Silibinin Upregulates CXCR4 Expression in Cultured Bone Marrow Cells (BMCs) Especially in Pulmonary Arterial Hypertension Rat Model

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Preparation
2.2. Bone Marrow Cell (BMC) Preparation
2.3. Reverse Transcription-Quantitative Polymerase Chain Reaction
2.4. FCM
2.5. Immunohistochemical Analysis
2.6. Hemodyamic Analysis
2.7. Statistical Analyses
3. Results
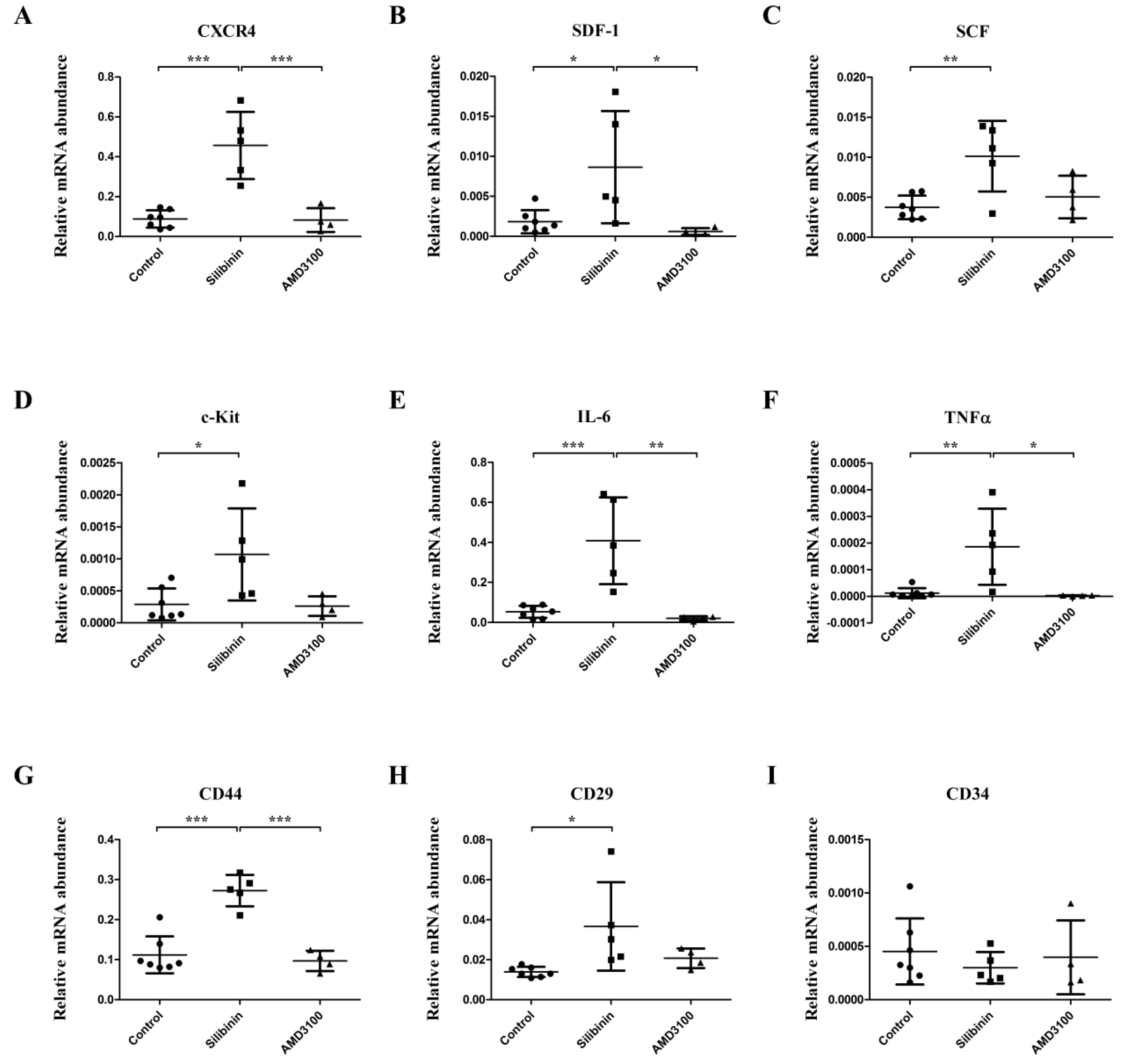
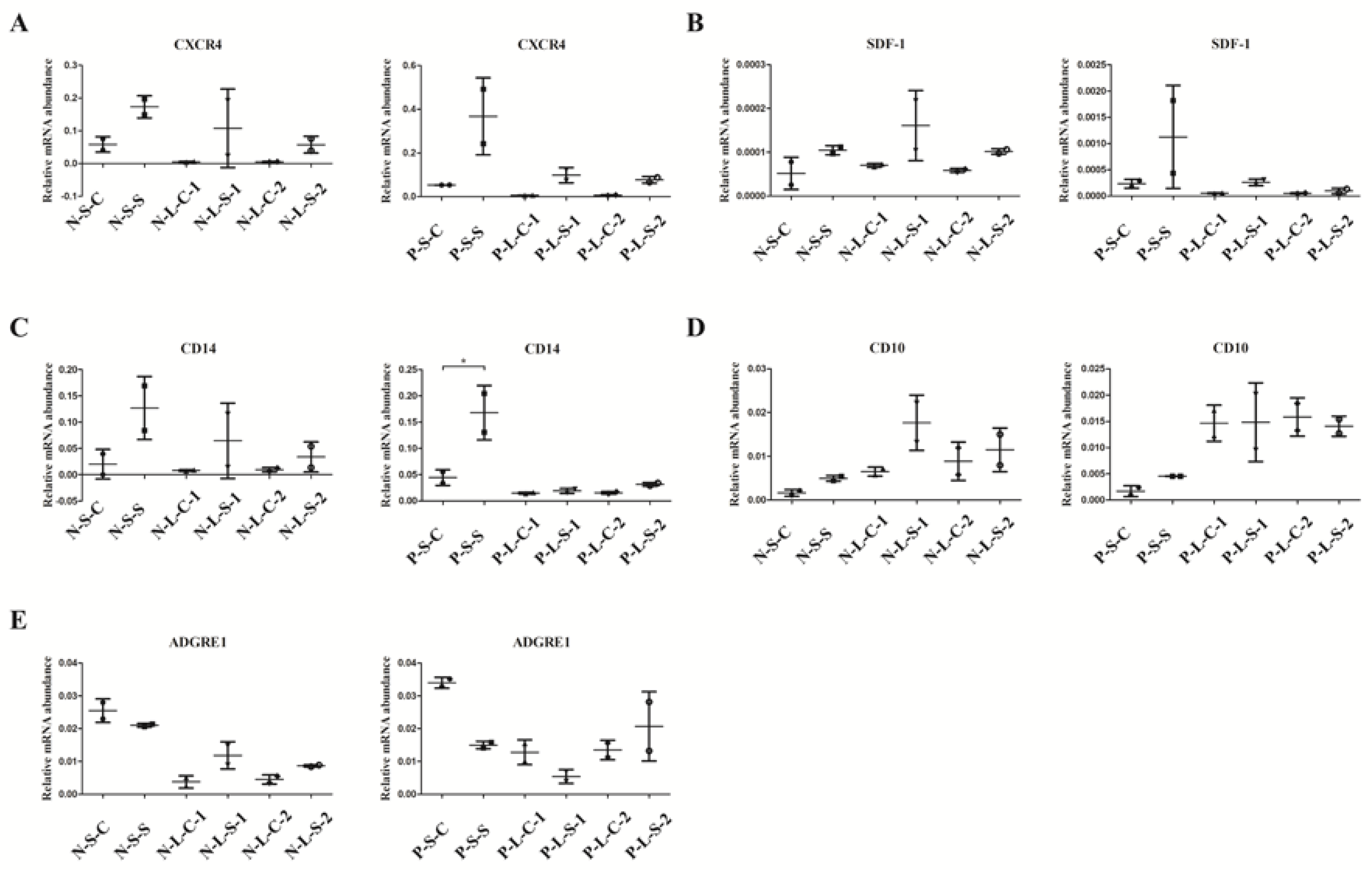
3.1. Effect of Silibinin in the BMCs of PAH Models in Gene Expression Level
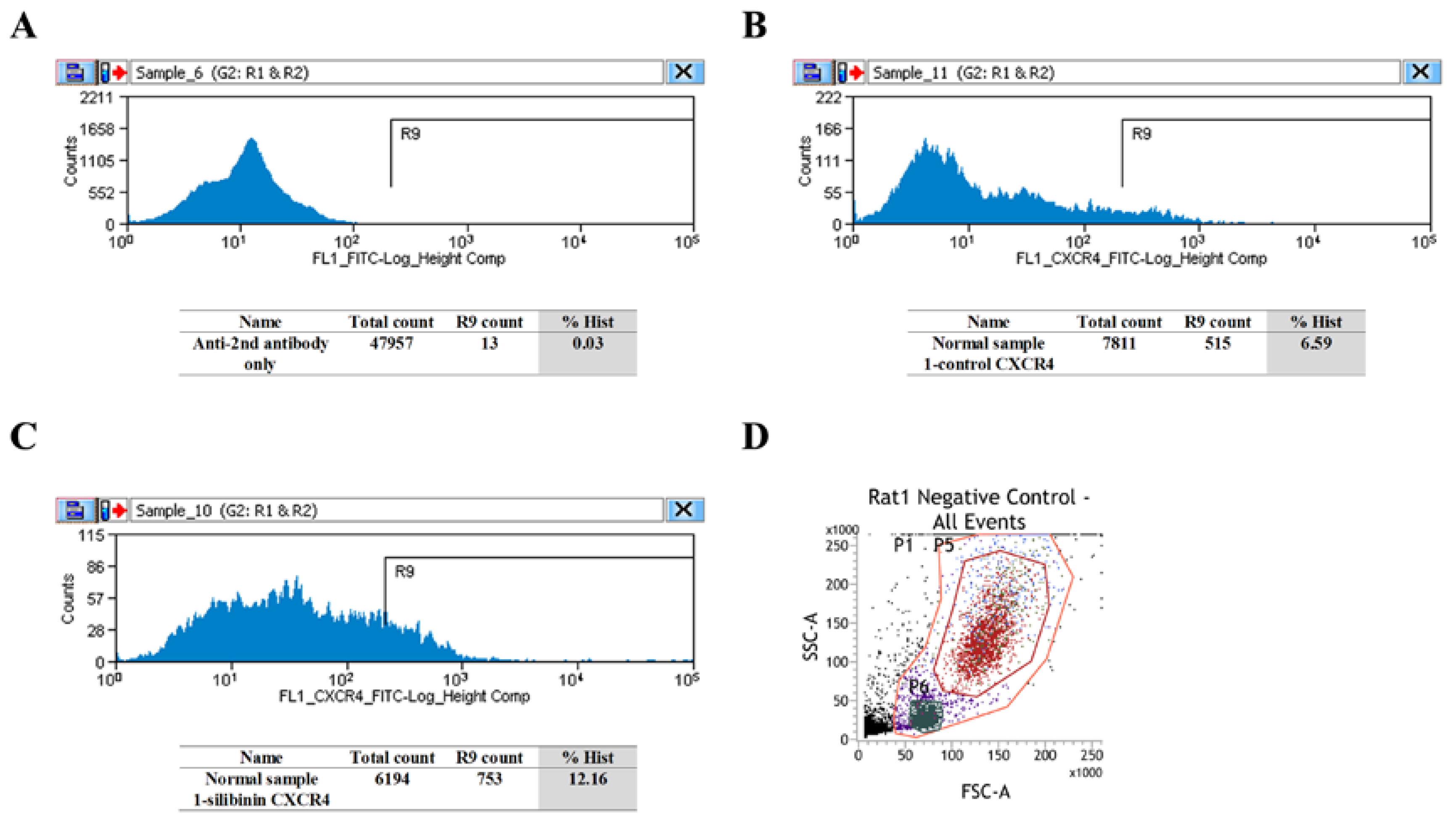
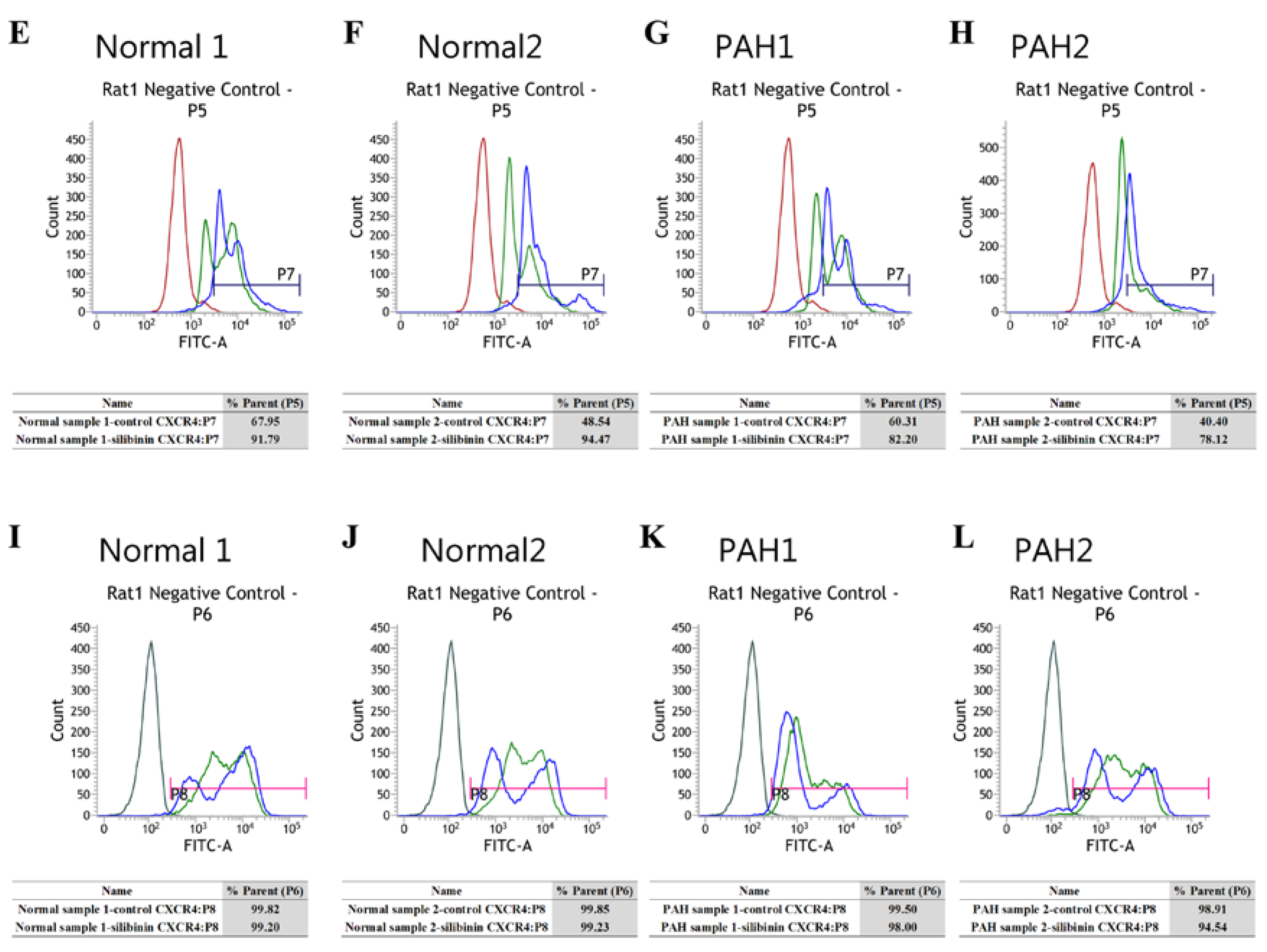
3.2. CXCR4-Positive Cell Population Association with Granulocytes/Monocyte-Macrophage
3.3. Immunohistochemical and Hemodynamic Studies
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tuder, R.M.; Archer, S.L.; Dorfmuller, P.; Erzurum, S.C.; Guignabert, C.; Michelakis, E.; Rabinovitch, M.; Schermuly, R.; Stenmark, K.R.; Morrell, N.W. Relevant issues in the pathology and pathobiology of pulmonary hypertension. J. Am. Coll. Cardiol. 2013, 62, D4–D12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ranchoux, B.; Antigny, F.; Rucker-Martin, C.; Hautefort, A.; Pechoux, C.; Bogaard, H.J.; Dorfmuller, P.; Remy, S.; Lecerf, F.; Plante, S.; et al. Endothelial-to-mesenchymal transition in pulmonary hypertension. Circulation 2015, 131, 1006–1018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galie, N.; Humbert, M.; Vachiery, J.L.; Gibbs, S.; Lang, I.; Torbicki, A.; Simonneau, G.; Peacock, A.; Vonk Noordegraaf, A.; Beghetti, M.; et al. 2015 esc/ers guidelines for the diagnosis and treatment of pulmonary hypertension: The joint task force for the diagnosis and treatment of pulmonary hypertension of the european society of cardiology (esc) and the european respiratory society (ers): Endorsed by: Association for european paediatric and congenital cardiology (aepc), international society for heart and lung transplantation (ishlt). Eur. Heart J. 2016, 37, 67–119. [Google Scholar] [PubMed]
- Chen, H.; Strappe, P.; Chen, S.; Wang, L.X. Endothelial progenitor cells and pulmonary arterial hypertension. Heart Lung Circ. 2014, 23, 595–601. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.X.; Zhang, F.R.; Shang, Y.P.; Zhu, J.H.; Xie, X.D.; Tao, Q.M.; Zhu, J.H.; Chen, J.Z. Transplantation of autologous endothelial progenitor cells may be beneficial in patients with idiopathic pulmonary arterial hypertension: A pilot randomized controlled trial. J. Am. Coll. Cardiol. 2007, 49, 1566–1571. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.X.; Pan, Y.Y.; Zhao, Y.Y.; Wang, X.X. Endothelial progenitor cell-based therapy for pulmonary arterial hypertension. Cell Transpl. 2013, 22, 1325–1336. [Google Scholar] [CrossRef] [Green Version]
- Miller, R.J.; Banisadr, G.; Bhattacharyya, B.J. Cxcr4 signaling in the regulation of stem cell migration and development. J. Neuroimmunol. 2008, 198, 31–38. [Google Scholar] [CrossRef] [Green Version]
- Kawaguchi, N.; Zhang, T.T.; Nakanishi, T. Involvement of cxcr4 in normal and abnormal development. Cells 2019, 8, 185. [Google Scholar] [CrossRef] [Green Version]
- Farkas, D.; Kraskauskas, D.; Drake, J.I.; Alhussaini, A.A.; Kraskauskiene, V.; Bogaard, H.J.; Cool, C.D.; Voelkel, N.F.; Farkas, L. Cxcr4 inhibition ameliorates severe obliterative pulmonary hypertension and accumulation of c-kit(+) cells in rats. PLoS ONE 2014, 9, e89810. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Wu, P.; Huang, F.; Xu, M.; Chen, M.; Huang, K.; Li, G.P.; Xu, M.; Yao, D.; Wang, L. Baicalin attenuates chronic hypoxia-induced pulmonary hypertension via adenosine a2a receptor-induced sdf-1/cxcr4/pi3k/akt signaling. J. Biomed. Sci. 2017, 24, 52. [Google Scholar] [CrossRef]
- Yu, L.; Hales, C.A. Effect of chemokine receptor cxcr4 on hypoxia-induced pulmonary hypertension and vascular remodeling in rats. Respir. Res. 2011, 12, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drummond, S.; Ramachandran, S.; Torres, E.; Huang, J.; Hehre, D.; Suguihara, C.; Young, K.C. Cxcr4 blockade attenuates hyperoxia-induced lung injury in neonatal rats. Neonatology 2015, 107, 304–311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, K.C.; Torres, E.; Hatzistergos, K.E.; Hehre, D.; Suguihara, C.; Hare, J.M. Inhibition of the sdf-1/cxcr4 axis attenuates neonatal hypoxia-induced pulmonary hypertension. Circ. Res. 2009, 104, 1293–1301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, T.; Kawaguchi, N.; Hayama, E.; Furutani, Y.; Nakanishi, T. High expression of cxcr4 and stem cell markers in a monocrotaline and chronic hypoxia-induced rat model of pulmonary arterial hypertension. Exp. Ther. Med. 2018, 15, 4615–4622. [Google Scholar] [CrossRef]
- Zhang, T.; Kawaguchi, N.; Yoshihara, K.; Hayama, E.; Furutani, Y.; Kawaguchi, K.; Tanaka, T.; Nakanishi, T. Silibinin efficacy in a rat model of pulmonary arterial hypertension using monocrotaline and chronic hypoxia. Respir. Res. 2019, 20, 79. [Google Scholar] [CrossRef] [Green Version]
- Surai, P.F. Silymarin as a natural antioxidant: An overview of the current evidence and perspectives. Antioxidants 2015, 4, 204–247. [Google Scholar] [CrossRef] [Green Version]
- Abenavoli, L.; Capasso, R.; Milic, N.; Capasso, F. Milk thistle in liver diseases: Past, present, future. Phytother. Res. PTR 2010, 24, 1423–1432. [Google Scholar] [CrossRef]
- Saller, R.; Meier, R.; Brignoli, R. The use of silymarin in the treatment of liver diseases. Drugs 2001, 61, 2035–2063. [Google Scholar] [CrossRef]
- Federico, A.; Dallio, M.; Loguercio, C. Silymarin/silybin and chronic liver disease: A marriage of many years. Molecules 2017, 22, 191. [Google Scholar] [CrossRef] [Green Version]
- Abenavoli, L.; Izzo, A.A.; Milic, N.; Cicala, C.; Santini, A.; Capasso, R. Milk thistle (silybum marianum): A concise overview on its chemistry, pharmacological, and nutraceutical uses in liver diseases. Phytother. Res. PTR 2018, 32, 2202–2213. [Google Scholar] [CrossRef]
- Singh, R.P.; Agarwal, R. Prostate cancer chemoprevention by silibinin: Bench to bedside. Mol. Carcinog. 2006, 45, 436–442. [Google Scholar] [CrossRef] [PubMed]
- Crocenzi, F.A.; Roma, M.G. Silymarin as a new hepatoprotective agent in experimental cholestasis: New possibilities for an ancient medication. Curr. Med. Chem. 2006, 13, 1055–1074. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, R.; Agarwal, C.; Ichikawa, H.; Singh, R.P.; Aggarwal, B.B. Anticancer potential of silymarin: From bench to bed side. Anticancer Res. 2006, 26, 4457–4498. [Google Scholar] [PubMed]
- Li, M.; Riddle, S.R.; Frid, M.G.; El Kasmi, K.C.; McKinsey, T.A.; Sokol, R.J.; Strassheim, D.; Meyrick, B.; Yeager, M.E.; Flockton, A.R.; et al. Emergence of fibroblasts with a proinflammatory epigenetically altered phenotype in severe hypoxic pulmonary hypertension. J. Immunol. 2011, 187, 2711–2722. [Google Scholar] [CrossRef] [PubMed]
- Hayashida, K.; Fujita, J.; Miyake, Y.; Kawada, H.; Ando, K.; Ogawa, S.; Fukuda, K. Bone marrow-derived cells contribute to pulmonary vascular remodeling in hypoxia-induced pulmonary hypertension. Chest 2005, 127, 1793–1798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDermott, D.H.; Pastrana, D.V.; Calvo, K.R.; Pittaluga, S.; Velez, D.; Cho, E.; Liu, Q.; Trout, H.H., 3rd; Neves, J.F.; Gardner, P.J.; et al. Plerixafor for the treatment of whim syndrome. N. Engl. J. Med. 2019, 380, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Lan, B.; Hayama, E.; Kawaguchi, N.; Furutani, Y.; Nakanishi, T. Therapeutic efficacy of valproic acid in a combined monocrotaline and chronic hypoxia rat model of severe pulmonary hypertension. PLoS ONE 2015, 10, e0117211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morimatsu, Y.; Sakashita, N.; Komohara, Y.; Ohnishi, K.; Masuda, H.; Dahan, D.; Takeya, M.; Guibert, C.; Marthan, R. Development and characterization of an animal model of severe pulmonary arterial hypertension. J. Vasc. Res. 2012, 49, 33–42. [Google Scholar] [CrossRef]
- Schutt, C. Cd14. Int. J. Biochem. Cell Biol. 1999, 31, 545–549. [Google Scholar] [CrossRef]
- De Clercq, E. The amd3100 story: The path to the discovery of a stem cell mobilizer (mozobil). Biochem. Pharmacol. 2009, 77, 1655–1664. [Google Scholar] [CrossRef] [Green Version]
- Kwon, S.G.; Park, I.; Kwon, Y.W.; Lee, T.W.; Park, G.T.; Kim, J.H. Role of stem cell mobilization in the treatment of ischemic diseases. Arch. Pharm. Res. 2019, 42, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Dar, A.; Schajnovitz, A.; Lapid, K.; Kalinkovich, A.; Itkin, T.; Ludin, A.; Kao, W.M.; Battista, M.; Tesio, M.; Kollet, O.; et al. Rapid mobilization of hematopoietic progenitors by amd3100 and catecholamines is mediated by cxcr4-dependent sdf-1 release from bone marrow stromal cells. Leukemia 2011, 25, 1286–1296. [Google Scholar] [CrossRef] [PubMed]
- Marini, O.; Costa, S.; Bevilacqua, D.; Calzetti, F.; Tamassia, N.; Spina, C.; De Sabata, D.; Tinazzi, E.; Lunardi, C.; Scupoli, M.T.; et al. Mature cd10(+) and immature cd10(-) neutrophils present in g-csf-treated donors display opposite effects on t cells. Blood 2017, 129, 1343–1356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waddell, L.A.; Lefevre, L.; Bush, S.J.; Raper, A.; Young, R.; Lisowski, Z.M.; McCulloch, M.E.B.; Muriuki, C.; Sauter, K.A.; Clark, E.L.; et al. Adgre1 (emr1, f4/80) is a rapidly-evolving gene expressed in mammalian monocyte-macrophages. Front. Immunol. 2018, 9, 2246. [Google Scholar] [CrossRef] [Green Version]
- Park, J.W.; Shin, N.R.; Shin, I.S.; Kwon, O.K.; Kim, J.S.; Oh, S.R.; Kim, J.H.; Ahn, K.S. Silibinin inhibits neutrophilic inflammation and mucus secretion induced by cigarette smoke via suppression of erk-sp1 pathway. Phytother. Res. PTR 2016, 30, 1926–1936. [Google Scholar] [CrossRef]
- Matias, M.L.; Gomes, V.J.; Romao-Veiga, M. Silibinin downregulates the nf-kappab pathway and nlrp1/nlrp3 inflammasomes in monocytes from pregnant women with preeclampsia. Molecules 2019, 24, 1548. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Liang, W.C.; Pan, W.L.; Law, W.K.; Hu, J.S.; Ip, D.T.; Waye, M.M.; Ng, T.B.; Wan, D.C. Silibinin, a novel chemokine receptor type 4 antagonist, inhibits chemokine ligand 12-induced migration in breast cancer cells. Phytomedicine 2014, 21, 1310–1317. [Google Scholar] [CrossRef]
- Hatse, S.; Princen, K.; Bridger, G.; De Clercq, E.; Schols, D. Chemokine receptor inhibition by amd3100 is strictly confined to cxcr4. FEBS Lett. 2002, 527, 255–262. [Google Scholar] [CrossRef] [Green Version]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, T.; Kawaguchi, N.; Tsuji, K.; Hayama, E.; Furutani, Y.; Sugiyama, H.; Nakanishi, T. Silibinin Upregulates CXCR4 Expression in Cultured Bone Marrow Cells (BMCs) Especially in Pulmonary Arterial Hypertension Rat Model. Cells 2020, 9, 1276. https://doi.org/10.3390/cells9051276
Zhang T, Kawaguchi N, Tsuji K, Hayama E, Furutani Y, Sugiyama H, Nakanishi T. Silibinin Upregulates CXCR4 Expression in Cultured Bone Marrow Cells (BMCs) Especially in Pulmonary Arterial Hypertension Rat Model. Cells. 2020; 9(5):1276. https://doi.org/10.3390/cells9051276
Chicago/Turabian StyleZhang, Tingting, Nanako Kawaguchi, Kunikazu Tsuji, Emiko Hayama, Yoshiyuki Furutani, Hisashi Sugiyama, and Toshio Nakanishi. 2020. "Silibinin Upregulates CXCR4 Expression in Cultured Bone Marrow Cells (BMCs) Especially in Pulmonary Arterial Hypertension Rat Model" Cells 9, no. 5: 1276. https://doi.org/10.3390/cells9051276