tRNA-Derived RNA Fragments Are Novel Biomarkers for Diagnosis, Prognosis, and Tumor Subtypes in Prostate Cancer

 and
and
Abstract
:1. Introduction
2. Methods
2.1. Data Collection
2.2. Identification and Quantification of 5′-tRFs in PRAD
2.3. Quantification of mRNA Expression Levels
2.4. Identification of 5′-tRFs Dysregulated in PRAD
2.5. Inference of Potential Functions of 5′-tRFs
2.6. Construction of Prognostic Predictor of 5′-tRFs
2.7. Identification of Tumor Subtypes Based on tRF Expression
2.8. Mutational Data Analysis
2.9. Statistical Analysis
3. Results
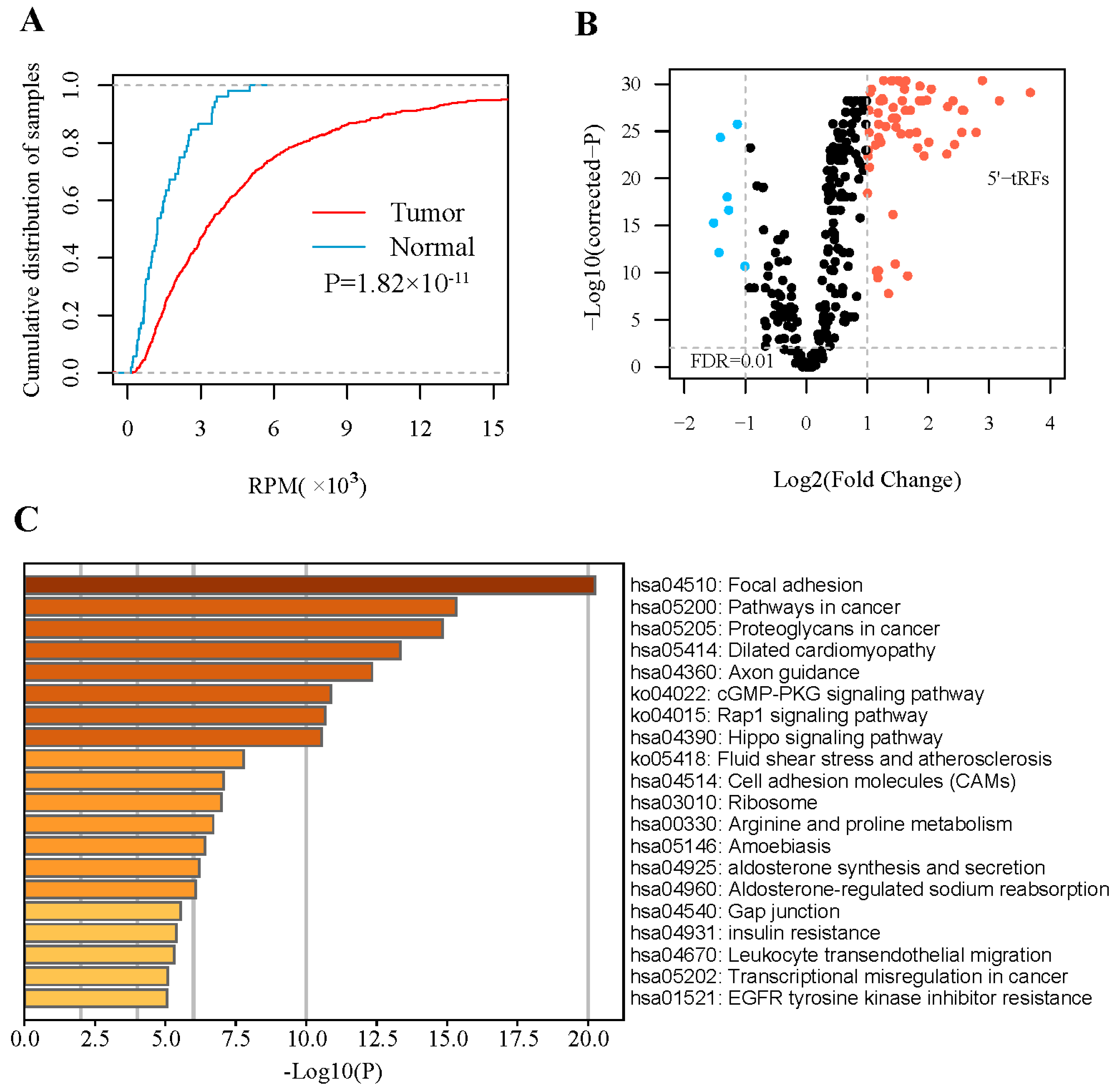
3.1. 5′- tRFs Are Dysregulated in PRAD
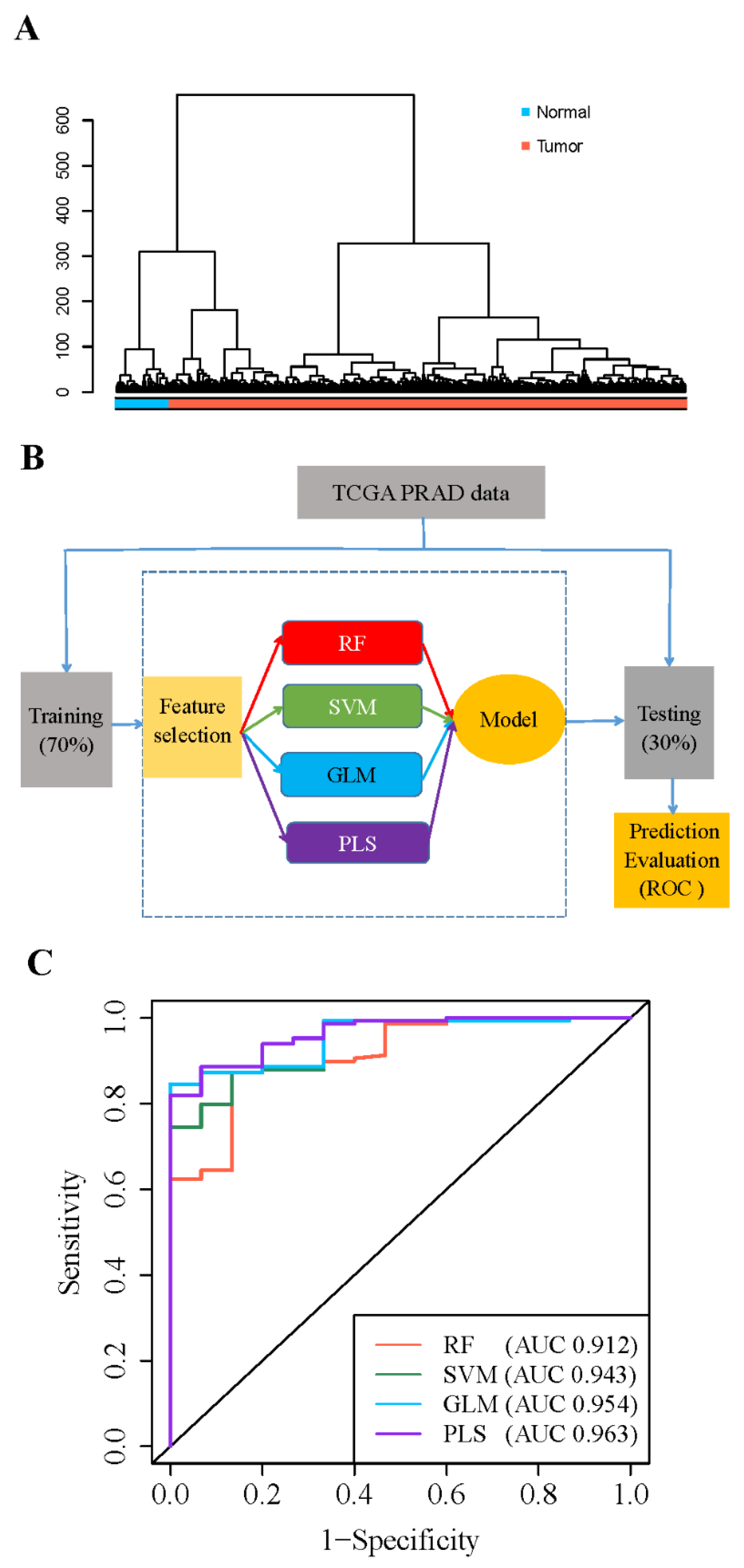
3.2. 5′- tRFs Are Novel Biomarkers for Diagnosis of PRAD
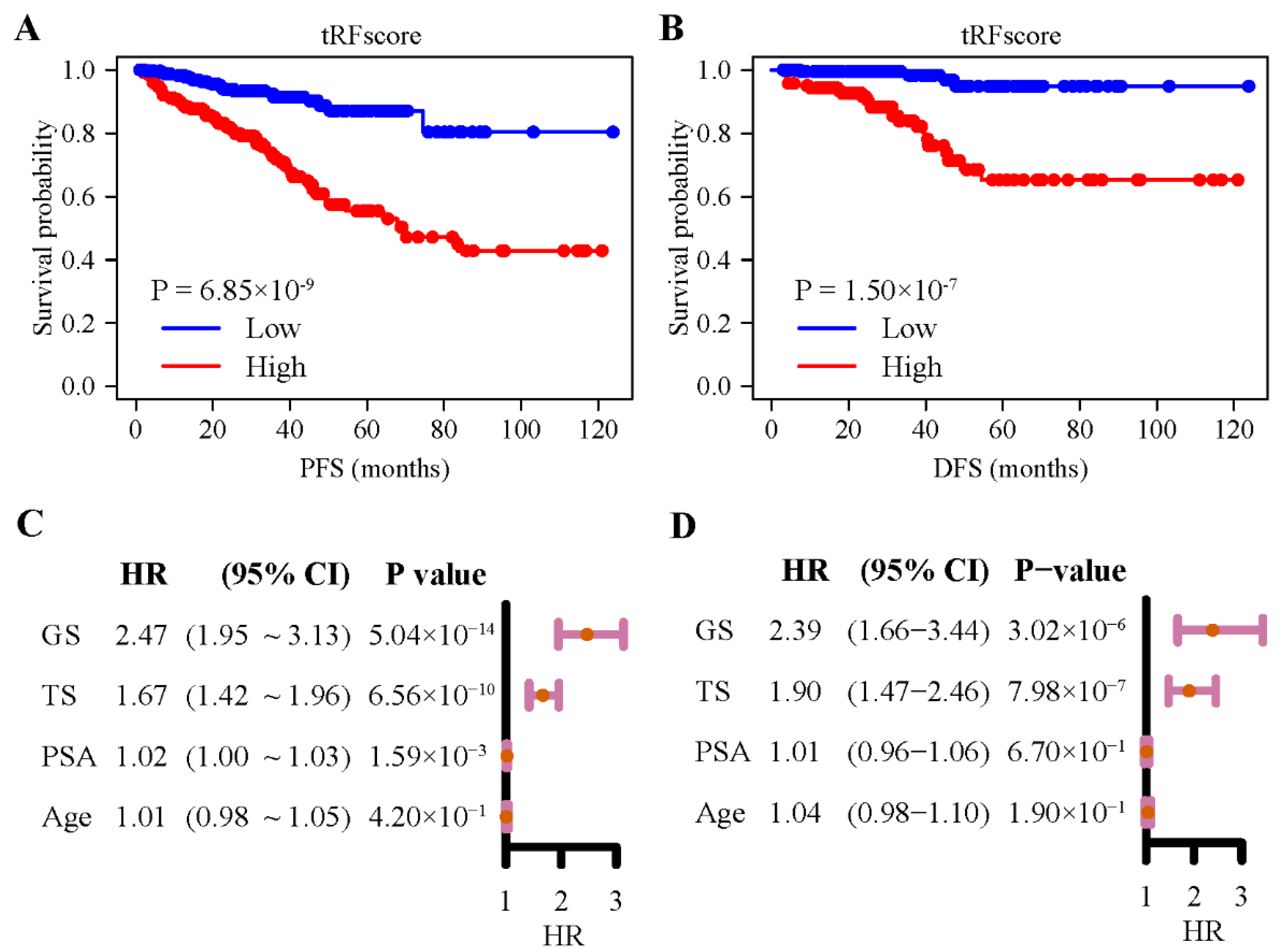
3.3. 5′- tRFs Are Novel Biomarkers for Prognosis of PRAD
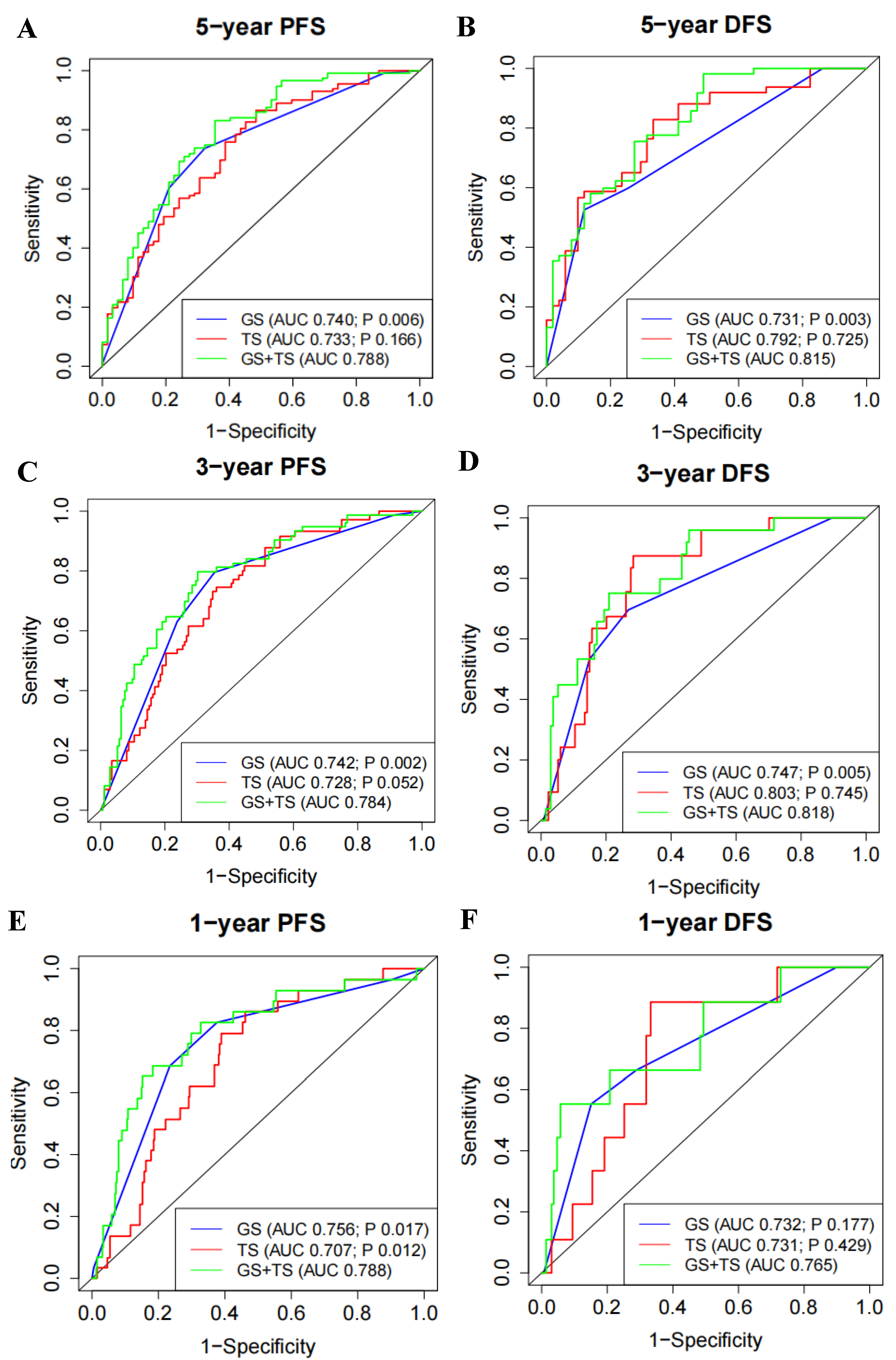
3.4. 5′- tRFs Provide Independent Prognostic Information for PRAD
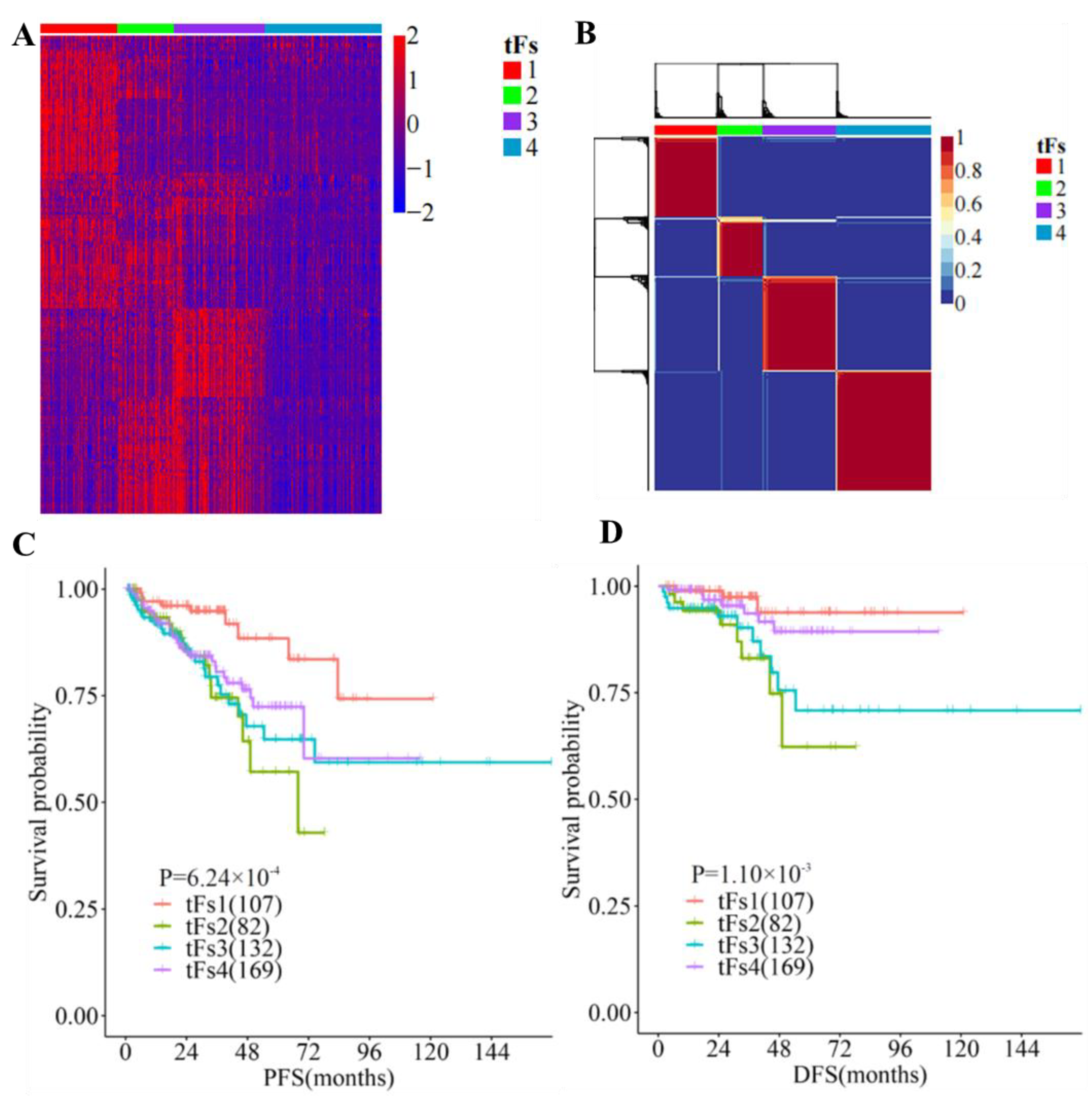
3.5. 5′- tRFs Are Novel Biomarkers for the Tumor Classification of PRAD
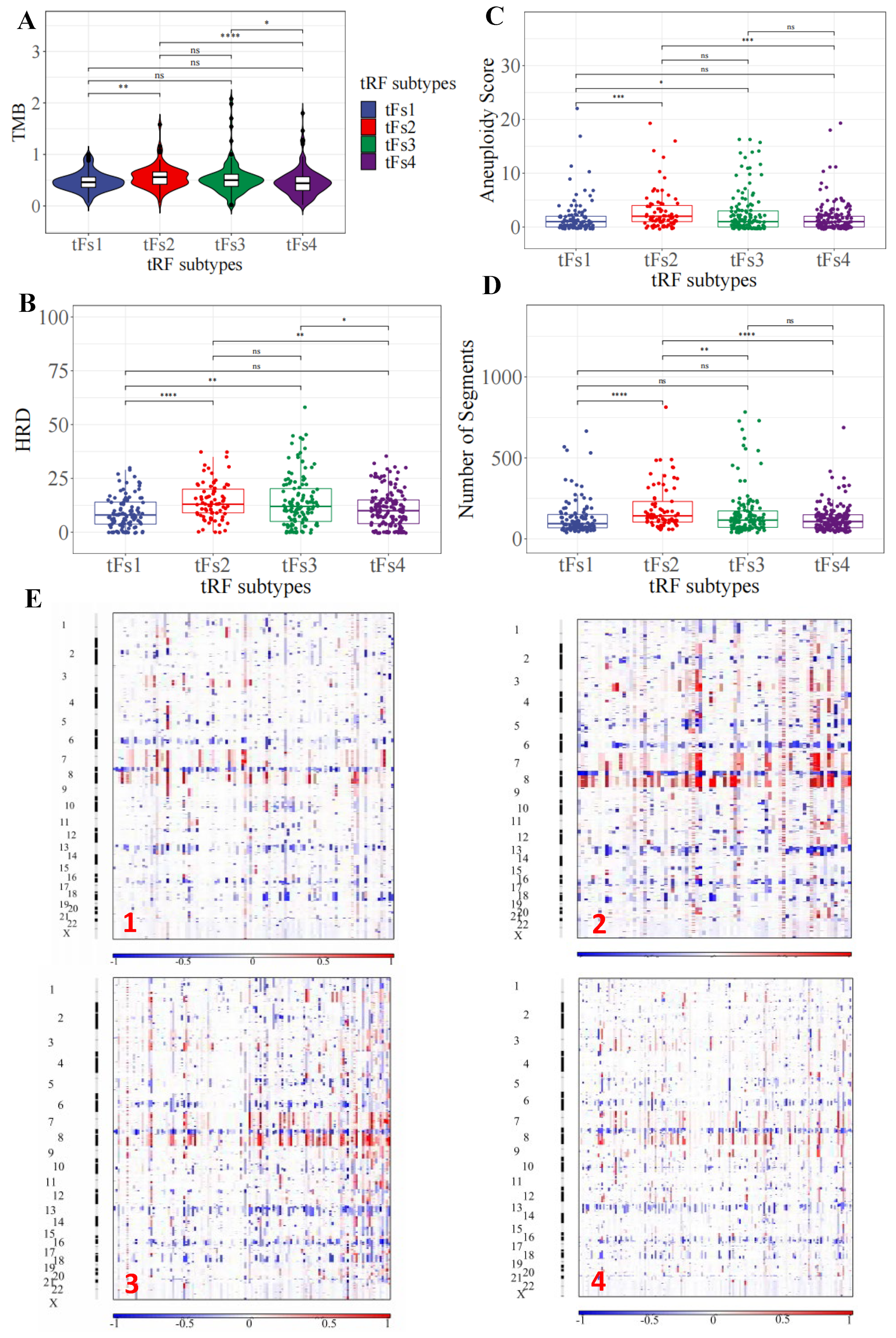
3.6. Genomic Landscapes of 5′-tRFs Tumor Subtypes of PRAD
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| tRFs | tRNA-derived RNA fragments |
| PRAD | prostate adenocarcinoma |
| PSA | serum prostate-specific antigen |
| ncRNAs | small noncoding RNAs |
| ANG | angiogenin |
| TCGA | The Cancer Genome Atlas |
| ICGC | International Cancer Genome Consortium |
| BWA | burrows-wheeler transform |
| RPM | reads per million |
| RPKM | read per kilobase per million |
| FDR | false discovery rate |
| RF | random forest |
| SVM | support vector machine |
| GLM | generalized linear model |
| PLS | partial least squares |
| ROC | receiver operation characteristic |
| GBA | guilt by association |
| GO | gene ontology |
| KEGG | Kyoto encyclopedia of genes and genomes |
| PFS | progression-free survival |
| HR | hazard ratio |
| CI | confidence interval |
| LASSOl | east absolute shrinkage and selection operator |
| NMF | non-negative matrix factorization |
| IQRs | interquartile ranges |
| SNP | single nucleotide polymorphism |
| INS | small insertions |
| DEL | deletions |
| TMB | tumor mutational burden |
| HRD | homologous recombination deficiency |
| AS | aneuploidy score |
| SD | standard deviation |
| TS | tRF score |
| GS | Gleason score |
| DFS | disease-free survival |
References
- Xia, C.; Dong, X.; Li, H.; Cao, M.; Sun, D.; He, S.; Yang, F.; Yan, X.; Zhang, S.; Li, N.; et al. Cancer statistics in China and United States, 2022: Profiles, trends, and determinants. Chin. Med. J. 2022, 135, 584–590. [Google Scholar] [CrossRef] [PubMed]
- De Angelis, R.; Sant, M.; Coleman, M.P.; Francisci, S.; Baili, P.; Pierannunzio, D.; Trama, A.; Visser, O.; Brenner, H.; Ardanaz, E.; et al. Cancer survival in Europe 1999–2007 by country and age: Results of EUROCARE-5-a population-based study. Lancet Oncol. 2014, 15, 23–34. [Google Scholar] [CrossRef]
- Virgo, K.S.; Basch, E.; Loblaw, D.A.; Oliver, T.K.; Rumble, R.B.; Carducci, M.A.; Nordquist, L.; Taplin, M.-E.; Winquist, E.; Singer, E.A. Second-Line Hormonal Therapy for Men with Chemotherapy-Naïve, Castration-Resistant Prostate Cancer: American Society of Clinical Oncology Provisional Clinical Opinion. J. Clin. Oncol. 2017, 35, 1952–1964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yadav, S.S.; Li, J.; Lavery, H.J.; Yadav, K.K.; Tewari, A.K. Next-generation sequencing technology in prostate cancer diagnosis, prognosis, and personalized treatment. Urol. Oncol. 2015, 33, 267.e1–267.e13. [Google Scholar] [CrossRef] [PubMed]
- Fujita, K.; Nonomura, N. Urinary biomarkers of prostate cancer. Int. J. Urol. 2018, 25, 770–779. [Google Scholar] [CrossRef] [Green Version]
- Di Minno, A.; Aveta, A.; Gelzo, M.; Tripodi, L.; Pandolfo, S.D.; Crocetto, F.; Imbimbo, C.; Castaldo, G. 8-Hydroxy-2-Deoxyguanosine and 8-Iso-Prostaglandin F2alpha: Putative Biomarkers to assess Oxidative Stress Damage Following Robot-Assisted Radical Prostatectomy (RARP). J. Clin. Med. 2022, 11, 6102. [Google Scholar] [CrossRef]
- Kristiansen, G. Diagnostic and prognostic molecular biomarkers for prostate cancer. Histopathology 2012, 60, 125–141. [Google Scholar] [CrossRef]
- Lee, Y.S.; Shibata, Y.; Malhotra, A.; Dutta, A. A novel class of small RNAs: tRNA-derived RNA fragments (tRFs). Genes Dev. 2009, 23, 2639–2649. [Google Scholar] [CrossRef] [Green Version]
- Yu, M.; Lu, B.; Zhang, J.; Ding, J.; Liu, P.; Lu, Y. tRNA-derived RNA fragments in cancer: Current status and future perspectives. J. Hematol. Oncol. 2020, 13, 121. [Google Scholar] [CrossRef]
- Phizicky, E.M.; Hopper, A.K. tRNA biology charges to the front. Genes Dev. 2010, 24, 1832–1860. [Google Scholar] [CrossRef]
- Sun, C.; Fu, Z.; Wang, S.; Li, J.; Li, Y.; Zhang, Y.; Yang, F.; Chu, J.; Wu, H.; Huang, X.; et al. Roles of tRNA-derived fragments in human cancers. Cancer Lett. 2018, 414, 16–25. [Google Scholar] [CrossRef]
- Yamasaki, S.; Ivanov, P.; Hu, G.-F.; Anderson, P. Angiogenin cleaves tRNA and promotes stress-induced translational repression. J. Cell Biol. 2009, 185, 35–42. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Yang, J.; Yu, M.; Yao, D.; Zhou, L.; Li, X.; Qiu, Q.; Lin, W.; Lu, B.; Chen, E.; et al. Global identification and characterization of tRNA-derived RNA fragment landscapes across human cancers. NAR Cancer 2020, 2, zcaa031. [Google Scholar] [CrossRef]
- Maute, R.L.; Schneider, C.; Sumazin, P.; Holmes, A.; Califano, A.; Basso, K.; Dalla-Favera, R. tRNA-derived microRNA modulates proliferation and the DNA damage response and is down-regulated in B cell lymphoma. Proc. Natl. Acad. Sci. USA 2013, 110, 1404–1409. [Google Scholar] [CrossRef] [Green Version]
- Goodarzi, H.; Liu, X.H.; Nguyen, H.C.B.; Zhang, S.; Fish, L.; Tavazoie, S.F. Endogenous tRNA-Derived Fragments Suppress Breast Cancer Progression via YBX1 Displacement. Cell 2015, 161, 790–802. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.K.; Fuchs, G.; Wang, S.; Wei, W.; Zhang, Y.; Park, H.; Roy-Chaudhuri, B.; Li, P.; Xu, J.; Chu, K.; et al. A transfer-RNA-derived small RNA regulates ribosome biogenesis. Nature 2017, 552, 57–62. [Google Scholar] [CrossRef]
- Chu, X.; He, C.; Sang, B.; Yang, C.; Yin, C.; Ji, M.; Qian, A.; Tian, Y. Transfer RNAs-derived small RNAs and their application potential in multiple diseases. Front. Cell Dev. Biol. 2022, 10, 954431. [Google Scholar] [CrossRef]
- Honda, S.; Loher, P.; Shigematsu, M.; Palazzo, J.P.; Suzuki, R.; Imoto, I.; Rigoutsos, I.; Kirino, Y. Sex hormone-dependent tRNA halves enhance cell proliferation in breast and prostate cancers. Proc. Natl. Acad. Sci. USA 2015, 112, E3816–E3825. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.; Lee, M.; Song, G.; Lim, W. tRNA(Lys)-Derived Fragment Alleviates Cisplatin-Induced Apoptosis in Prostate Cancer Cells. Pharmaceutics 2021, 13, 55. [Google Scholar] [CrossRef]
- Balatti, V.; Nigita, G.; Veneziano, D.; Drusco, A.; Stein, G.S.; Messier, T.L.; Farina, N.H.; Lian, J.B.; Tomasello, L.; Liu, C.-G.; et al. tsRNA signatures in cancer. Proc. Natl. Acad. Sci. USA 2017, 114, 8071–8076. [Google Scholar] [CrossRef]
- Pekarsky, Y.; Balatti, V.; Palamarchuk, A.; Rizzotto, L.; Veneziano, D.; Nigita, G.; Rassenti, L.Z.; Pass, H.I.; Kipps, T.J.; Liu, C.-G.; et al. Dysregulation of a family of short noncoding RNAs, tsRNAs, in human cancer. Proc. Natl. Acad. Sci. USA 2016, 113, 5071–5076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, D.; Sun, X.; Zhou, L.; Amanullah, M.; Pan, X.; Liu, Y.; Liang, M.; Liu, P.; Lu, Y. OncotRF: An online resource for exploration of tRNA-derived fragments in human cancers. RNA Biol. 2020, 17, 1081–1091. [Google Scholar] [CrossRef] [PubMed]
- Van Dam, S.; Vosa, U.; van der Graaf, A.; Franke, L.; de Magalhaes, J.P. Faculty Opinions recommendation of Gene co-expression analysis for functional classification and gene-disease predictions. Brief Bioinform. 2018, 19, 575–592. [Google Scholar] [CrossRef] [PubMed]
- Knijnenburg, T.A.; Wang, L.; Zimmermann, M.T.; Chambwe, N.; Gao, G.F.; Cherniack, A.D.; Fan, H.; Shen, H.; Way, G.P.; Greene, C.S.; et al. Genomic and Molecular Landscape of DNA Damage Repair Deficiency across The Cancer Genome Atlas. Cell Rep. 2018, 23, 239–254.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, A.M.; Shih, J.; Ha, G.; Gao, G.F.; Zhang, X.; Berger, A.C.; Schumacher, S.E.; Wang, C.; Hu, H.; Liu, J.; et al. Genomic and Functional Approaches to Understanding Cancer Aneuploidy. Cancer Cell 2018, 33, 676–689.e3. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Heyer, W.-D. Homologous recombination in DNA repair and DNA damage tolerance. Cell Res. 2008, 18, 99–113. [Google Scholar] [CrossRef] [Green Version]
- Ok Atilgan, A.; Ozdemir, B.H.; Yilmaz Akcay, E.; Tepeoglu, M.; Borcek, P.; Dirim, A. Association between focal adhesion kinase and matrix metalloproteinase-9 expression in prostate adenocarcinoma and their influence on the progression of prostatic adenocarcinoma. Ann. Diagn. Pathol. 2020, 45, 151480. [Google Scholar] [CrossRef]
- Marx, A.; Schumann, A.; Höflmayer, D.; Bady, E.; Hube-Magg, C.; Möller, K.; Tsourlakis, M.C.; Steurer, S.; Büscheck, F.; Eichenauer, T.; et al. Up regulation of the Hippo signalling effector YAP1 is linked to early biochemical recurrence in prostate cancers. Sci. Rep. 2020, 10, 8916. [Google Scholar] [CrossRef]
- Munkley, J.; Mills, I.G.; Elliott, D.J. The role of glycans in the development and progression of prostate cancer. Nat. Rev. Urol. 2016, 13, 324–333. [Google Scholar] [CrossRef]
- Fay, E.K.; Graff, J.N. Immunotherapy in Prostate Cancer. Cancers 2020, 12, 1752. [Google Scholar] [CrossRef]
- Karnes, R.J.; Cheville, J.C.; Ida, C.M.; Sebo, T.J.; Nair, A.A.; Tang, H.; Munz, J.-M.; Kosari, F.; Vasmatzis, G. The Ability of Biomarkers to Predict Systemic Progression in Men with High-Risk Prostate Cancer Treated Surgically Is Dependent on ERG Status. Cancer Res. 2010, 70, 8994–9002. [Google Scholar] [CrossRef] [Green Version]
- Feng, F.Y.; Brenner, J.C.; Hussain, M.; Chinnaiyan, A.M. Molecular Pathways: Targeting ETS Gene Fusions in Cancer. Clin. Cancer Res. 2014, 20, 4442–4448. [Google Scholar] [CrossRef] [Green Version]
- Ray-Coquard, I.; Pautier, P.; Pignata, S.; Pérol, D.; González-Martín, A.; Berger, R.; Fujiwara, K.; Vergote, I.; Colombo, N.; Mäenpää, J.; et al. Olaparib plus Bevacizumab as First-Line Maintenance in Ovarian Cancer. New Engl. J. Med. 2019, 381, 2416–2428. [Google Scholar] [CrossRef]
- Zhao, E.Y.; Shen, Y.; Pleasance, E.; Kasaian, K.; Leelakumari, S.; Jones, M.; Bose, P.; Ch’Ng, C.; Reisle, C.; Eirew, P.; et al. Homologous Recombination Deficiency and Platinum-Based Therapy Outcomes in Advanced Breast Cancer. Clin. Cancer Res. 2017, 23, 7521–7530. [Google Scholar] [CrossRef] [Green Version]
- Sun, C.; Yang, F.; Zhang, Y.; Chu, J.; Wang, J.; Wang, Y.; Zhang, Y.; Li, J.; Li, Y.; Fan, R.; et al. tRNA-Derived Fragments as Novel Predictive Biomarkers for Trastuzumab-Resistant Breast Cancer. Cell. Physiol. Biochem. 2018, 49, 419–431. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| tF-1 (n = 107) | tF-2 (n = 82) | tF-3 (n = 132) | tF-4 (n = 169) | p Value | |
|---|---|---|---|---|---|
| Age | 60.60 ± 6.72 | 61.41 ± 6.37 | 61.60 ± 6.78 | 60.54 ± 7.01 | 0.744 |
| PSA | 9.22 ± 9.42 | 11.68 ± 8.83 | 12.74 ± 15.95 | 10.53 ± 11.8 | 0.020 |
| Gleason score | 2.07 × 10−6 | ||||
| 6 | 17 (15.89%) | 2 (2.44%) | 13 (9.85%) | 13 (7.69%) | |
| 7 | 69 (64.49%) | 36 (43.90%) | 51 (38.64%) | 87 (51.48%) | |
| 8 | 13 (12.15%) | 9 (10.98%) | 17 (12.88%) | 25 (14.79%) | |
| 9/10 | 8 (7.47%) | 35 (42.68%) | 51 (38.63%) | 44 (26.04%) | |
| Grading group | 3.93 × 10−7 | ||||
| 1 | 17 (15.89%) | 2 (2.44%) | 13 (9.85%) | 13 (7.69%) | |
| 2 | 47 (43.93%) | 15 (18.29%) | 31 (23.48%) | 51 (30.18%) | |
| 3 | 22 (20.56%) | 21 (25.61%) | 20 (15.15%) | 36 (21.30%) | |
| 4 | 13 (12.15%) | 9 (10.98%) | 17 (12.88%) | 25 (14.79%) | |
| 5 | 8 (7.47%) | 35 (42.68%) | 51 (38.64%) | 44 (26.04%) |
| tF-1 (n = 89) | tF-2 (n = 52) | tF-3 (n = 38) | tF-4 (n = 147) | p Value | |
|---|---|---|---|---|---|
| ERG (fusion) | 1.24 × 10−4 | ||||
| Yes | 34(38.20%) | 12(23.08%) | 16(42.10%) | 84(57.14%) | |
| No | 55(61.80%) | 40(76.92%) | 22(57.90%) | 63(42.86%) | |
| FOXA1 (mutation) | |||||
| Yes | 5(5.62%) | 5(9.62%) | 1(2.63%) | 2(1.36%) | 0.038 |
| No | 84(94.38%) | 47(90.38%) | 37(97.37%) | 145(98.64%) | |
| KMT2D (mutation) | 0.0018 | ||||
| Yes | 1(1.12%) | 7(13.46%) | 1(2.63%) | 2(1.36%) | |
| No | 88(98.88%) | 45(86.54%) | 37(97.37%) | 145(98.64%) | |
| ZMYM3 (mutation) | 0.063 | ||||
| Yes | 0(0.00%) | 3(5.77%) | 1(2.63%) | 2(1.36%) | |
| No | 89(100%) | 49(94.23%) | 37(97.37%) | 145(98.64%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, W.; Yu, M.; Cheng, S.; Zhou, X.; Li, J.; Lu, Y.; Liu, P.; Ding, S. tRNA-Derived RNA Fragments Are Novel Biomarkers for Diagnosis, Prognosis, and Tumor Subtypes in Prostate Cancer. Curr. Oncol. 2023, 30, 981-999. https://doi.org/10.3390/curroncol30010075
Liu W, Yu M, Cheng S, Zhou X, Li J, Lu Y, Liu P, Ding S. tRNA-Derived RNA Fragments Are Novel Biomarkers for Diagnosis, Prognosis, and Tumor Subtypes in Prostate Cancer. Current Oncology. 2023; 30(1):981-999. https://doi.org/10.3390/curroncol30010075
Chicago/Turabian StyleLiu, Weigang, Mengqian Yu, Sheng Cheng, Xiaoxu Zhou, Jia Li, Yan Lu, Pengyuan Liu, and Shiping Ding. 2023. "tRNA-Derived RNA Fragments Are Novel Biomarkers for Diagnosis, Prognosis, and Tumor Subtypes in Prostate Cancer" Current Oncology 30, no. 1: 981-999. https://doi.org/10.3390/curroncol30010075