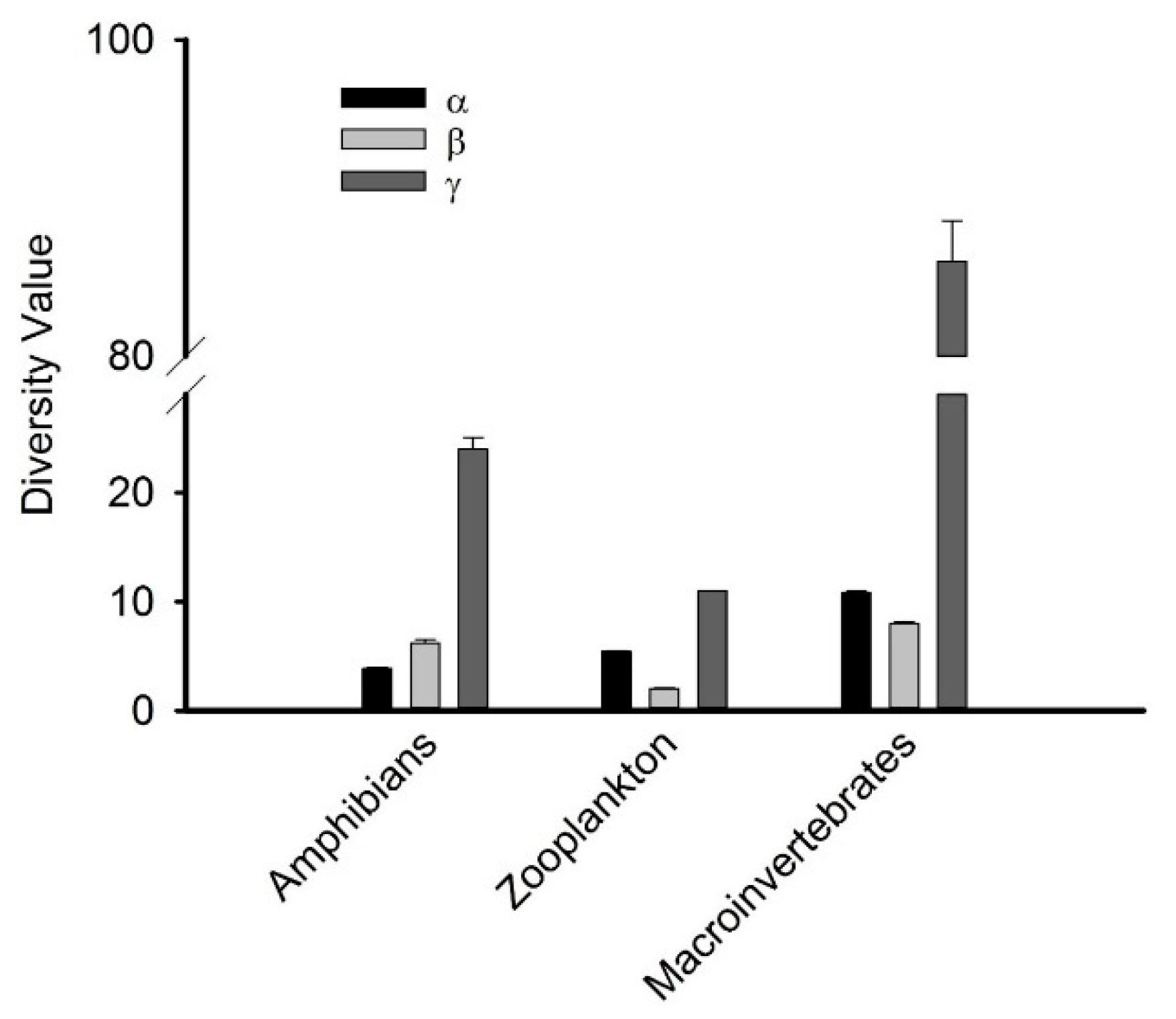{kind=link}
{kind=link}
| Taxa | Variables | Adj R2 | F | p Value |
|---|---|---|---|---|
| Amphibians | 0.15 | 1.304 | 0.047 | |
| Space (a + b) | 0.11 | 1.53 | 0.012 | |
| Environment (b + c) | 0.03 | 1.26 | 0.093 | |
| Environment|Space (a) | 0.03 | 1.36 | 0.308 | |
| Space|Environment (b) | 0.12 | 1.09 | 0.057 * | |
| Zooplankton | 0.05 | 1.06 | 0.41 | |
| Space (a + b) | 0.00 | 0.95 | 0.573 | |
| Environment (b + c) | 0.00 | 1.08 | 0.37 | |
| Environment|Space (a) | 0.05 | 1.13 | 0.31 | |
| Space|Environment (b) | 0.10 | 1.05 | 0.38 | |
| Macroinvertebrates | 0.144 | 1.22 | 0.058 * | |
| Space (a + b) | 0.02 | 1.00 | 0.50 | |
| Environment (b + c) | 0.14 | 1.26 | 0.056 * | |
| Environment|Space (a) | 0.12 | 1.58 | 0.003 | |
| Space|Environment (b) | 0.00 | 1.19 | 0.09 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tornwall, B.M.; Pitt, A.L.; Brown, B.L.; Hawley-Howard, J.; Baldwin, R.F. Diversity Patterns Associated with Varying Dispersal Capabilities as a Function of Spatial and Local Environmental Variables in Small Wetlands in Forested Ecosystems. Forests 2020, 11, 1146. https://doi.org/10.3390/f11111146
Tornwall BM, Pitt AL, Brown BL, Hawley-Howard J, Baldwin RF. Diversity Patterns Associated with Varying Dispersal Capabilities as a Function of Spatial and Local Environmental Variables in Small Wetlands in Forested Ecosystems. Forests. 2020; 11(11):1146. https://doi.org/10.3390/f11111146
Chicago/Turabian StyleTornwall, Brett M., Amber L. Pitt, Bryan L. Brown, Joanna Hawley-Howard, and Robert F. Baldwin. 2020. "Diversity Patterns Associated with Varying Dispersal Capabilities as a Function of Spatial and Local Environmental Variables in Small Wetlands in Forested Ecosystems" Forests 11, no. 11: 1146. https://doi.org/10.3390/f11111146