Changes in Spruce Growth and Biomass Allocation Following Thinning and Guying Treatments


1
Forest Research, Northern Research Station, Roslin, Midlothian, Scotland EH25 9SY, UK
2
European Forest Institute, 69, Route d’Arcachon, 33612 Cestas-Pierroton, France
*
Author to whom correspondence should be addressed.
Forests 2019, 10(3), 253; https://doi.org/10.3390/f10030253
Submission received: 28 January 2019
/
Revised: 5 March 2019
/
Accepted: 8 March 2019
/
Published: 13 March 2019
(This article belongs to the Section Forest Ecology and Management)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:When forest stands are thinned, the retained trees are exposed to increased light and greater mechanical strain from the wind. The consequent greater availability of photosynthate and localised mechanical strain in stems and roots are both known to increase cambial growth in conifers, but their relative importance has not previously been quantified. Light availability and wind movement were manipulated in a 10-year-old stand of Sitka spruce trees on an exposed upland site. Treatments were “Control”—no change in spacing or wind loading; “Thinned”—light availability and wind loading increased by removing neighbouring trees; “Thinned and guyed”—light increased and wind loading reduced by removing neighbouring trees and guying stems with wires. Twelve trees per treatment were maintained and monitored for four years before harvesting and removal of cross-sectional stem samples from four heights for measurement of radial growth response. Root systems were excavated from each treatment for observations of associated root growth responses. The “Thinned” treatment and “Thinned and guyed” treatment showed no significant stem growth response in the first year after treatment, but very large increases in the second and subsequent years. There were much larger growth responses in the “Thinned” treatment than in the “Thinned and guyed” treatment, especially in the lower stem. Similar growth responses were observed in the structural roots, close to the stem base. These increases in stem and root growth in response to wind movement corresponded with a reduction of branch growth. Such changes in allocation have implications for the hydraulic and biomechanical requirements of trees, and should be incorporated into tree growth and stability models.
1. Introduction
Forest thinning to reduce stand density commonly results in large observable growth responses, as a result of increased light availability and increased photosynthetic area [1,2,3]. At the same time, trees must also stabilise against a greater vulnerability to overturning from increased wind exposure and increasing above-ground (stem, branch and leaf) mass [4]. Gardiner et al. [5] showed a linear increase in maximum bending moment (wind force x distance from centre of force to ground) of Sitka spruce (Picea sitchensis (Bong.) Carr.) trees with an increasing ratio of spacing to height. Trees have been observed to acclimate to a new wind regime within a few years of thinning treatments [6,7], and exposed trees achieve this by allocating a greater proportion of available biomass to the stem base and below-ground [8,9,10,11].
In experiments to study the growth of Sitka spruce seedlings that were mechanically stimulated either in wind tunnels or by attaching them to flexing machines, Stokes at al. [12,13] observed the development of an elliptical cross-sectional shape at the base of the stem with increased thickening in the wind direction axis, and greatest thickening of structural roots on the windward and leeward sides of the tree. Conversely, a reduction of stem and root secondary thickening was induced in a number of experiments by preventing stem sway [14,15,16,17]. Fayle [14] removed soil from around the structural roots at the base of 3.4 m tall, Scots pine (Pinus sylvestris L.) trees to allow increased wind movement, and guyed others to prevent movement. Trees with increased movement showed a large increase in stem radial growth, in comparison to guyed trees. Such growth responses increase the strength and rigidity of trees [18] and so reduce wind movement and vulnerability to wind damage [11,19,20]. It may also act to compensate for the reduced stem sapwood hydraulic conductivity that can result from xylem damage induced by stem flexing [21], in order to maintain sapwood water transport [22].
To understand the effect of thinning in terms of biomass allocation and wind stability, it is necessary to separate the responses to increased light from any concurrent responses to increased mechanical strain that will be greatest towards the base of the tree [23,24]. Previous studies of responses to stand thinning have not included treatments that increase light availability while controlling wind-induced mechanical strain. Therefore, in this paper we describe a novel field experiment that examined the effects of increased mechanical strain and light exposure on tree growth and development. The experiment, conducted between 1996 and 1999, has been analysed now to inform the incorporation of wind acclimation effects into wind risk models. Changes in stem, root and branch growth of Sitka spruce trees were examined following thinning and guying treatments in which light availability was increased and wind movement was either increased or decreased. This investigation was designed to improve our understanding of tree physiology and growth responses to changes in light availability and wind exposure, and to explore the implications of increased exposure on growth, wind stability and management of natural and plantation forests. The hypotheses that we test are as follows:
- Forest trees respond through enhanced radial growth to increased wind loading and light availability following thinning, with the greatest increase in growth towards the base of the stem.
- If tree movement is reduced while light availability is increased, the increase in radial growth will be more even up the stem.
2. Materials and Methods
The experiment was conducted within a forest plantation located at Crookburn Hill in Kershope Forest, Cumbria, North England (55°06′ N, 2°47′ W, 220 m altitude). The site was flat and had an area of an approximately 1500 m2, on a uniform gentle slope of less than 3° with a south-east aspect. Windiness, as measured by the Detailed Aspect Method of Scoring (DAMS) system [25], was DAMS 15 (a typical exposure for upland conifer forests in the UK). Soil was predominantly surface-water gley [26]; i.e., a “stagno-orthic gley” soil as classified by Avery [27]. Trees were Sitka spruce (Queen Charlotte Islands provenance), planted on the site as “1 + 1” transplants [28] in early 1986. They were “flat-planted” without site cultivation, at an average spacing of 1.7 m. The stand had not been thinned or “brashed” (to remove lower branches) before experimental treatments were applied.
2.1. Experiment Details
Treatments were applied when the trees were ten-years old, between 26 and 29 February 1996. The experiment consisted of two adjacent blocks, each containing three approximately rectangular plots, i.e., one plot per treatment in each block. The plots each had an area of 180 m2, and each contained 90 trees prior to treatment. The three treatments were located randomly within each block. Six experimental trees were selected randomly from a central area of 30 trees in each plot, using a table of pre-generated random numbers. Unthinned buffers (approximately 3.4 m width) consisting of two lines of trees, were left around the central treatment area within each plot. Treatments (see Figure 1) were applied to the trees as follows:
- “Control” trees were labelled and left untreated, with no thinning of the plot. These trees had no change in wind loading or light availability.
- “Thinned” treatment trees experienced both increased wind loading and light availability. They were exposed by removing all immediately neighbouring trees (4 trees) from around each experimental tree (see Figure 1), which reduced the density (trees ha−1) to approximately 25% of the control plot density.
- “Thinned and Guyed” treatment trees were exposed in the same way as in the Thinned treatment, i.e., the plots were thinned by removal of four immediate neighbour trees around each experimental tree. These were then guyed to stabilise them against wind movement. This treatment increased light availability while restricting wind movement. Trees were guyed by attaching three steel wires (3 mm diameter) to separate eye-screws fixed to each tree at 0.5 × tree height (Figure 1) and secured to 0.75 m long, steel ground-anchors hammered 0.65 m into the ground at a distance of 0.5 × tree height from the tree base. Guy wires were positioned at 120° spacing around the trees. The guy wires were tightened to similar tension on each tree and were re-tightened as necessary in each year of the experiment to minimise tree movement. Eye-screws were used in preference to bands for attachment of the guy wires to avoid constricting tree growth. Previous experience in a study by Blackburn [24] showed that eye-screws only had a very localised direct growth effect on Sitka spruce trees; i.e., only a minor change in radial growth for a few millimetres up and down the stem from the point of attachment.
Following treatment, the mean distance from experimental trees to the four closest neighbouring trees (stem centre to stem centre) was 1.7 m in the “Control” treatment (equivalent to approximately 3460 stems ha−1), and 3.4 m in the “Thinned” and “Thinned and Guyed” treatments (equivalent to approximately 860 stems ha−1). At the time of treatment, trees had a mean height of 5.3 m and mean DBH (stem diameter measured 1.3 m above ground level) of 7.5 cm. Applying the relationship from Gardiner et al. [5] for Sitka spruce trees grown in Kershope forest, we estimate that the wind loading on the “Thinned” and “Thinned and guyed” treatment trees was approximately double that of the loading on “Control” treatment trees.
2.2. Climate Data
A 3-cup switching cup anemometer with wind vane (A100R and W200G, Vector Instruments, Rhyl, North Wales, UK) was installed on a 10 m tall mast situated on open ground 100 m to the east of the experiment site, and wind data were recorded for the four years of the experiment. The anemometer gave 1 count for a passage of 0.8 m of air, with a threshold of 0.2 m s−1 and a maximum speed of >75 m s−1 and an accuracy of ±1% between 10 and 55 m s−1. The wind vane had a resolution of 22.5° ± 3° with a low-speed threshold of 0.3 m s−1. The outputs from the anemometer and wind vane were logged every 8 s and stored every 30 min in a data logger (HAL01, Holtech Associates, Harwood-in-Teesdale, County Durham, UK) as frequency distributions. These data were analysed using Oriana 2.0 software (KCS, Pentraeth, UK) to provide wind speed frequency distribution, a wind rose, and the mean and median wind direction. Total annual rainfall and mean air temperature data were taken from the closest UK Met Office meteorological station 30 km to the north-east, at Eskdalemuir (55°18′44″ N, 3°12′22″ W). This station has similar elevation and aspect to the experiment site.
2.3. Assessment and Analysis
Tree heights (m) and DBH (cm) were measured when the treatments were applied and then in February of each of the next two years. All experimental trees were harvested four growing seasons after application of the treatments. Measurements at harvest included tree height (m), and total fresh (undried) branch mass including needles (kg). Cross-sectional sample “discs” were cut from the stem at four “heights”, corresponding with 0.1 m, 1.3 m, 3.0 m and 4.5 m above ground level. Tree stumps of the experimental trees from one block (i.e., six trees per treatment) were extracted by lifting vertically with a hand winch suspended under an aluminium scaffold pole tripod, and roots were cleaned using hand tools. Tree stumps were not extracted from the second block due to operational constraints. Cross-sectional samples were cut from the 10 largest structural roots of each tree at 0.25 m, 0.50 m and 0.75 m horizontal distance from the stump centre. Annual growth rings were measured on stem and root cross-sections using the WinDendro software (Régent Instruments, Chicoutimi, Quebec, Canada) [29]. These were measured on each stem section on four radii corresponding with the north, south, east and west sides of the tree. Growth rings of roots were measured from the upper-side outer ring to the anatomical centre of the root. Stem growth ring measurements from the four directions were averaged for each section, and root growth ring measurements were averaged for all sections from each growth year and for each distance from the stem centre. Relative growth responses were calculated as follows:
where is mean growth of the two years before treatment, and is growth during the year i after treatment. Radial growth after treatment was analysed as repeated measures series [6,30]. As root excavation was possible for only one block (i.e., one plot per treatment), root radial growth measurements were included for non-statistical comparison only. For each stem section height (0.1, 1.3, 3.0, 4.5), we modelled the above-ground measurements of growth, looking at the difference and pairwise comparisons between “Control”, “Guyed”, and “Thinned and Guyed” treatments for years 1996, 1997, 1998 and 1999. Analysis was performed at the tree level, and used a linear mixed model with fixed treatment and year effects plus their interaction, with random block and plot effects. For the repeated measurements of growth, we used an autoregressive method that assumes a linear dependence on previous values plus a stochastic term, with an unstructured correlation matrix to allow separate correlations between individual years. The model was fitted using procedure MIXED in the SAS statistical software (version 13.1, SAS Institute Inc. Cary, NC, USA) [31].
3. Results
3.1. Climate
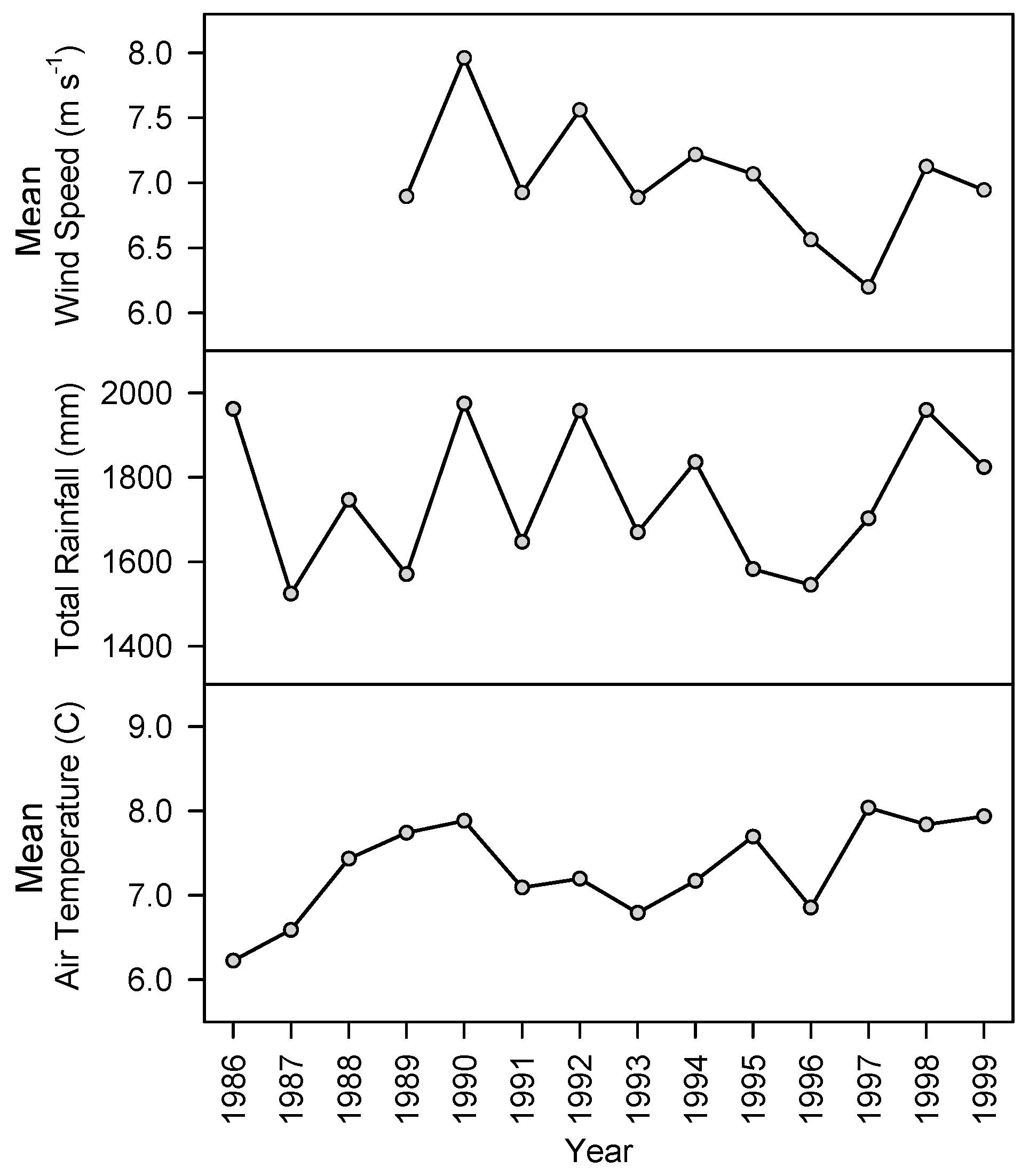
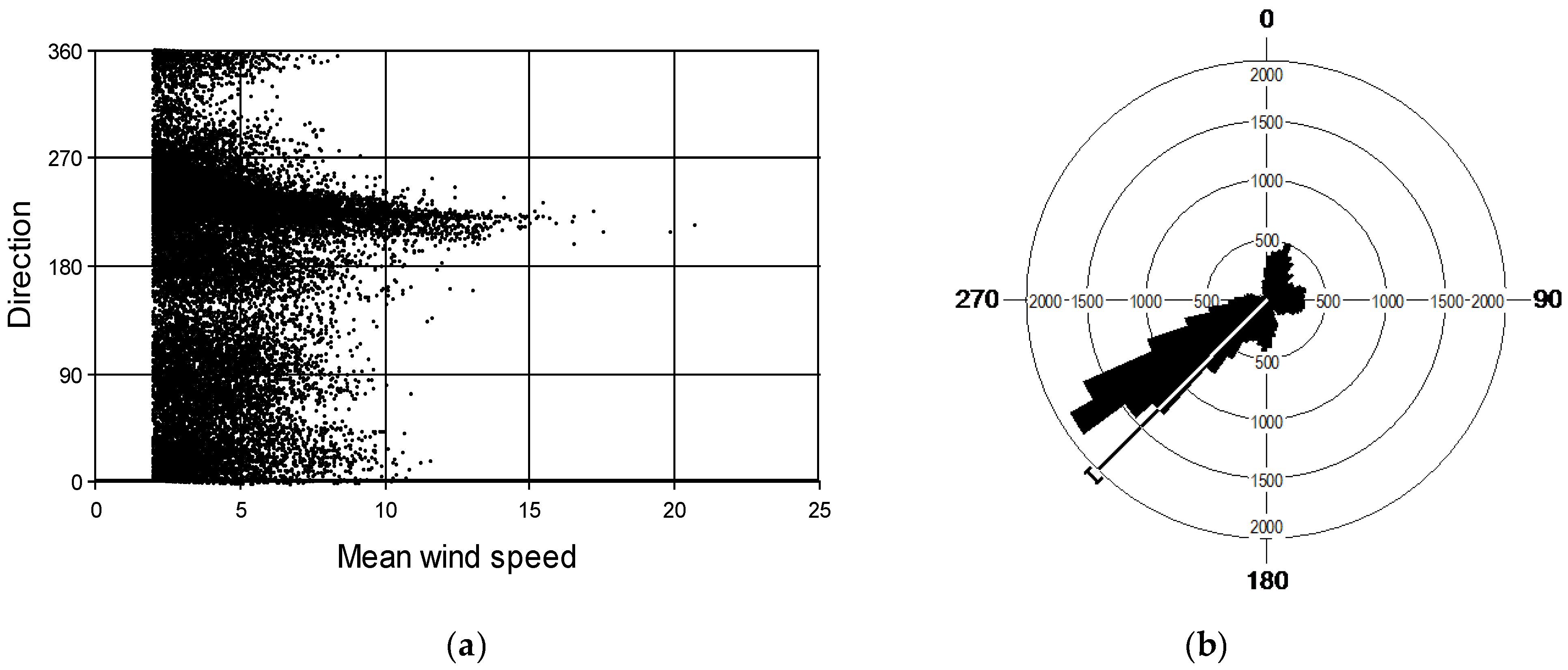
Annual rainfall during the life of the trees in the experiment was between 1500 and 2000 mm and annual mean air temperature was between 6 °C and 8 °C (Figure 2). Annual mean wind speed at the experimental site was between 6 m s−1 and 8 m s−1. Mean wind direction between 2 February 1996 and 3 April 2000 was from the south-west; mean 225°, median 234°, standard error of the mean 0.8° (Figure 3).
3.2. Tree Height
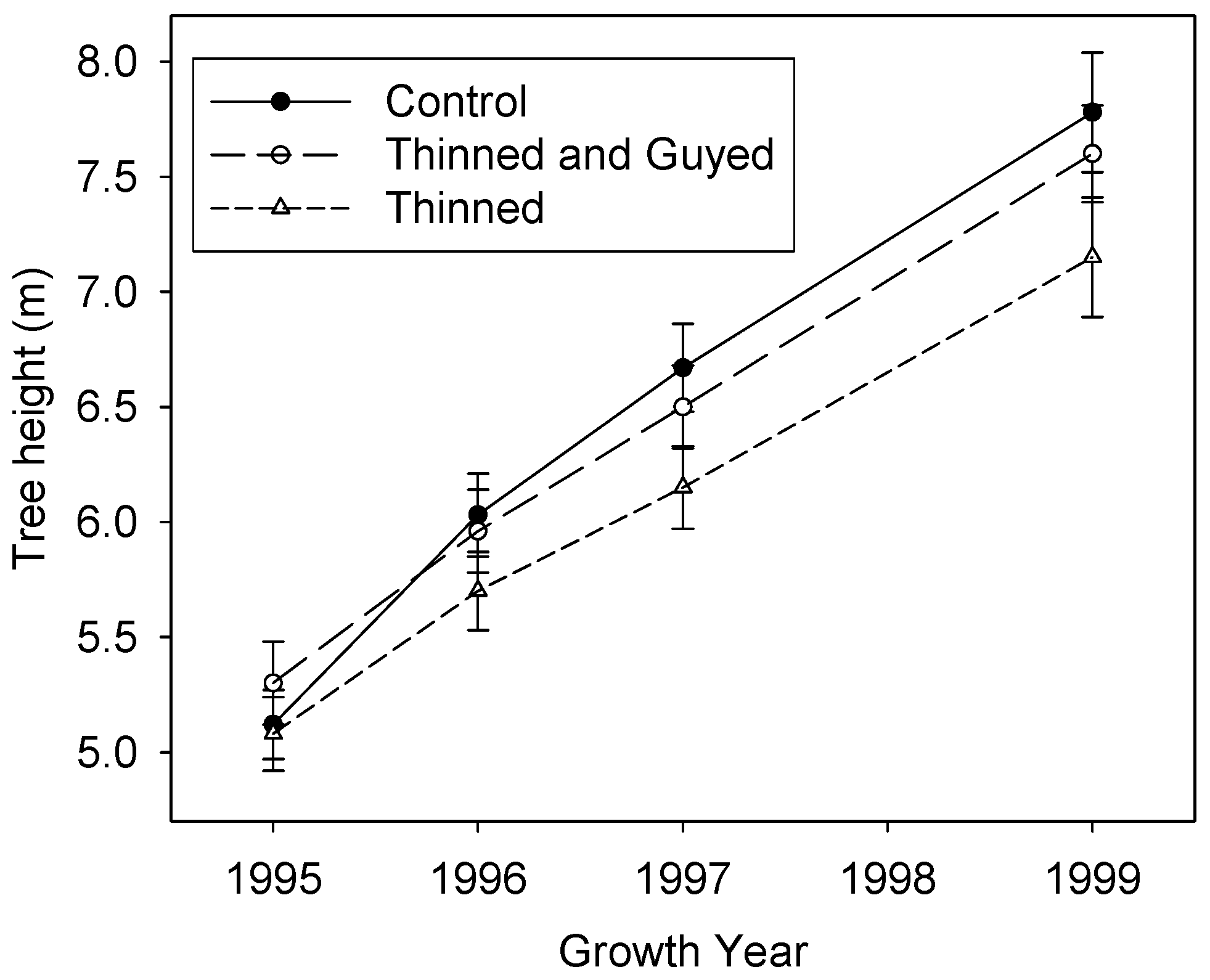
By the end of the experiment (Figure 4), “Control” trees were the tallest (mean 7.78 m, s.e. 0.26), with “Thinned and Guyed” trees slightly smaller (mean 7.60 m, s.e. 0.21), and “Thinned” trees were the shortest (mean 7.15 m, s.e. 0.26). Control trees grew on average 28% more in height than Thinned trees in the four years of the experiment but differences were not significant at the p < 0.05 level.
3.3. Analysis of Radial Growth
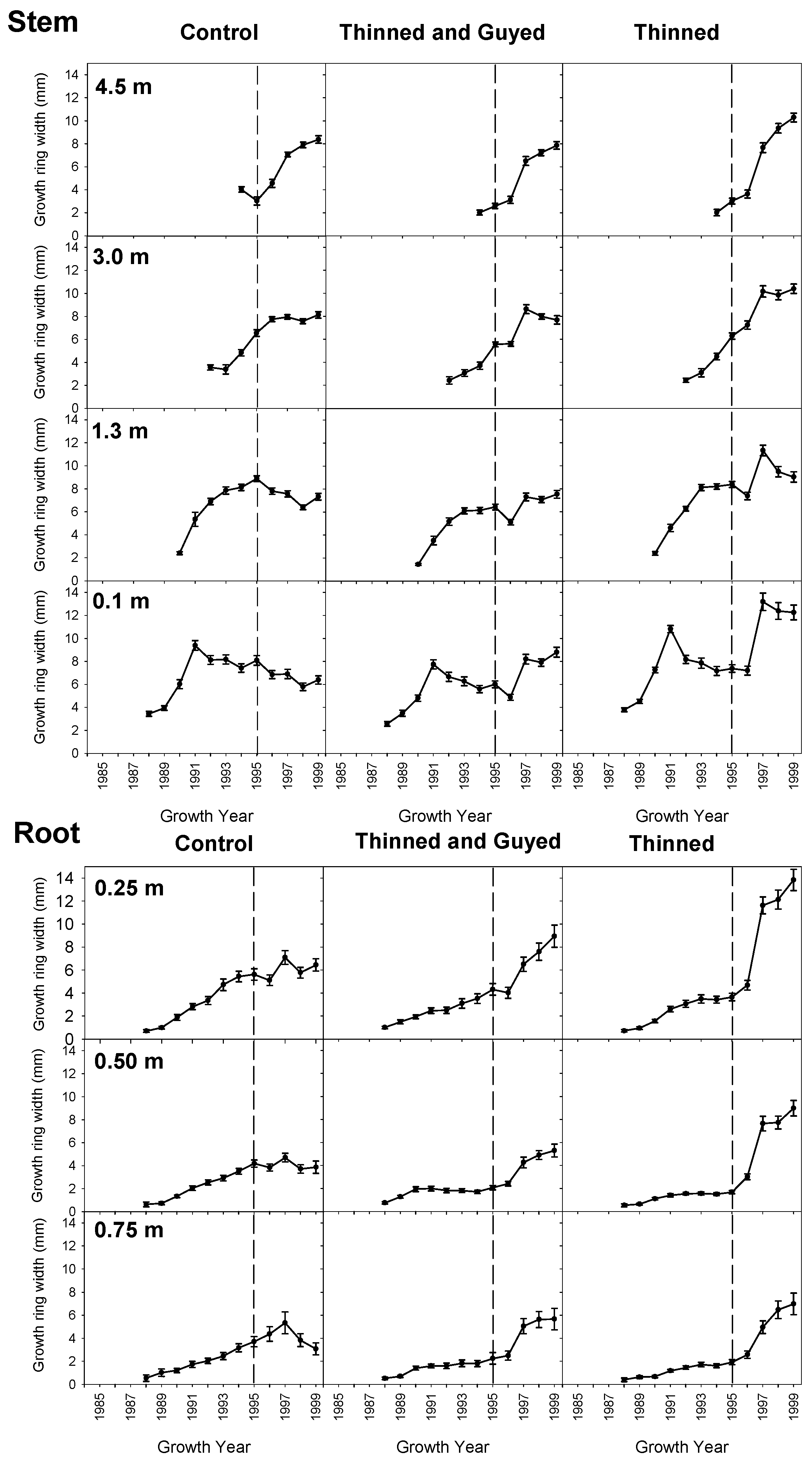
All trees showed a rapid increase in radial growth at the stem base (0.1m) for the first four years of growth after planting on site, with a growth peak in 1991, followed by a slow decline in growth rate before treatments were applied (Figure 5). In the untreated “Control trees”, the initial peak in stem radial growth was progressively later at increasing heights on the stem; 1991 at 0.1 m, 1995 at 1.3 m, 1997 at 3.0 m. Roots in the “Control” treatment showed a peak of radial growth simultaneously in 1997 at all distances from the stem (0.25, 0.50 and 0.75 m).
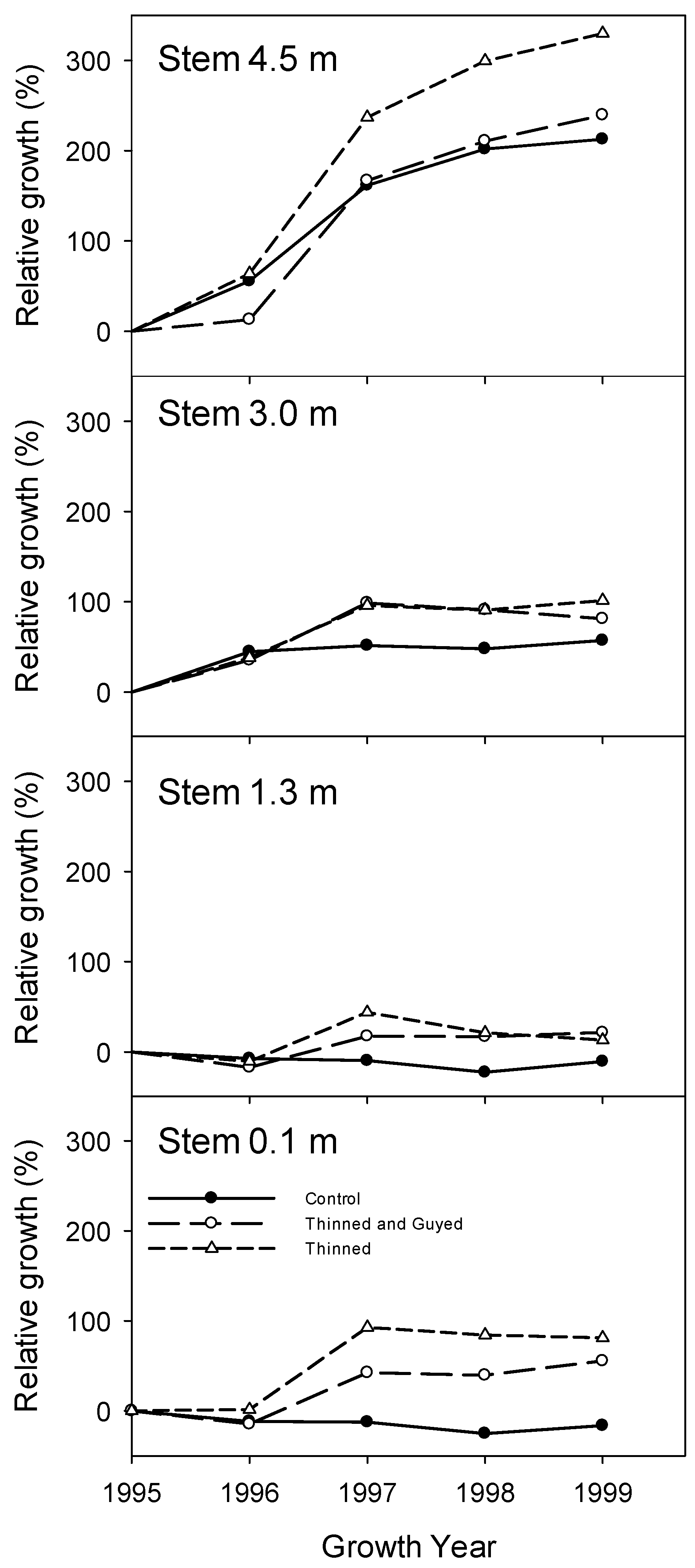
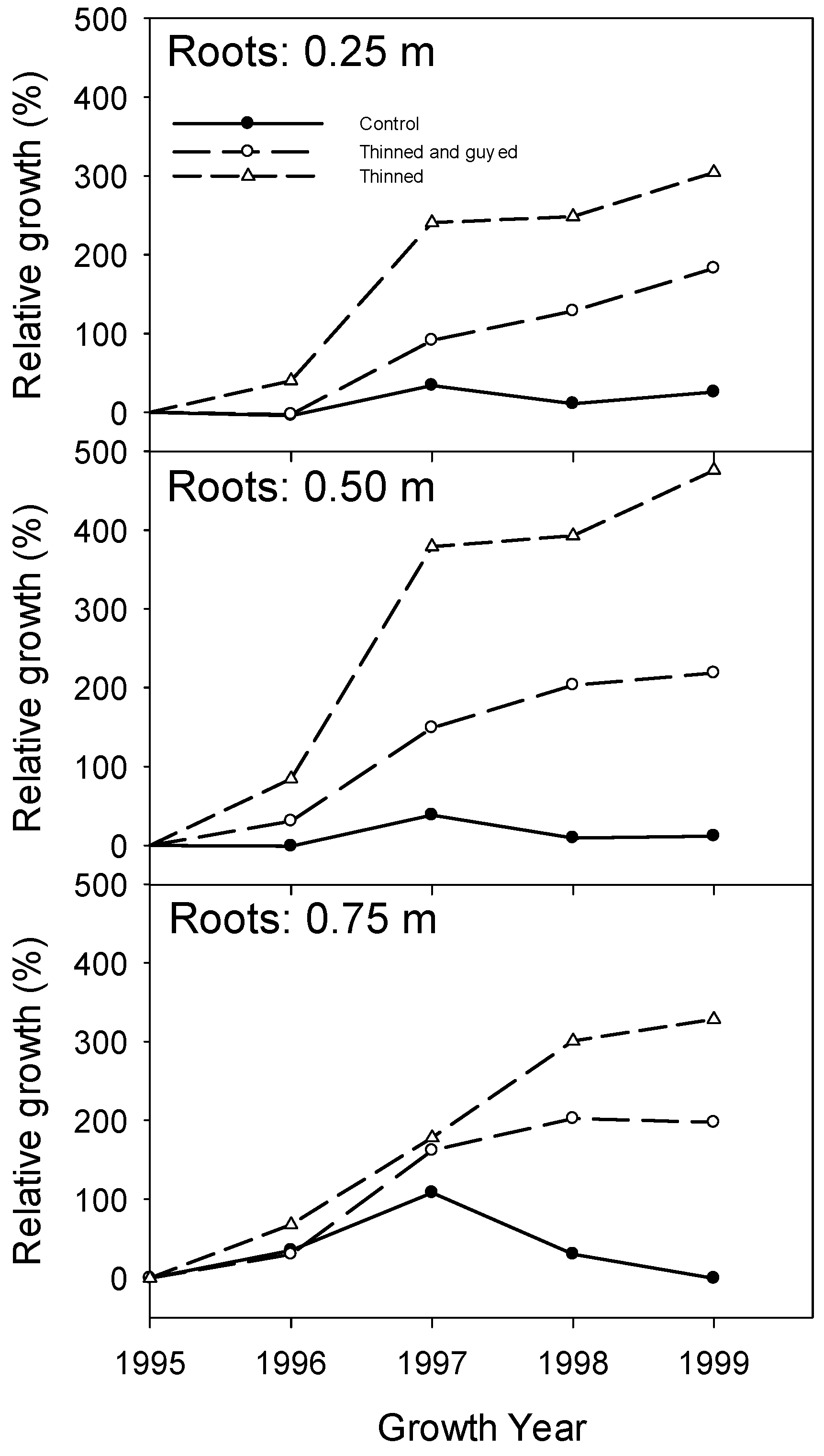
In the first growing season following treatment, there was a small decrease of stem radial growth at 0.1 m and 1.3 m in all treatments (Figure 5). However, in the second growing season following treatment, trees in both the “Thinned”, and “Thinned and Guyed” treatments showed large increases in stem-relative growth at 0.1 and 1.3 m above ground (Figure 6), and this corresponded with an observed increase in root growth at all distances along the root (Figure 7). The differences were greatest at 0.1 m height on the stem, and “Thinned” treatment trees showed a considerably greater growth response after treatment (93% increase) than ‘Thinned and Guyed’ treatment trees (42% increase) (Figure 6). Differences between treatments in relative growth rate were largely maintained during the 1997, 1998 and 1999 growth years, and these differences were significant at 0.1 m (p = 0.0023). Differences at other heights on the stem were not significant.
3.4. Branch Mass
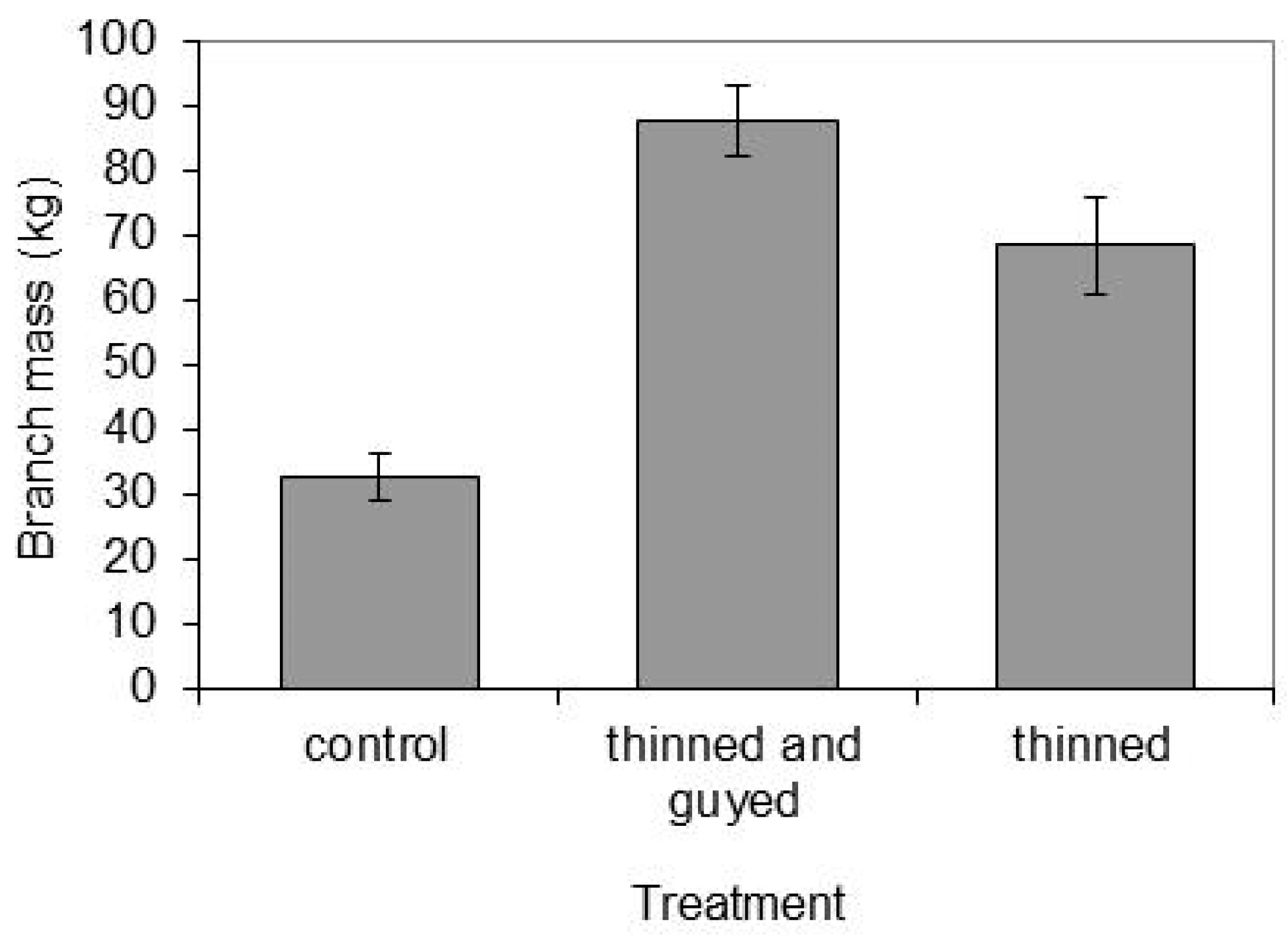
At the end of the experiment, fresh branch mass, including needles, varied significantly (p < 0.001) between treatments with trees in the “Control” treatment having the smallest branch mass, and trees in the “Thinned and Guyed” treatment having the greatest branch mass (Figure 8). The ”Thinned and Guyed“ trees (mean 87.7 kg, standard error 5.6) had on average >19 kg more branch mass than the ”Thinned“ treatment (mean 68.5 kg, standard error 7.5), which had more than twice the branch mass of the ”Control“ treatment (mean 32.6 kg, standard error 3.6) (Figure 8).
4. Discussion
Our original hypotheses appear to have been largely confirmed. The “Thinned” treatment responded with greater increased growth in the lower parts of the stem, in comparison to both the “Control” and the “Thinned and guyed” treatment. The “Thinned and guyed” treatment also responded with increased lower stem growth relative to the “Control” treatment, but considerably less than occurred in the “Thinned” treatment, making the increase in radial growth more even up the stem.
Despite little response in the growth season directly following the thinning treatment, there was a strong response to treatments in the second year. A similar strong response in the second growing season following loading of branches in Scots pine during the dormant winter period was found by Lunqvist and Valinger [32]. Other authors have found comparable delays in response to thinning treatments, with the duration of the delay varying between root and shoot. Urban et al. [9] used dendrochronological techniques to examine the response of 120-year-old white spruce (Picea glauca (Moench) Voss) to increased light and wind stress following removal of trees in a road clearing operation. Their study showed that the ‘release’ of stem growth (at 1.3 m) was delayed by between 3 and 9 years, while release of root growth was immediate. Therefore, trees appear to initially allocate photosynthates for below-ground growth at the expense of above-ground growth. Ruel et al. [6] found a similar immediate response in structural roots of 17- and 22-year-old balsam fir (Abies balsamea (L.) Mill.) to stand thinning, with a significant increase in stem growth (at 1.3 m) only after 2 years following thinning. The below-ground response described by Ruel et al. [6] was in the first full growing season following the year of treatment, and as the thinning treatment was applied in the summer, the response is comparable to the present experiment where the largest response followed the first dormancy period (autumn and winter) after treatment. Our results are also similar to those of Kneeshaw et al. [8] who observed a root radial growth response in Douglas fir (Pseudotsuga menziesii (Mirb.) Franco) and lodgepole pine (Pinus contorta Dougl.) trees in the year following stand thinning; a one-year delay before a stem radial growth response, and two- to three-year growth reductions in branch radial growth and leader height growth. An explanation for the variation in timing of response between these studies may be that to re-establish adequate root anchorage, larger thinned trees would have to allocate the greatest proportion of available carbohydrates below-ground for longer periods of time than smaller trees. Alternatively, the variation might result from the relative difference in wind exposure between the different experiments, with stem growth delay being longest in trees with the largest relative change in exposure.
The minimal response in the first growing season following thinning is explainable by the timing of needle formation. As the next cohort of new needles would have been pre-formed in buds before the February treatment in our study [33], there would be little opportunity to increase leaf area in the first year. Thinning would have provided an immediate benefit from an increase in available light in the first year, but the effect would be small compared to the effect of an increase in photosynthetic area in subsequent years [34]. A requirement for increased photosynthate supply to the most mechanically stressed parts of the tree would be expected to lead to reduced shoot extension and reduced radial growth of the less stressed parts of the tree [35,36] until subsequent years when photosynthetic area increases.
Radial growth was considerably greater in the “Thinned” than in the “Thinned and Guyed” treatment both in the lower stem and the roots. As trees in these two treatments would be expected to have the same amount of photosynthate available for cambial growth, the greater growth in trees that experienced increased mechanical strain requires explanation. A possible explanation for some of this difference is that the density of wood in the new growth rings might have been reduced in trees that experienced increased mechanical stress. Koga et al. [37] examined the effects of stand thinning on balsam fir (Abies balsamea L. Mill.). They found an immediate growth response to increased movement in the lower stem corresponding with an increase in earlywood rather than latewood. As earlywood has lower density than latewood, the resulting smaller latewood percentage gave lower ring densities. Therefore, the tree may be able to increase its overall growth response to mechanical loading to some extent by delaying its switch from earlywood to latewood to later in the growth season [35]. However, an explanation that may better fit with the large growth response and our whole-tree observations, is that more photosynthate was made available to stem and roots as a result of reduced height growth [7] and the considerably reduced allocation to branch growth (Figure 8) in the “Thinned” treatment compared to the ‘Thinned and Guyed’ treatment.
Wind not only increases mechanical loading on the tree’s component parts but also decreases humidity around the leaves [38]. The tree can respond to this by increasing leaf diffusive resistance or decreasing stomatal conductance, which in turn decreases photosynthetic rate and growth [38]. However, on the exposed upland site described here, “Thinned” trees that were allowed to move in the wind and were estimated to have experienced almost double the maximum bending moment [5], increased their radial growth faster than those that were restrained by guy wires. Therefore, any potential growth reduction effect from increased wind movement around needles was more than offset by increased growth in response to improved light availability, and the need to allocate growth to stabilise the tree and to transport more water to the developing crown [21,22].
In the “Thinned and Guyed” treatment, we speculate that the increase in radial growth is partly a response to the increased water requirements of a larger crown [39]. Whereas the ‘Thinned’ trees had to account for the changing hydraulic demands of an increasing photosynthetic area and increased mechanical strain, the ”Thinned and guyed“ trees were only required to adjust stem radial growth for changed hydraulic requirements [22] and therefore could allocate more available photosynthate to developing their crowns.
Peltola et al. [10] reported that the fastest diameter growth of Scots pine was close to the stem base for a period between seven and nine years after thinning. Similarly, Koga et al. [37] found that balsam fir showed a positive increase in stem base radial growth that lasted for seven years after stand thinning. Kneeshaw et al. [8] found that stem and root radial increment peaked four years after thinning of Douglas fir and lodgepole pine stands. None of the previous studies included a comparison of trees in a thinned treatment with trees in a similarly thinned, but mechanically restrained, control treatment. Our study advances understanding of the growth responses to both the increased light and the increased mechanical strain that trees experience following thinning treatments, and the interactions between these responses. To better understand the above- and below-ground acclimation of trees to changed wind exposure, further controlled experiments, similar to the one presented here, should be conducted on a range of species and monitored for longer time periods. However, the results of our experiment reinforce and highlight the importance of incorporating biomechanical acclimation of trees into forest growth and wind risk models [19,36,40].
5. Conclusions
Forest trees respond to increased light availability after stand thinning with large increases in stem cambial growth in the second and subsequent growth years following treatment. Our experiment showed considerably greater growth of the lower stem of thinned trees when they were allowed to move freely in the wind compared to when their movement was restricted by guying. Observations indicated that this difference was mirrored below ground with faster thickening of structural roots in thinned trees allowed to move freely. The acclimation of trees to wind movement through increased cambial growth in stems and roots appeared to rely on photosynthates that would otherwise have been allocated to branch growth. These growth responses would allow trees to rapidly improve their stability following an increase in wind exposure. Our results indicate that the substantial wind acclimation effects on tree growth and development should be incorporated into future revisions of forest growth and wind risk models.
Author Contributions
B.C.N. and B.A.G. were responsible for in the establishment of the experiment, collection, capture and analysis of the data, and writing of the manuscript. T.C. was responsible for statistical analysis and contributed to writing the manuscript.
Funding
This research was funded by the GB Forestry Commission and by the European Commission as part of the “TreeArch” project (AIR3-CT93-1269).
Acknowledgments
We thank Forestry Commission England for permission to run this experiment in Kershope Forest. We are grateful to M. P. Coutts for initial discussions on the experimental concept, to D. Ray for making part of his experimental area available, to C. N. Nielson for his leadership of the EC TreeArch project, and to D. Moore for inspiration during the fieldwork. Thank you also to the staff of Forest Research Technical Support Unit for monitoring the experiment, and to Shaun Mochan and Joanne Forbes for their help with the harvesting and measuring of trees.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kramer, H. Crown development in conifer stands in Scotland as influenced by initial spacing and subsequent thinning treatment. Forestry 1966, 39, 40–58. [Google Scholar] [CrossRef]
- Valinger, E.; Elfving, B.; Mörling, T. Twelve-year growth response of Scots pine to thinning and nitrogen fertilisation. For. Ecol. Manag. 2000, 134, 45–53. [Google Scholar] [CrossRef]
- Olivar, J.; Bogino, S.; Rathgeber, C.; Bonnesoeur, V.; Bravo, F. Thinning has a positive effect on growth dynamics and growth–climate relationships in Aleppo pine (Pinus halepensis) trees of different crown classes. Ann. For. Sci. 2014, 71, 395–404. [Google Scholar] [CrossRef]
- Hale, S.; Gardiner, B.A.; Wellpott, A.; Nicoll, B.C.; Achim, A. Wind loading of trees: Influence of tree size and competition. Eur. J. For. Res. 2012, 131, 203–217. [Google Scholar] [CrossRef]
- Gardiner, B.A.; Stacey, G.R.; Belcher, R.E.; Wood, C.J. Field and wind tunnel assessments of the implications of respacing and thinning for tree stability. Forestry 1997, 70, 233–252. [Google Scholar] [CrossRef] [Green Version]
- Ruel, J.-C.; Larouche, C.; Achim, A. Changes in root morphology after precommercial thinning in balsam fir stands. Can. J. For. Res. 2003, 33, 2452–2459. [Google Scholar] [CrossRef]
- Mitchell, S. Stem growth responses in Douglas-fir and Sitka spruce following thinning: Implications for assessing wind-firmness. For. Ecol. Manag. 2000, 135, 105–114. [Google Scholar] [CrossRef]
- Kneeshaw, D.D.; Williams, H.; Nikinmaa, E.; Messier, C. Patterns of above- and below-ground response of understory conifer release 6 years after partial cutting. Can. J. For. Res. 2002, 32, 255–265. [Google Scholar] [CrossRef] [Green Version]
- Urban, S.T.; Lieffers, V.J.; MacDonald, S.E. Release in radial growth in the trunk and structural roots of white spruce as measured by dendrochronology. Can. J. For. Res. 1994, 24, 1550–1556. [Google Scholar] [CrossRef]
- Peltola, H.; Miina, J.; Rouvinen, I.; Kellomäki, S. Effect of early thinning on the diameter growth distribution along the stem of Scots pine. Silva Fenn. 2002, 36, 813–825. [Google Scholar] [CrossRef]
- Nicoll, B.C.; Ray, D. Adaptive growth of tree root systems in response to wind action and site conditions. Tree Physiol. 1996, 16, 899–904. [Google Scholar] [CrossRef]
- Stokes, A.; Nicoll, B.C.; Coutts, M.P.; Fitter, A.H. Responses of young Sitka spruce clones to mechanical perturbation and nutrition: Effects on biomass allocation, root development, and resistance to bending. Can. J. For. Res. 1997, 27, 1049–1057. [Google Scholar] [CrossRef]
- Stokes, A.; Fitter, A.H.; Coutts, M.P. Responses of young trees to wind: Effects on root growth. In Wind and Trees; Coutts, M.P., Grace, J., Eds.; Cambridge University Press: Cambridge, UK, 1995; pp. 264–275. [Google Scholar]
- Fayle, D.C.F. Stem sway affects ring width and compression wood formation in exposed root bases. For. Sci. 1976, 22, 193–194. [Google Scholar]
- Fayle, D.C.F. Radial Growth in Tree Roots. Distribution, Timing, Anatomy; Tech. Rep. No. 9; Faculty of Forestry, University of Toronto: Toronto, ON, Canada, 1968; p. 183. [Google Scholar]
- Jacobs, M.R. The effect of wind sway on the form and development of Pinus radiata D. Don. Aust. J. Bot. 1954, 2, 35–51. [Google Scholar] [CrossRef]
- Wilson, B.F. Distribution of secondary thickening in tree root systems. In The Development and Function of Roots; Torrey, J.G., Clarkson., D.T., Eds.; Academic Press: New York, NY, USA, 1975; pp. 197–219. [Google Scholar]
- Niez, B.; Dlouha, J.; Moulia, B.; Badel, E. Water-stressed or not, the mechanical acclimation is a priority requirement for trees. Trees 2018, 33, 279–291. [Google Scholar] [CrossRef]
- Mitchell, S.J. Wind as a natural disturbance agent in forests: A synthesis. Forestry 2013, 86, 147–157. [Google Scholar] [CrossRef]
- Nicoll, B.C.; Gardiner, B.A.; Peace, A.J. Improvements in anchorage provided by the acclimation of forest trees to wind stress. Forestry 2008, 81, 389–398. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Silins, U.; Lieffers, V.J.; Man, R. Stem hydraulic properties and growth in lodgepole pine stands following thinning and sway treatment. Can. J. For. Res. 2003, 33, 1295–1303. [Google Scholar] [CrossRef]
- Badel, E.; Ewers, F.W.; Cochard, H.; Telewski, F.W. Acclimation of mechanical and hydraulic functions in trees: Impact of the thigmomorphogenetic process. Front. Plant Sci. 2015, 6, 1–12. [Google Scholar] [CrossRef]
- Wood, C.J. Understanding wind forces on trees. In Wind and Trees; Coutts, M.P., Grace, J., Eds.; Cambridge University Press: Cambridge, UK, 1995; pp. 133–164. [Google Scholar]
- Blackburn, G.R.A. The Growth and Mechanical Response of Trees to Wind Loading. Ph.D Thesis, School of Biological Sciences, University of Manchester, Manchester, UK, 1997. [Google Scholar]
- Quine, C.P.; White, I.M.S. Revised Windiness Scores for the Windthrow Hazard Classification: The Revised Scoring Method; Forestry Commission: Farnham, UK, 1993; Volume 230. [Google Scholar]
- Kennedy, F. The Identification of Soils for Forest Management. Forestry Commission Field Guide; Forestry Commission: Edinburgh, UK, 2002; p. 56. [Google Scholar]
- Avery, B.W. Soils of the British Isles; CAB International: Wallingford, UK, 1990; p. 463. [Google Scholar]
- Aldhous, J.R. Nursery Practice. Forestry Commission Bulletin 43; HMSO: London, UK, 1972. [Google Scholar]
- Guay, R.; Gagnon, R.; Morin, H. A new automatic and interactive tree-ring measurement system based on a line scan camera. For. Chron. 1992, 68, 138–141. [Google Scholar] [CrossRef]
- Meredith, M.P.; Stehman, S.V. Repeated measures experiments in forestry:focus on analysis of response curves. Can. J. For. Res. 1991, 21, 957–965. [Google Scholar] [CrossRef]
- SAS. SAS Institute Inc. SAS/STAT® 13.1 User’s Guide: The MIXED Procedure (Chapter); SAS Institute Inc.: Cary, NC, USA, 2013. [Google Scholar]
- Lundqvist, L.; Valinger, E. Stem diameter growth of Scots pine trees after increased mechanical load in the crown during dormancy and (or) growth. Ann. Bot. 1996, 77, 59–62. [Google Scholar] [CrossRef]
- Cannell, M.G.R.; Willett, S.C. Rates and times at which needles are initiated in buds on differing provenances of Pinus contorta and Picea sitchensis in Scotland. Can. J. For. Res. 1975, 5, 367–380. [Google Scholar] [CrossRef]
- Ginn, S.E.; Seiler, J.R.; Cazell, B.H.; Kreh, R.E. Physiological and growth responses of eight-year-old Loblolly pine stands to thinning. For. Sci. 1991, 37, 1030–1040. [Google Scholar]
- Meng, S.X.; Lieffers, V.J.; Reid, D.E.; Rudnicki, M.; Silins, U.; Jin, M. Reducing stem bending increases the height growth of tall pines. J. Exp. Bot. 2006, 57, 3175–3182. [Google Scholar] [CrossRef] [Green Version]
- Bonnesoeur, V.; Constant, T.; Moulia, B.; Fournier, M. Forest trees filter chronic wind-signals to acclimate to high winds. New Phytol. 2016, 210, 850–860. [Google Scholar] [CrossRef] [Green Version]
- Koga, S.; Zhang, S.Y.; Bégin, J. Effects of precommercial thinning on annual radial growth and wood density in balsam fir (Abies balsamea). Wood Fiber Sci. 2002, 34, 625–642. [Google Scholar]
- Grace, J.; Malcolm, D.C.; Bradbury, I.K. The effect of wind and humidity on leaf diffusive resistance in Sitka spruce seedlings. J. Appl. Ecol. 1975, 12, 931–940. [Google Scholar] [CrossRef]
- Beauchamp, K.; Mencuccini, M.; Perks, M.; Gardiner, B. The regulation of sapwood area, water transport and heartwood formation in Sitka spruce. Plant Ecol. Divers. 2012, 6, 45–56. [Google Scholar] [CrossRef]
- Gardiner, B.; Byrne, K.; Hale, S.; Kamimura, K.; Peltola, H.; Rual, J.-C. A review of mechanistic modelling of wind damage risk to forests. Forestry 2008, 81, 447–463. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
The experimental site and treatments. From top: “Thinned” treatment; “Thinned and Guyed” treatment; Cables attached to a guyed tree.
Figure 1.
The experimental site and treatments. From top: “Thinned” treatment; “Thinned and Guyed” treatment; Cables attached to a guyed tree.

Figure 2.
Annual mean wind speed (m s−1) at the experiment site at Crookburn, Kershope, and total annual rainfall and mean air temperature at Eskdalemuir, for the duration of the tree growth periods. Eskdalemuir meteorological data supplied by the UK Met Office.
Figure 2.
Annual mean wind speed (m s−1) at the experiment site at Crookburn, Kershope, and total annual rainfall and mean air temperature at Eskdalemuir, for the duration of the tree growth periods. Eskdalemuir meteorological data supplied by the UK Met Office.

Figure 3.
(a) Wind speed (m s−1) frequency distribution and (b) wind rose for Crookburn, Kershope between February 1996 and April 2000. The line on the wind rose indicates the mean wind direction, with bars showing 95% confidence intervals.
Figure 3.
(a) Wind speed (m s−1) frequency distribution and (b) wind rose for Crookburn, Kershope between February 1996 and April 2000. The line on the wind rose indicates the mean wind direction, with bars showing 95% confidence intervals.

Figure 4.
Mean tree height (± standard error) at the end of 1995, 1996, 1997 and 1999 growth years.

Figure 5.
Mean (± standard error) radial growth in the stem (0.1 m, 1.3 m, 3.0 m and 4.5 m) in each of the treatments between the 1990 and 1999 growth years. Radial growth in structural roots from one plot (at 0.25 m, 0.50 m and 0.75 m from the tree centre) are shown for comparison. The dashed vertical lines mark the last growth year before treatment.
Figure 5.
Mean (± standard error) radial growth in the stem (0.1 m, 1.3 m, 3.0 m and 4.5 m) in each of the treatments between the 1990 and 1999 growth years. Radial growth in structural roots from one plot (at 0.25 m, 0.50 m and 0.75 m from the tree centre) are shown for comparison. The dashed vertical lines mark the last growth year before treatment.

Figure 6.
The relative (to size in 1995) diameter growth response following treatment, in stems at 0.1 m, 1.3 m, 3.0 m and 4.5 m above ground-level in growth years 1996, 1997, 1998 and 1999. Differences between treatments were significant at 0.1 m for 1997, 1998 and 1999 growth years (overall p = 0.004).
Figure 6.
The relative (to size in 1995) diameter growth response following treatment, in stems at 0.1 m, 1.3 m, 3.0 m and 4.5 m above ground-level in growth years 1996, 1997, 1998 and 1999. Differences between treatments were significant at 0.1 m for 1997, 1998 and 1999 growth years (overall p = 0.004).

Figure 7.
The relative diameter growth response following treatments, in structural roots at 0.25 m, 0.5 m and 0.75 m from the tree centre in growth years 1996, 1997, 1998 and 1999.
Figure 7.
The relative diameter growth response following treatments, in structural roots at 0.25 m, 0.5 m and 0.75 m from the tree centre in growth years 1996, 1997, 1998 and 1999.

Figure 8.
Branch mass at harvesting compared between treatments. Error bars are ±1 standard error. Differences between all treatments were significant at p < 0.001.
Figure 8.
Branch mass at harvesting compared between treatments. Error bars are ±1 standard error. Differences between all treatments were significant at p < 0.001.

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Nicoll, B.C.; Connolly, T.; Gardiner, B.A. Changes in Spruce Growth and Biomass Allocation Following Thinning and Guying Treatments. Forests 2019, 10, 253. https://doi.org/10.3390/f10030253
AMA Style
Nicoll BC, Connolly T, Gardiner BA. Changes in Spruce Growth and Biomass Allocation Following Thinning and Guying Treatments. Forests. 2019; 10(3):253. https://doi.org/10.3390/f10030253
Chicago/Turabian StyleNicoll, Bruce C., Thomas Connolly, and Barry A. Gardiner. 2019. "Changes in Spruce Growth and Biomass Allocation Following Thinning and Guying Treatments" Forests 10, no. 3: 253. https://doi.org/10.3390/f10030253
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.