Constraints to Vegetation Growth Reduced by Region-Specific Changes in Seasonal Climate

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. CMIP5
- The models had monthly data of near-surface air temperature (output variable name in the standard output is tas; the other variables are showed the same way hereafter), precipitation (pr), surface downwelling shortwave radiation (rsds), and Leaf Area Index (LAI) (lai) data for the specific years (1875–2005: historical; and 2006–2099: Representative Concentration Pathway (RCP) 8.5).
- The land sub-model had year-to-year changes in LAI.
2.2. GIMMS-LAI3G
2.3. CRU/CRUNCEP
2.4. NDP026
2.5. Limiting Factor Analysis
2.6. Estimation of Precipitation Equivalent Water
2.7. Mann–Kendall Test
3. Results
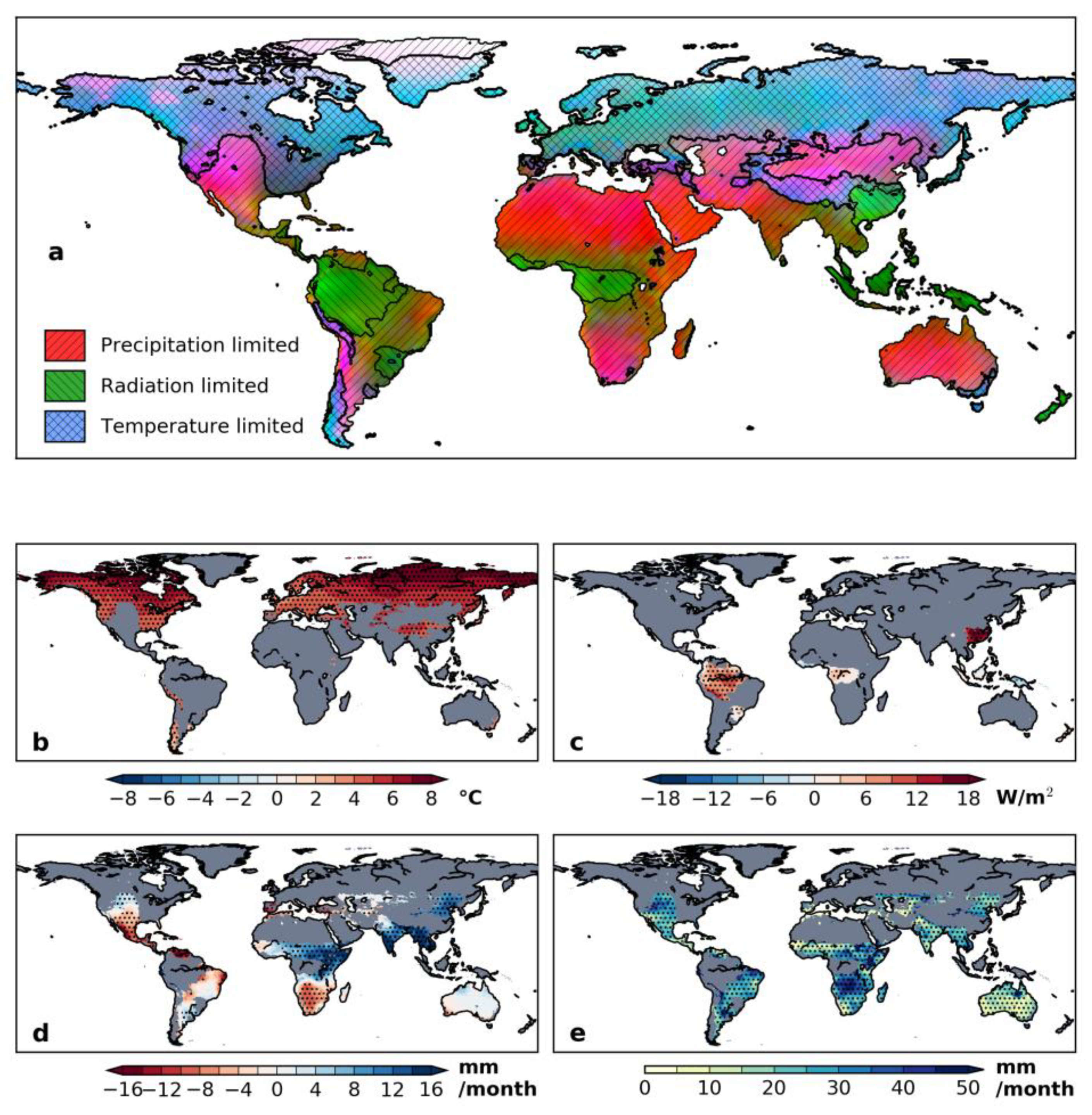
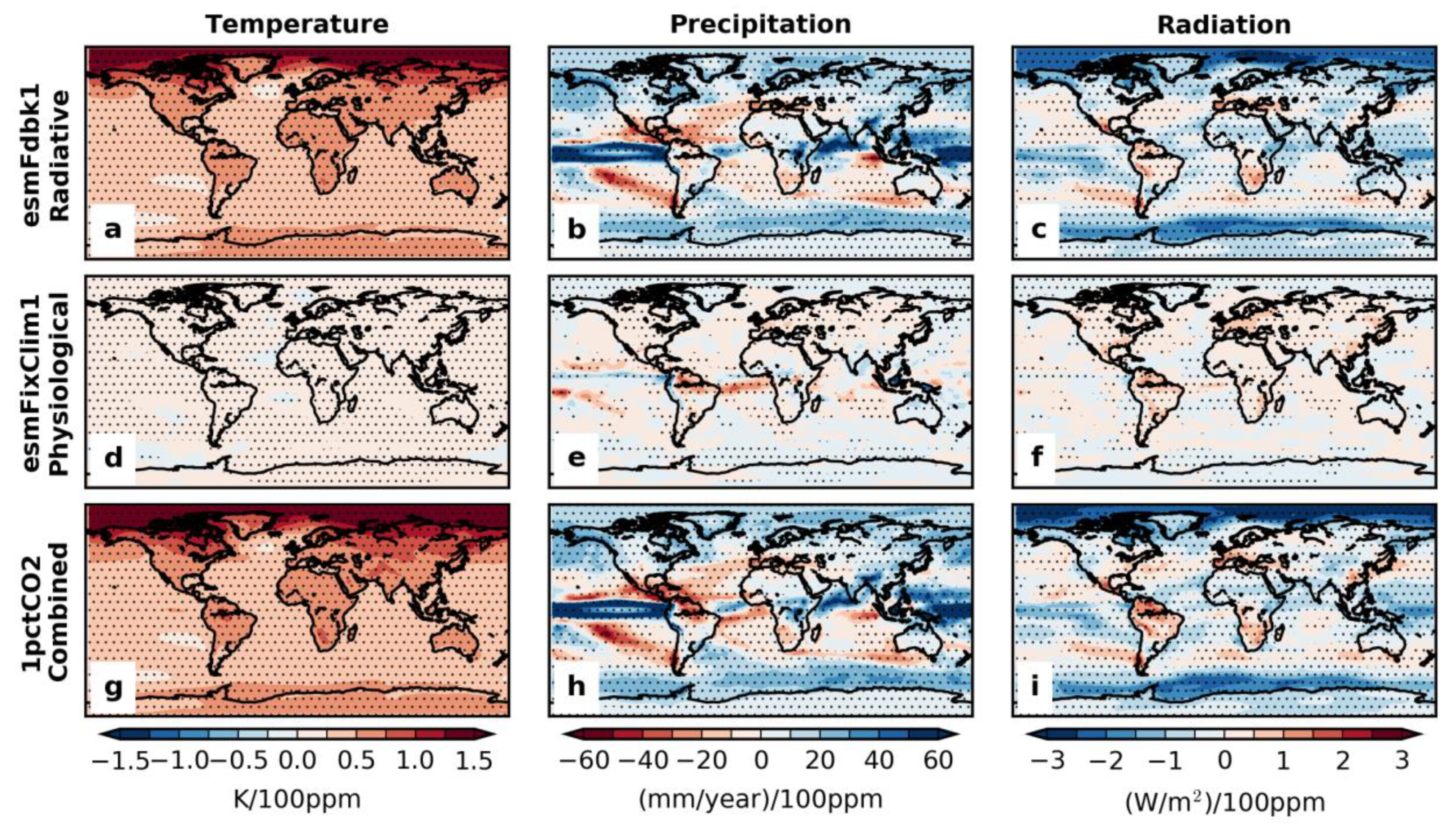
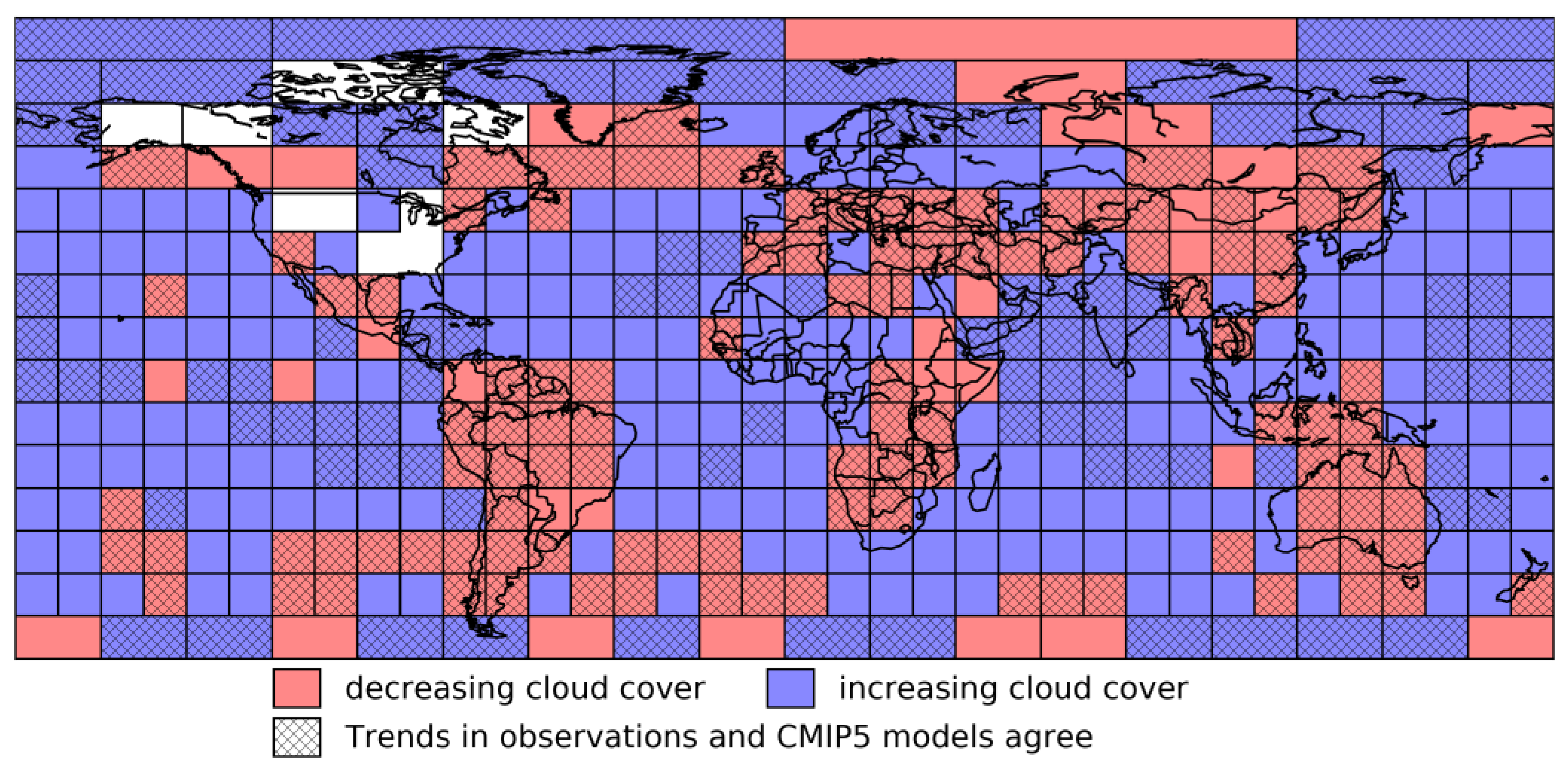
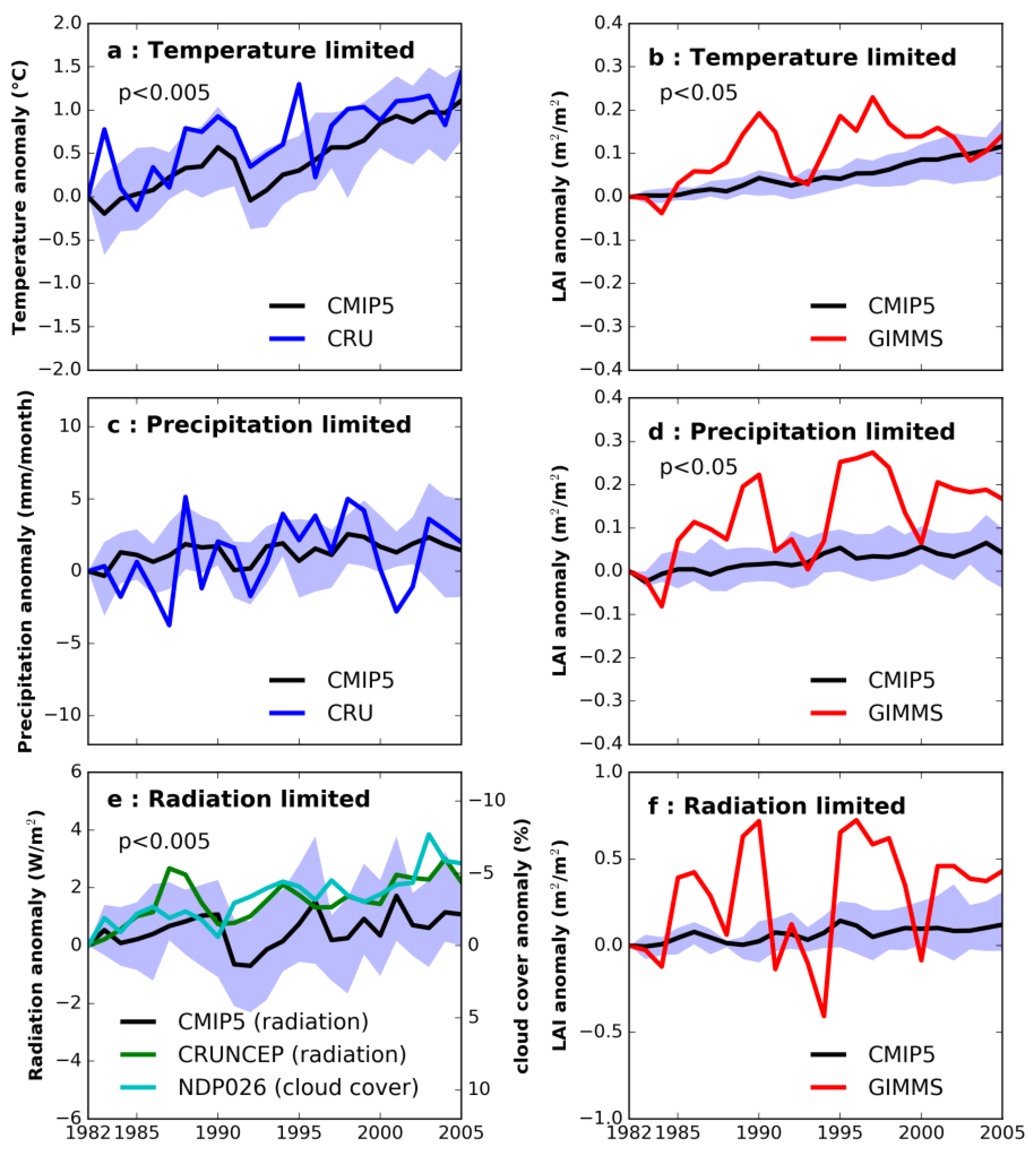
3.1. Annual Mean Trend and Climate Feedback to Vegetation
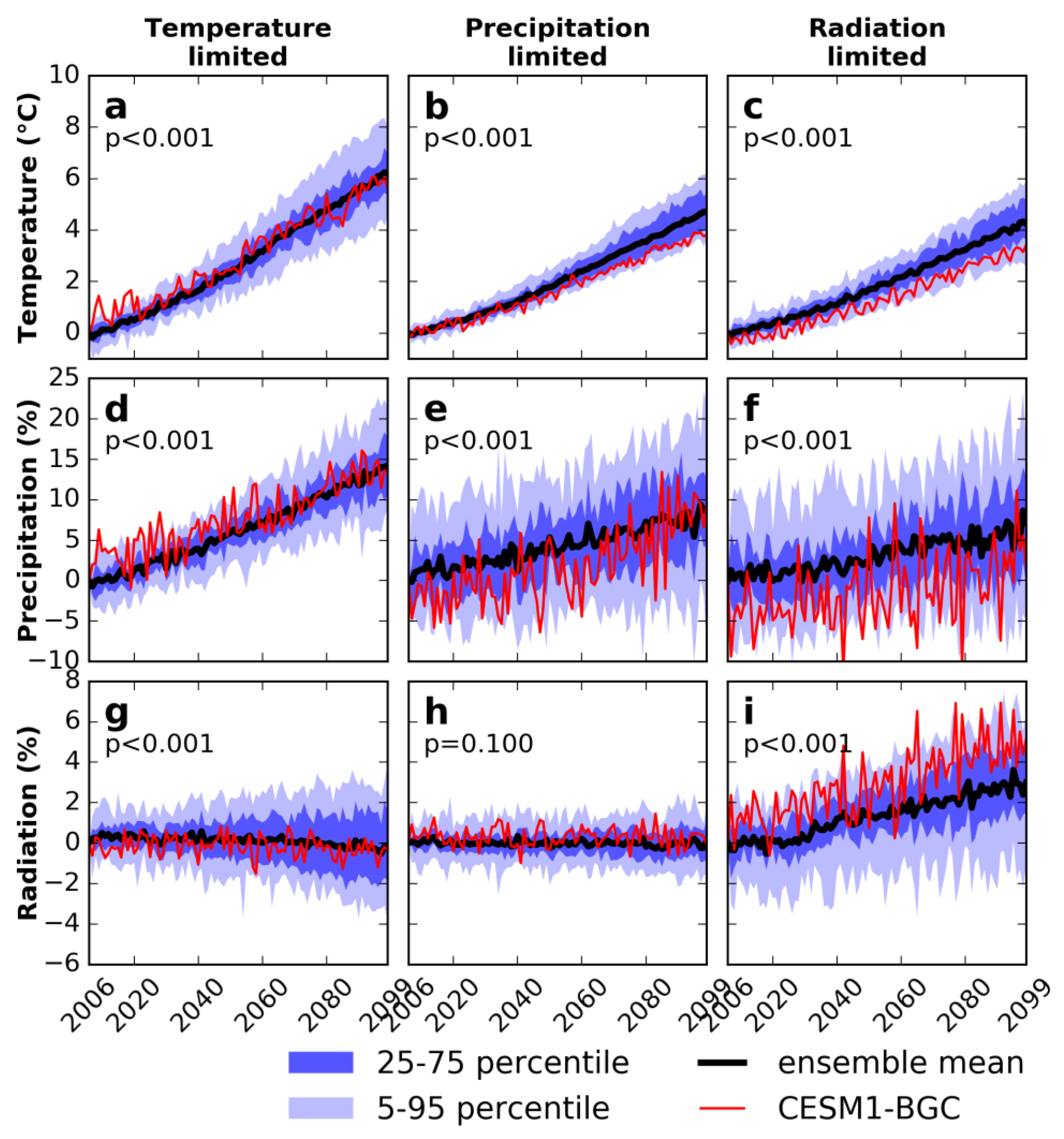
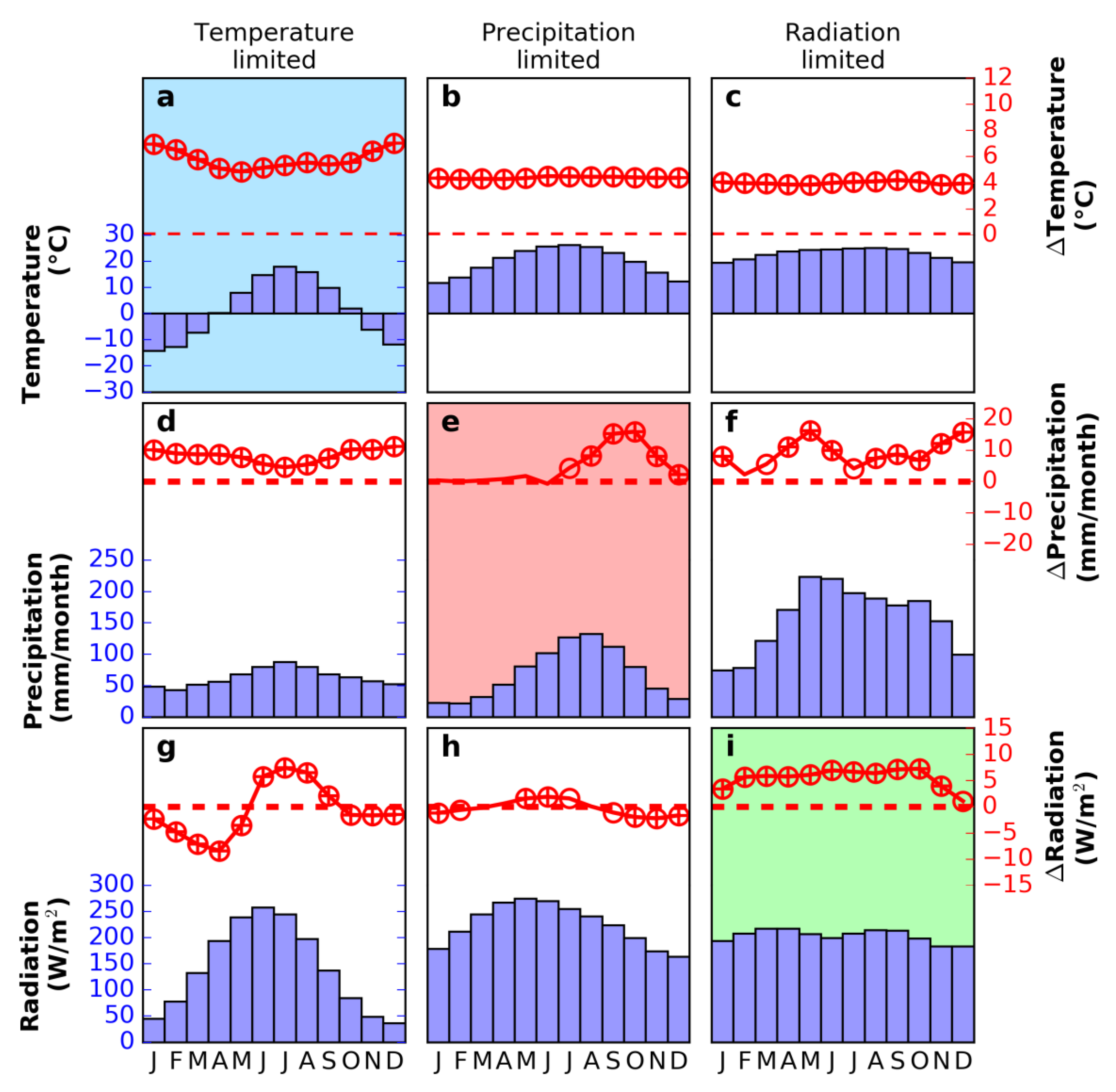
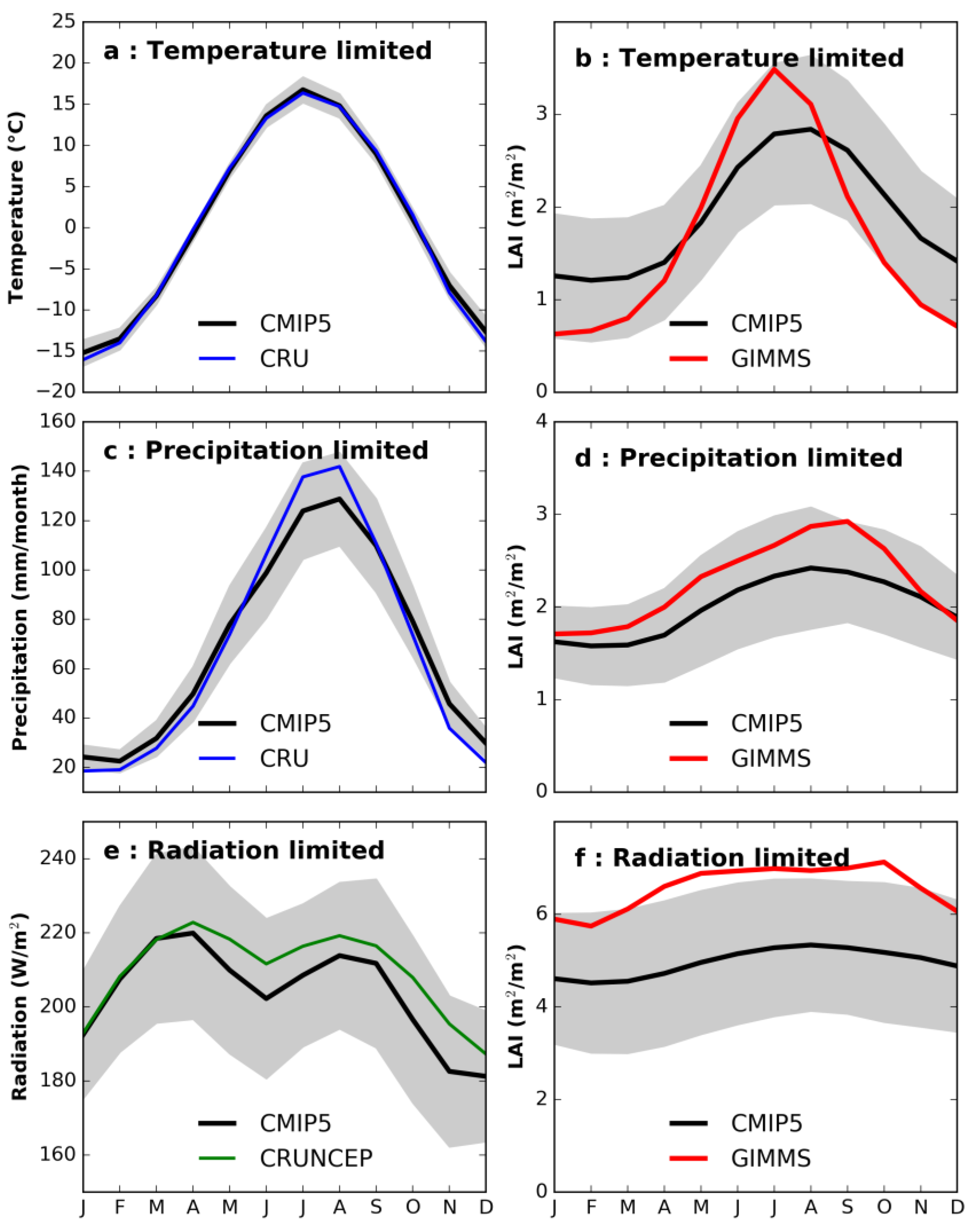
3.2. Seasonal Trend in Climate and Vegetation
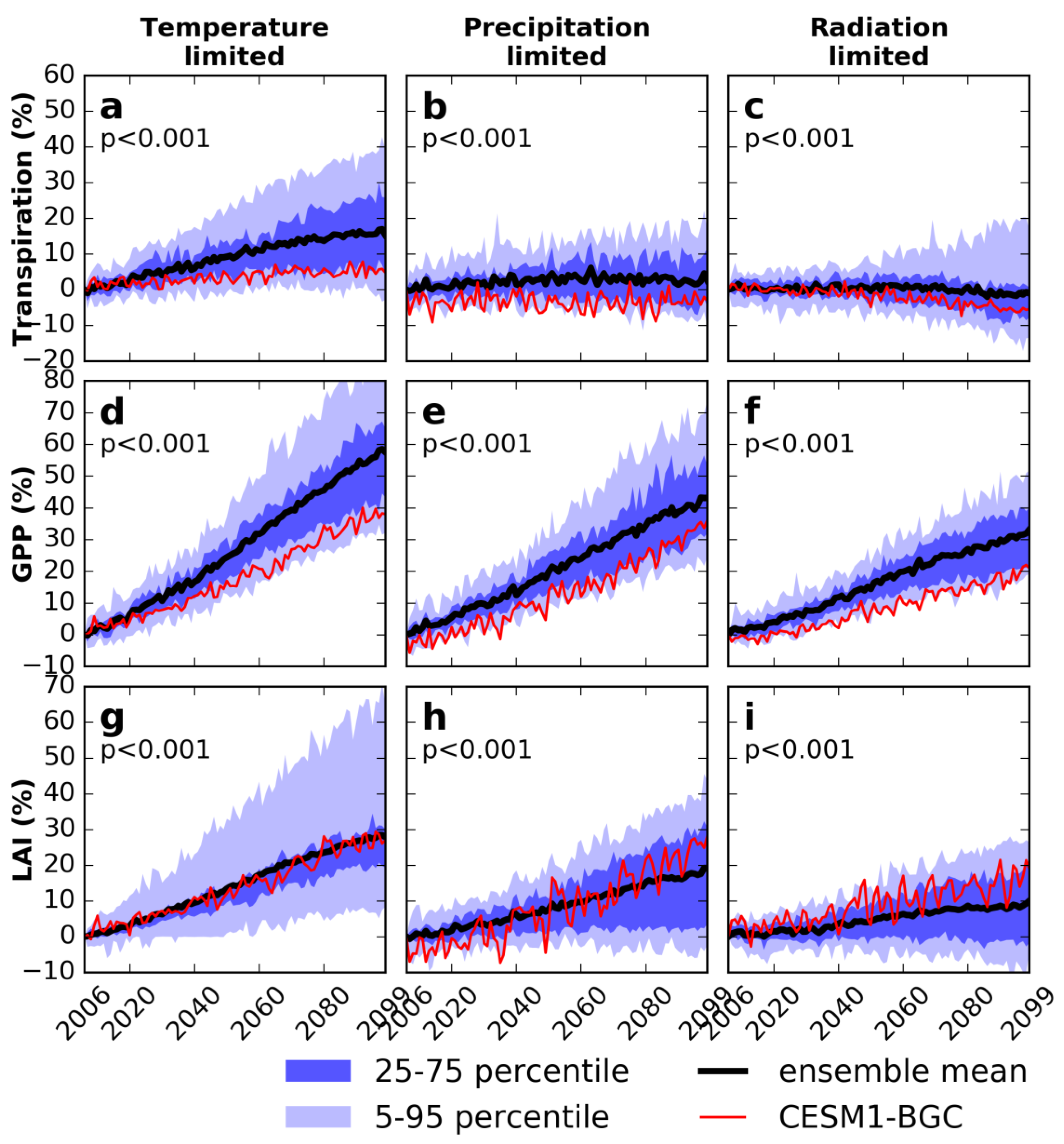
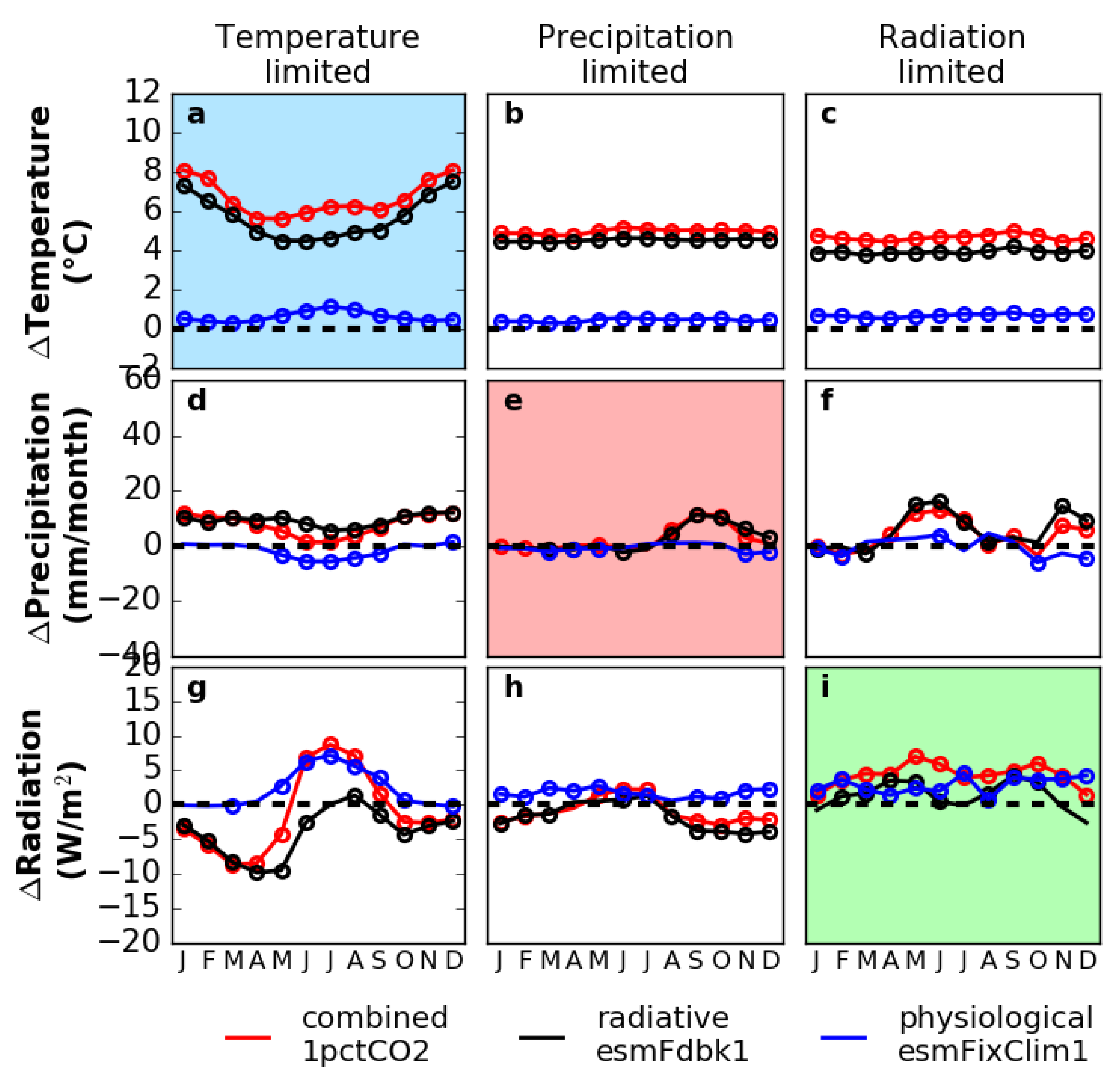
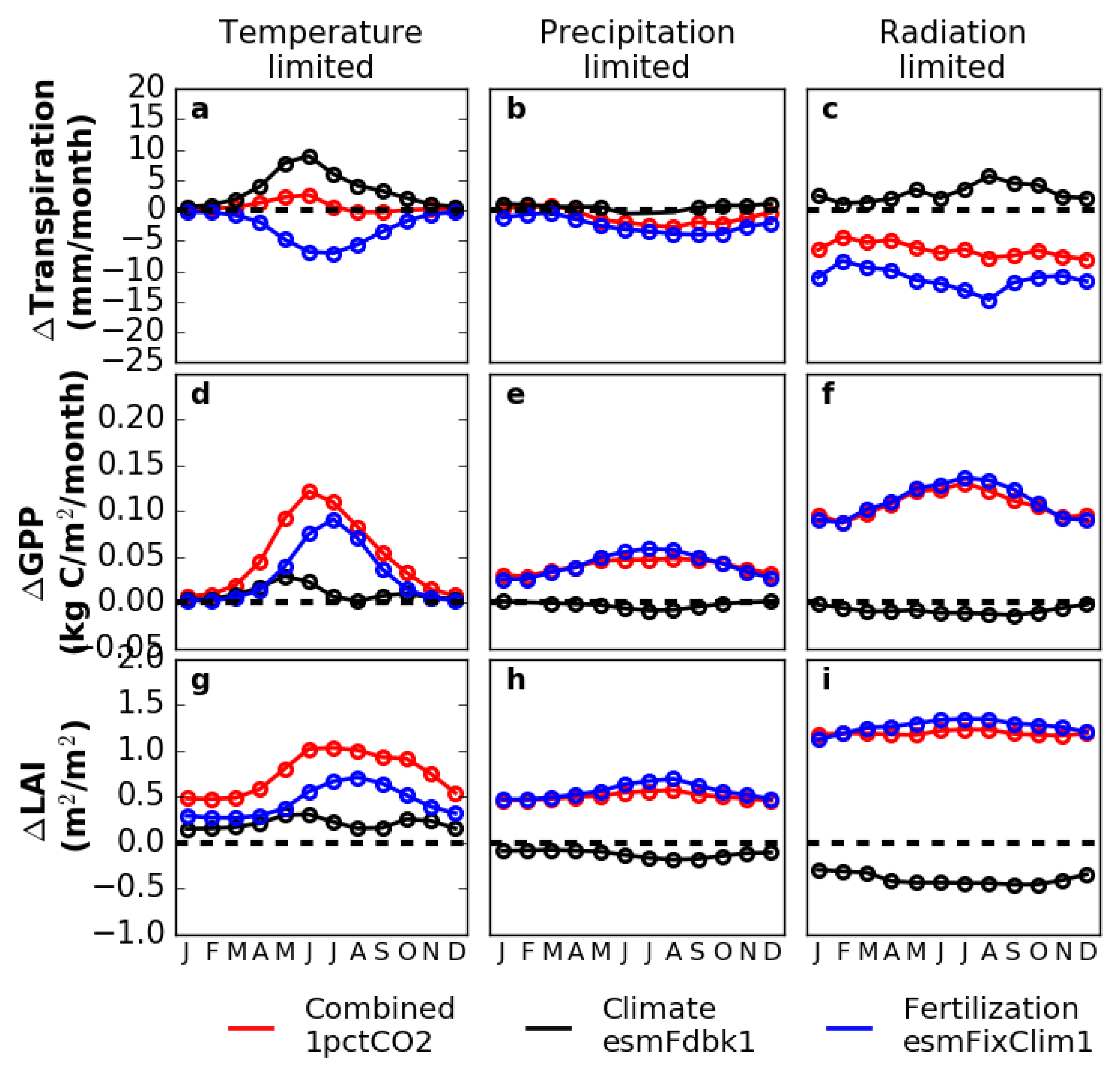
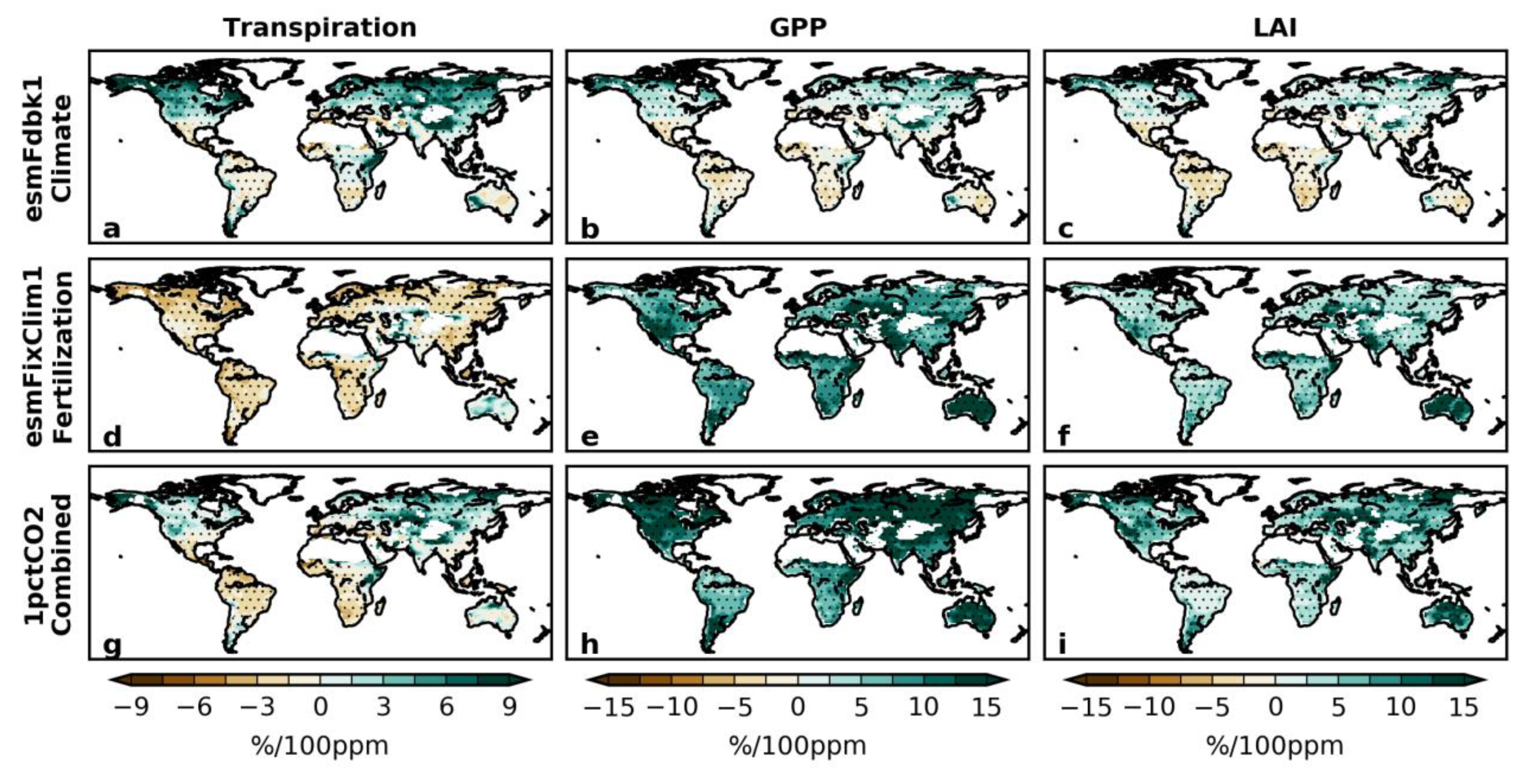
3.3. Sensitivity Experiments for Climate and Vegetation

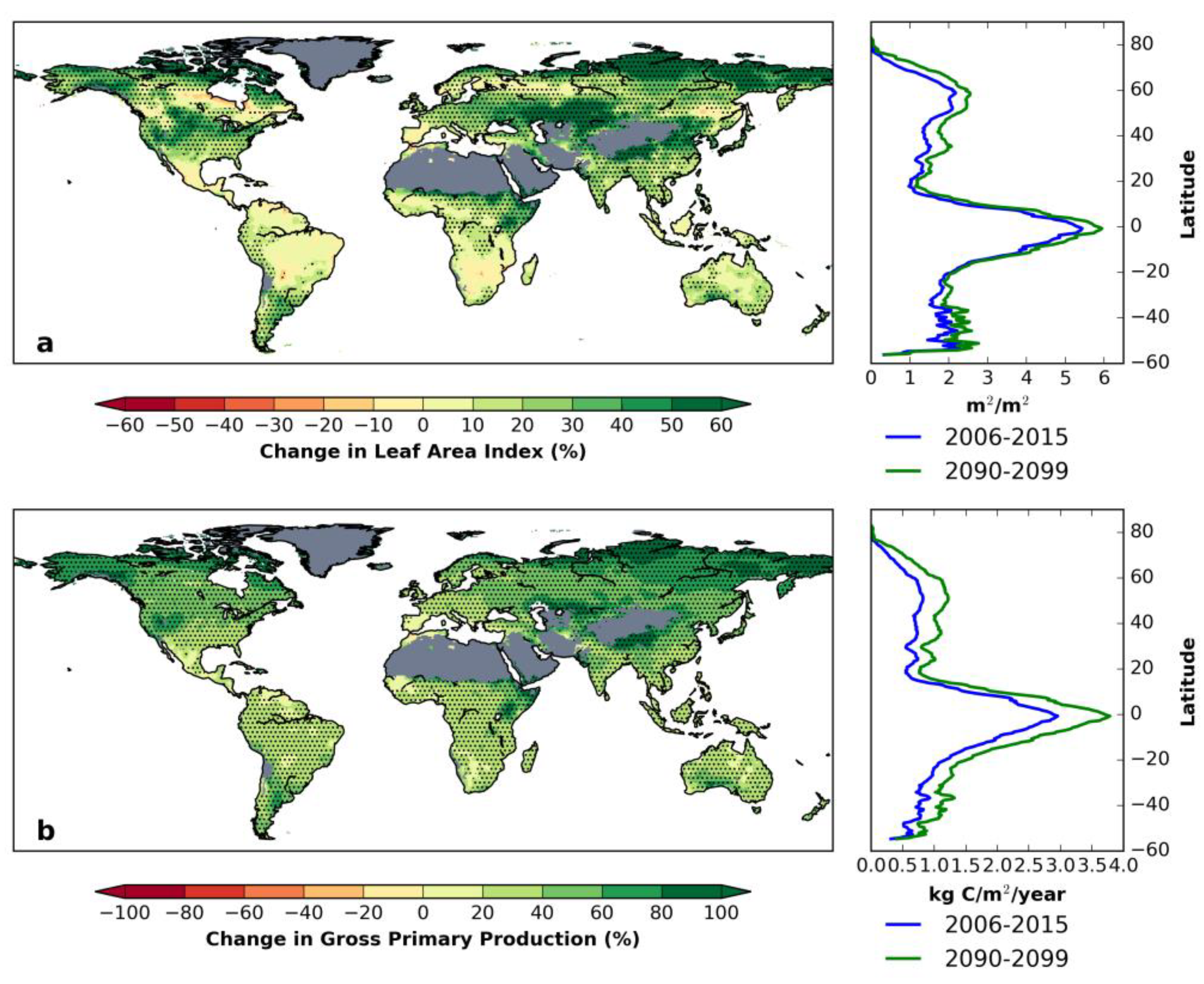
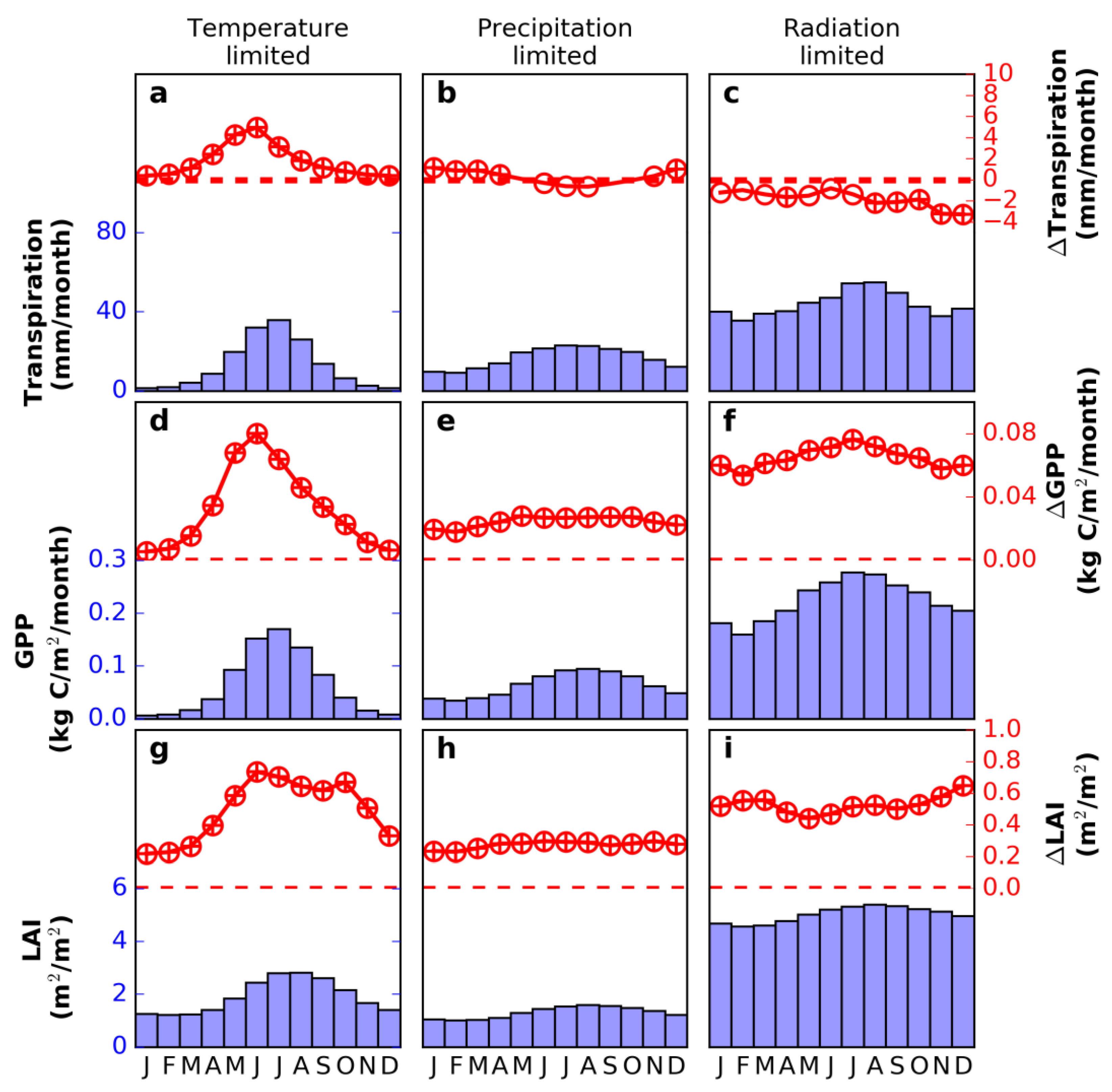
3.4. Decomposing Vegetation Growth Into Three Factors
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Friedlingstein, P.; Meinshausen, M.; Arora, V.K.; Jones, C.D.; Anav, A.; Liddicoat, S.K.; Knutti, R. Uncertainties in CMIP5 climate projections due to carbon cycle feedbacks. J. Clim. 2013, 27, 511–526. [Google Scholar] [CrossRef]
- Zhu, Z.; Piao, S.; Myneni, R.B.; Huang, M.; Zeng, Z.; Canadell, J.G.; Ciais, P.; Sitch, S.; Friedlingstein, P.; Arneth, A.; et al. Greening of the Earth and its drivers. Nat. Clim. Chang. 2016, 6, 791–795. [Google Scholar] [CrossRef]
- Mao, J.; Ribes, A.; Yan, B.; Shi, X.; Thornton, P.E.; Séférian, R.; Ciais, P.; Myneni, R.B.; Douville, H.; Piao, S.; et al. Human-induced greening of the northern extratropical land surface. Nat. Clim. Chang. 2016, 6, 959–963. [Google Scholar] [CrossRef]
- Donohue, R.J.; Roderick, M.L.; McVicar, T.R.; Farquhar, G.D. Impact of CO2 fertilization on maximum foliage cover across the globe’s warm, arid environments. Geophys. Res. Lett. 2013, 40, 3031–3035. [Google Scholar] [CrossRef]
- Pan, Y.; Birdsey, R.A.; Fang, J.; Houghton, R.; Kauppi, P.E.; Kurz, W.A.; Phillips, O.L.; Shvidenko, A.; Lewis, S.L.; Canadell, J.G.; et al. A large and persistent carbon sink in the world’s forests. Science 2011, 333, 988–993. [Google Scholar] [CrossRef] [PubMed]
- Ciais, P.; Sabine, C.; Bala, G.; Bopp, L.; Brovkin, V.; Canadell, J.; Chhabra, A.; DeFries, R.; Galloway, J.; Heimann, M.; et al. Carbon and other biogeochemical cycles. In Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Stocker, T.F., Qin, D., Plattner, G.K., Tignor, M., Allen, S.K., Boschung, J., Nauels, A., Xia, Y., Bex, V., Midgley, P.M., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013; pp. 465–570. ISBN 9781107661820. [Google Scholar]
- Schimel, D.; Stephens, B.B.; Fisher, J.B. Effect of increasing CO2 on the terrestrial carbon cycle. Proc. Natl. Acad. Sci. USA 2015, 112, 436–441. [Google Scholar] [CrossRef] [PubMed]
- Devaraju, N.; Bala, G.; Caldeira, K.; Nemani, R. A model based investigation of the relative importance of CO2-fertilization, climate warming, nitrogen deposition and land use change on the global terrestrial carbon uptake in the historical period. Clim. Dyn. 2016, 47, 173–190. [Google Scholar] [CrossRef]
- Nemani, R.R.; Keeling, C.D.; Hashimoto, H.; Jolly, W.M.; Piper, S.C.; Tucker, C.J.; Myneni, R.B.; Running, S.W. Climate-driven increases in global terrestrial net primary production from 1982 to 1999. Science 2003, 300, 1560–1563. [Google Scholar] [CrossRef] [PubMed]
- Norby, R.J.; Zak, D.R. Ecological Lessons from Free-Air CO2 Enrichment (FACE) Experiments. Annu. Rev. Ecol. Evol. Syst. 2011, 42, 181–203. [Google Scholar] [CrossRef]
- Keenan, T.F.; Hollinger, D.Y.; Bohrer, G.; Dragoni, D.; Munger, J.W.; Schmid, H.P.; Richardson, A.D. Increase in forest water-use efficiency as atmospheric carbon dioxide concentrations rise. Nature 2013, 499, 324–327. [Google Scholar] [CrossRef] [PubMed]
- Bonan, G.B.; Pollard, D.; Thompson, S.L. Effects of boreal forest vegetation on global climate. Nature 1992, 359, 716–718. [Google Scholar] [CrossRef]
- Myneni, R.B.; Keeling, C.D.; Tucker, C.J.; Asrar, G.; Nemani, R.R. Increased plant growth in the northern high latitudes from 1981 to 1991. Nature 1997, 386, 698–702. [Google Scholar] [CrossRef]
- Smith, W.K.; Reed, S.C.; Cleveland, C.C.; Ballantyne, A.P.; Anderegg, W.R.L.; Wieder, W.R.; Liu, Y.Y.; Running, S.W. Large divergence of satellite and Earth system model estimates of global terrestrial CO2 fertilization. Nat. Clim. Chang. 2015, 6, 306–310. [Google Scholar] [CrossRef]
- Lemordant, L.; Gentine, P.; Swann, A.S.; Cook, B.I.; Scheff, J. Critical impact of vegetation physiology on the continental hydrologic cycle in response to increasing CO2. Proc. Natl. Acad. Sci. USA 2018, 115, 4093–4098. [Google Scholar] [CrossRef] [PubMed]
- Chadwick, R.; Good, P.; Martin, G.; Rowell, D.P. Large rainfall changes consistently projected over substantial areas of tropical land. Nat. Clim. Chang. 2015, 6, 177. [Google Scholar] [CrossRef]
- Kumar, S.; Allan, R.P.; Zwiers, F.; Lawrence, D.M.; Dirmeyer, P.A. Revisiting trends in wetness and dryness in the presence of internal climate variability and water limitations over land. Geophys. Res. Lett. 2015, 42, 10867–10875. [Google Scholar] [CrossRef]
- Polson, D.; Hegerl, G.C. Strengthening contrast between precipitation in tropical wet and dry regions. Geophys. Res. Lett. 2017, 44, 365–373. [Google Scholar] [CrossRef]
- Taylor, K.E.; Stouffer, R.J.; Meehl, G.A.; Taylor, K.E.; Stouffer, R.J.; Meehl, G.A. An overview of CMIP5 and the experiment design. Bull. Am. Meteorol. Soc. 2012, 93, 485–498. [Google Scholar] [CrossRef]
- Riahi, K.; Rao, S.; Krey, V.; Cho, C.; Chirkov, V.; Fischer, G.; Kindermann, G.; Nakicenovic, N.; Rafaj, P. RCP 8.5—A scenario of comparatively high greenhouse gas emissions. Clim. Chang. 2011, 109, 33–57. [Google Scholar] [CrossRef]
- Christensen, J.H.; Krishna, K.K.; Aldrian, E.; An, S.-I.; Cavalcanti, I.F.A.; de Castro, M.; Dong, W.; Goswami, P.; Hall, A.; Kanyanga, J.K.; et al. Climate Phenomena and their Relevance for Future Regional Climate Change. In Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Stocker, T.F., Qin, D., Plattner, G.K., Tignor, M., Allen, S.K., Boschung, J., Nauels, A., Xia, Y., Bex, V., Midgley, P.M., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013; pp. 1217–1308. ISBN 978-1-107-66182-0. [Google Scholar]
- Dekker, S.C.; Groenendijk, M.; Booth, B.B.B.; Huntingford, C.; Cox, P.M. Spatial and temporal variations in plant water-use efficiency inferred from tree-ring, eddy covariance and atmospheric observations. Earth Syst. Dyn. 2016, 7, 525–533. [Google Scholar] [CrossRef]
- Boucher, O.; Randall, D.; Artaxo, P.; Bretherton, C.; Feingold, G.; Forster, P.; Kerminen, V.M.; Kondo, Y.; Liao, H.; Lohmann, U.; et al. Clouds and aerosols. In Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Stocker, T.F., Qin, D., Plattner, G.K., Tignor, M., Allen, S.K., Boschung, J., Nauels, A., Xia, Y., Bex, V., Midgley, P.M., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013; pp. 571–658. ISBN 9781107661820. [Google Scholar]
- Zhu, Z.; Bi, J.; Pan, Y.; Ganguly, S.; Anav, A.; Xu, L.; Samanta, A.; Piao, S.; Nemani, R.R.; Myneni, R.B. Global Data Sets of Vegetation Leaf Area Index (LAI)3g and Fraction of Photosynthetically Active Radiation (FPAR)3g Derived from Global Inventory Modeling and Mapping Studies (GIMMS) Normalized Difference Vegetation Index (NDVI)3g for the period 1981 to 2. Remote Sens. 2013, 5, 927–948. [Google Scholar] [CrossRef]
- New, M.; Lister, D.; Hulme, M.; Makin, I. A high-resolution data set of surface climate over global land areas. Clim. Res. 2002, 21, 1–25. [Google Scholar] [CrossRef]
- Eastman, R.; Warren, S.G. A 39-yr survey of cloud changes from land stations worldwide 1971–2009: Long-term trends, relation to aerosols, and expansion of the tropical belt. J. Clim. 2013, 26, 1286–1303. [Google Scholar] [CrossRef]
- Warren, S.G.; Eastman, R.M.; Hahn, C.J.; Warren, S.G.; Eastman, R.M.; Hahn, C.J. A Survey of Changes in Cloud Cover and Cloud Types over Land from Surface Observations, 1971–96. J. Clim. 2007, 20, 717–738. [Google Scholar] [CrossRef]
- Zelinka, M.D.; Klein, S.A.; Taylor, K.E.; Andrews, T.; Webb, M.J.; Gregory, J.M.; Forster, P.M.; Zelinka, M.D.; Klein, S.A.; Taylor, K.E.; et al. Contributions of Different Cloud Types to Feedbacks and Rapid Adjustments in CMIP5. J. Clim. 2013, 26, 5007–5027. [Google Scholar] [CrossRef]
- Cao, L.; Bala, G.; Caldeira, K. Climate response to changes in atmospheric carbon dioxide and solar irradiance on the time scale of days to weeks. Environ. Res. Lett. 2012, 7, 034015. [Google Scholar] [CrossRef]
- Drake, B.G.; Gonzàlez-Meler, M.A.; Long, S.P. More efficient plants: A consequence of rising atmospheric CO2? Annu. Rev. Plant Biol. 1997, 48, 609–639. [Google Scholar] [CrossRef]
- Kendall, M.G. Rank Correlation Methods; Griffin: London, UK, 1975; ISBN 9780852641996. [Google Scholar]
- Mystakidis, S.; Davin, E.L.; Gruber, N.; Seneviratne, S.I. Constraining future terrestrial carbon cycle projections using observation-based water and carbon flux estimates. Glob. Chang. Biol. 2016, 22, 2198–2215. [Google Scholar] [CrossRef]
- Mahowald, N.; Lo, F.; Zheng, Y.; Harrison, L.; Funk, C.; Lombardozzi, D.; Goodale, C. Projections of leaf area index in earth system models. Earth Syst. Dyn. 2016, 7, 211–229. [Google Scholar] [CrossRef]
- Norris, J.R.; Allen, R.J.; Evan, A.T.; Zelinka, M.D.; O’Dell, C.W.; Klein, S.A. Evidence for climate change in the satellite cloud record. Nature 2016, 536, 72–75. [Google Scholar] [CrossRef]
- Cao, L.; Bala, G.; Caldeira, K.; Nemani, R.; Ban-Weiss, G. Importance of carbon dioxide physiological forcing to future climate change. Proc. Natl. Acad. Sci. USA 2010, 107, 9513–9518. [Google Scholar] [CrossRef] [PubMed]
- Forkel, M.; Carvalhais, N.; Rödenbeck, C.; Keeling, R.; Heimann, M.; Thonicke, K.; Zaehle, S.; Reichstein, M. Enhanced seasonal CO2 exchange caused by amplified plant productivity in northern ecosystems. Science 2016, 351, 696–699. [Google Scholar] [CrossRef] [PubMed]
- Bathiany, S.; Claussen, M.; Brovkin, V.; Bathiany, S.; Claussen, M.; Brovkin, V. CO2-induced Sahel greening in three CMIP5 earth system models. J. Clim. 2014, 27, 7163–7184. [Google Scholar] [CrossRef]
- Wang, L.; Cole, J.N.S.; Bartlett, P.; Verseghy, D.; Derksen, C.; Brown, R.; von Salzen, K. Investigating the spread in surface albedo for snow-covered forests in CMIP5 models. J. Geophys. Res. Atmos. 2016, 121, 1104–1119. [Google Scholar] [CrossRef]
- Loranty, M.M.; Berner, L.T.; Goetz, S.J.; Jin, Y.; Randerson, J.T. Vegetation controls on northern high latitude snow-albedo feedback: Observations and CMIP5 model simulations. Glob. Chang. Biol. 2014, 20, 594–606. [Google Scholar] [CrossRef] [PubMed]
- Dirmeyer, P.A.; Brubaker, K.L.; Dirmeyer, P.A.; Brubaker, K.L. Characterization of the Global Hydrologic Cycle from a Back-Trajectory Analysis of Atmospheric Water Vapor. J. Hydrometeorol. 2007, 8, 20–37. [Google Scholar] [CrossRef]
- Franks, P.J.; Adams, M.A.; Amthor, J.S.; Barbour, M.M.; Berry, J.A.; Ellsworth, D.S.; Farquhar, G.D.; Ghannoum, O.; Lloyd, J.; McDowell, N.; et al. Sensitivity of plants to changing atmospheric CO2 concentration: From the geological past to the next century. New Phytol. 2013, 197, 1077–1094. [Google Scholar] [CrossRef]
- Maxbauer, D.P.; Royer, D.L.; LePage, B.A. High Arctic forests during the middle Eocene supported by moderate levels of atmospheric CO2. Geology 2014, 42, 1027–1030. [Google Scholar] [CrossRef]
- Kamae, Y.; Watanabe, M. Tropospheric adjustment to increasing CO2: Its timescale and the role of land-sea contrast. Clim. Dyn. 2013, 41, 3007–3024. [Google Scholar] [CrossRef]
- Modak, A.; Bala, G.; Cao, L.; Caldeira, K. Why must a solar forcing be larger than a CO2 forcing to cause the same global mean surface temperature change? Environ. Res. Lett. 2016, 11, 044013. [Google Scholar] [CrossRef]
- Le Quéré, C.; Peters, G.P.; Andres, R.J.; Andrew, R.M.; Boden, T.A.; Ciais, P.; Friedlingstein, P.; Houghton, R.A.; Marland, G.; Moriarty, R.; et al. Global carbon budget 2013. Earth Syst. Sci. Data 2014, 6, 235–263. [Google Scholar] [CrossRef]
- Graven, H.D.; Keeling, R.F.; Piper, S.C.; Patra, P.K.; Stephens, B.B.; Wofsy, S.C.; Welp, L.R.; Sweeney, C.; Tans, P.P.; Kelley, J.J.; et al. Enhanced seasonal exchange of CO2 by Northern ecosystems since 1960. Science 2013, 341, 1085–1089. [Google Scholar] [CrossRef] [PubMed]
- Reichstein, M.; Bahn, M.; Ciais, P.; Frank, D.; Mahecha, M.D.; Seneviratne, S.I.; Zscheischler, J.; Beer, C.; Buchmann, N.; Frank, D.C.; et al. Climate extremes and the carbon cycle. Nature 2013, 500, 287–295. [Google Scholar] [CrossRef] [PubMed]
- Salzmann, U.; Haywood, A.M.; Lunt, D.J. The past is a guide to the future? Comparing Middle Pliocene vegetation with predicted biome distributions for the twenty-first century. Philos. Trans. R. Soc. A Math. Phys. Eng. Sci. 2009, 367, 189–204. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Wang, Y.P.; Matear, R.J.; Pitman, A.J.; Dai, Y.J. Nitrogen and phosphorous limitations significantly reduce future allowable CO2 emissions. Geophys. Res. Lett. 2014, 41, 632–637. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model | Modeling Group | Land Component | N Cycle | Dynamic Vegetation |
|---|---|---|---|---|
| bcc-csm1-1 | Beijing Climate Center, China Meteorological Administration, CHINA | AVIM1.0 | N | N |
| bcc-csm1-1-m | Meteorological Administration, CHINA | AVIM1.0 | N | N |
| BNU-ESM | Beijing Normal University, CHINA | CoLM3 & BNU DGVM (C/N) | - | - |
| CanESM2 | Canadian Center for Climate Modelling and Analysis, CANADA | CLASS2.7 & CTEM1 | N | N |
| CESM1-CAM5 | Community Earth System Model Contributors, NSF-DOE-NCAR, USA | CLM4 | Y | N |
| CESM1-WACCM | Community Earth System Model Contributors, NSF-DOE-NCAR, USA | CLM4 | Y | N |
| CESM1-BGC | Community Earth System Model Contributors, NSF-DOE-NCAR, USA | CLM4 | Y | N |
| GFDL-CM3 | NOAA Geophysical Fluid Dynamics Laboratory, USA | LM3 | N | Y |
| GFDL-ESM2G | NOAA Geophysical Fluid Dynamics Laboratory, USA | LM3 | N | Y |
| GFDL-ESM2M | NOAA Geophysical Fluid Dynamics Laboratory, USA | LM3 | N | Y |
| HadGEM2-ES | Met Office Hadley Centre, UNITED KINDOM | MOSES2 & TRIFFID | N | Y |
| INMCM4 | Russia | |||
| IPSL-CM5A-LR | Institut Pierre-Simon Laplace, FRANCE | ORCHIDEE | N | N |
| IPSL-CM5A-MR | Institut Pierre-Simon Laplace, FRANCE | ORCHIDEE | N | N |
| MIROC5 | JAMSTEC, University of Tokyo, and NIES, JAPAN | MATSIRO & SEIB-DGVM | N | Y |
| MIROC-ESM-CHEM | JAMSTEC, University of Tokyo, and NIES, JAPAN | MATSIRO & SEIB-DGVM | N | Y |
| MIROC-ESM | JAMSTEC, University of Tokyo, and NIES, JAPAN | MATSIRO & SEIB-DGVM | N | Y |
| MPI-ESM-LR | Max Planck Institute for Meteorology, GERMANY | JSBACH | N | Y |
| MPI-ESM-MR | Max Planck Institute for Meteorology, GERMANY | JSBACH | N | Y |
| MPI-ESM1 | Max Planck Institute for Meteorology, GERMANY | JSBACH | N | Y |
| NorESM1-ME | Norwegian Climate Centre, NORWAY | CLM4 | Y | N |
| NorESM1-M | Norwegian Climate Centre, NORWAY | CLM4 | Y | N |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hashimoto, H.; Nemani, R.R.; Bala, G.; Cao, L.; Michaelis, A.R.; Ganguly, S.; Wang, W.; Milesi, C.; Eastman, R.; Lee, T.; et al. Constraints to Vegetation Growth Reduced by Region-Specific Changes in Seasonal Climate. Climate 2019, 7, 27. https://doi.org/10.3390/cli7020027
Hashimoto H, Nemani RR, Bala G, Cao L, Michaelis AR, Ganguly S, Wang W, Milesi C, Eastman R, Lee T, et al. Constraints to Vegetation Growth Reduced by Region-Specific Changes in Seasonal Climate. Climate. 2019; 7(2):27. https://doi.org/10.3390/cli7020027
Chicago/Turabian StyleHashimoto, Hirofumi, Ramakrishna R. Nemani, Govindasamy Bala, Long Cao, Andrew R. Michaelis, Sangram Ganguly, Weile Wang, Cristina Milesi, Ryan Eastman, Tsengdar Lee, and et al. 2019. "Constraints to Vegetation Growth Reduced by Region-Specific Changes in Seasonal Climate" Climate 7, no. 2: 27. https://doi.org/10.3390/cli7020027