
Expression of Endogenous Putative TSH Binding Protein in Orbit

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Tissues Studied; In Vitro Culture and Ex Vivo Samples
2.2. In Vitro Adipogenesis
2.3. QPCR Analysis
2.4. Production of Antibody Specific for TSHRv
2.5. Western Blot Analysis
2.6. Measurement of TSHR Activation
2.7. Statistical Analysis
3. Results
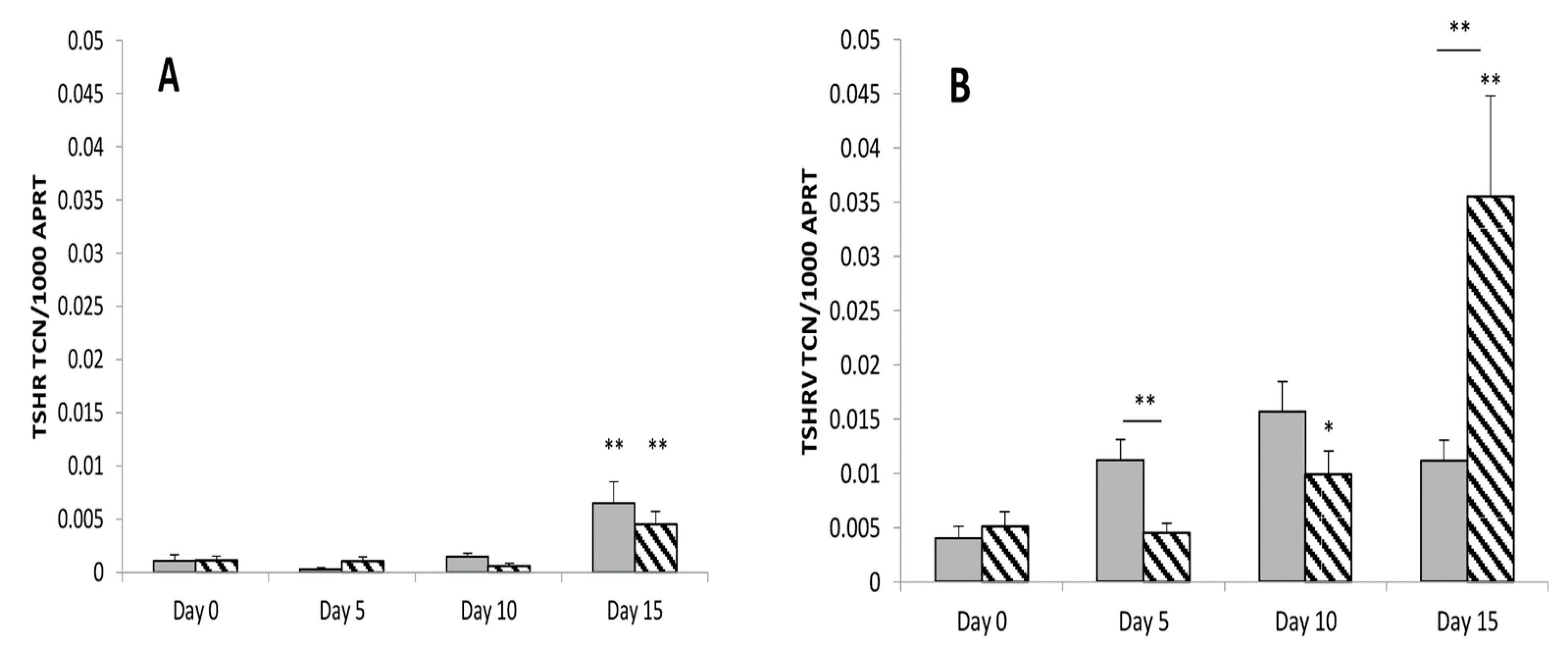
3.1. TSHRv Transcripts Are Expressed at Higher Levels than Full-Length TSHR
3.2. There Is No Role of Thyrostimulin in GO
3.3. Transcripts Expression of Thyrostimulin and TSHRv in Ex Vivo Samples
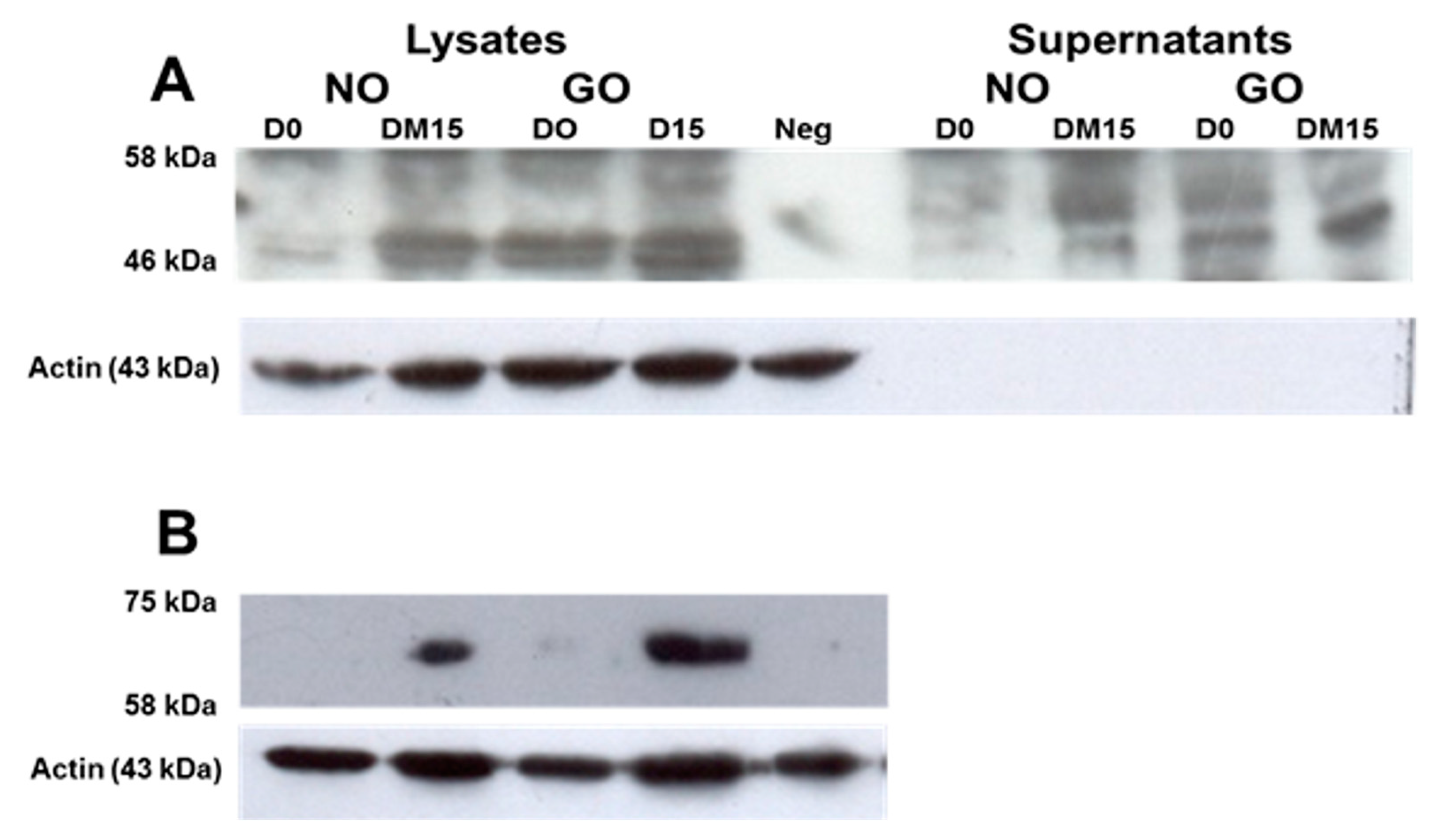
3.4. TSHRv and TSHR Protein Are Detectable in OF
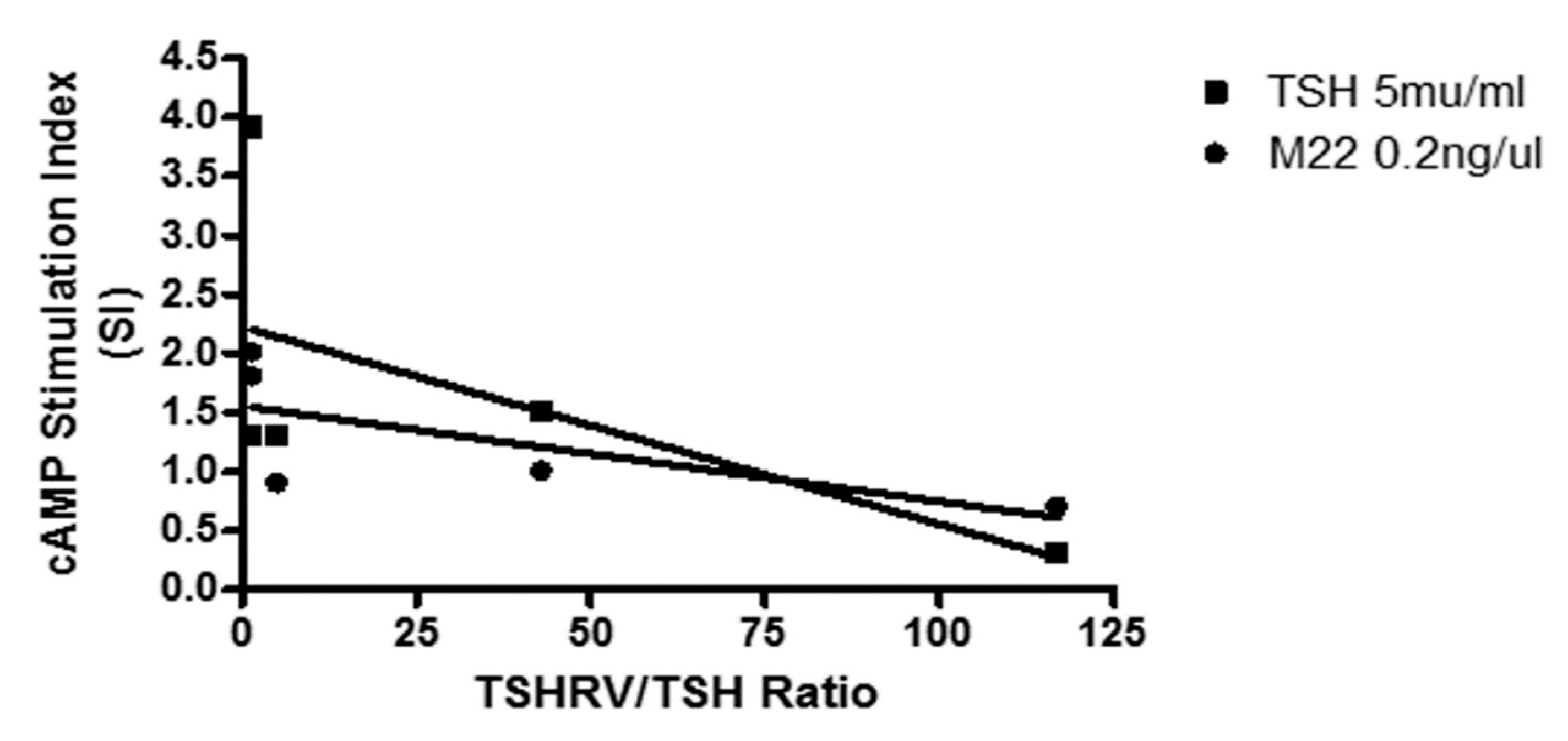
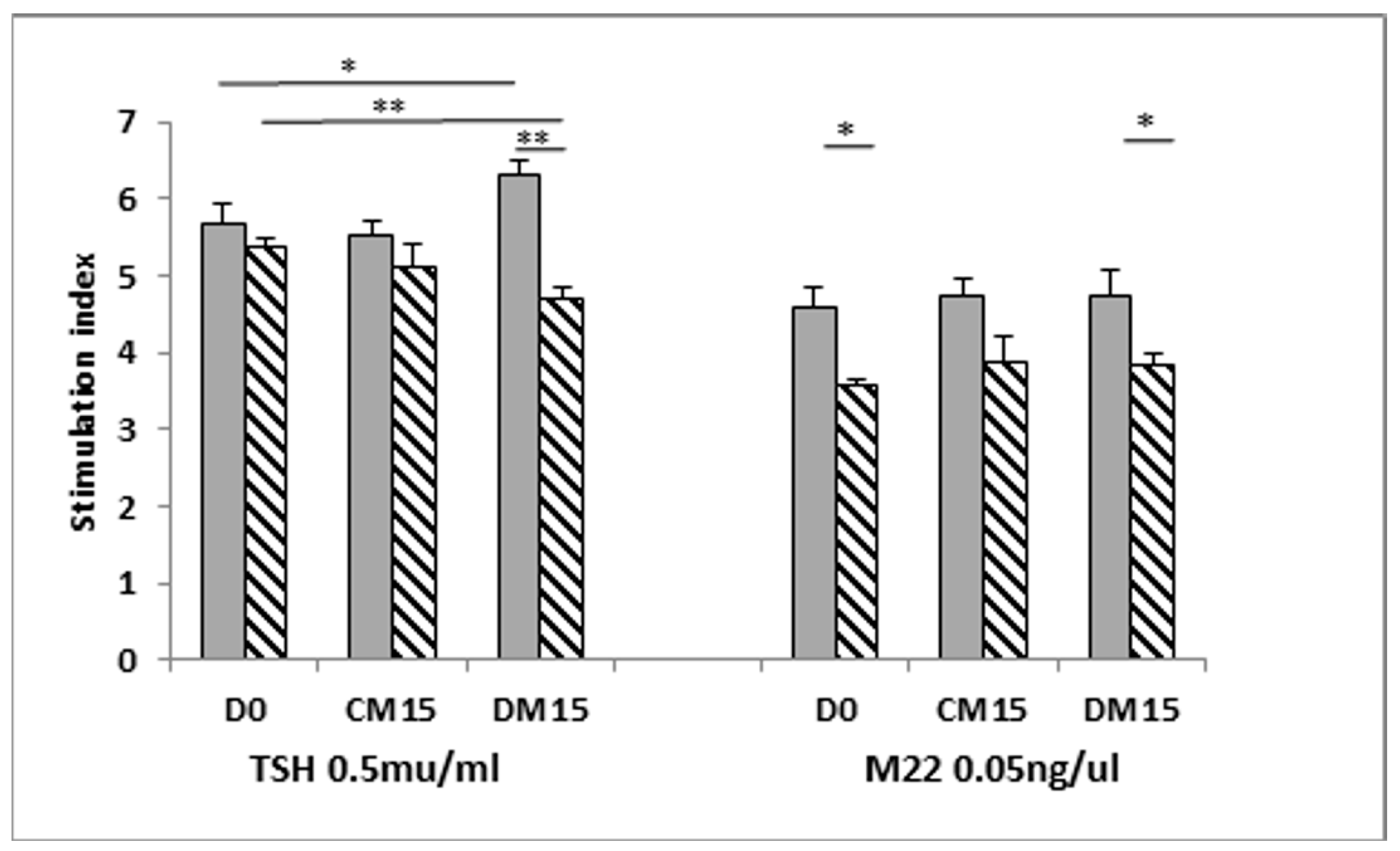
3.5. Is the TSHRv Functional?
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| GO | Graves’ Orbitopathy |
| TSHR | Thyrotropin receptor |
| TSHRv | TSHR variants |
| OF | Orbital preadipocytes/fibroblasts |
| TSAB | Thyroid stimulating antibodies |
| GD | Graves’ disease |
References
- Reddy, S.V.; Jain, A.; Yadav, S.B.; Sharma, K.; Bhatia, E. Prevalence of Graves’ ophthalmopathy in patients with Graves’ disease presenting to a referral centre in north India. Indian J. Med. Res. 2014, 139, 99–104. [Google Scholar]
- Tellez, M.; Cooper, J.; Edmonds, C. Graves’ ophthalmopathy in relation to cigarette smoking and ethnic origin. Clin. Endocrinol. 1992, 36, 291–294. [Google Scholar] [CrossRef] [PubMed]
- Wiersinga, W.M.; Bartalena, L. Epidemiology and prevention of Graves’ ophthalmopathy. Thyroid 2002, 12, 855–860. [Google Scholar] [CrossRef]
- Tanda, M.L.; Piantanida, E.; Liparulo, L.; Veronesi, G.; Lai, A.; Sassi, L.; Pariani, N.; Gallo, D.; Azzolini, C.; Ferrario, M.; et al. Prevalence and natural history of Graves’ orbitopathy in a large series of patients with newly diagnosed graves’ hyperthyroidism seen at a single center. J. Clin. Endocrinol. Metab. 2013, 98, 1443–1449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crisp, M.; Starkey, K.J.; Lane, C.; Ham, J.; Ludgate, M. Adipogenesis in thyroid eye disease. Investig. Ophthalmol. Vis. Sci. 2000, 41, 3249–3255. [Google Scholar]
- Kumar, S.; Coenen, M.J.; Scherer, P.E.; Bahn, R.S. Evidence for enhanced adipogenesis in the orbits of patients with Graves’ ophthalmopathy. J. Clin. Endocrinol. Metab. 2004, 89, 930–935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bahn, R.S.; Dutton, C.M.; Natt, N.; Joba, W.; Spitzweg, C.; Heufelder, A.E. Thyrotropin receptor expression in Graves’ orbital adipose/connective tissues: Potential autoantigen in Graves’ ophthalmopathy. J. Clin. Endocrinol. Metab. 1998, 83, 998–1002. [Google Scholar] [CrossRef]
- Roselli-Rehfuss, L.; Robbins, L.S.; Cone, R.D. Thyrotropin receptor messenger ribonucleic acid is expressed in most brown and white adipose tissues in the guinea pig. Endocrinology 1992, 130, 1857–1861. [Google Scholar]
- Khoo, D.H.; Ho, S.C.; Seah, L.L.; Fong, K.S.; Tai, E.S.; Chee, S.P.; Eng, P.H.; Aw, S.E.; Fok, A.C. The combination of absent thyroid peroxidase antibodies and high thyroid-stimulating immunoglobulin levels in Graves’ disease identifies a group at markedly increased risk of ophthalmopathy. Thyroid 1999, 9, 1175–1180. [Google Scholar] [CrossRef]
- Wiersinga, W.M. Quality of life in Graves’ ophthalmopathy. Best Pract. Res. Clin. Endocrinol. Metab. 2012, 26, 359–370. [Google Scholar] [CrossRef]
- Coulter, I.; Frewin, S.; Krassas, G.E.; Perros, P. Psychological implications of Graves’ orbitopathy. Eur. J. Endocrinol. Eur. Fed. Endocr. Soc. 2007, 157, 127–131. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Baker, G.; Janus, D.; Paddon, C.A.; Fuhrer, D.; Ludgate, M. Biological effects of thyrotropin receptor activation on human orbital preadipocytes. Investig. Ophthalmol. Vis. Sci. 2006, 47, 5197–5203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Paddon, C.; Lewis, M.D.; Grennan-Jones, F.; Ludgate, M. Gsalpha signalling suppresses PPARgamma2 generation and inhibits 3T3L1 adipogenesis. J. Endocrinol. 2009, 202, 207–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakabayashi, K.; Matsumi, H.; Bhalla, A.; Bae, J.; Mosselman, S.; Hsu, S.Y.; Hsueh, A.J. Thyrostimulin, a heterodimer of two new human glycoprotein hormone subunits, activates the thyroid-stimulating hormone receptor. J. Clin. Investig. 2002, 109, 1445–1452. [Google Scholar] [CrossRef] [PubMed]
- Bassett, J.H.; van der Spek, A.; Logan, J.G.; Gogakos, A.; Bagchi-Chakraborty, J.; Murphy, E.; van Zeijl, C.; Down, J.; Croucher, P.I.; Boyde, A.; et al. Thyrostimulin Regulates Osteoblastic Bone Formation During Early Skeletal Development. Endocrinology 2015, 156, 3098–3113. [Google Scholar] [CrossRef] [Green Version]
- Okada, S.L.; Ellsworth, J.L.; Durnam, D.M.; Haugen, H.S.; Holloway, J.L.; Kelley, M.L.; Lewis, K.E.; Ren, H.; Sheppard, P.O.; Storey, H.M.; et al. A glycoprotein hormone expressed in corticotrophs exhibits unique binding properties on thyroid-stimulating hormone receptor. Mol. Endocrinol. 2006, 20, 414–425. [Google Scholar] [CrossRef] [Green Version]
- Paschke, R.; Metcalfe, A.; Alcalde, L.; Vassart, G.; Weetman, A.; Ludgate, M. Presence of nonfunctional thyrotropin receptor variant transcripts in retroocular and other tissues. J. Clin. Endocrinol. Metab. 1994, 79, 1234–1238. [Google Scholar]
- Quellari, M.; Desroches, A.; Beau, I.; Beaudeux, E.; Misrahi, M. Role of cleavage and shedding in human thyrotropin receptor function and trafficking. Eur. J. Biochem. 2003, 270, 3486–3497. [Google Scholar] [CrossRef] [PubMed]
- Rapoport, B.; McLachlan, S.M. The thyrotropin receptor in Graves’ disease. Thyroid 2007, 17, 911–922. [Google Scholar] [CrossRef]
- Zhao, S.X.; Tsui, S.; Cheung, A.; Douglas, R.S.; Smith, T.J.; Banga, J.P. Orbital fibrosis in a mouse model of Graves’ disease induced by genetic immunization of thyrotropin receptor cDNA. J. Endocrinol. 2011, 210, 369–377. [Google Scholar] [CrossRef]
- Moshkelgosha, S.; So, P.W.; Diaz-Cano, S.; Banga, J.P. Preclinical models of Graves’ disease and associated secondary complications. Curr. Pharm. Des. 2015, 21, 2414–2421. [Google Scholar] [CrossRef] [PubMed]
- Fuhrer, D.; Lewis, M.D.; Alkhafaji, F.; Starkey, K.; Paschke, R.; Wynford-Thomas, D.; Eggo, M.; Ludgate, M. Biological activity of activating thyroid-stimulating hormone receptor mutants depends on the cellular context. Endocrinology 2003, 144, 4018–4030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, C.; Morgenthaler, N.G.; Lee, S.; Llewellyn, D.H.; Clifton-Bligh, R.; John, R.; Lazarus, J.H.; Chatterjee, V.K.; Ludgate, M. Development of a luminescent bioassay for thyroid stimulating antibodies. J. Clin. Endocrinol. Metab. 1999, 84, 374–377. [Google Scholar] [CrossRef] [PubMed]
- Draman, M.S.; Grennan-Jones, F.; Zhang, L.; Taylor, P.N.; Tun, T.K.; McDermott, J.; Moriarty, P.; Morris, D.; Lane, C.; Sreenan, S.; et al. Effects of prostaglandin F(2alpha) on adipocyte biology relevant to graves’ orbitopathy. Thyroid 2013, 23, 1600–1608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lantz, M.; Vondrichova, T.; Capretz, A.; Nilsson, E.; Frenander, C.; Bondeson, A.G.; Ridderstrale, M.; Aberg, M.; Asman, P.; Groop, L.; et al. Thyrostimulin (a TSH-like Hormone) expression in orbital and thyroid tissue. Thyroid 2007, 17, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.C.; Hsu, P.J.; Wu, F.J.; Li, S.H.; Lu, C.H.; Luo, C.W. Thyrostimulin, but not thyroid-stimulating hormone (TSH), acts as a paracrine regulator to activate the TSH receptor in mammalian ovary. J. Biol. Chem. 2010, 285, 3758–3765. [Google Scholar] [CrossRef] [Green Version]
- Bodo, E.; Kromminga, A.; Biro, T.; Borbiro, I.; Gaspar, E.; Zmijewski, M.A.; van Beek, N.; Langbein, L.; Slominski, A.T.; Paus, R. Human female hair follicles are a direct, nonclassical target for thyroid-stimulating hormone. J. Investig. Dermatol. 2009, 129, 1126–1139. [Google Scholar] [CrossRef]
- Nagasaki, H.; Wang, Z.; Jackson, V.R.; Lin, S.; Nothacker, H.P.; Civelli, O. Differential expression of the thyrostimulin subunits, glycoprotein alpha2 and beta5 in the rat pituitary. J. Mol. Endocrinol. 2006, 37, 39–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Starkey, K.J.; Janezic, A.; Jones, G.; Jordan, N.; Baker, G.; Ludgate, M. Adipose thyrotrophin receptor expression is elevated in Graves’ and thyroid eye diseases ex vivo and indicates adipogenesis in progress in vivo. J. Mol. Endocrinol. 2003, 30, 369–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valyasevi, R.W.; Erickson, D.Z.; Harteneck, D.A.; Dutton, C.M.; Heufelder, A.E.; Jyonouchi, S.C.; Bahn, R.S. Differentiation of human orbital preadipocyte fibroblasts induces expression of functional thyrotropin receptor. J. Clin. Endocrinol. Metab. 1999, 84, 2557–2562. [Google Scholar] [CrossRef]
- Bell, A.; Gagnon, A.; Grunder, L.; Parikh, S.J.; Smith, T.J.; Sorisky, A. Functional TSH receptor in human abdominal preadipocytes and orbital fibroblasts. Am. J. Physiol. Cell Physiol. 2000, 279, C335–C340. [Google Scholar] [CrossRef] [PubMed]
- van Zeijl, C.J.; Fliers, E.; van Koppen, C.J.; Surovtseva, O.V.; de Gooyer, M.E.; Mourits, M.P.; Wiersinga, W.M.; Miltenburg, A.M.; Boelen, A. Thyrotropin receptor-stimulating Graves’ disease immunoglobulins induce hyaluronan synthesis by differentiated orbital fibroblasts from patients with Graves’ ophthalmopathy not only via cyclic adenosine monophosphate signaling pathways. Thyroid 2011, 21, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Neumann, S.; Pope, A.; Geras-Raaka, E.; Raaka, B.M.; Bahn, R.S.; Gershengorn, M.C. A Drug-Like Antagonist Inhibits Thyrotropin Receptor–Mediated Stimulation of cAMP Production in Graves’ Orbital Fibroblasts. Thyroid 2012, 22, 839–843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chazenbalk, G.D.; Jaume, J.C.; McLachlan, S.M.; Rapoport, B. Engineering the human thyrotropin receptor ectodomain from a non-secreted form to a secreted, highly immunoreactive glycoprotein that neutralizes autoantibodies in Graves’ patients’ sera. J. Biol. Chem. 1997, 272, 18959–18965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagayama, Y.; Wadsworth, H.L.; Russo, D.; Chazenbalk, G.D.; Rapoport, B. Binding domains of stimulatory and inhibitory thyrotropin (TSH) receptor autoantibodies determined with chimeric TSH-lutropin/chorionic gonadotropin receptors. J. Clin. Investig. 1991, 88, 336–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ploski, R.; Brand, O.J.; Jurecka-Lubieniecka, B.; Franaszczyk, M.; Kula, D.; Krajewski, P.; Karamat, M.A.; Simmonds, M.J.; Franklyn, J.A.; Gough, S.C.; et al. Thyroid stimulating hormone receptor (TSHR) intron 1 variants are major risk factors for Graves’ disease in three European Caucasian cohorts. PLoS ONE 2010, 5, e15512. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession Number | Forward | Reverse | Amplicon Size (bp) | |
|---|---|---|---|---|
| hAPRT | NM_001030018.1 | GCTGCGTGCTCATCCGAAAG | CCTTAAGCGAGGTCAGCTCC | 247 |
| hLPL | NC_000008.11 | GAGATTTCTCTGTATGGACC | CTGCAAATGAGACACTTTCTC | 275 |
| hTSHR | NM_000369 | GTGTCACTGCCCTTCCATCCA | GGGGCTATTCAAGGCATTCACAGA | 254 |
| hTSHRv | NM_001018036 | CCTCCTAAAGTTCCTTGGCATT | AGGACTTTCTTCCAAGAGGTAG | 338 |
| hα2 | NM_130769.3 | CTCGGAAGTGATGCCTATGGC | CTAGTAGCGAGAGAGGCGAC | 400 |
| hβ5 | AF403430 | ATGAAGCTGGCATTCCTCTTC | CTGTTGGGCAGCTTGACAGTC | 296 |
| Day of Culture | Treatment | cAMP Stimulation Index (SI) | |
|---|---|---|---|
| NO | GO | ||
| Day 0 | TSH | 1.30 ± 0.11 | 0.29 ± 0.08 * |
| M22 | 0.84 ± 0.03 | 0.75 ± 0.02 * | |
| Day 15 | TSH | 3.93 ± 1.64 | 1.55 ± 0.45 |
| M22 | 1.89 ± 0.15 | 1.05 ± 0.28 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Draman, M.S.; Grennan-Jones, F.; Taylor, P.; Muller, I.; Evans, S.; Haridas, A.; Morris, D.S.; Rees, D.A.; Lane, C.; Dayan, C.; et al. Expression of Endogenous Putative TSH Binding Protein in Orbit. Curr. Issues Mol. Biol. 2021, 43, 1794-1804. https://doi.org/10.3390/cimb43030126
Draman MS, Grennan-Jones F, Taylor P, Muller I, Evans S, Haridas A, Morris DS, Rees DA, Lane C, Dayan C, et al. Expression of Endogenous Putative TSH Binding Protein in Orbit. Current Issues in Molecular Biology. 2021; 43(3):1794-1804. https://doi.org/10.3390/cimb43030126
Chicago/Turabian StyleDraman, Mohd Shazli, Fiona Grennan-Jones, Peter Taylor, Ilaria Muller, Sam Evans, Anjana Haridas, Daniel S. Morris, D. Aled Rees, Carol Lane, Colin Dayan, and et al. 2021. "Expression of Endogenous Putative TSH Binding Protein in Orbit" Current Issues in Molecular Biology 43, no. 3: 1794-1804. https://doi.org/10.3390/cimb43030126