
Rhizosphere Soil Microbial Survival States and N-Related Process during Riparian Plant Dormancy: Influences of Plant Locations and Plant Species


Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
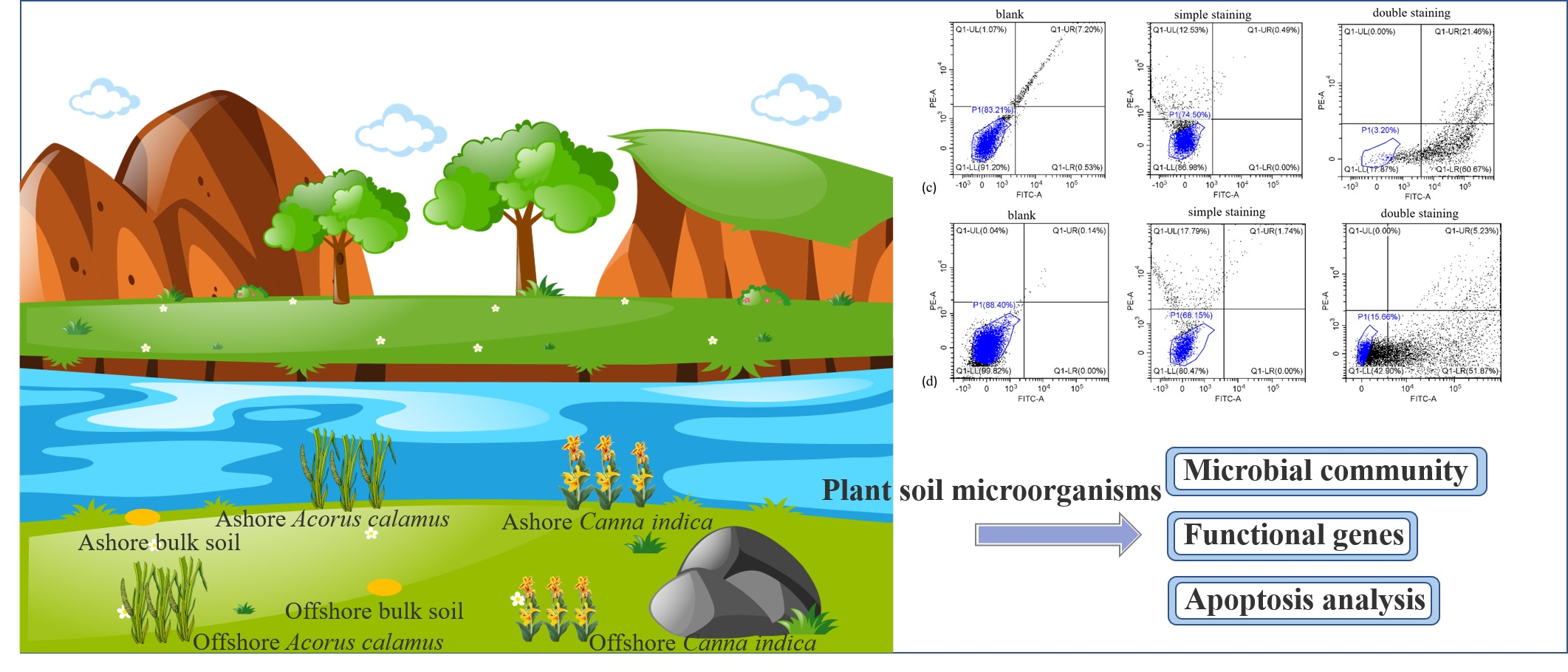
2.1. Study Area and Description
2.2. Sample Collection
2.3. Soil Physiochemical Analysis
2.4. DNA Extraction, Quantitative PCR, and Illumina MiSeq Sequencing
2.5. Flow Cytometry Analysis
2.6. Data Analyses
3. Results and Discussion
3.1. Soil Physiochemical Properties
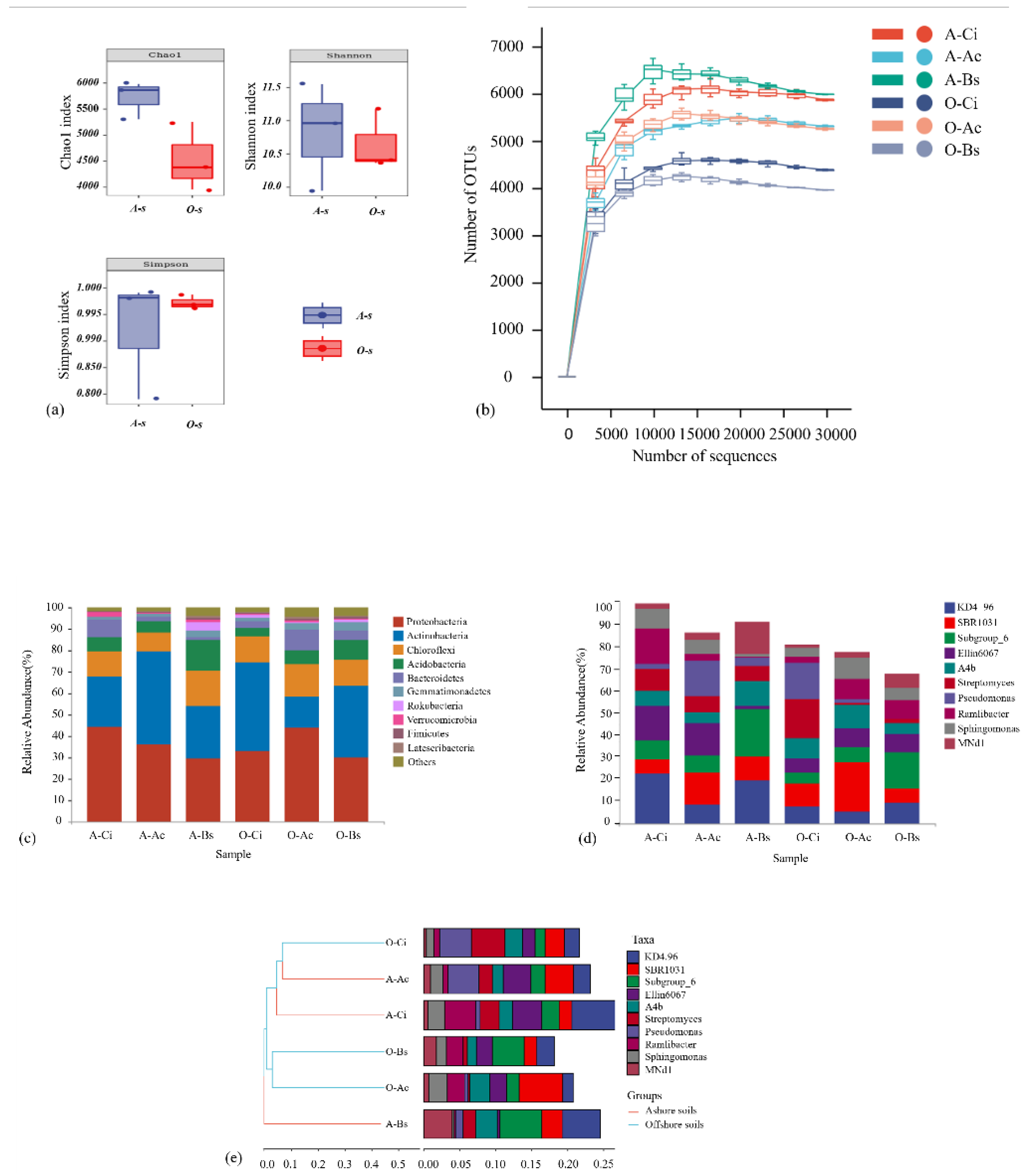
3.2. Soil Microbial Community Structures and Diversity
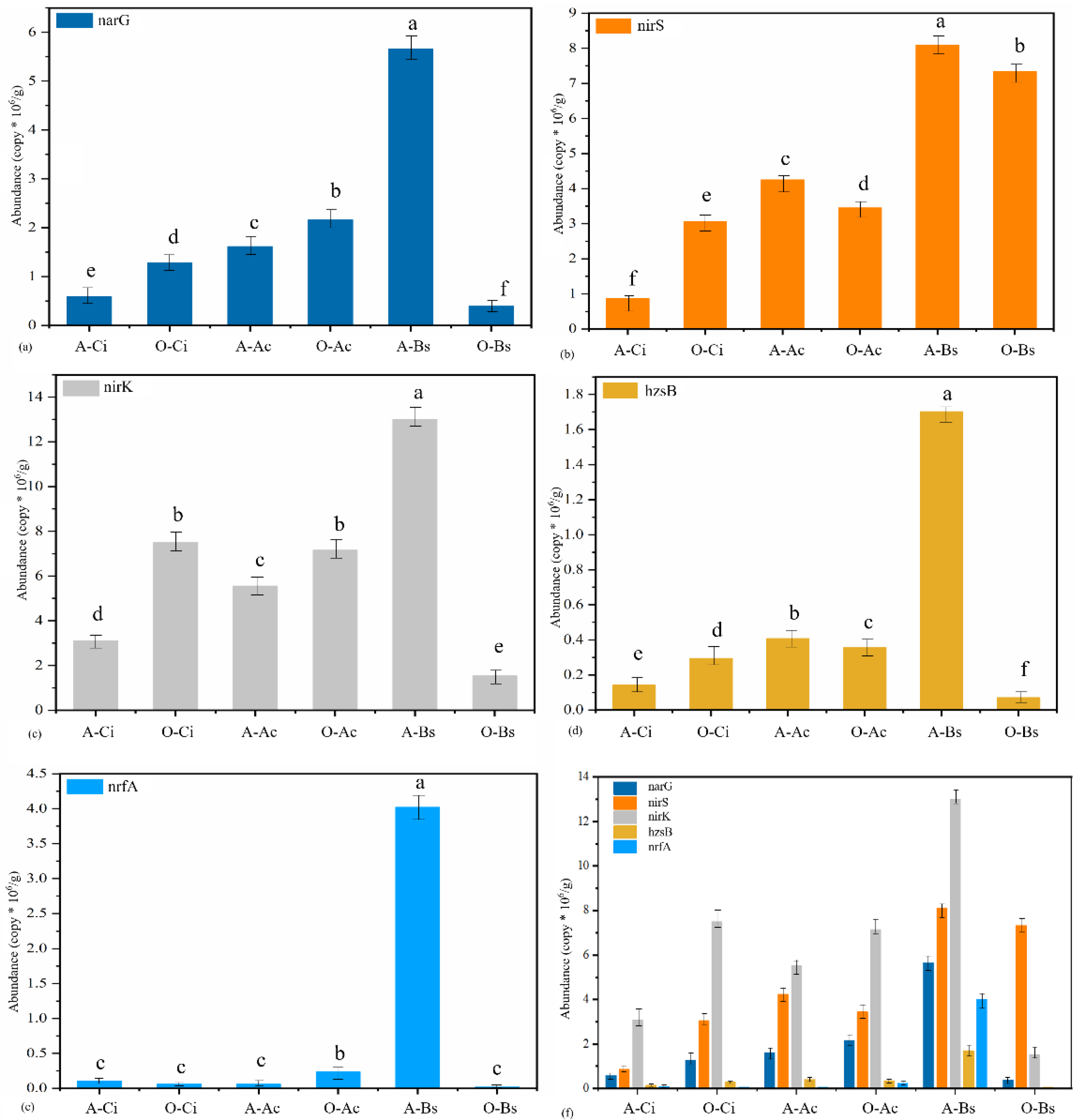
3.3. Functional Gene Abundance of N-Reduction Pathways
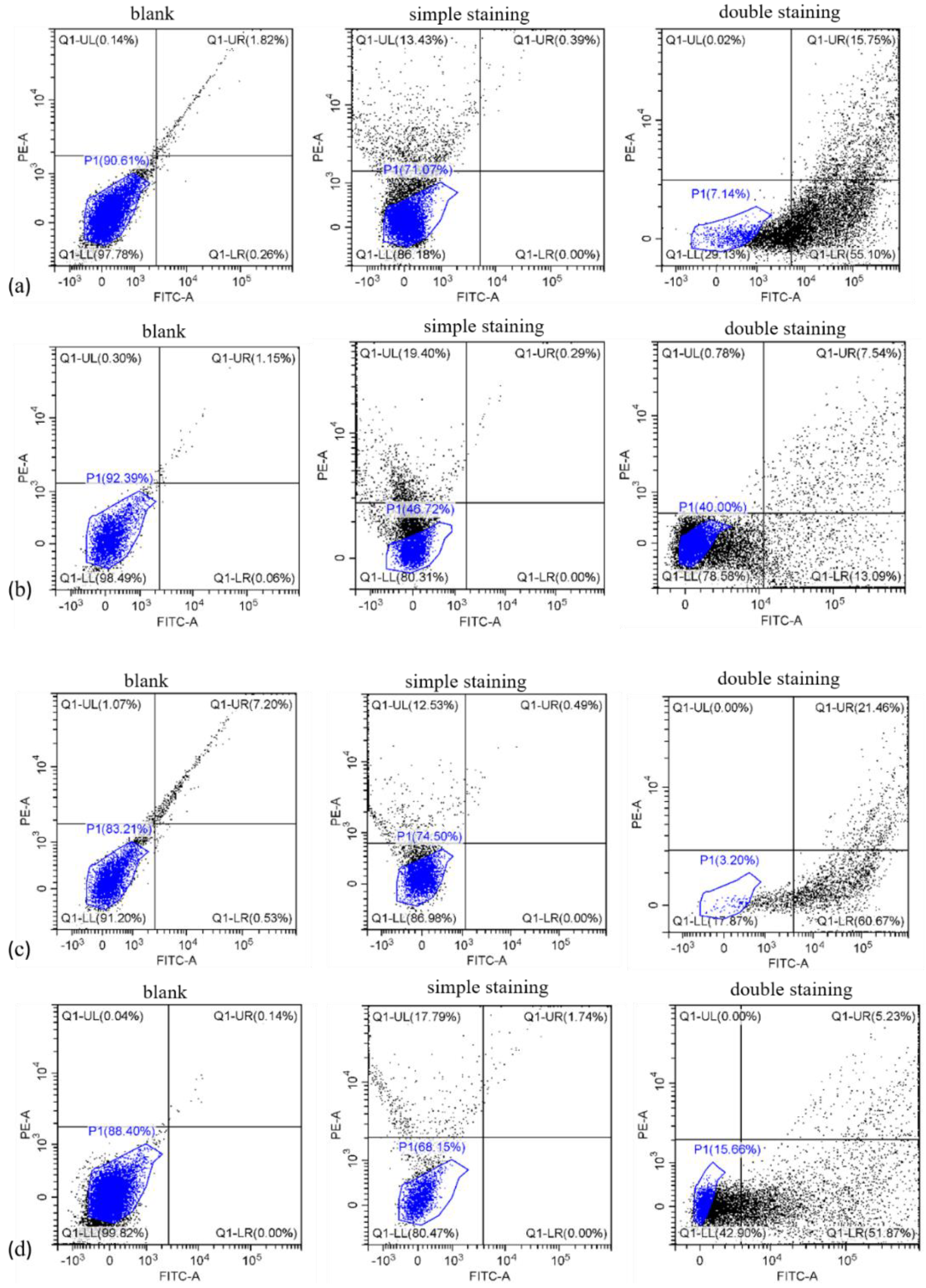
3.4. Cell Apoptosis Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nilsson, C.; Berggren, K. Alterations of riparian ecosystems caused by river regulation. Bioscience 2000, 50, 783–792. [Google Scholar] [CrossRef]
- Naiman, R.J.; Decamps, H. The ecology of interfaces: Riparian zones. Annu. Rev. Ecol. Syst. 1997, 28, 621–658. [Google Scholar] [CrossRef] [Green Version]
- Lyu, C.J.; Li, X.J.; Yuan, P.; Song, Y.H.; Gao, H.J.; Liu, X.L.; Liu, R.X.; Yu, H.B. Nitrogen retention effect of riparian zones in agricultural areas: A meta-analysis. J. Clean. Prod. 2021, 315, 128143. [Google Scholar] [CrossRef]
- Chen, F.X.; Lu, S.Y.; Hu, X.Z.; He, Q.; Feng, C.P.; Xu, Q.J.; Chen, N.; Ngo, H.H.; Guo, H.M. Multi-dimensional habitat vegetation restoration mode for lake riparian zone, Taihu, China. Ecol. Eng. 2019, 134, 56–64. [Google Scholar] [CrossRef]
- Xia, H.J.; Kong, W.J.; Zhou, G.; Sun, O.J. Impacts of landscape patterns on water-related ecosystem services under natural restoration in Liaohe River Reserve, China. Sci. Total Environ. 2021, 792, 148290. [Google Scholar] [CrossRef]
- Xia, J.H.; Yan, Z.M. Advances in research of ecological riparian zones and its trend of development. J. Hohai Univ. 2004, 32, 252–255. [Google Scholar]
- Ye, F.; Ma, M.H.; Wu, S.J.; Jiang, Y.; Zhu, G.B.; Zhang, H.; Wang, Y. Soil properties and distribution in the riparian zone: The effects of fluctuations in water and anthropogenic disturbances. Eur. J. Soil Sci. 2019, 70, 664–673. [Google Scholar] [CrossRef]
- Guo, E.; Chen, L.; Sun, R.; Wang, Z. Effects of riparian vegetation patterns on the distribution and potential loss of soil nutrients: A case study of the Wenyu River in Beijing. Front. Environ. Sci. Eng. 2015, 9, 279–287. [Google Scholar] [CrossRef]
- Martin, T.L.; Trevors, J.T.; Kaushik, N.K. Soil microbial diversity, community structure and denitrification in a temperate riparian zone. Biodivers. Conserv. 1999, 8, 1057–1078. [Google Scholar] [CrossRef]
- Greet, J.; Cousens, R.D.; Webb, J.A. Seasonal timing of inundation affects riparian plant growth and flowering: Implications for riparian vegetation composition. Plant Ecol. 2013, 214, 87–101. [Google Scholar] [CrossRef]
- Neilen, A.D.; Chen, C.R.; Parker, B.M.; Faggotter, S.J.; Burford, M.A. Differences in nitrate and phosphorus export between wooded and grassed riparian zones from farmland to receiving waterways under varying rainfall conditions. Sci. Total Environ. 2017, 598, 188–197. [Google Scholar] [CrossRef]
- Kolbjørn Jensen, J.; Engesgaard, P.; Johnsen, A.R.; Marti, V.; Nilsson, B. Hydrological mediated denitrification in groundwater below a seasonal flooded restored riparian zone. Water Res. 2017, 53, 2074–2094. [Google Scholar] [CrossRef]
- Welsh, M.K.; Vidon, P.G.; McMillan, S.K. Changes in riparian hydrology and biogeochemistry following storm events at a restored agricultural stream. Environ. Sci. Process. Impacts 2019, 21, 677–691. [Google Scholar] [CrossRef]
- Torsvik, V.; Øvreås, L. Microbial diversity and function in soil: From genes to ecosystems. Curr. Opin. Microbiol. 2002, 5, 240–245. [Google Scholar] [CrossRef]
- Knowles, R. Denitrification. Microbiol. Rev. 1982, 46, 43. [Google Scholar] [CrossRef]
- Kuenen, J.G. Anammox bacteria: From discovery to application. Nat. Rev. Microbiol. 2008, 6, 320–326. [Google Scholar] [CrossRef]
- Rütting, T.; Boeckx, P.; Müller, C.; Klemedtsson, L. Assessment of the importance of dissimilatory nitrate reduction to ammonium for the terrestrial nitrogen cycle. Biogeosciences 2011, 8, 1779–1791. [Google Scholar] [CrossRef] [Green Version]
- Reinhold-Hurek, B.; Bünger, W.; Burbano, C.S.; Sabale, M.; Hurek, T. Roots shaping their microbiome: Global hotspots for microbial activity. Annu. Rev. Phytopathol. 2015, 53, 403–424. [Google Scholar] [CrossRef] [PubMed]
- Shelef, O.; Gross, A.; Rachmilevitch, S. Role of plants in a constructed wetland: Current and new perspectives. Water 2013, 5, 405–419. [Google Scholar] [CrossRef]
- Mohanram, S.; Kumar, P. Rhizosphere microbiome, revisiting the synergy of plant-microbe interactions. Ann. Microbiol. 2019, 69, 307–320. [Google Scholar] [CrossRef]
- Yin, X.; Liu, G.; Peng, L.; Hua, Y.; Wan, X.; Zhou, W.; Zhao, J.; Zhu, D. Microbial community of nitrogen cycle-related genes in aquatic plant rhizospheres of Lake Liangzi in winter. J. Basic Microbiol. 2018, 58, 998–1006. [Google Scholar] [CrossRef]
- Chaudhary, D.R.; Gautam, R.K.; Yousuf, B.; Mishra, A.; Jha, B. Nutrients, microbial community structure and functional gene abundance of rhizosphere and bulk soils of halophytes. Appl. Soil Ecol. 2015, 91, 16–26. [Google Scholar] [CrossRef]
- Li, J.; Chen, Q.; Zhao, C.S.; Feng, Y. Influence of plants and environmental variables on the diversity of soil microbial communities in the Yellow River Delta Wetland, China. Chemosphere 2021, 274, 129967. [Google Scholar] [CrossRef]
- Drenovsky, R.E.; Vo, D.; Graham, K.J.; Scow, K.M. Soil water content and organic carbon availability are major determinants of soil microbial community composition. Microb. Ecol. 2004, 48, 424–430. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.Y.; O’Conner, P.J.; Zhang, J.Y.; Ye, X.X. Linking soil nutrient cycling and microbial community with vegetation cover in riparian zone. Geoderma 2021, 384, 114801. [Google Scholar] [CrossRef]
- Veihmeyer, F.J.; Hendrickson, A.H. Soil moisture in relation to plant growth. Annu. Rev. Plant Physiol. 1950, 1, 285–304. [Google Scholar] [CrossRef]
- Barriuso, J.; Solano, B.R.; Lucas, J.A.; Lobo, A.P.; García-Villaraco, A.; Gutiérrez Mañero, F.J. Ecology, genetic diversity and screening strategies of plant growth promoting rhizobacteria (PGPR). J. Plant Nutr. 2008, 4, 1–17. [Google Scholar]
- Lu, B.; Qian, J.; Wang, P.; Wang, C.; Hu, J.; Li, K.; He, X.; Jin, W. Effect of perfluorooctanesulfonate (PFOS) on the rhizosphere soil nitrogen cycling of two riparian plants. Sci. Total Environ. 2020, 741, 140494. [Google Scholar] [CrossRef]
- Lu, B.; Xu, Z.; Li, J.; Chai, X. Removal of water nutrients by different aquatic plant species: An alternative way to remediate polluted rural rivers. Ecol. Eng. 2018, 110, 18–26. [Google Scholar] [CrossRef]
- Chaparro, J.M.; Badri, D.V.; Vivanco, J.M. Rhizosphere microbiome assemblage is affected by plant development. ISME J. 2014, 8, 790–803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frey, S.D.; Lee, J.; Melillo, J.M.; Six, J. The temperature response of soil microbial efficiency and its feedback to climate. Nat. Clim. Chang. 2013, 3, 395–398. [Google Scholar] [CrossRef]
- Vilchez, S.; Manzanera, M. Biotechnological uses of desiccation-tolerant microorganisms for the rhizoremediation of soils subjected to seasonal drought. Appl. Microbiol. Biotechnol. 2011, 91, 1297–1304. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Ji, Z.; Shao, Y.; Guo, C.; Zhou, H.; Liu, L.; Qu, Y. Seasonal variations of soil bacterial communities in Suaeda wetland of Shuangtaizi River estuary, Northeast China. J. Environ. Sci. 2020, 97, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Li, S.; Zhang, Y.; Cheng, H.; Zhou, H.; Qiu, L.; Diao, X. Seasonal variation of anaerobic ammonium oxidizing bacterial community and abundance in tropical mangrove wetland sediments with depth. Appl. Soil Ecol. 2018, 130, 149–158. [Google Scholar] [CrossRef]
- Zhang, L.H.; Song, L.P.; Xu, G.; Chen, P.; Sun, J.N.; Shao, H.B. Seasonal dynamics of rhizosphere soil microbial abundances and enzyme activities under different vegetation types in the coastal zone, shandong, China. CLEAN–Soil Air Water. 2014, 42, 1115–1120. [Google Scholar] [CrossRef] [Green Version]
- Qian, J.; Lu, B.; Chen, H.; Wang, P.; Wang, C.; Li, K.; Tian, X.; Jin, W.; He, X.; Chen, H. Phytotoxicity and oxidative stress of perfluorooctanesulfonate to two riparian plants: Acorus calamus and Phragmites communis. Ecotoxicol. Environ. Saf. 2019, 180, 215–226. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.M.; Cai, W.J.; Li, J.H. Effects of saltwater incursion on the microbiological characteristics and denitrification in a riparian rhizosphere soil in Chongming Island of Shanghai, East China. Ying Yong Sheng Tai Xue Bao. 2012, 23, 1083–1089. [Google Scholar]
- Verhoeven, J.T.; Arheimer, B.; Yin, C.; Hefting, M.M. Regional and global concerns over wetlands and water quality. Trends Ecol. Evol. 2006, 21, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Qian, J.; Zheng, H.; Wang, P.; Liao, X.; Wang, C.; Hou, J.; Ao, Y.; Shen, M.; Liu, J.; Li, K. Assessing the ecohydrological separation hypothesis and seasonal variations in water use by Ginkgo biloba L. in a subtropical riparian area. J. Hydrol. 2017, 553, 486–500. [Google Scholar] [CrossRef]
- Qian, J.; Jin, W.; Hu, J.; Wang, P.; Wang, C.; Lu, B.; Li, K.; He, X.; Tang, S. Stable isotope analyses of nitrogen source and preference for ammonium versus nitrate of riparian plants during the plant growing season in Taihu Lake Basin. Sci. Total Environ. 2021, 763, 143029. [Google Scholar] [CrossRef] [PubMed]
- Pastor, A.; Peipoch, M.; Cañas, L.; Chappuis, E.; Ribot, M.; Gacia, E.; Riera, J.L.; Marti, E.; Sabater, F. Nitrogen stable isotopes in primary uptake compartments across streams differing in nutrient availability. Environ. Sci. Technol. 2013, 47, 10155–10162. [Google Scholar] [CrossRef]
- Kenkel, N.C.; Podani, J. Plot size and estimation efficiency in plant community studies. J. Veg. Sci. 1991, 2, 539–544. [Google Scholar] [CrossRef]
- Reddy, K.R.; Clark, M.W.; Delaune, R.D.; Kongchum, M. Physicochemical characterization of wetland soils. In Methods in Biogeochemistry of Wetlands; DeLaune, R.D., Reddy, K.R., Richardson, C.J., Megonigal, J.P., Eds.; Soil Science Society of America: Madison, WI, USA, 2013; Volume 10, pp. 41–54. [Google Scholar]
- Wang, J.; Song, Y.; Ma, T.; Raza, W.; Li, J.; Howland, G.J.; Huang, Q.; Shen, Q. Impacts of inorganic and organic fertilization treatments on bacterial and fungal communities in a paddy soil. Appl. Soil Ecol. 2017, 112, 42–50. [Google Scholar] [CrossRef]
- Bray, J.R.; Curtis, J.T. An ordination of the upland forest communities of southern wisconsin. Ecol. Monogr. 1957, 27, 326–349. [Google Scholar] [CrossRef]
- Huson, D.H.; Mitra, S.; Ruscheweyh, H.J.; Weber, N.; Schuster, S.C. Integrative analysis of environmental sequences using MEGAN4. Genome Res. 2011, 21, 1552–1560. [Google Scholar] [CrossRef] [Green Version]
- Asnicar, F.; Weingart, G.; Tickle, T.L.; Huttenhower, C.; Segata, N. Compact graphical representation of phylogenetic data and metadata with GraPhlAn. PeerJ 2015, 3, e1029. [Google Scholar] [CrossRef]
- Zaura, E.; Keijser, B.J.; Huse, S.M.; Crielaard, W. Defining the healthy “core microbiome” of oral microbial communities. BMC Microbiol. 2009, 9, 259. [Google Scholar] [CrossRef] [Green Version]
- Douglas, G.M.; Maffei, V.J.; Zaneveld, J.; Yurgel, S.N.; Brown, J.R.; Taylor, C.M.; Huttenhower, C.; Langile, M.G. PICRUSt2: An improved and customizable approach for metagenome inference. BioRxiv 2020. [Google Scholar] [CrossRef] [Green Version]
- Balakumbahan, R.; Rajamani, K.; Kumanan, K. Acorus calamus: An overview. J. Med. Plants Res. 2010, 4, 2740–2745. [Google Scholar]
- Hardie, K.A.Y.; Leyton, L. The influence of vesicular-arbuscular mycorrhiza on growth and water relations of red clover: I. In phosphate deficient soil. New Phytol. 1981, 89, 599–608. [Google Scholar] [CrossRef]
- Allen, M.F. Influence of vesicular-arbuscular mycorrhizae on water movement through Bouteloua gracilis (H.B.K.) Lag ex Steud. New Phytol. 1982, 91, 191–196. [Google Scholar] [CrossRef] [Green Version]
- Kothari, S.K.; Marschner, H.; George, E. Effect of VA mycorrhizal fungi and rhizosphere microorganisms on root and shoot morphology, growth and water relations in maize. New Phytol. 1990, 116, 303–311. [Google Scholar] [CrossRef]
- Carminati, A.; Moradi, A.B.; Vetterlein, D.; Vontobel, P.; Lehmann, E.; Weller, U.; Vogel, H.; Oswald, S.E. Dynamics of soil water content in the rhizosphere. Plant Soil 2010, 332, 163–176. [Google Scholar] [CrossRef]
- Reichman, G.A.; Grunes, D.L.; Viets, F.G., Jr. Effect of soil moisture on ammonification and nitrification in two northern plains soils. Soil Sci. Soc. Am. J. 1966, 30, 363–366. [Google Scholar] [CrossRef]
- Papendick, R.I.; Campbell, G.S. Theory and measurement of water potential. In Water Potential Relations in Soil Microbiology; Parr, J.F., Gardner, W.R., Elliott, L.F., Eds.; Soil Science Society of America: Madison, WI, USA, 1981; Volume 9, pp. 1–22. [Google Scholar]
- Csonka, L.N. Physiological and genetic responses of bacteria to osmotic stress. Microbiol. Rev. 1989, 53, 121–147. [Google Scholar] [CrossRef]
- Angst, G.; Mueller, K.E.; Nierop, K.G.J.; Simpson, M.J. Plant- or microbial-derived? A review on the molecular composition of stabilized soil organic matter. Soil Biol. Biochem. 2021, 156, 108189. [Google Scholar] [CrossRef]
- Desnues, C.; Michotey, V.D.; Wieland, A.; Zhizang, C.; Fourçans, A.; Duran, R.; Bonin, P.C. Seasonal and diel distributions of denitrifying and bacterial communities in a hypersaline microbial mat (Camargue, France). Water Res. 2007, 41, 3407–3419. [Google Scholar] [CrossRef] [PubMed]
- Graham, D.W.; Trippett, C.; Dodds, W.K.; O’Brien, J.M.; Banner, E.B.; Head, I.M.; Smith, M.S.; Yang, R.K.; Knapp, C.W. Correlations between in situ denitrification activity and nir-gene abundances in pristine and impacted prairie streams. Environ. Pollut. 2010, 158, 3225–3229. [Google Scholar] [CrossRef] [Green Version]
- Fang, J.H.; Dong, J.Y.; Li, C.C.; Chen, H.; Wang, L.F.; Lyu, T.S.; Huan, H.; Liu, J. Response of microbial community composition and function to emergent plant rhizosphere of a constructed wetland in northern China. Appl. Soil Ecol. 2021, 168, 104141. [Google Scholar] [CrossRef]
- Tanner, C.C. Plants for constructed wetland treatment systems—A comparison of the growth and nutrient uptake of eight emergent species. Ecol. Eng. 1996, 7, 59–83. [Google Scholar] [CrossRef]
- Dandie, C.E.; Wertz, S.; Leclair, C.L.; Goyer, C.; Burton, D.L.; Patten, C.L.; Zebarth, B.J.; Trevors, J.T. Abundance, diversity and functional gene expression of denitrifier communities in adjacent riparian and agricultural zones. FEMS Microbiol. Ecol. 2011, 77, 69–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, Z.; Guo, L.; Zhang, Q.; Liu, G.; Liu, W. Edaphic conditions regulate denitrification directly and indirectly by altering denitrifier abundance in wetlands along the Han River, China. Environ. Sci. Technol. 2017, 51, 5483–5491. [Google Scholar] [CrossRef]
- Yan, L.; Xie, C.; Liang, A.; Jiang, R.; Che, S. Effects of revetments on soil denitrifying communities in the urban river-riparian interface. Chemosphere 2021, 263, 128077. [Google Scholar] [CrossRef]
- Ligi, T.; Truu, M.; Truu, J.; Nõlvak, H.; Kaasik, A.; Mitsch, W.J.; Mander, Ü. Effects of soil chemical characteristics and water regime on denitrification genes (nirS, nirK, and nosZ) abundances in a created riverine wetland complex. Ecol. Eng. 2014, 72, 47–55. [Google Scholar] [CrossRef]
- Ulrich, D.E.; Sevanto, S.; Ryan, M.; Albright, M.B.; Johansen, R.B.; Dunbar, J.M. Plant-microbe interactions before drought influence plant physiological responses to subsequent severe drought. Sci. Rep. 2019, 9, 249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hallin, S.; Jones, C.M.; Schloter, M.; Philippot, L. Relationship between N-cycling communities and ecosystem functioning in a 50-year-old fertilization experiment. ISME J. 2009, 3, 597–605. [Google Scholar] [CrossRef]
- Ryser, P.; Kamminga, A.T. Root survival of six cool-temperate wetland graminoids in autumn and early winter. Plant Ecol. Divers. 2009, 2, 27–35. [Google Scholar] [CrossRef]
- Marschner, P.; Marhan, S.; Kandeler, E. Microscale distribution and function of soil microorganisms in the interface between rhizosphere and detritusphere. Soil Biol. Biochem. 2012, 49, 174–183. [Google Scholar] [CrossRef]
- Zhou, Y.; Coventry, D.R.; Gupta, V.V.; Fuentes, D.; Merchant, A.; Kaiser, B.N.; Li, J.; Wei, Y.; Liu, H.; Wang, Y.; et al. The preceding root system drives the composition and function of the rhizosphere microbiome. Genome Biol. 2020, 21, 89. [Google Scholar] [CrossRef]
- Houlden, A.; Timms-Wilson, T.M.; Day, M.J.; Bailey, M.J. Influence of plant developmental stage on microbial community structure and activity in the rhizosphere of three field crops. FEMS Microbiol. Ecol. 2008, 65, 193–201. [Google Scholar] [CrossRef] [Green Version]
- Kerr, J.F.R.; Wyllie, A.H.; Currie, A.R. Apoptosis: A basic biological phenomenon with wideranging implications in tissue kinetics. Br. J. Cancer 1972, 26, 239–257. [Google Scholar] [CrossRef] [Green Version]
- Edinger, A.L.; Thompson, C.B. Death by design: Apoptosis, necrosis and autophagy. Curr. Opin. Cell Biol. 2004, 16, 663–669. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, C.; Qian, J.; Hu, J.; Huang, Y.; Lu, B.; He, Y. Rhizosphere Soil Microbial Survival States and N-Related Process during Riparian Plant Dormancy: Influences of Plant Locations and Plant Species. Diversity 2022, 14, 611. https://doi.org/10.3390/d14080611
Zhang C, Qian J, Hu J, Huang Y, Lu B, He Y. Rhizosphere Soil Microbial Survival States and N-Related Process during Riparian Plant Dormancy: Influences of Plant Locations and Plant Species. Diversity. 2022; 14(8):611. https://doi.org/10.3390/d14080611
Chicago/Turabian StyleZhang, Chao, Jin Qian, Jing Hu, Yuanyuan Huang, Bianhe Lu, and Yuxuan He. 2022. "Rhizosphere Soil Microbial Survival States and N-Related Process during Riparian Plant Dormancy: Influences of Plant Locations and Plant Species" Diversity 14, no. 8: 611. https://doi.org/10.3390/d14080611