Biogeography, Systematics, and Ecomorphology of Pacific Island Anoles
by
 ,
,
John G. Phillips
1,2,* ,
,
Sarah E. Burton
1,3,
Margarita M. Womack
4,
Evan Pulver
1,5 and
Kirsten E. Nicholson
1 1
Department of Biology, Central Michigan University, 2100 Biosciences, Mount Pleasant, MI 48859, USA
2
Department of Biological Sciences, University of Idaho, 875 Perimeter Dr., Moscow, ID 83844, USA
3
Department of Fisheries and Wildlife, Michigan State University, 480 Wilson Rd., East Lansing, MI 48824, USA
4
3612 Woodley Rd NW, District of Columbia, Washington, DC 20016, USA
5
3759 Farm to Market Rd. 1488 #175, The Woodlands, TX 77382, USA
*
Author to whom correspondence should be addressed.
Diversity 2019, 11(9), 141; https://doi.org/10.3390/d11090141
Submission received: 1 July 2019
/
Revised: 12 August 2019
/
Accepted: 15 August 2019
/
Published: 21 August 2019
(This article belongs to the Special Issue Systematics and Conservation of Neotropical Amphibians and Reptiles)
Abstract
:Anoles are regarded as important models for understanding dynamic processes in ecology and evolution. Most work on this group has focused on species in the Caribbean Sea, and recently in mainland South and Central America. However, the Eastern Tropical Pacific (ETP) is home to seven species of anoles from three unique islands (Islas Cocos, Gorgona, and Malpelo) that have been largely overlooked. Four of these species are endemic to single islands (Norops townsendi on Isla Cocos, Dactyloa agassizi on Isla Malpelo, D. gorgonae and N. medemi on Isla Gorgona). Herein, we present a phylogenetic analysis of anoles from these islands in light of the greater anole phylogeny to estimate the timing of divergence from mainland lineages for each species. We find that two species of solitary anoles (D. agassizi and N. townsendi) diverged from mainland ancestors prior to the emergence of their respective islands. We also present population-wide morphological data suggesting that both display sexual size dimorphism, similar to single-island endemics in the Caribbean. All lineages on Isla Gorgona likely arose during past connections with South America, and ecologically partition their habitat. Finally, we highlight the importance of conservation of these species and island fauna in general.
1. Introduction
Island ecosystems often display highly diverse communities, distinct from their continental counterparts. This has led to a rich body of work examining communities on islands to understand their ecological dynamics and the processes that promoted their formation [1,2,3]. As a result, islands have served as model systems for examining evolutionary processes like community assembly and speciation. Particularly in taxonomic groups with reduced lability, an island–mainland divide can promote allopatric speciation, leading to island endemism [4,5,6]. Many archipelagos that have featured prominently in the annals of evolutionary biology are of volcanic origin (e.g., the Canary, Galápagos, and Hawaiian Islands), where the nature of their formation necessitates any inhabitants to have almost certainly arrived via overwater dispersal [7], but the process of community assembly on continental islands can be different. While dispersal is a viable explanation for communities on continental islands, any historical connection to the mainland yields the possibility of a vicariant origin for those species. In some cases, support for both options has been presented for a variety of taxa inhabiting the same island or archipelago, with disagreement among biologists [8] and sources within. These studies on island biogeography have produced important, although sometimes controversial ideas [9,10] and additional studies may continue to influence our current understanding of island communities.
One group of organisms that has been influential in our understanding of island biogeography and community assembly therein, are anole lizards (family Dactyloidae). Anoles are highly diverse with 429 species, [11,12] and display a variety of morphological and ecological differences. They inhabit a wide breadth of microhabitats from forest floor to canopy, exhibit a concomitant array of morphological features adapted to exploiting those niches, and display equally diverse behaviors and ecological characteristics [13]. Studies of Caribbean anoles have figured prominently in the literature with respect to novel ecological and evolutionary patterns [14,15,16] and have played a role in testing hypotheses of island colonization [17,18]. Caribbean anole research has supported overwater dispersal as the generally accepted paradigm for the biogeographic origin Caribbean anoles [19,20]. However, this line of inquiry has been limited to the Caribbean islands, and studies of anoles in Pacific Islands have been lacking.
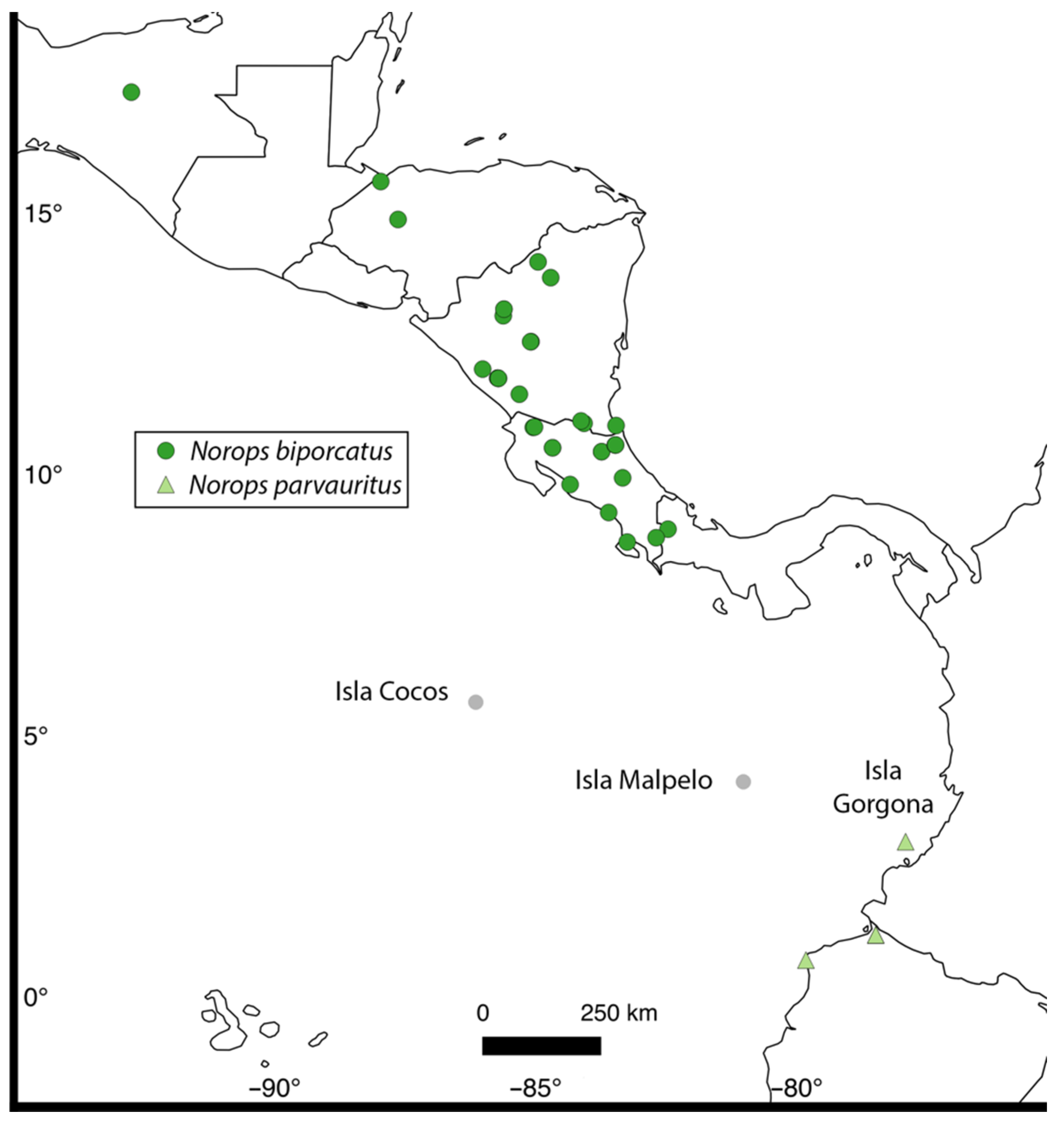
The paucity of work on Pacific anoles may be attributable to a lack of taxa and islands as only a combined seven species of anoles (Figure 1) are known from the Eastern Tropical Pacific (ETP) across three islands (Islas Cocos, Gorgona, and Malpelo; Figure 2 and Figure 3). The islands are variable in ecology and geology, which may impact the evolution of their biotic communities. Anoles occupying these islands span the extreme ends of the dactyloid phylogeny [20,21], and are highly variable in ecology and natural history (reviewed below). Genetic analyses of these species may shed light on the evolutionary patterns and processes in the Pacific Islands, as well as provide a framework for the comparison of evolutionary patterns to the Caribbean anoles to better understand island evolution. Among the Pacific islands inhabited by anoles, only two contain a single species of anole or “solitary anoles” (sensu Schoener [22]): Isla Malpelo, Colombia is occupied by Dactyloa agassizi, and Isla Cocos, Costa Rica is inhabited by Norops townsendi. The third Pacific anole island, Isla Gorgona off Colombia, is occupied by five species: D. chocorum, D. gorgonae, D. princeps, N. medemi, and N. parvauritus (Figure 1). Below we briefly review each island in terms of geologic origin and known ecomorphology of their anole communities.
While there is debate within the anole community as to the generic level nomenclature of anoles, herein we opt for the eight-genus model proposed by Nicholson et al. [11,18], highlighting the evolutionary distinctness among anoles. Our study illustrates the utility of treating Norops and Dactyloa as the distinct non-sister genera separated by as much as 31–87 million years ([11,23]; although Román-Palacios et al. [24] performed more rigorous comparative analyses on the dating methods suggesting this basal divergence likely occurred 51–65 Mya). If considered the same genus, a reader who is unfamiliar with the intricacies of anole taxonomy may erroneously assume that solitary anoles D. agassizi and N. townsendi are comparable in morphology and ecology, which is a misleading assumption. Each species is more closely related to separate clades of solitary Caribbean anoles which make for more appropriate comparisons of ecomorphological evolution on islands (D. extrema, D. luciae, and D. roquet for D. agassizi and N. concolor, N. lineatus, and N. pinchoti for N. townsendi; [25]).
1.1. Isla Malpelo
Isla Malpelo (3.5 km2, Figure 2a) is a remnant volcanic plug (hardened magma within the vent of a volcano) and is estimated to be 15–17 million years old [26], representing an older stage in the life of a volcanic island. A younger Malpelo was considerably larger and most likely vegetated [27], but now the floral communities of Malpelo’s steep slopes are sparse, which likely contributes to a species-poor ecosystem [28,29,30,31]. No terrestrial vertebrates are found on Malpelo except for three endemic species of lizards: an anole (Dactyloa agassizi), a galliwasp (Diploglossus millepunctatus), and a gecko (Phyllodactylus transversalis). Without sufficient primary producers to anchor the trophic pyramid of Isla Malpelo’s ecosystem, all three lizards are dependent upon the colonies of a unique keystone species, the Nazca Booby (Sula granti), which seasonally nests on the island [32]. Lizards and crabs (Geocarcinus malpilensis) compete inter- and intra-specifically for booby fecal matter, unprotected eggs, and dropped or regurgitated fish. Dactyloa agassizi is the most abundant lizard species on the island with an estimated population of 140,000–206,000 [33]. Its natural history and behavior are unique among anoles, in that individual males do not defend territories or display aggression toward one other [34], (pers. obs. KEN and MMW). This may be due to a lack of importance of territory, given the stochastic nature of nutrient availability and the lack of habitat heterogeneity. They are also very curious and unafraid of humans, readily approaching and investigating them, presumably in search of food [34], (pers. obs. KEN and MMW). Dactyloa agassizi belongs to a clade of anoles (Dactyloa, sensu Nicholson et al. [21]) ranging from northern Costa Rica to central South America [35,36,37]. A recent study examined the divergence of D. agassizi from mainland D. insignis, finding this split to predate the origin of Isla Malpelo [24]. Dactyloa insignis is one of the northernmost-ranging Dactyloa species, so the mainland ancestor of D. agassizi may have originated in northern Costa Rica.
1.2. Isla Cocos
Isla Cocos (24 km2, Figure 2b) also has a single endemic anole, Norops townsendi. The environment of Isla Cocos is dramatically different from Malpelo [38], (pers. obs. KEN and MMW), being a highly vegetated and lush tropical island with cloud forest around its highest peak. Similar to Isla Malpelo, Cocos is volcanic in origin, although its origin is estimated to be much younger at 1.9–2.4 Mya [39]. Isla Cocos likely represents how Malpelo might have appeared earlier in its geologic life, while Cocos will likely more closely resemble Malpelo over time. Norops townsendi is found throughout the island from sea level to its peak (Cerro Yglesias, 575 m) and is very abundant [38], (pers. obs. KEN and MMW). The species is most often observed on the trunks of the trees with no obvious preference for tree species on which to perch but has been observed very high up in the canopy, as well as occasionally capturing food items on the ground (pers. obs. KEN and MMW). These observations suggest N. townsendi to be an ecological generalist, although we did not collect sufficient data to specifically address the question of its trophic ecology. Norops townsendi was previously thought to be morphologically allied to N. polylepis from Costa Rica [40], but phylogenetic analyses have since suggested it is allied closer with N. poecilopus, a semi-aquatic anole from Panama [21,41,42,43], but see [20].
1.3. Isla Gorgona
Isla Gorgona (13.3 km2, Figure 2c) is only 35 km from the coast of Colombia and, like Isla Cocos, is a small tropical island that is separated from the mainland by a deep marine channel (~270 m; [44]). The island falls entirely within Gorgona National Natural Park which covers ~600 km2 [45]. It has a complex geologic history perhaps dating back to 90 Mya [46,47,48,49]. While Isla Gorgona formed separately from the mainland, there is evidence to suggest multiple connections and subsequent separations from the mainland [50]. However, the timing of the connections is unclear. Most recently, Gorgona was connected to mainland South America from 17,000–11,000 years ago during the Pleistocene glaciation [51,52], which may explain some of the biotic similarities between the island and mainland communities [53,54]. The dynamic geologic history sets the stage for different components of Isla Gorgona’s biotic community to have originated via different modes: dispersal or vicariance. However, without more refined dates of the connection history we cannot test hypotheses regarding these alternatives, but we can provide interpretations of divergence dates estimated from our data.
Five anole species inhabit the island, representing a diverse range of morphological and ecological modalities as well as phylogenetic affinities (Figure 4). Dactyloa gorgonae and Norops medemi are endemic to Isla Gorgona, while D. chocorum, D. princeps, and N. parvauritus have populations in mainland South America in addition to the island. The two species of Norops (N. parvauritus and N. medemi) differ greatly from each other in ecomorphology in that Norops medemi is smaller and found low on the trunks of trees and is darkly colored and patterned, similar to the tree trunks on which they are found. On the other hand, one of the largest members of Norops, the brilliant green N. parvauritus has a robust body with slender limbs. This species often goes unobserved as it frequents the forest canopy and is ecologically similar to the Dactyloa species. This species was previously recognized as a subspecies of the widespread N. biporcatus (ranging from Mexico to central South America) until it was recently elevated to species status [55]. In addition to examining the timing of N. parvauritus on Isla Gorgona, we also used this opportunity to evaluate this proposed taxonomic change. Armstead et al. [55] had limited genetic sampling of both N. biporcatus (N = 5) and N. parvauritus (N = 3), with no individuals from Isla Gorgona, the type locality for Norops biporcatus parvauritus. Herein we address both deficiencies by presenting a broad genetic data across the range including from Isla Gorgona. While Armstead et al. (2017) also presented a morphological analysis to support the species elevation, past work in mainland anoles has shown that delineating species using morphology without range-wide genetic sampling can lead to the violation of a monophyletic species concept (e.g., N. quaggulus, [43,56]).
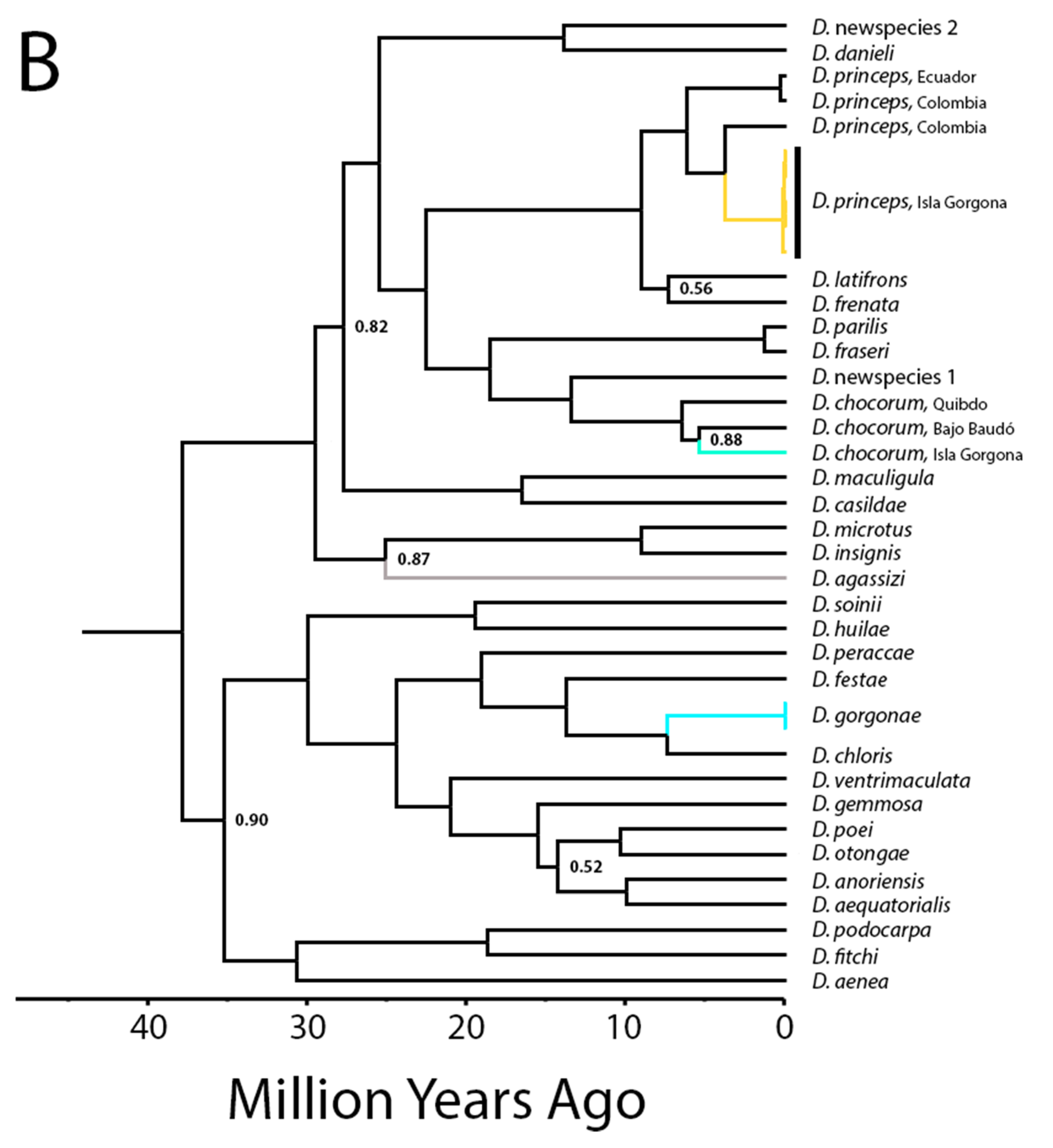
The other three species are in the genus Dactyloa (D. chocorum, D. gorgonae, and D. princeps) and are all highly arboreal, but differ in size, coloration, and perch preferences (pers. obs., this study). Dactyloa gorgonae is unique among all anoles in that it is bright blue in color, smaller, and slimmer than the other two Dactyloa, and believed to be most closely related to a mainland species (D. chloris), to which it is morphologically similar except in coloration. Locals on Isla Gorgona believe that D. gorgonae is only found within the small inhabited area on the island, where it is observed in the mostly open branches of trees, but that assertion is likely due to frequency of observations and insufficient sampling effort elsewhere on the island. The forest on the island is dense, and their blue coloration is quite cryptic within the foliage of the trees (pers. obs. KEN and MMW). Dactyloa princeps was observed frequently on the trunks of trees while D. chocorum was observed rarely (only once during our visit). Both D. princeps and D. chocorum have populations on the mainland, but no studies have examined the morphological differences among island populations.
Understanding the biogeographic history of these Pacific anoles is a crucial initial step in setting the stage for future ecological and evolutionary studies of these species. Investigating the timing and route(s) of colonization for each island, combined with knowledge of each island’s likely environment at the time of colonization, will establish the historical context for the unique extant communities. We investigated the biogeography of these Pacific Island anoles using molecular data to address the timing of divergence from mainland ancestors and to evaluate the route of colonization of each island by each of the seven species. This information was used to compare divergence of island lineages to geologic models for each island and to investigate hypotheses of overwater dispersal to each island. Islas Cocos and Malpelo are both volcanic in origin and could only have been colonized via over water dispersal, but we sought to investigate the timing and route of potential pathways. We also investigated the timing of divergence from mainland populations for all five species on Isla Gorgona and compared them to evaluate if their colonization was temporally proximate.
Finally, we collected observations and ecomorphological measurements for comparing to those recorded for Caribbean anoles to examine the relationship in ecomorphological patterns between solitary anoles. Sexual size dimorphism is evident in many solitary species of birds [57,58], mammals [59], and lizards [22,60,61]. Solitary anoles in the Caribbean typically display sexual size dimorphism, presumably to reduce intraspecific competition [22], a trend that likewise holds in mainland populations with species-poor anole communities [62]. Given that sexual size dimorphism has not been fully evaluated in Pacific species, we seek to determine if D. agassizi and N. townsendi also adhere to this pattern.
2. Methods
Each island was visited in July or August 2004, for 3–10 days. Because of permit restrictions, collection of samples for genetic analysis was restricted to only five individuals per species for all islands. In the case of Dactyloa gorgonae, collection was further limited to N = 2 by station officials on Isla Gorgona. Cryptic and arboreal niches allowed for collection of only one D. chocorum and one N. parvauritus, but D. gorgonae, D. princeps, and N. medemi were more common and useful for population-wide morphological data (see Supplementary Table S1 for specimen data). For both N. townsendi (Isla Cocos) and D. agassizi (Isla Malpelo) full populations were observed to assess habitat use and captured and measured when possible (see Morphology and Ecology section below). Additionally, to evaluate the taxonomic status of N. parvauritus, samples of N. biporcatus were collected throughout Central America (Mexico through Panama, Figure 3, Supplementary Table S2). We complied with all applicable Animal Care guidelines (CMU-IACUC # 10–02).
2.1. DNA Sequencing and Phylogenetic Reconstruction
Tissue samples for each species were collected and preserved in 95% EtOH. Voucher specimens were fixed in formalin, stored in EtOH, and deposited with the Universidad de los Andes, Colombia. Qiagen DNEasy kits were used for DNA extraction. PCR amplification of several contiguous mitochondrial genes (ND2, tRNATrp, tRNAAla, tRNAAsn, tRNACys, tRNATyr, and partial CO1) used previously in the literature for phylogenetic reconstruction of mainland anoles [42,43] was conducted. Genes were amplified using Empirical Bioscience Mean Green Master Mix and the same cycling conditions and primers as Nicholson et al. [21]. PCR products were purified using Qiagen QIAquick purification kits, followed by sequencing at Michigan State University’s (MSU, East Lansing, MI, USA) sequencing core. Sequence data were edited and aligned using MUSCLE, then combined with published sequences for individuals of the same species from mainland Central or South America and closely related species (Supplementary Table S1). Our newly collected data were combined with published data for 75 Dactyloa and Norops species plus 10 non-Dactyloid outgroup taxa (Table 1 and Supplementary Table S1) for a total of 1480 aligned base pairs of sequence data (Genbank vouchers in Supplementary Table S1).
Phylogenetic analyses were conducted to investigate the placement of taxa among established species of anoles for which comparative data were available. Phylogenetic reconstructions were conducted using BEAST2 for Bayesian analyses under a lognormal relaxed clock model and a Yule Speciation prior [63]. PARTITIONFINDER was used to select models of evolution as well as to examine the suitability of partitioning. In all cases each gene (including tRNAs) was entered as a potential partition for the first, second, and third codon positions. Branch lengths were unlinked, all models of evolution available in BEAST2 were tested, and a BIC information criterion and greedy algorithm were used. The PARTITIONFINDER analysis recommended a GTR + Γ model of evolution for the mtDNA segment without partitioning. The same alignment for each dating method was run for 20 million generations with 20% burn in removed via TreeAnnotator [63] after evaluating that the analysis had reached stationarity using Tracer v.1.6.0 [64].
We estimated the date of origin for each island species by analyzing our data using BEAST2. We used two approaches to estimating the age of divergence between each island lineage and its mainland relatives: (1) using the Macey et al. [65] calibration rate of 0.65 mutations per 100 bp per million years, used by several mtDNA studies of anoles and related taxa [66,67,68,69,70,71] and (2) implementing the same mutation rate in addition to three calibration dates for Iguanian fossils used by Prates et al. [37] and sources within. We used this approach following Román-Palacios et al.’s [24] findings that implementing multiple dating strategies can avoid the pitfalls of trees based only on a single mutation rate [72].
From our phylogenetic reconstruction we compared divergence dates between the island and mainland lineages to the estimated timing of origin for each island; our null hypothesis was that each species’ origin would post-date the emergence of each island but had no a priori expectations or hypotheses regarding when each species may have originated. For some species (N. medemi, N. townsendi) for which we had multiple samples within an island, we were also able to estimate the coalescence of species on each island, making the assumption that this date represents the youngest possible age of colonization.
2.2. Morphology and Ecology
On each island, observations were made for all species during diurnal and nocturnal survey periods (except N. agassizi, as we were not allowed on Isla Malpelo at night). Individuals were captured with collapsible fishing poles fitted with string lassos in order to record population-wide morphological data. Measurements were taken on all captured individuals (SVL, mass, fore- and hind-limb length, head width, depth and length, number of lamellae rows) as well as recording ecological data (perch height, perch diameter, perch type; Supplementary Tables S3–S5).
Student’s t-test was used to compare differences between the sexes for each species (although fore- and hind-limb length, head width, depth and length, were all determined to covary with SVL and were omitted from the sexual dimorphism analysis). Additionally, ecological and morphological data for N. townsendi were collected at four different locations on the island, including the highest point (Cerro Yglesias) and a small islet (Manuelita) that is disconnected from Isla Cocos. One-way ANOVA assuming unequal variance was implemented to test for differences among sites, which may suggest separate populations. This was followed by a t-test for any variables that displayed a significant difference (p < 0.05) to evaluate pairwise relationships between populations. To account for multiple tests in all cases, we applied a sequential Bonferroni correction. The same process was used to evaluate the differences in habitat use (perch height, perch diameter) among three of the Isla Gorgona species (D. gorgonae, D. princeps, N. medemi).
3. Results
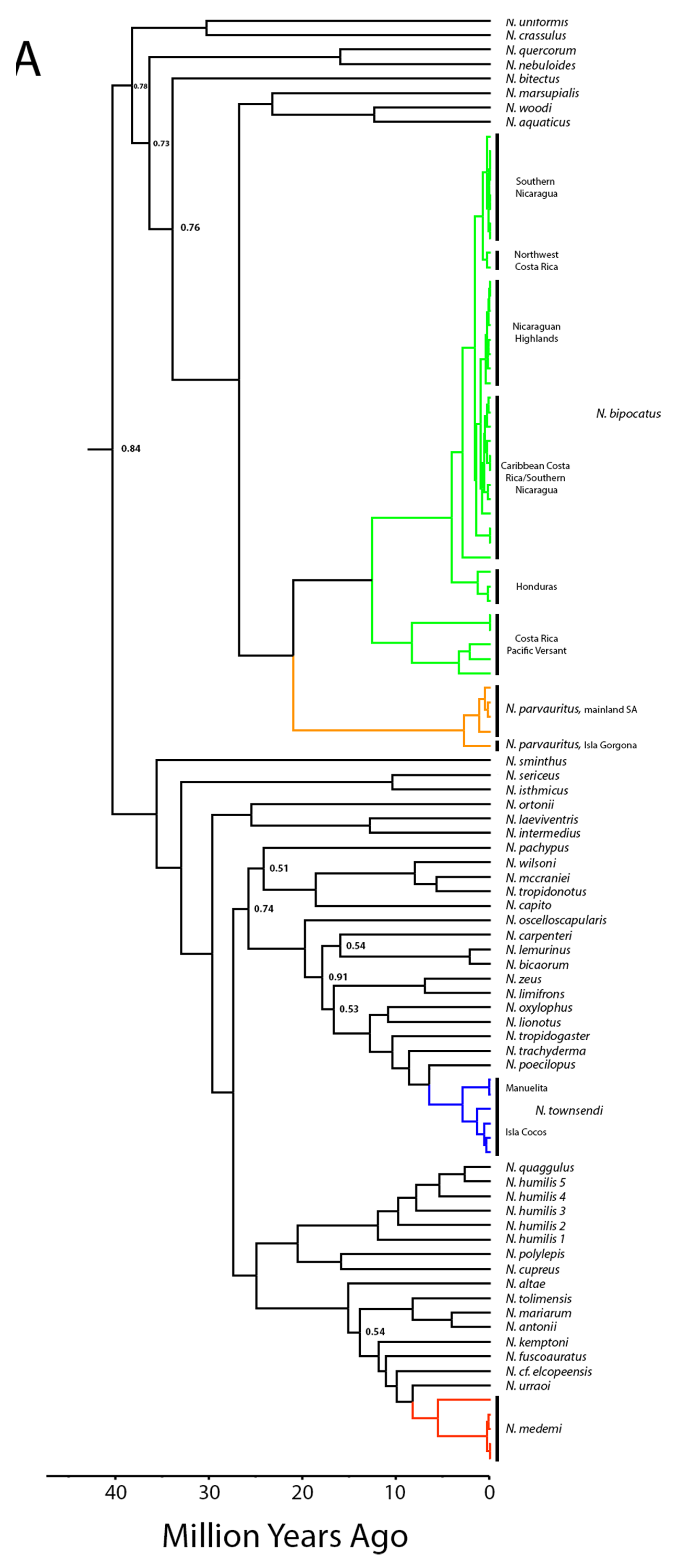
The results of our phylogenetic reconstruction are depicted in Figure 4. All island taxa were reconstructed within their previously assigned genera (Dactyloa and Norops), and all relationships described herein have high support (posterior probability > 0.88). For Isla Gorgona, D. chocorum and D. princeps were both nested within clades of mainland conspecifics. The D. princeps clade was recovered to be sister to D. frenatus and D. chocorum sister to D. fraseri in all analyses. As previously hypothesized, D. gorgonae was placed sister to D. chloris. Within Norops, the island N. parvauritus was placed within a clade of mainland N. parvauritus from Ecuador, all of which is sister to N. biporcatus. Norops townsendi was recovered as sister to N. poecilopus and a clade of semiaquatic anoles that also include non-aquatic N. trachyderma and N. tropidogaster, and endemic N. medemi was placed sister to N. urraoi (a newly described species from the northwestern Andes) as suggested by Grisales-Martínez et al. [41], and nested within a clade of species from northern Colombia, Panama, and Costa Rica (N. fuscoauratus, N. kemptoni, and N. “newspecies,” (believed to be N. cf. elcopeensis per [73]). There was also a deeper divergence within N. medemi, suggesting the species has been on Isla Gorgona at least for 6.1 Mya (5–8 Mya, 95% CI).
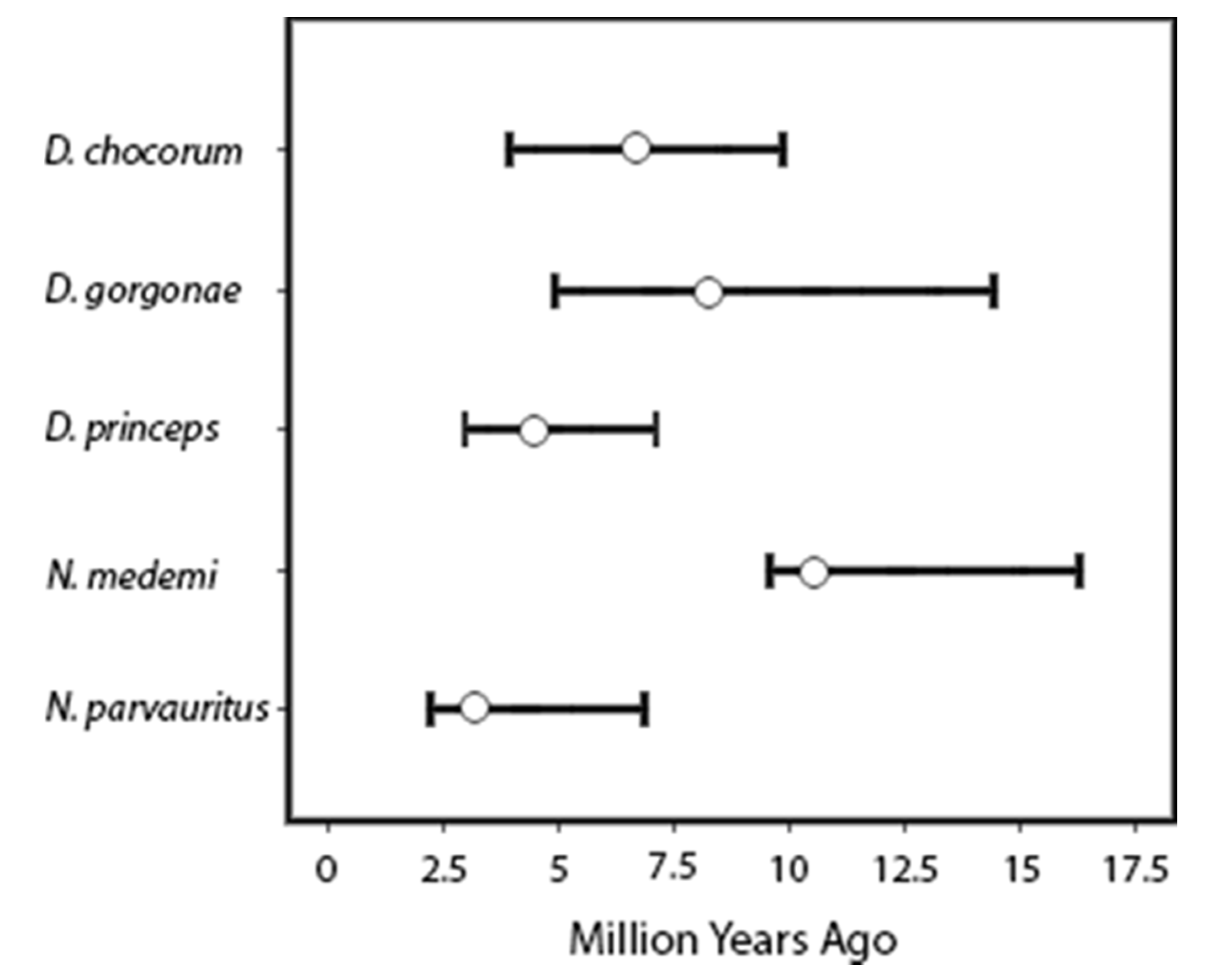
Node dating analyses were similar when using fossil dates + mutation rate versus the molecular calibration rate alone (Figure 4, Dactyloa chloris 1). These estimated dates of D. agassizi (as suggested in Román-Palacios et al. [24]) and N. townsendi predate the predicted origin for Islas Malpelo and Cocos respectively, while divergence dates for the Isla Gorgona taxa from their mainland counterparts are all younger than the island (all estimates within the past 10 My; Table 1, Figure 5).
Morphological Data
Males were significantly larger than females in D. agassizi, D. princeps, and N. townsendi (Table 2). Most of the morphological variables (SVL, head, and limb measurements) are likely the products of having a larger body size and therefore cannot be characterized as independently evolving traits, all three species unquestionably display sexual size dimorphism, similar to solitary anoles in the Caribbean (Table 3). Norops medemi on the other hand displayed no significant differences between sexes with the exception of males having one more row of toe lamellae on average. However, sample sizes for this species were very small (N = 5 for each sex). Because of small sample sizes for D. chocorum and N. parvauritus, and only one female measured for D. gorgonae, those three species are omitted from further morphological analysis.
Comparative sample sizes from the Caribbean were small for solitary relatives to D. agassizi and N. townsendi without wide-scale population data available but serve as an exploratory indication of variation within and between species. Both sexes of D. agassizi are considerably larger than their Caribbean counterparts, although the magnitude of sexual size dimorphism is similar to D. luciae (and also the distantly related N. concolor). Norops townsendi displayed a lower magnitude of sexual size dimorphism than any other species of solitary Norops, although the difference was still significant (Table 2 and Table 3).
The four populations of N. townsendi displayed significant differences among each other. Two of these populations (Chatham Trail and Playa) showed few differences except for Playa individuals being more massive with longer hindlimbs. Mass could be attributed to differences in prey availability between the sites, so we also re-analyzed our populations with these two sites combined to treat as a single population (hereafter referred to as the “main” population). In all morphological measurements, Cerro Yglesias individuals were significantly smaller than the main population, while Manuelita individuals were significantly larger with a greater number of toe lamellae (Table 4). The main population occupied perches >1 m higher than either the cerro or the islet, which may provide an explanation for the divergence in morphological evolution among the populations.
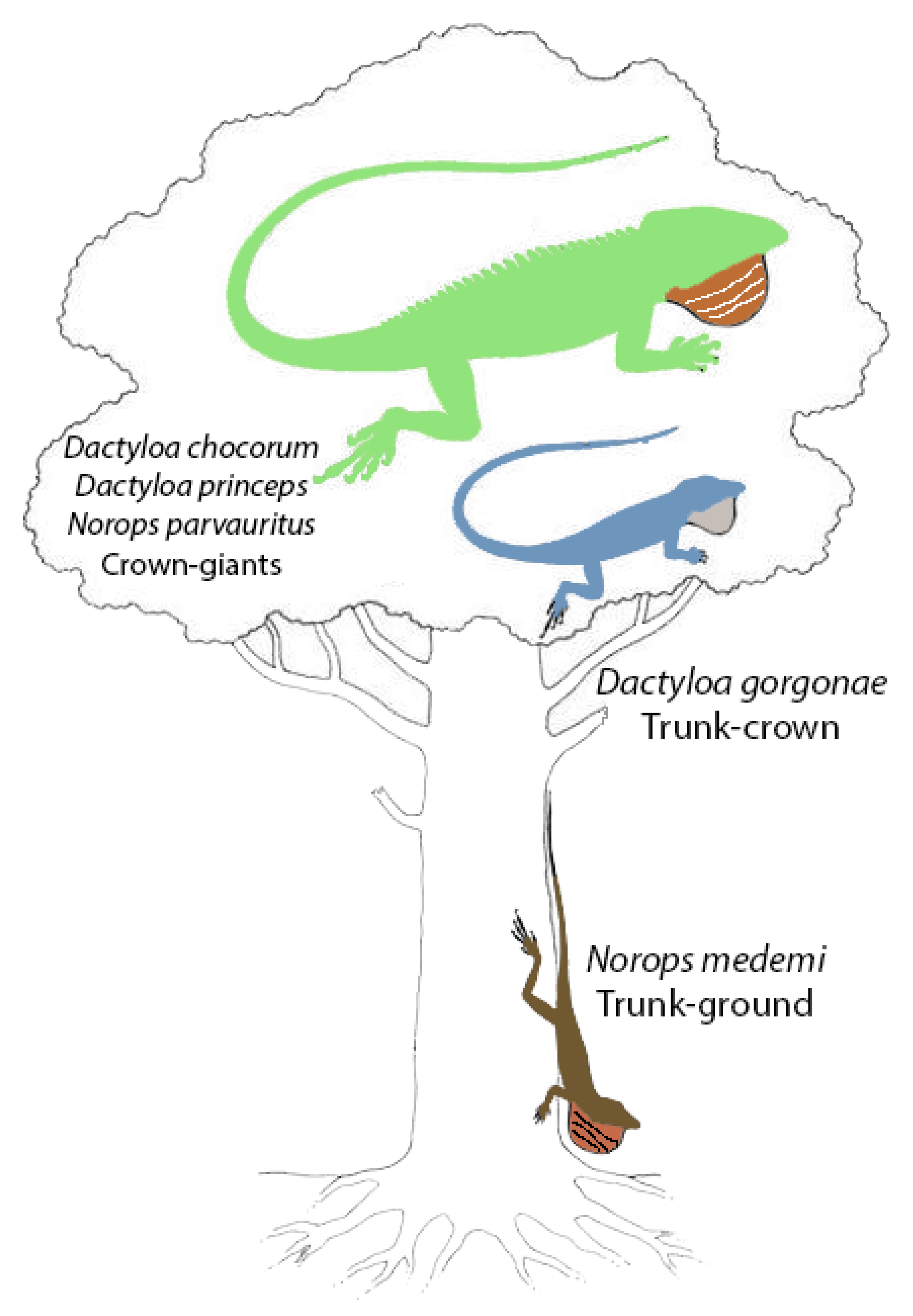
Perch height, diameter, type, and position were difficult to evaluate objectively for D. agassizi because the surface of the island consists of bare rock with nothing to perch upon except occasional boulders and pebbles. We observed some individuals perching on boulders, but most were on the ground or on pebbles, rendering a perch height mean of 1.5 m and perch diameter that was not quantifiable (often individuals were on the ground); perch type could only be described as rock. Alternatively, perch height, diameter, type, and position were measurable for N. townsendi because Isla Cocos had a variety of available perch heights and types. Norops townsendi was observed throughout the island on everything from sand to branches well within the canopy, making averages or generalizations difficult. Canopy height extended well over 20 m (the upper limit of our measuring abilities) and animals were often observed as high, or within the canopy. The aforementioned lack of samples for two of the Isla Gorgona species, inhibits our ability to evaluate habitat use across the local anole community, especially since the low number (1) of D. chocorum and N. parvauritus observed is likely due to their preferred niche in the canopy. The forest of Isla Cocos is similar to non-climax lowland rainforest habitats of Central America in having a range of tree species of varying heights and diameters with substantial undercover shrubs providing a variety of perches and niches. However, we found that D. gorgonae occupies higher perches than either D. princeps or N. medemi, while D. princeps occupied significantly larger perches than N. medemi (Table 5), likely owing to a much larger body size and longer limbs (Table 2). This habitat partitioning aligns with Nicholson et al.’s [21] ecomode classification, where each species occupies a different ecomode: D. gorgonae = trunk-crown, D. princeps = crown giant, N. medemi = trunk ground (Figure 6). Dactyloa chocorum and N. parvauritus are also both considered crown giants, but D. princeps clearly utilizes habitat lower down in the forest.
4. Discussion
4.1. Biogeographic and Phylogenetic Interpretations
We estimated the dates of divergence from mainland populations for Pacific Island anoles from molecular data. The range of estimated dates is consistent with past findings: the ancestors of the included species have a long history on the mainland [20,21,24], are derived from clades with disparate physical and temporal origins. The genus Dactyloa represents one of the first splits in the anole tree and arose in the northern South America, while Norops (+Ctenonotus) is one of the last clades to diverge and occurred approximately 20 million years later [24] in the Caribbean, followed by dispersal to Central America before dispersing south into South America [21].
We interpret our data as follows, beginning with the multi-species island of Gorgona. Isla Gorgona has a complex geologic history leading to its aerial emergence potentially by 27 Mya [50]. The island is close to the mainland (although currently separated by a deep channel) and because of previous contact with the mainland, it could have been easily colonized early by overland dispersal prior to its separation from the mainland, or via overwater dispersal when the sea levels were lower, causing the landmasses to be more easily reachable. Three of the five Isla Gorgona species do not appear to differ morphologically from their mainland counterparts. However, denser sampling in mainland Colombia and Ecuador might be able to elucidate closer relationships than our present sampling allows (see N. urraoi example, [41]). Dactyloa chocorum occurs both on mainland South America and on Isla Gorgona and there have been no identified morphological distinctions recorded to date. Dactyloa gorgonae only differs from mainland D. chloris in being solid blue. Dactyloa princeps was always believed to be closely related to D. frenata with some investigators debating whether any features distinguish the two, unpubl. per [35,79,80], although our phylogenetic estimates support evolutionary distinctness. The lack of obvious morphological differences from mainland relatives may be due to low selection pressure, suggesting the environment on the island is strongly similar to the mainland environment from which it is derived, or to a recent arrival on the island that has yet to produce substantial change.
The two Norops species’ colonization of Isla Gorgona are even more interesting considering the lack of solid hypotheses regarding the origin and biogeography of Norops as a whole. Early hypotheses suggested that South American Norops couldn’t have invaded from Central America until after the closure of the Panamanian Portal which was believed to have occurred ~3.5 Mya [10]. However, the timing of this event is believed to have occurred multiple times with several estimates at 23–25 Mya ([81,82] with some controversy on this date reviewed in Elmer et al. [83]), 15 Mya [84], and 3.5 Mya [85,86]. More recent work has postulated that perhaps the Norops clade was widespread across Central America and South America before the continents were separated by the eastward movement of the proto-Greater Antillean land arc [21]. The estimated dates of divergence for N. parvauritus (3.2 Mya) and coalescence for N. medemi (6.6 Mya, Table 1) would lend support for the Nicholson et al. [21] biogeographic hypothesis, although more testing is required to investigate that hypothesis. Our sampling of Isla Gorgona was low because of the cryptic nature of some species combined with government permitting restrictions, so these dating estimates are tentative and should be tested in future with additional data. However, our findings of habitat partitioning among three of the Isla Gorgona species is of interest and warrants further study.
The biogeographic scenarios for the two solitary anoles (D. agassizi—Isla Malpelo; N. townsendi—Isla Cocos) are more difficult to interpret. In both cases, the estimated timing of divergence from mainland relatives (D. agassizi: 22.8 Mya; N. townsendi: 7.6 Mya) considerably predate the islands’ estimated origin (Malpelo: 15–17 Mya; Cocos: 1.9–2.4 Mya). One explanation for both solitary anoles could be that each species originated on the mainland prior to dispersal to each island, followed by subsequent extinction of the species on the mainland, a conclusion also reached by Román-Palacios et al. [24]. The route of dispersal to the islands from the mainland appears to be fairly straightforward from the standpoint of currents, as currents move virtually straight to each island from the mainland at regular intervals throughout the year [29,50], making over-water dispersal plausible. For many over-water dispersal scenarios it may seem improbable for anoles to disperse over great expanses of water, but other species made the same trip and colonized the same islands, and similar dispersal events have been observed and hypothesized for anoles and other lizard fauna in the Caribbean [19,67,87].
Our calculated divergence estimate for D. agassizi is similar to Román-Palacios et al. [24], who found that D. agassizi and its sister species, D. insignis, diverged during the Oligocene, 22–26 Ma. We recovered D. agassizi sister to a sister-pair of D. insignis and D. microtus with a mean divergence of 31 (fossil 95% 22–38) or 29 Ma (rate 95% 23–34). Román-Palacios et al. [24] did not include D. microtus in their phylogeny, so this is not a contradiction of their topology, although our dates suggest an older divergence, which is concurrent with the assessment that rate-based methods can overestimate the ages of shallower nodes if gene saturation is in effect [72]. Poe et al. [20] placed D. agassizi as sister to a clade of 14 other species of Dactyloa, but did not include D. insignis in their analysis. They also included N. townsendi within a group of anoles aligned with N. trachyderma. We note that Poe et al. [20] also included other loci in their analysis, which might account for the differences seen here.
Prior to this study, no one had published an estimated age of colonization for N. townsendi. Our analyses suggest a split within N. townsendi around 3.5 Mya that separates individuals on Isla Cocos from those on Manuelita Islet (Figure 4). This is problematic in that the divergence predates the origin of the island, but our limitations on sample collection might be influencing our results. A more fine-scale sampling effort could provide a more reliable estimate of the coalescence between the two lineages (incidentally, the Manuelita population was significantly larger than any of the three populations on Isla Cocos, Table 4). The phylogenetic placement of N. townsendi was unknown until recently but was presumed to be within the mainland Norops clade [88,89]. Our analyses also placed it sister to N. poecilopus, a semi-aquatic anole from Panamá (100% posterior probability support), concurring with other recent topologies [21]. This clade also contains two additional semi-aquatic species (N. lionotus and N. oxylophus). If N. townsendi evolved from a semi-aquatic ancestor, it may have had more opportunities for rafting, and also makes biogeographic sense, as the currents could have carried an ancestor from Panama to Isla Cocos. Other semi-aquatic anoles (N. lynchi, N. macrolepis, N. rivalis) occur in coastal South America that are missing from our phylogeny, providing other possible relatives. If one of these shares a more recent common ancestor with N. townsendi, it may result in a more recent estimate of colonization closer to the origin of Isla Cocos.
The oceanic currents may have played a significant role in the colonization of the islands in the eastern Pacific region by anoles and other taxa [35,90,91]. El Niño Southern Oscillation (ENSO) cycles are believed to have played an important role in island colonization in the past three to five million years [92,93], which could explain dispersal events to Isla Cocos and even the Galápagos Archipelago [3]. As suggested by Román-Palacios et al. [24], this is too recent to influence the divergence of D. agassizi, but the Colombian Current as part of the Humboldt Current has been active for over 100 million years [94], and likely was an important factor in the colonization of Isla Malpelo by all inhabitants. For both D. agassizi and N. townsendi, divergences predating island emergence strongly suggest dispersal from the mainland (likely lower Central America given their closest ancestor as suggested by our phylogenetic reconstruction). This could have taken place via island hopping, possibly using seamounts that were previously aerial [24,95,96,97]. Recent work in the ETP suggests such a scenario for previously emergent landmasses that are part of the Galápagos archipelago [98], therefore, we cannot discount the possibility that there are other seamounts in the ETP that were previously emergent and could have served as the “stepping-stones” to Isla Cocos and/or Malpelo.
4.2. Taxonomic Implications
Our findings support Armstead et al. [55] in their recognition of Norops parvauritus as a distinct species from N. biporcatus. The age of the divide between the two species is older than many other species pairs, so we conclude this taxonomic change was warranted. Our inclusion of a specimen from Isla Gorgona (the type locality) was highly important to confirm the specific status of N. parvauritus. Fine-scale sampling throughout Mesoamerica also suggests geographic structure in N. biporcatus, with some division aligning with the continental divide in Costa Rica and the Nicaraguan highlands (Figure 4, Supplementary Figures S1 and S2).
Given the placement of the Isla Gorgona N. parvauritus as sister to the rest of its species cannot rule out the possibility of N. parvauritus first colonizing Isla Gorgona and then back-colonizing the mainland after ancestral N. biporcatus populations had been extirpated. In this way, Isla Gorgona may have served as a refugium for at least one species, but perhaps also for N. medemi. While N. biporcatus is still wide-ranging for Neotropical anoles, the recently elevated N. parvauritus represents what would have been a disjunct portion of its range. This pattern may carry over to other lineages of Neotropical reptiles as we note that there are also disjunct populations of the colubrid snake Leptophis ahaetulla in Northwest Ecuador and Isla Gorgona [99], which are yet to be analyzed in a phylogeographic context.
4.3. Natural History Observations
Both D. agassizi and N. townsendi can be described as ecological generalists unlike solitary Lesser Antillean anoles [100]. Norops townsendi appears to clearly conform to this generalization, given that is was found ubiquitously on Isla Cocos in terms of perch height, diameter, and type, as briefly noted by Carpenter [38]. Dactyloa agassizi has distinctly unique behavioral characteristics that challenge the original meaning of generalist as applied to the Lesser Antillean anoles. The food web of Isla Malpelo appears to be very tight given the lack of vegetation and scarcity of obvious food resources for all the island inhabitants. Everything seems to revolve around the birds that visit the island, particularly the Nazca Booby (Sula granti). Any food dropped by the birds, and all waste products were immediately consumed by all D. agassizi, as well as Diploglossus millepunctatus lizards and Geocarcinus malpilensis crabs which congregate en masse around these resources. The Geocarcinus and Diploglossus attempt to capture any D. agassizi that venture too close [34], pers. obs., and the crabs will also consume eggs of all species [34]. Rand et al. [34] and Wolda [31] reported that D. agassizi ate both insects and seeds, and that individuals would quickly consume potential food items revealed when rocks were turned over. All D. agassizi departed from typical anole behavior in being very curious, climbing on, and licking the observers (clearly investigating food possibilities), never displaying any defensive behaviors or territoriality. Therefore, insofar as their environment allowed them, D. agassizi appeared to be an ecological generalist. Isla Malpelo is unlike most other islands that are home to anoles, so it is not surprising that D. agassizi departs from the general patterns observed for solitary Caribbean anoles.
4.4. Conservation of Island Fauna
While these Pacific Island anoles represent novel diversity unseen elsewhere, it is fortunate that all three islands are strictly controlled and monitored, virtually ensuring the conservation of the species. Tourists are somewhat rare to both Islas Cocos and Malpelo, and mostly consist of individuals seeking to scuba dive in the waters around the islands. It requires special permission to step onto the islands and is virtually impossible to be allowed onto Malpelo unless undertaking research. More tourists visit Isla Gorgona, which is also considerably closer to the mainland and faster to visit. Even then, they are only allowed to walk along trails accompanied by a guide and walk within the grounds of the former prison which is slowly being taken over by the forest. These strict controls are highly beneficial to the conservation of each island’s flora and fauna and no development of any kind is allowed. Thus, the future seems cautiously bright for the conservation status of these islands.
Only one species (D. agassizi) has estimates of population size [32], suggesting 140,000–206,000 individuals. None of the seven species of anoles discussed in this study has received any IUCN designation. However, we currently have no reason to believe any of these species/populations is currently at a high risk of extinction/extirpation because of the restricted access of these islands by the Colombian and Costa Rican governments. It is important that these policies continue, as island species are generally more predisposed to extirpation because of high endemism and smaller population sizes. [6]. In particular, tropical islands should be a priority in conservation since they serve as hotspots for biodiversity in many clades [101,102]. Anthropogenic disturbances have contributed to the extinction of other oceanic taxa [103,104], so the protection of these islands and their native ecosystems needs to continue for the conservation of not just these anoles but all flora and fauna on these islands.
5. Conclusions
Herein, we have estimated the divergence from the mainland for seven species of anole lizards in the ETP. Dactyloa agassizi and N. townsendi both diverge prior to the emergence of their islands (Islas Malpelo and Cocos respectively), and display sexual size dimorphism, similar to Caribbean relatives. Norops townsendi additionally displayed divergent patterns of morphological evolution in different populations across Isla Cocos. All lineages on Isla Gorgona appear to originate within the past 10 My, but geologic history of past connections with the mainland is unclear. Three of the species D. gorgonae, D. princeps, and N. medemi display ecological partitioning. This biogeographic and ecomorphological work set the stage for future questions in the realm of island biogeography and community ecology.
Supplementary Materials
The following are available online at https://www.mdpi.com/1424-2818/11/9/141/s1, Figure S1: Bayesian phylogeny as calculated by using a mutation rate only; Figure S2: Bayesian phylogeny as calculated by both mutation rate and fossil calibrations, displaying node bars for 95% confidence intervals; Table S1: Species used in phylogenetic analyses with Genbank numbers; Table S2: Locality data for all Norops biporcatus and N. parvauritus samples used in this study; Table S3: Ecological and morphological data from Dactyloa agassizi used in this study; Table S4: Ecological and morphological data from Norops townsendi used in this study. Table S5: Ecological and morphological data from five species of anoles on Isla Gorgona used in this study.
Author Contributions
K.E.N. designed the study. J.G.P., S.E.B., M.M.W., and K.E.N. collected specimens. J.G.P., S.E.B., E.P., and K.E.N. conducted labwork. J.G.P. and K.E.N. conducted analysis. J.G.P. and K.E.N. led the writing with contributions from all authors.
Funding
This work was supported in part by a grant K.E.N. (labwork, analysis and writing: NSF DEB 0949359 and internal funding from Central Michigan University).
Acknowledgments
The authors would like to thank Andrew C. Kramer and A. Scott McNaught for statistical advice. We would like to thank Mahmood Sasa for logistical and permitting support in Costa Rica, particularly for Isla Cocos. We also thank our Colombian colleagues for logistical support and permitting, especially in accessing Islas Gorgona and Malpelo. We thank Natasha Bloch and Daniel Medina for field assistance on Isla Gorgona, as well as Jeremy Gibson-Brown and the Nicholson lab for editorial comments. Mainland sampling of Norops biporcatus was assisted by Sarah Burton, Jenny Gubler, David Laurencio, Lenin Obando, and Javier Sunyer. Additional tissues were provided/collected by Carlos A. Andino, Atanasio Baldonado, Cesar A Cerrato-Mendoza, Sebastian Charley, Wes Chun, Ignacio Cruz, Gabriela Diaz, Stephen Doucette-Riise, Sergio Gonzalez, Lorraine P Ketzler, Francisco Lopez, Ileana R Luque-Montes, Melissa Medina-Flores, Aaron Mendoza, Wendy Naira-Mejia, Ronald Picado, Jay M. Savage, Josiah H Townsend, Scott L. Travers, Laurie J. Vitt and Larry David Wilson. This study conformed to all IACUC regulations and specimens were collected with the appropriate permits (in Costa Rica: permit number ACMIC-002-003, Resolucíones 239-2008-SINAC, 040–2009-SINAC, 017-2010-SINAC and 005–2011-SINAC and MINAET Permit no. 029–2011-SINAC; in Colombia: resolución número 2388, in Nicaragua under DGPN/DB-10–2010 and in Panama under ANAM Scientific Permit No. SEX/A-50-12).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Chiba, S. Ecological and morphological patterns in communities of land snails of the genus Mandarina from the Bonin Islands. J. Evol. Biol. 2004, 17, 131–143. [Google Scholar] [CrossRef] [PubMed]
- Gillespie, R. Community assembly through adaptive radiation in Hawaiian spiders. Science 2004, 303, 356–359. [Google Scholar] [CrossRef] [PubMed]
- Parent, C.E.; Caccone, A.; Petren, K. Colonization and diversification of Galápagos terrestrial fauna: A phylogenetic and biogeographical synthesis. Philos. Trans. R. Soc. B Biol. Sci. 2008, 363, 3347–3361. [Google Scholar] [CrossRef] [PubMed]
- Carlquist, S.J. Island Biology; Columbia University Press: New York, NY, USA, 1974. [Google Scholar]
- Hendriks, K.P.; Alciatore, G.; Schilthuizen, M.; Etienne, R.S. Phylogeography of Bornean land snails suggests long-distance dispersal as a cause of endemism. J. Biogeogr. 2019, 46, 932–944. [Google Scholar] [CrossRef]
- Whittaker, R.J.; Fernández-Palacios, J.M. Island Biogeography: Ecology, Evolution, and Conservation, 2nd ed.; Oxford University Press: Oxford, UK, 2007. [Google Scholar]
- Ali, J.R.; Aitchison, J.C. Exploring the combined role of eustasy and oceanic island thermal subsidence in shaping biodiversity on the Galápagos. J. Biogeogr. 2014, 41, 1227–1241. [Google Scholar] [CrossRef]
- Williams, E.E. A critique of Guyer and Savage (1986): Cladistic relationships among anoles (Sauria: Iguanidae): Are the data available to reclassify the anoles? In Biogeography of the West Indies: Past, Present, and Future; Sandhill Crane Press: Gainesville, FL, USA, 1989; pp. 286–341. [Google Scholar]
- Crother, B.I.; Parenti, L.R. Assumptions Inhibiting Progress in Comparative Biology; CRC Press: Boca Raton, FL, USA, 2016. [Google Scholar]
- Guyer, C.; Savage, J.M. Cladistic relationships among anoles (Sauria: Iguanidae). Syst. Zool. 1986, 35, 509–531. [Google Scholar] [CrossRef]
- Nicholson, K.E.; Crother, B.I.; Guyer, C.; Savage, J.M. Translating a clade-based classification into one that is valid under the international code of zoological nomenclature: The case of the lizards of the family Dactyloidae (Order Squamata). Zootaxa 2018, 4461, 573–586. [Google Scholar] [CrossRef] [PubMed]
- Uetz, P.; Freed, P.; Hošek, J. The Reptile Database. Available online: http://www.reptile-database.org (accessed on 29 June 2019).
- Losos, J.B. Lizards in an Evolutionary Tree: Ecology and Adaptive Radiation of Anoles; University of California Press: Berkeley, CA, USA, 2009. [Google Scholar]
- Irschick, D.L.; Vitt, L.J.; Zani, P.A.; Losos, J.B. A comparison of evolutionary radiations in mainland and West Indian Anolis lizards. Ecology 1997, 78, 2191–2203. [Google Scholar] [CrossRef]
- Irschick, D.L.; Losos, J.B. A comparative analysis of the ecological significance of maximal locomotor performance in Caribbean Anolis lizards. Evolution 1998, 52, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Losos, J.B. The evolution of convergent community structure in Caribbean Anolis communities. Syst. Biol. 1992, 41, 403–420. [Google Scholar] [CrossRef]
- Losos, J.B.; Warheit, K.I.; Schoener, T.W. Adaptive differentiation following experimental island colonization in Anolis lizards. Nature 1997, 387, 70–73. [Google Scholar] [CrossRef]
- Williams, E.E. The ecology of colonization as seen in the zoogeography of Anolis lizards on small islands. Q. Rev. Biol. 1969, 44, 345–389. [Google Scholar] [CrossRef]
- Hedges, S.B.; Hass, C.A.; Maxson, L.R. Caribbean biogeography: Molecular evidence for dispersal in West Indian terrestrial vertebrates. Proc. Natl. Acad. Sci. USA 1992, 89, 1909–1913. [Google Scholar] [CrossRef]
- Poe, S.; Nieto-Montes de Oca, A.; Torres-Carvajal, O.; de Queiroz, K.; Velasco, J.A.; Truett, B.; Gray, L.N.; Ryan, M.J.; Köhler, G.; Ayala-Varela, F.; et al. A phylogenetic, biogeographic, and taxonomic study of all extant species of Anolis (Squamata; Iguanidae). Syst. Biol. 2017, 66, 663–697. [Google Scholar] [CrossRef]
- Nicholson, K.E.; Crother, B.I.; Guyer, C.; Savage, J.M. It is time for a new classification of anoles. Zootaxa 2012, 3477, 1–108. [Google Scholar] [CrossRef]
- Schoener, T.W. Size patterns in West Indian Anolis lizards: I. size and species diversity. Syst. Biol. 1969, 18, 386–401. [Google Scholar] [CrossRef]
- Blankers, T.; Townsend, T.M.; Pepe, K.; Reeder, T.W.; Wiens, J.J. Contrasting global-scale evolutionary radiations: Phylogeny, diversification, and morphological evolution in the major clades of iguanian lizards. Biol. J. Linn. Soc. 2013, 108, 127–143. [Google Scholar] [CrossRef]
- Román-Palacios, C.; Tavera, J.; Castañeda, D.R. When did anoles diverge? An analysis of multiple dating strategies. Mol. Phylogenet. Evol. 2018, 127, 655–668. [Google Scholar] [CrossRef]
- Poe, S.; Goheen, J.R.; Hulebak, E.P. Convergent exaptation and adaptation in solitary island lizards. Proc. R. Soc. B Biol. Sci. 2007, 274, 2231–2237. [Google Scholar] [CrossRef] [Green Version]
- Hoernle, K.; van den Bogaard, P.; Werner, R.; Lissinna, B.; Hauff, F.; Alvarado, G.; Garbe-Schönberg, D. Missing history (16–17 Ma) of the Galápagos hotspot: Implications for the tectonic and biological evolution of the Americas. Geology 2002, 30, 795–798. [Google Scholar] [CrossRef]
- Stead, J.A. Field observations of the geology of Malpelo Island. Smithson. Contrib. Zool. 1975, 176, 17–26. [Google Scholar]
- Gorman, G.C.; Chorba, T.L. Terrestrial biology of Malpelo Island: A historical review. Smithson. Contrib. Zool. 1975, 176, 9–12. [Google Scholar]
- Hertlein, L.G. Contribution to the biogeography of Cocos Island, including a bibliography. Proc. Calif. Acad. Sci. USA 1963, 32, 219–289. [Google Scholar]
- Kiester, A.R.; Hoffman, J.A. Reconnaissance and mapping of Malpelo Island. Smithson. Contrib. Zool. 1975, 176, 13–20. [Google Scholar]
- Wolda, H. The ecosystem on Malpelo Island. Smithson. Contrib. Zool. 1975, 176, 21–26. [Google Scholar]
- López-Victoria, M.; Wolters, V.; Werding, B. Nazca Booby (Sula granti) inputs maintain the terrestrial food web of Malpelo Island. J. Ornithol. 2009, 150, 865–870. [Google Scholar] [CrossRef]
- López-Victoria, M. The lizards of Malpelo (Colombia): Some topics on their ecology and threats. Caldesia 2006, 28, 129–134. [Google Scholar]
- Rand, A.S.; Gorman, G.C.; Rand, W.M. Natural history, behavior, and ecology and Anolis agassizi. Smithson. Contrib. Zool. 1975, 176, 27–38. [Google Scholar]
- Castañeda, M.D.R.; de Queiroz, K. Phylogenetic relationships of the Dactyloa clade of Anolis lizards based on nuclear and mitochondrial DNA sequence data. Mol. Phylogenetics Evol. 2011, 61, 784–800. [Google Scholar] [CrossRef]
- Castañeda, M.D.R.; de Queiroz, K. Phylogeny of the Dactyloa clade of Anolis lizards: New insights from combining morphological and molecular data. Bull. Mus. Comp. Zool. 2013, 160, 345–398. [Google Scholar] [CrossRef]
- Prates, I.; Rodrigues, M.T.; Melo-Sampaio, P.R.; Carnaval, A.C. Phylogenetic relationships of Amazonian anole lizards (Dactyloa): Taxonomic implications, new insights about phenotypic evolution and the timing of diversification. Mol. Phylogenetics Evol. 2015, 82, 258–268. [Google Scholar] [CrossRef]
- Carpenter, C.C. The display of the Cocos Island anole. Herpetologica 1965, 21, 256–260. [Google Scholar]
- Castillo, P.; Batiza, R.; Vanko, D.; Malavassi, E.; Barquero, J.; Fernandez, E. Anomalously young volcanoes on old hot-spot traces. I. Geology and petrology of Cocos Island. Geology and petrology of Cocos Island. Geol. Soc. Am. Bull. 1998, 100, 1400–1414. [Google Scholar] [CrossRef]
- Poe, S. Phylogeny of anoles. Herpetol. Monogr. 2004, 18, 37–89. [Google Scholar] [CrossRef]
- Grisales-Martínez, F.A.; Velasco, J.A.; Bolívar, W.; Williams, E.E.; Daza, J.M. The taxonomic and phylogenetic status of some poorly known Anolis species from the Andes of Colombia with the description of a nomen nudum taxon. Zootaxa 2017, 4303, 213–230. [Google Scholar] [CrossRef]
- Nicholson, K.E.; Guyer, C.; Phillips, J.G. Biogeographic origin of mainland Norops (Squamata: Dactyloidae). In Assumptions Inhibiting Progress in Comparative Biology; CRC Press: Boca Raton, FL, USA, 2017; pp. 169–184. [Google Scholar]
- Phillips, J.G.; Deitloff, J.; Guyer, C.; Hutteman, S.; Nicholson, K.E. Biogeography and evolution of a widespread Central American lizard species complex: Norops humilis, (Squamata: Dactyloidae). BMC Evol. Biol. 2015, 15, 143. [Google Scholar] [CrossRef]
- Castro, F.; Ayala, S.; Carvajal, H. Los saurios de las islas Gorgona y Gorgonilla. In Gorgona; Universidad de los Andes: Cundinamarca, Colombia, 1979; pp. 189–218. [Google Scholar]
- UASPNN. El Sistema de Parques Nacionales Naturales de Colombia; Unidad Administrativa Especial del Sistema de Parques Nacionales Naturales (UASPNN), Ministerio del Medio Ambiente. Editorial Nomos: Bogotá, Colombia, 1998. [Google Scholar]
- Echeverría, L.M.; Aitken, B. Pyroclastic rocks: Another manifestation of ultramafic volcanism of Gorgona Island. Colombia. Contrib. Mineral. Petrol. 1986, 92, 428–436. [Google Scholar] [CrossRef]
- Gomez, H. Algunos aspectos neotectónicos hacia el suroeste del Litoral Pacífico colombiano. Rev. CIAF 1986, 11, 281–296. [Google Scholar]
- Kerr, A.C. La Isla de Gorgona, Colombia: A petrological enigma? Lithos 2005, 84, 77–101. [Google Scholar] [CrossRef]
- Serrano, L.; Ferrari, L.; Martínez, M.L.; Petrone, C.M.; Jaramillo, C. An integrative geologic, geochronologic and geochemical study of Gorgona Island, Colombia: Implications for the formation of the Caribbean Large Igneous Province. Earth Planet. Sci. Lett. 2011, 309, 324–336. [Google Scholar] [CrossRef]
- Von Prahl, H.; Guhl, F.; Grögl, M. Gorgona; Futura Groupo Editorial Ltd.: Bogota, Colombia, 1979. [Google Scholar]
- Aguirre, J.; Rangel, O. La isla Gorgona y sus ecosistemas. In Colombia Pacífico; Fondo para la Protección del Medio Ambiente (FEN): Bogotá, Colombia, 1993; Volume I, pp. 106–170. [Google Scholar]
- Alberico, M. Biogeografía terrestre. Capitulo XI. In Isla de Gorgona; Banco Popular: Bogotá, Colombia, 1986; pp. 223–244. [Google Scholar]
- Murillo, M.T.; Lozano, G. Hacia la realización de una flórula del Parque Nacional Natural Islas de Gorgona y Gorgonilla, Cauca, Colombia. Rev. Acad. Col. Cie. Exactas Físicas y Naturales 1989, 12, 277–304. [Google Scholar]
- Yockteng, R.; Cavelier, J. Diversidad y mecanismos de dispersión de árboles de la Isla Gorgona y de los bosques húmedos tropicales del Pacífico colombo-ecuatoriano. Rev. Biol. Trop. 1998, 46, 45–53. [Google Scholar]
- Armstead, J.V.; Ayala-Varela, F.; Torres-Carvajal, O.; Ryan, M.J.; Poe, S. Systematics and ecology of Anolis biporcatus (Squamata: Iguanidae). Salamandra 2017, 53, 285–293. [Google Scholar]
- Köhler, G.; McCranie, J.R.; Nicholson, K.E.; Kreutz, J. Geographic variation in hemipenial morphology in Norops humilis (Peters 1863), and the systematic status of Norops quaggulus (Cope 1885) (Reptilia, Squamata, Polychrotidae). Senckenbergiana Biol. 2003, 82, 213–222. [Google Scholar]
- Selander, R.K. Sexual dimorphism and differential niche utilization in birds. Condor 1966, 68, 113–151. [Google Scholar] [CrossRef]
- Clegg, S.M.; Owens, P.F. The ‘island rule’ in birds: Medium body size and its ecological explanation. Proc. R. Soc. Lond. Ser. B Biol. Sci. 2002, 269, 1359–1365. [Google Scholar] [CrossRef]
- Dayan, T.; Simberloff, D. Size patterns among competitors: Ecological character displacement and character release in mammals, with special reference to island populations. Mammal Rev. 1998, 28, 99–124. [Google Scholar] [CrossRef]
- Schoener, T.W. The ecological significance of sexual dimorphism in size in the lizard Anolis conspersus. Science 1967, 155, 474–477. [Google Scholar] [CrossRef]
- Schoener, T.W. Size patterns in West Indian Anolis lizards. II. Correlations with the sizes of particular sympatric species-displacement and convergence. Am. Nat. 1970, 104, 155–174. [Google Scholar] [CrossRef]
- Velasco, J.A.; Poe, S.; González-Salazar, C.; Flores-Villela, O. Solitary ecology as a phenomenon extending beyond insular systems: Exaptive evolution in Anolis lizards. Biol. Lett. 2019, 15, 20190056. [Google Scholar] [CrossRef]
- Bouckaert, R.; Heled, J.; Kühnert, D.; Vaughan, T.; Wu, C.; Xie, D.; Suchard, M.A.; Rambaut, A.; Drummond, A.J. BEAST 2: A software platform for Bayesian evolutionary analysis. PLoS Comput. Biol. 2014, 10, e1003537. [Google Scholar]
- Rambaut, A.; Suchard, M.A.; Xie, D.; Drummond, A.J. Tracer v.1.6. 2014. Available online: http://tree.bio.ed.ac.uk/software/tracer/ (accessed on 9 August 2018).
- Macey, J.R.; Schulte II, J.A.; Ananjeva, N.B.; Larson, A.; Rastegar-Pouyani, N.; Shammakov, S.M.; Papenfuss, T.J. Phylogenetic relationships among Agamid Lizards of the Laudakia caucasia species group: Testing hypotheses of biogeographic fragmentation and an area cladogram for the Iranian Plateau. Mol. Phylogenetics Evol. 1998, 10, 118–131. [Google Scholar] [CrossRef]
- Glor, R.E.; Kolbe, J.J.; Powell, R.; Larson, A.; Losos, J.B. Phylogenetic analysis of ecological and morphological diversification in Hispaniolan trunk-ground anoles (Anolis cybotes group). Evolution 2003, 57, 2383–2397. [Google Scholar] [CrossRef]
- Glor, R.E.; Losos, J.B.; Larson, A. Out of Cuba: Overwater dispersal and speciation among lizards in the Anolis carolinensis subgroup. Mol. Ecol. 2005, 14, 2419–2432. [Google Scholar] [CrossRef]
- Guarnizo, C.E.; Werneck, F.P.; Giugliano, L.G.; Santos, M.G.; Fenker, J.; Sousa, L.; D’Angiolella, A.B.; dos Santos, A.R.; Strüssmann, C.; Rodrigues, M.T.; et al. Cryptic lineages and diversification of an endemic lizard (Squamata, Dactyloidae) of the Cerrado hotspot. Mol. Phylogenetics Evol. 2016, 94, 279–289. [Google Scholar] [CrossRef]
- Ng, J.; Glor, R.E. Genetic differentiation among populations of a Hispaniolan trunk anole that exhibit geographical variation in dewlap color. Mol. Ecol. 2011, 20, 4302–4317. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, R.G.; Strickland, T.R.; Kolbe, J.J.; Falk, B.G.; Perry, G.; Revell, L.J.; Losos, J.B. Archipelagic genetics in a widespread Caribbean anole. J. Biogeogr. 2017, 44, 2631–2647. [Google Scholar] [CrossRef]
- Torres-Carvajal, O.; de Queiroz, K. Phylogeny of hoplocercine lizards (Squamata: Iguania) with estimates of relative divergence times. Mol. Phylogenetics Evol. 2009, 50, 31–43. [Google Scholar] [CrossRef]
- Zheng, Y.; Peng, R.; Kuro-o, M.; Zeng, X. Exploring patterns and extent of bias in estimating divergence time from mitochondrial DNA sequence data in a particular lineage: A case study of salamanders (Order Caudata). Mol. Biol. Evol. 2011, 28, 2521–2535. [Google Scholar] [CrossRef]
- Poe, S.; Scarpetta, S.; Schaad, E.W. A new species of Anolis (Squamata: Iguanidae) from Panama. Amphib. Reptile Conserv. 2015, 9, 1–13. [Google Scholar]
- Macedonia, J.M.; Clark, D.L. Headbob display structure in the naturalized Anolis lizards of Bermuda: Sex, context, and population effects. J. Herpetol. 2003, 37, 266–277. [Google Scholar] [CrossRef]
- Thorpe, R.S.; Barlow, A.; Malhotra, A.; Surget-Groba, Y. Widespread parallel population adaptation to climate variation across a radiation: Implications for adaptation to climate change. Mol Ecol. 2015, 24, 1019–1030. [Google Scholar] [CrossRef] [PubMed]
- Calderón-Espinosa, M.L.; Forero, A.B. Morphological diversification in solitary endemic anoles: Anolis concolor and Anolis pinchoti from San Andrés and Providence Islands, Colombia. South Am. J. Herpetol. 2011, 6, 205–210. [Google Scholar] [CrossRef]
- Gartner, G.E.A.; Gamble, T.; Jaffe, A.L.; Harrison, A.; Losos, J.B. Left-right dewlap asymmetry and phylogeography of Anolis lineatus on Aruba and Curaçao. Biol. J. Linn. Soc. 2013, 110, 409–426. [Google Scholar] [CrossRef]
- Williams, E.E. Ecomorphs, faunas, island size, and diverse end points in island radiations of Anolis. In Lizard Ecology: Studies of a Model Organism; Harvard University Press: Cambridge, MA, USA, 1983; pp. 326–370. [Google Scholar]
- Savage, J.M.; Talbot, J.J. The giant anoline lizards of Costa Rica and western Panama. Copeia 1978, 1978, 480–492. [Google Scholar] [CrossRef]
- Williams, E.E. New or problematic Anolis from Colombia. V. Anolis danieli, a new species of the latifrons species group and a reassessment of Anolis apollinaris Boulenger, 1919. Breviora 1988, 489, 1–25. [Google Scholar]
- Farris, D.W.; Jaramillo, C.; Bayona, G.; Restrepo-Moreno, S.A.; Montes, C.; Cardona, A.; Mora, A.; Speakman, R.J.; Glascock, M.D.; Valencia, V. Fracturing of the Panamanian Isthmus during initial collision with South America. Geology 2011, 39, 1007–1010. [Google Scholar] [CrossRef]
- Montes, C.; Cardona, A.; Jaramillo, C.; Pardo, A.; Silva, J.C.; Valencia, V.; Ayala, C.; Pérez-Angel, L.C.; Rodriguez-Parra, L.A.; Ramirez, V.; et al. Middle Miocene closure of the Central American seaway. Science 2012, 348, 226–229. [Google Scholar] [CrossRef]
- Elmer, K.R.; Bonett, R.M.; Wake, D.B.; Lougheed, S.C. Early Miocene origin and cryptic diversification of South American salamanders. BMC Evol. Biol. 2013, 13, 59. [Google Scholar] [CrossRef]
- Montes, C.; Cardona, A.; McFadden, R.; Morón, S.E.; Silva, C.A.; Restrepo-Moreno, S.; Bayona, G.A. Evidence for middle Eocene and younger land emergence in central Panama: Implications for Isthmus closure. Geol. Soc. Am. Bull. 2012, 124, 780–799. [Google Scholar] [CrossRef]
- Coates, A.G.; Obando, J.A. The geologic evolution of the Central American isthmus. In Evolution and Environment in Tropical America; University of Chicago Press: Chicago, IL, USA, 1996; pp. 21–56. [Google Scholar]
- MacFadden, B.J. Extinct mammalian biodiversity of the ancient New World tropics. Trends Ecol. Evol. 2006, 21, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Censky, E.J.; Hodge, K.; Dudley, J. Over-water dispersal of lizards due to hurricanes. Nature 1988, 395, 556. [Google Scholar] [CrossRef]
- Etheridge, R.E. The Relationships of the Anoles (Reptilia: Sauria: Iguanidae): An Interpretation Based on Skeletal Morphology. Ph.D. Thesis, University Microfilms, Ann Arbor, MI, USA, 1959. [Google Scholar]
- Savage, J.M.; Guyer, C. Infrageneric classification and species composition of the anole genera, Anolis, Ctenonotus, Dactyloa, Norops, and Semiurus (Sauria: Iguanidae). Amphib. Reptil. 1989, 10, 105–116. [Google Scholar] [CrossRef]
- Caccone, A.; Gibbs, J.P.; Ketmaier, V.; Suatoni, E.; Powell, J.R. Origin and evolutionary relationships of giant Galapagos tortoises. Proc. Natl. Acad. Sci. USA 1999, 96, 13223–13228. [Google Scholar] [CrossRef] [PubMed]
- Caccone, A.; Gentile, G.; Gibbs, J.P.; Fritts, T.H.; Snell, H.L.; Betts, J.; Powell, J.R. Phylogeography and history of giant Galapagos tortoises. Evolution 2002, 56, 2052–2066. [Google Scholar]
- Huber, M.; Caballero, R. Eocene El Nino: Evidence for robust tropical dynamics in the “hothouse”. Science 2003, 299, 877–881. [Google Scholar] [CrossRef] [PubMed]
- Fedorov, A.V.; Dekens, P.S.; McCarthy, M.; Ravelo, A.C.; Barreiro, M.; Pacanowski, R.C.; Philander, S.G. The Pliocene paradox (mechanisms for a permanent El Niño). Science 2006, 312, 1485–1489. [Google Scholar] [CrossRef] [PubMed]
- Hartley, A.J.; Chong, G.; Houston, J.; Mather, A.E. 150 million years of climate stability: Evidence from the Atacama Desert, northern Chile. J. Geol. Soc. 2005, 162, 421–444. [Google Scholar] [CrossRef]
- Bessudo, S.; Soler, G.A.; Klimley, A.P.; Ketchum, J.T.; Hearn, A.; Arauz, R. Residency of the scalloped hammerhead shark (Sphyrna lewini) at Malpelo Island and evidence of migration to other islands in the Eastern Tropical Pacific. Environ. Biol. Fishes 2011, 91, 165–176. [Google Scholar] [CrossRef]
- Jiménez, L.; Acosta, A.; Chong, N. Population structure of Megabalanus peninsularis in Malpelo Island, Colombia. Rev. Biol. Trop. 2016, 51, 461–468. [Google Scholar]
- O’Connor, M.; Bruno, J.; Gaines, S. Temperature control of larval dispersal and the implications for marine ecology, evolution, and conservation. Proc. Natl. Acad. Sci. USA 2007, 104, 1266–1271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orellana-Rovirosa, F.; Richards, M. Emergence/subsidence histories along the Carnegie and Cocos Ridges and their bearing upon biological speciation in the Galápagos. Geochem. Geophys. Geosystems 2018, 19, 4099–4129. [Google Scholar] [CrossRef]
- Savage, J.M. The Amphibians and Reptiles of Costa Rica: A Herpetofauna Between Two Continents, Between Two Seas; University of Chicago Press: Chicago, IL, USA, 2002; p. 954. [Google Scholar]
- Losos, J.B.; de Queiroz, K. Evolutionary consequences of ecological release in Caribbean Anolis lizards. Biol. J. Linn. Soc. 1997, 61, 459–483. [Google Scholar] [CrossRef]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fonseca, G.A.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Cubas, J.; Irl, S.D.; Villafuerte, R.; Bello-Rodríguez, V.; Rodríguez-Luengo, J.L.; del Arco, M.; Martín-Esquivel, J.L.; González-Mancebo, J.M. Endemic plant species are more palatable to introduced herbivores than non-endemics. Proc. R. Soc. Lond. Ser. B Biol. Sci. 2019, 286, 20190136. [Google Scholar] [CrossRef]
- Regnier, C.; Achaz, G.; Lambert, A.; Cowie, R.H.; Bouchet, P.; Fontaine, B. Mass extinction in poorly known taxa. Proc. Natl. Acad. Sci. USA 2015, 112, 7761–7766. [Google Scholar] [CrossRef] [Green Version]
- Solem, A. How many Hawaiian land snail species are left and what can we do for them? Bish. Mus. Occas. Pap. 1990, 30, 27–40. [Google Scholar]
Figure 1.
Seven species of anoles found on the islands in the Eastern Tropical Pacific (ETP). Top row, left to right: Dactyloa agassizi (Isla Malpelo), Dactyloa gorgonae (Isla Gorgona), Norops townsendi (Isla Cocos). Bottom row: Norops medemi, Norops parvauritus, Dactyloa princeps, Dactyloa chocorum (all Isla Gorgona).
Figure 1.
Seven species of anoles found on the islands in the Eastern Tropical Pacific (ETP). Top row, left to right: Dactyloa agassizi (Isla Malpelo), Dactyloa gorgonae (Isla Gorgona), Norops townsendi (Isla Cocos). Bottom row: Norops medemi, Norops parvauritus, Dactyloa princeps, Dactyloa chocorum (all Isla Gorgona).

Figure 2.
Three islands that are home to the anoles in the ETP. (a) Isla Malpelo, (b) Isla Cocos, (c) Isla Gorgona.
Figure 2.
Three islands that are home to the anoles in the ETP. (a) Isla Malpelo, (b) Isla Cocos, (c) Isla Gorgona.

Figure 3.
Map of the Eastern Tropical Pacific (ETP) with the three islands in this study. Mainland sampling localities for Norops biporcatus and N. parvauritus included in the molecular phylogeny are also shown.
Figure 3.
Map of the Eastern Tropical Pacific (ETP) with the three islands in this study. Mainland sampling localities for Norops biporcatus and N. parvauritus included in the molecular phylogeny are also shown.

Figure 4.
Bayesian reconstruction of Dactyloa and Norops anoles dated using a combination of fossil calibrations and a mtDNA mutation rate. All lineages of Pacific Island anoles are colored, and posterior values are present on all nodes with <0.95 probability. (A) Norops clades pertinent to this study. (B) Dactyloa clades pertinent to this study.
Figure 4.
Bayesian reconstruction of Dactyloa and Norops anoles dated using a combination of fossil calibrations and a mtDNA mutation rate. All lineages of Pacific Island anoles are colored, and posterior values are present on all nodes with <0.95 probability. (A) Norops clades pertinent to this study. (B) Dactyloa clades pertinent to this study.


Figure 5.
Range of divergence dates (95% CI) for each of the five anole lineages on Isla Gorgona from a mainland ancestor. Mean node age is represented by an open circle.
Figure 5.
Range of divergence dates (95% CI) for each of the five anole lineages on Isla Gorgona from a mainland ancestor. Mean node age is represented by an open circle.

Figure 6.
The Isla Gorgona anole community depicted by habitat in accordance with ecomode designation by Nicholson et al. [21]. Figure modified from [13], which was modified from [78].


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Mean dates and 95% confidence intervals for select nodes relevant to Pacific Island anoles, using multiple dating methods (fossil-fossil calibrations + mutation rate; rate = mutation rate alone).
Table 1.
Mean dates and 95% confidence intervals for select nodes relevant to Pacific Island anoles, using multiple dating methods (fossil-fossil calibrations + mutation rate; rate = mutation rate alone).
| Node | Fossil | 95% | Rate | 95% |
|---|---|---|---|---|
| Dactyloa agassizi + Dactyloa insignis/Dactyloa microtus | 29.2 | 21.6–37.2 | 33.4 | 22.4–36.4 |
| Dactyloa chocorum from Isla Gorgona vs. mainland | 6.3 | 3.8–9.9 | 6.7 | 4.1–9.1 |
| All Dactyloa chocorum coalescence | 7.6 | 4.8–11.2 | 7.3 | 5.3–10.5 |
| Dactyloa princeps from Isla Gorgona vs. mainland | 4.5 | 2.7–7.1 | 3.0 | 2.8–6.5 |
| All Dactyloa princeps coalescence | 7.3 | 5.1–10.5 | 7.4 | 5.2–9.4 |
| Dactyloa chloris + Dactyloa gorgonae | 8.6 | 4.9–13.2 | 12.8 | 5.3–12.2 |
| Norops medemi coalescence (within Isla Gorgona) | 6.6 | 5.2–9.6 | 6.1 | 5.0–8.0 |
| Norops medemi + Norops urraoi | 9.6 | 10.2–16.3 | 8.1 | 7.7–11.8 |
| Norops townsendi coalescence (Isla Cocos vs. Manuelita) | 3.5 | 2.2–4.7 | 3.1 | 2.4–4.9 |
| Norops poecilopus + Norops townsendi | 7.6 | 5.1–10.1 | 6.7 | 5.6–10.0 |
| Norops parvauritus coalescence | 3.2 | 2.4–6.6 | 3.8 | 2.0–4.8 |
| Norops biporcatus coalescence | 14.7 | 11.6–20.7 | 12.5 | 11.4–18.3 |
| Norops biporcatus + Norops parvauritus | 24.4 | 20.2–32.7 | 21.3 | 19.4–29.7 |
Table 2.
Average morphological measurements by sex for four species of Pacific Island anoles. (SVL = snout-vent length, # lamelle = # of lamellar rows). P-values from pairwise t-tests between the sexes are listed below the means. Standard deviation is included for each mean value in parentheses. Significant values after Bonferroni correction are bolded and italicized.
Table 2.
Average morphological measurements by sex for four species of Pacific Island anoles. (SVL = snout-vent length, # lamelle = # of lamellar rows). P-values from pairwise t-tests between the sexes are listed below the means. Standard deviation is included for each mean value in parentheses. Significant values after Bonferroni correction are bolded and italicized.
| Sex/Species | N | SVL (mm) | Mass (g) | # Lamelle | PH (m) | PD (m) | # Mites |
|---|---|---|---|---|---|---|---|
| Isla Malpelo | |||||||
| Dactyloa agassizi, female | 18 | 67.3 (± 5.9) | 7.6 (± 2.2) | 36.8 (± 1.7) | 1.4 (± 1.2) | 0.3 (± 0.3) | |
| Dactyloa agassizi, male | 29 | 92.9 (± 9.0) | 24.1 (± 6.9) | 37.1 (± 1.7) | 1.5 (± 1.1) | 0.3 (± 0.4) | |
| p | <0.0001 | <0.0001 | 0.53 | 0.76 | 0.88 | ||
| Isla Cocos | |||||||
| Norops townsendi, female | 16 | 46.6 (± 4.2) | 2.1 (± 0.7) | 18.5 (± 1.6) | 0.9 (± 0.6) | 0.07 (± 0.06) | 5.2 (± 6.4) |
| Norops townsendi, male | 61 | 50.5 (± 4.0) | 2.6 (± 0.8) | 18.5 (± 1.2) | 1.4 (± 1.2) | 0.09 (± 1.0) | 7.9 (± 13.3) |
| p | 0.002 | 0.05 | 0.83 | 0.12 | 0.53 | 0.52 | |
| Isla Gorgona | |||||||
| Dactyloa princeps, female | 10 | 102.1 (± 14.0) | 21.1 (± 8.4) | 22.9 (± 0.9) | 2.4 (± 1.3) | 17.3 (± 9.6) | |
| Dactyloa princeps, male | 9 | 139.8 (± 7.9) | 49.1 (± 7.3) | 24.0 (± 1.1) | 3.6 (± 1.3) | 11.4 (± 4.9) | |
| p | <0.0001 | <0.0001 | 0.04 | 0.07 | 0.12 | ||
| Norops medemi, female | 5 | 51.0 (± 2.0) | 2.3 (± 0.2) | 15.0 (± 0.0) | 3.1 (± 2.3) | 5.2 (± 4.5) | |
| Norops medemi, male | 5 | 48.8 (± 3.9) | 1.9 (± 0.3) | 15.8 (± 0.5) | 2.0 (± 0.2) | 5.9 (± 7.7) | |
| p | 0.30 | 0.02 | 0.01 | 0.37 | 0.87 |
Table 3.
Average SVL by sex for solitary species of Dactyloa and Norops. Standard deviation is provided when available.
Table 3.
Average SVL by sex for solitary species of Dactyloa and Norops. Standard deviation is provided when available.
| Species | Male SVL (mm) | Female SVL (mm) | F/M Ratio | Reference |
|---|---|---|---|---|
| Dactyloa agassizi | 92.9 (± 9.0) | 67.3 (± 5.9) | 0.72 | This study |
| Dactyloa extrema | 65.9 | 52 | 0.79 | [74] |
| Dactyloa luciae | 72.6 | 52.1 | 0.72 | [75] |
| Dactyloa roquet | 67.3 (± 7.5) | 52.1 (± 6.3) | 0.77 | [22] |
| Norops concolor | 69.3 (± 0.7) | 50.1 (± 0.3) | 0.72 | [76] |
| Norops lineatus | 69.1 | 58.1 | 0.84 | [77] |
| Norops pinchoti | 47.7 (± 0.1) | 40.2 (± 0.2) | 0.84 | [76] |
| Norops townsendi | 50.5 (± 4.0) | 46.6 (± 4.2) | 0.92 | This study |
Table 4.
(A) Average morphological and ecological measurements for four populations of Norops townsendi on Isla Cocos. (B) Results for pairwise comparisons of all variables between each population. Significant differences following Bonferroni correction are bolded. (C) Results for pairwise comparisons of all variables between each population, while treating the Playa and Trail sites as a single population. Significant values after Bonferroni correction are bolded and italicized. (SVL = snout-vent length, Fore = forelimb length, Hind = hindlimb length, HW = head width, HH = head height, HL = head length, PH = perch height, PD = perch diameter).
Table 4.
(A) Average morphological and ecological measurements for four populations of Norops townsendi on Isla Cocos. (B) Results for pairwise comparisons of all variables between each population. Significant differences following Bonferroni correction are bolded. (C) Results for pairwise comparisons of all variables between each population, while treating the Playa and Trail sites as a single population. Significant values after Bonferroni correction are bolded and italicized. (SVL = snout-vent length, Fore = forelimb length, Hind = hindlimb length, HW = head width, HH = head height, HL = head length, PH = perch height, PD = perch diameter).
| Population | SVL (mm) | mass (g) | Fore (mm) | Hind (mm) | HW (mm) | HH (mm) | HL (mm) | # lamellae | PH (m) | PD (cm) | # mites |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Cerro Yglesias | 45.9 (± 3.0) | 1.6 (± 0.4) | 21.5 (± 1.4) | 40.4 (± 3.5) | 7.0 (± 0.5) | 5.4 (± 0.6) | 12.4 (± 1.2) | 17.7 (± 1.2) | 0.8 (± 0.1) | 11.2 (± 9.9) | 7.7 (± 8.9) |
| Playa by Station | 48 (± 2.4) | 2.4 (± 0.4) | 22.9 (± 1.8) | 40.1 (± 2.8) | 7.6 (± 0.4) | 5.7 (± 0.4) | 13.3 (± 0.8) | 18.4 (± 1.2) | 1.8 (± 0.9) | 8.3 (± 12.1) | N/A |
| Chatham Trail | 48.9 (± 3.3) | 2.1 (± 0.3) | 22.8 (± 1.3) | 40.8 (± 1.6) | 7.5 (± 0.4) | 5.6 (± 0.3) | 13.2 (± 0.5) | 18.2 (± 1.3) | 1.7 (± 1.5) | 8.2 (± 6.8) | 10.4 (± 15.8) |
| Playa + Trail | 48.3 (± 2.8) | 2.3 (± 0.4) | 22.8 (± 1.6) | 40.3 (± 2.4) | 7.5 (± 0.4) | 5.7 (± 0.4) | 13.3 (± 0.7) | 18.3 (± 1.2) | 1.8 (± 1.2) | 8.3 (± 9.9) | 10.4 (± 15.8) |
| Manuelita Islet | 55.2 (± 3.2) | 3.5 (± 0.7) | 26.4 (± 2.3) | 45.9 (± 3.9) | 8.3 (± 0.6) | 6.3 (± 0.4) | 14.5 (± 1.0) | 19.5 (± 0.9) | 0.4 (± 0.1) | 6.4 (± 6.6) | 3.4 (± 6.5) |
| Population | SVL | mass | Fore | Hind | HW | HH | HL | lamellae | PH | PD | mites |
| Playa v Trail | 0.32 | 0.004 | 0.10 | 0.002 | 0.23 | 0.61 | 0.05 | 0.69 | 0.34 | 0.17 | N/A |
| Playa v Cerro | 0.003 | 0.0003 | 0.002 | 0.30 | 0.001 | 0.06 | 0.001 | 0.21 | 0.03 | 0.69 | N/A |
| Cerro v Trail | 0.003 | 0.00001 | 0.02 | 0.31 | 0.01 | 0.19 | 0.04 | 0.15 | 0.0002 | 0.14 | 0.61 |
| Playa v Islet | < 0.00001 | < 0.00001 | < 0.00001 | 0.00001 | 0.0002 | < 0.00001 | 0.0003 | 0.0006 | 0.0001 | 0.20 | N/A |
| Trail v Islet | < 0.00001 | < 0.00001 | < 0.00001 | < 0.00001 | 0.00001 | < 0.00001 | 0.00002 | 0.004 | < 0.00001 | 0.77 | 0.09 |
| Cerro v Islet | < 0.00001 | < 0.00001 | < 0.00001 | 0.001 | 0.00001 | 0.00005 | 0.00003 | 0.0001 | < 0.00001 | 0.12 | 0.15 |
| Population | SVL | mass | Fore | Hind | HW | HH | HL | lamellae | PH | PD | mites |
| Main v Cerro | 0.02 | 0.00001 | 0.02 | 0.97 | 0.0004 | 0.07 | 0.001 | 0.14 | 0.01 | 0.36 | 0.60 |
| Main v Islet | < 0.00001 | < 0.00001 | < 0.00001 | < 0.00001 | < 0.00001 | < 0.00001 | < 0.00001 | 0.0004 | < 0.00001 | 0.46 | 0.08 |
| Cerro v Islet | < 0.00001 | < 0.00001 | < 0.00001 | 0.001 | 0.00001 | 0.00005 | 0.00003 | 0.0001 | < 0.00001 | 0.12 | 0.15 |
Table 5.
Mean values and pairwise comparisons of habitat use in Isla Gorgona anoles. Bold values are statistically significant. * Denotes that only one individual was observed and is not believed to reflect typical habitat use.
Table 5.
Mean values and pairwise comparisons of habitat use in Isla Gorgona anoles. Bold values are statistically significant. * Denotes that only one individual was observed and is not believed to reflect typical habitat use.
| Species | N | Perch Height (m) | Perch Diameter (cm) |
|---|---|---|---|
| Dactyloa chocorum | 1 * | 2.2 | 12.0 |
| Dactyloa gorgonae | 8 | 15.6 (± 7.3) | 18.2 (± 7.1) |
| Dactyloa princeps | 21 | 3.1 (± 1.5) | 14.5 (± 7.9) |
| Norops medemi | 30 | 3.4 (± 2.6) | 7.9 (± 5.3) |
| Norops parvauritus | 1 * | 4.4 | 93.0 |
| D. gorgonae vs. D. princeps | < 0.00001 | 0.26 | |
| D. gorgonae vs. N. medemi | < 0.00001 | 0.10 | |
| D. princeps vs. N. medemi | 0.66 | 0.04 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Phillips, J.G.; Burton, S.E.; Womack, M.M.; Pulver, E.; Nicholson, K.E. Biogeography, Systematics, and Ecomorphology of Pacific Island Anoles. Diversity 2019, 11, 141. https://doi.org/10.3390/d11090141
AMA Style
Phillips JG, Burton SE, Womack MM, Pulver E, Nicholson KE. Biogeography, Systematics, and Ecomorphology of Pacific Island Anoles. Diversity. 2019; 11(9):141. https://doi.org/10.3390/d11090141
Chicago/Turabian StylePhillips, John G., Sarah E. Burton, Margarita M. Womack, Evan Pulver, and Kirsten E. Nicholson. 2019. "Biogeography, Systematics, and Ecomorphology of Pacific Island Anoles" Diversity 11, no. 9: 141. https://doi.org/10.3390/d11090141
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.