
Reprogrammed CD8+ T-Cells Isolated from the Mouse Spleen Increase the Number of Immune Cells with Antitumor Activity and Decrease the Amount of Cancer Stem Cells †

 , , ,
, , ,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Lewis Lung Carcinoma Cell Line and Tumor Model
2.3. Isolation of Mononuclear Cells
2.4. Flow Cytometry
2.5. Magnetic Separation of CD8+ T-Cells
2.6. Reprogramming of Spleen CD8+ T-Cells
2.7. CD8+ T-Cells Injection
2.8. Detection of the CCR7 Expression, Cytotoxicity, and Apoptosis of rsCD8+T-cells in Vitro
2.9. Histology of the Lungs
2.10. Assessment of Tumor Growth
2.11. Assessment of Tumor Volume
2.12. Statistical Analysis
3. Results
3.1. Study of Detection of the CCR7 Expression, Cytotoxicity, and Apoptosis of rsCD8+T-Cells In Vitro
3.2. Migration of rsCD8+ T-Cells Isolated from Mouse Spleen into the Lungs of Mice with LLC
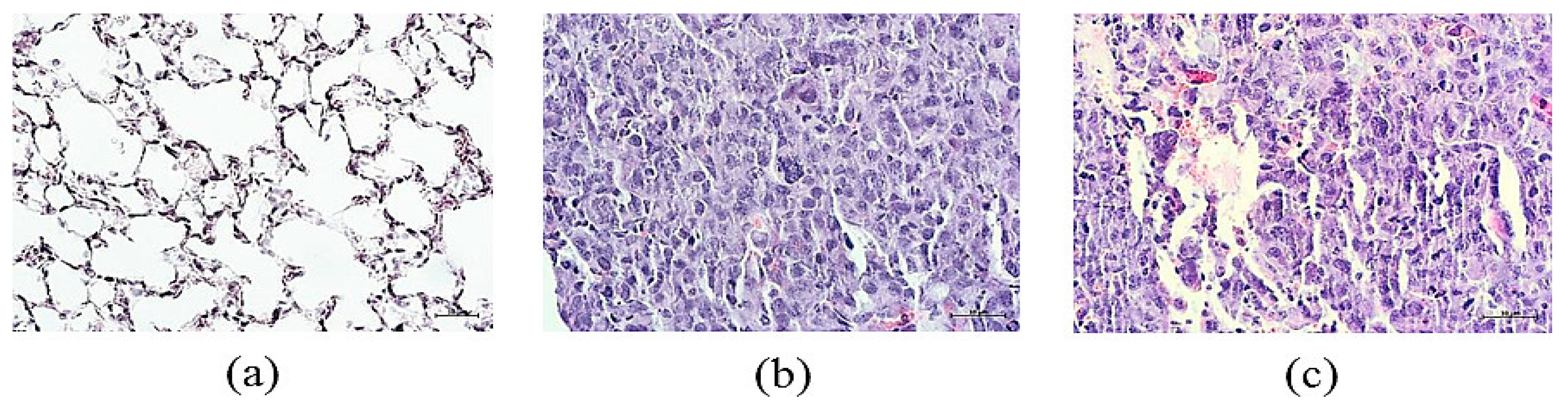
3.3. Lung Histology and Tumor Growth after Cell Therapy with rsCD8+ T-Cells
3.3.1. Lung Histology
3.3.2. Tumor Growth
3.4. The rsCD8+ T-Cells Change the Amount of Cancer Cells and Cancer Stem Cells in the Lungs and Blood of Tumor-Bearing Mice
3.5. Effect of rsCD8+ T-Cells on the Content of CD8+ T-Cells in the Lungs and Blood of Tumor-Bearing Mice
3.6. Effect of rsCD8+ T-Cells on the Content of CD4+ T-Cells in the Blood and Lungs of Tumor-Bearing Mice
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Stapelfeld, C.; Dammann, C.; Maser, E. Sex-specificity in lung cancer risk. Int. J. Cancer 2020, 146, 2376–2382. [Google Scholar] [CrossRef] [PubMed]
- Poncette, L.; Bluhm, J.; Blankenstein, T. The role of CD4 T cells in rejection of solid tumors. Curr. Opin. Immunol. 2022, 74, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Raskov, H.; Orhan, A.; Christensen, J.P.; Gögenur, I. Cytotoxic CD8+ T-cells in cancer and cancer immunotherapy. Br. J. Cancer. 2021, 124, 359–367. [Google Scholar] [CrossRef] [PubMed]
- Skurikhin, E.G.; Pershina, O.; Ermakova, N.; Pakhomova, A.; Widera, D.; Zhukova, M.; Pan, E.; Sandrikina, L.; Kogai, L.; Kushlinskii, N.; et al. Reprogrammed CD8+ T-Lymphocytes Isolated from Bone Marrow Have Anticancer Potential in Lung Cancer. Biomedicines. 2022, 10, 1450. [Google Scholar] [CrossRef] [PubMed]
- Skurikhin, E.G.; Pershina, O.; Ermakova, N.; Pakhomova, A.; Zhukova, M.; Pan, E.; Sandrikina, L.; Widera, D.; Kogai, L.; Kushlinskii, N.; et al. Cell Therapy with Human Reprogrammed CD8+ T-Cells Has Antimetastatic Effects on Lewis Lung Carcinoma in C57BL/6 Mice. Int. J. Mol. Sci. 2022, 23, 15780. [Google Scholar] [CrossRef] [PubMed]
- Cardiff, R.D.; Miller, C.H.; Munn, R.J. Manual hematoxylin and eosin staining of mouse tissue sections. Cold Spring Harb. Protoc. 2014, 2014, 655–658. [Google Scholar] [CrossRef] [PubMed]
- Hather, G.; Liu, R.; Bandi, S.; Mettetal, J.; Manfredi, M.; Shyu, W.C.; Donelan, J.; Chakravarty, A. Growth rate analysis and efficient experimental design for tumor xenograft studies. Cancer Inform. 2014, 13, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Tomayko, M.M.; Reynolds, C.P. Determination of subcutaneous tumor size in athymic (nude) mice. Cancer Chemother. Pharmacol. 1989, 24, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Czystowska, M.; Gooding, W.; Szczepanski, M.J.; Lopez-Abaitero, A.; Ferris, R.L.; Johnson, J.T.; Whiteside, T.L. The immune signature of CD8(+)CCR7(+) T cells in the peripheral circulation associates with disease recurrence in patients with HNSCC. Clin. Cancer Res. 2013, 19, 889–899. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Values/Parameters | Intact Control | LLC | rsCD8+ T-Cells |
|---|---|---|---|
| Tumor volume, mm3 | 0 | 4.72 ± 2.85 # | 2.09±0.09 #,* |
| The average number of metastases | 0 | 2.60 ± 0.35 #,1 | 0.17±0.17 #,* |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Skurikhin, E.G.; Pershina, O.; Zhukova, M.; Pakhomova, A.; Ermakova, N.; Widera, D.; Pan, E.; Sandrikina, L.; Kogai, L.; Kushlinskii, N.; et al. Reprogrammed CD8+ T-Cells Isolated from the Mouse Spleen Increase the Number of Immune Cells with Antitumor Activity and Decrease the Amount of Cancer Stem Cells. Med. Sci. Forum 2023, 21, 40. https://doi.org/10.3390/ECB2023-14132
Skurikhin EG, Pershina O, Zhukova M, Pakhomova A, Ermakova N, Widera D, Pan E, Sandrikina L, Kogai L, Kushlinskii N, et al. Reprogrammed CD8+ T-Cells Isolated from the Mouse Spleen Increase the Number of Immune Cells with Antitumor Activity and Decrease the Amount of Cancer Stem Cells. Medical Sciences Forum. 2023; 21(1):40. https://doi.org/10.3390/ECB2023-14132
Chicago/Turabian StyleSkurikhin, Evgenii G., Olga Pershina, Mariia Zhukova, Angelina Pakhomova, Natalia Ermakova, Darius Widera, Edgar Pan, Lubov Sandrikina, Lena Kogai, Nikolai Kushlinskii, and et al. 2023. "Reprogrammed CD8+ T-Cells Isolated from the Mouse Spleen Increase the Number of Immune Cells with Antitumor Activity and Decrease the Amount of Cancer Stem Cells" Medical Sciences Forum 21, no. 1: 40. https://doi.org/10.3390/ECB2023-14132