A Systematic Review of MicroRNAs Involved in Cervical Cancer Progression

 , , , ,
, , , ,  and
and
Abstract
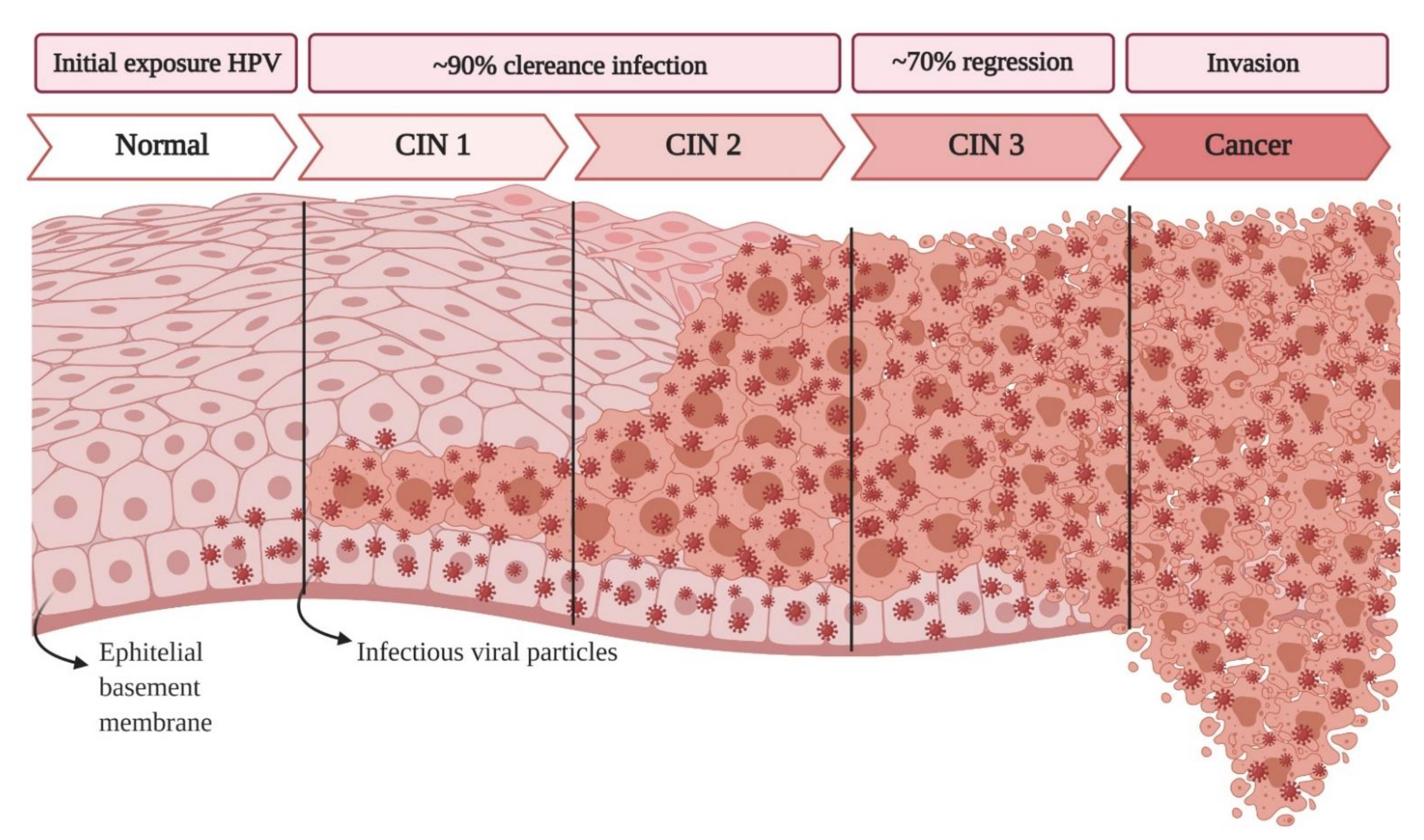
:1. Introduction
2. Materials and Methods
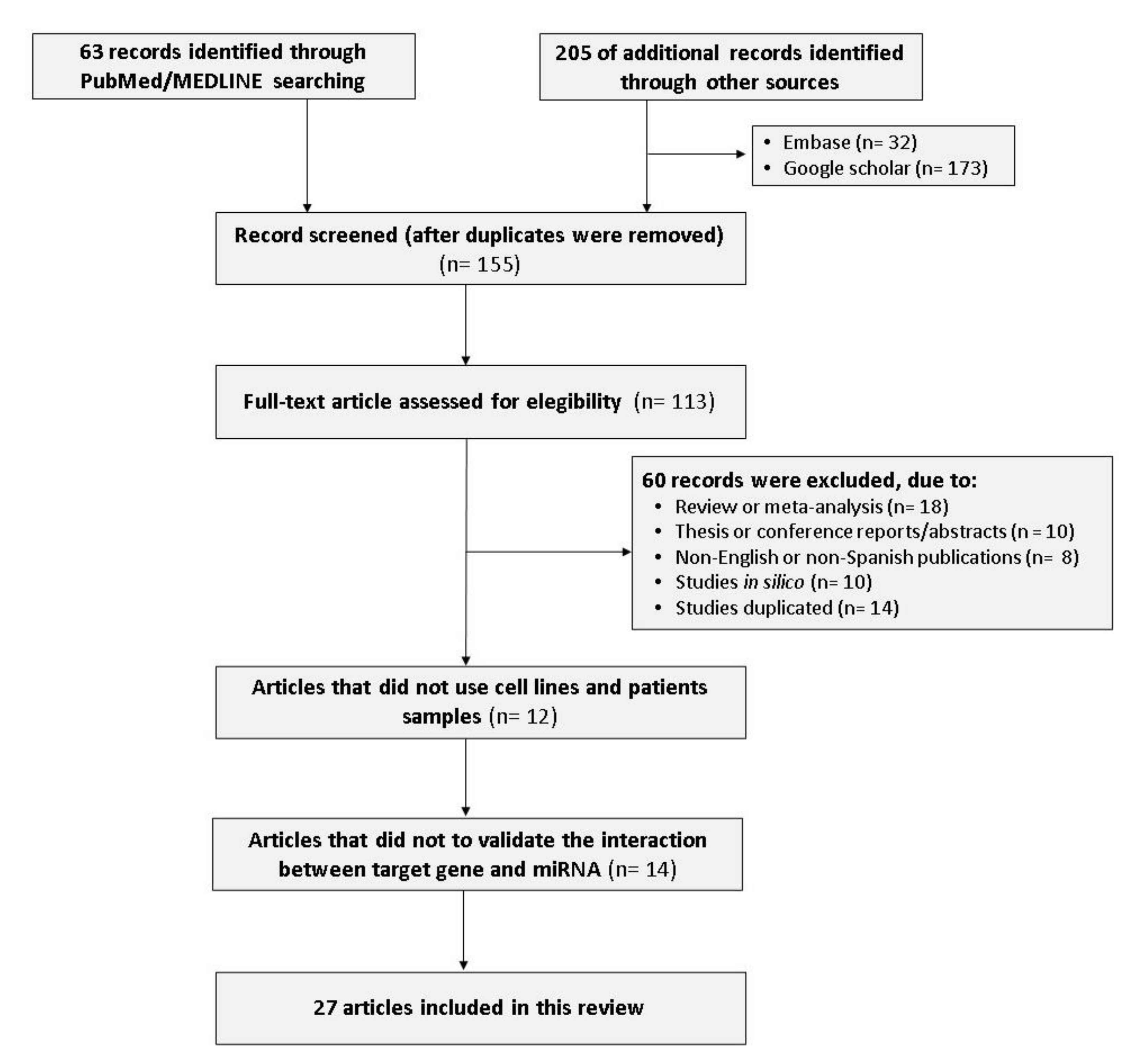
2.1. Search
2.2. Inclusion and Exclusion Criteria
2.3. Data Extraction
3. Results
3.1. Study Population
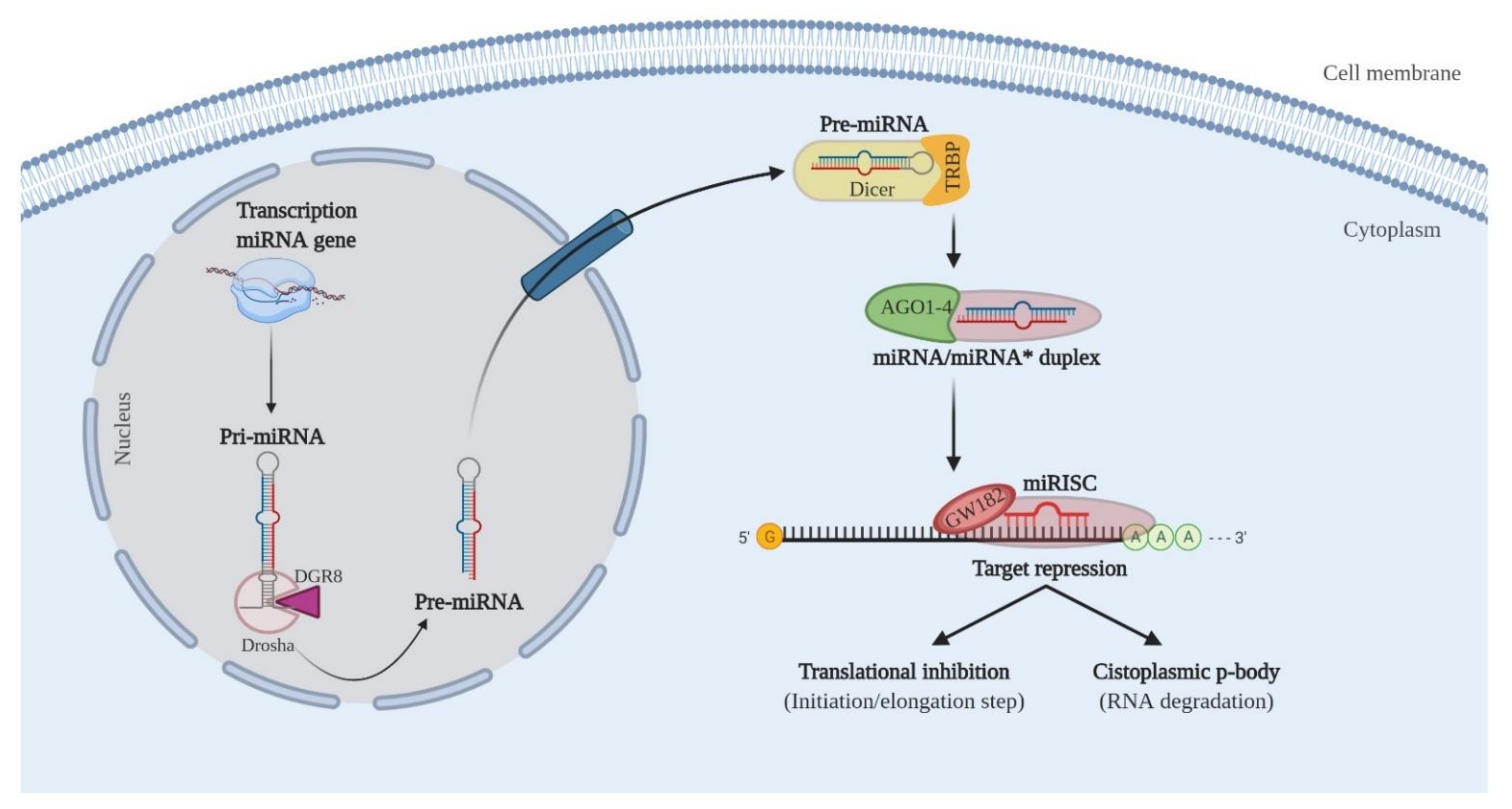
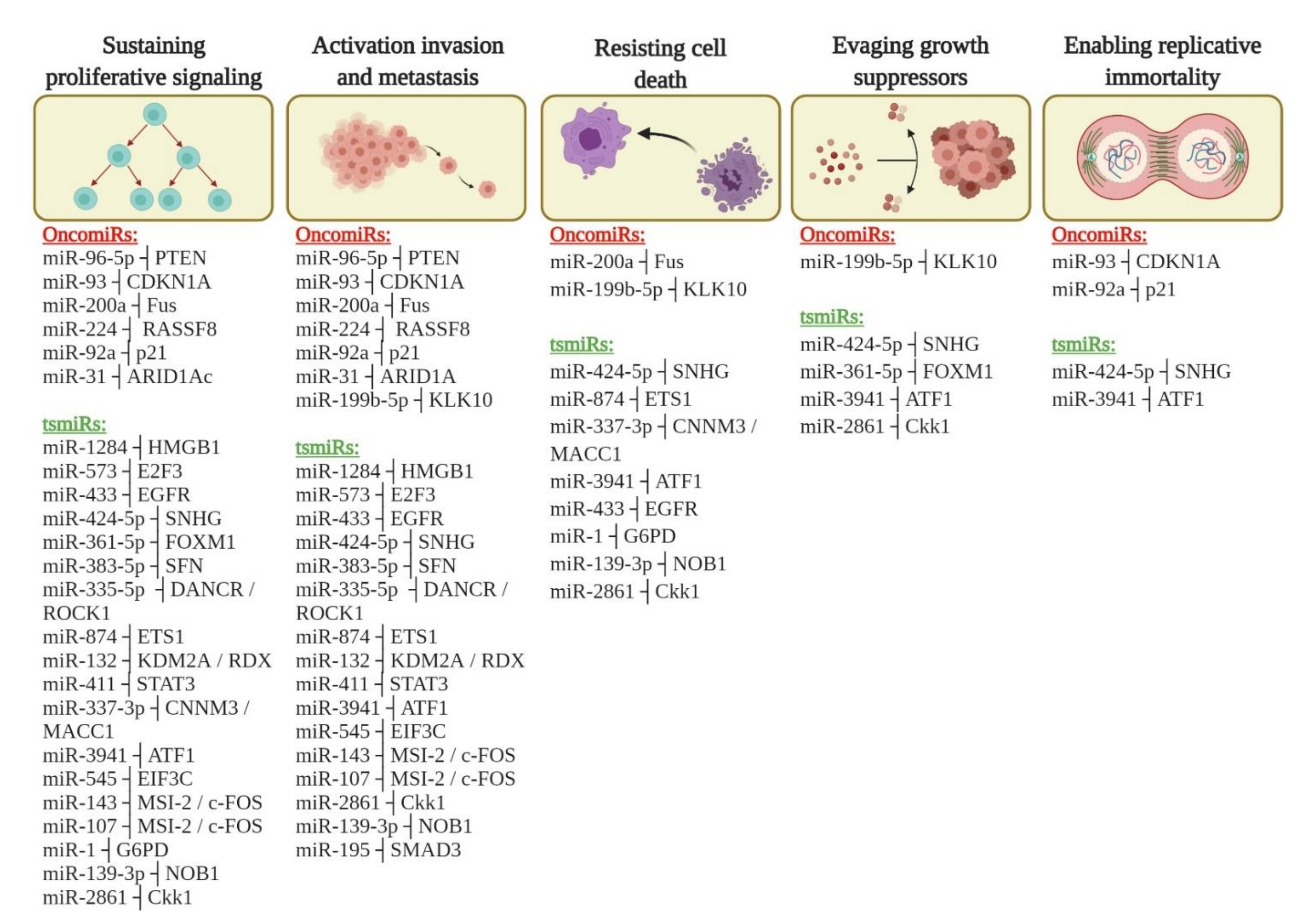
3.2. miRNAs Involved in Cervical Cancer Progression
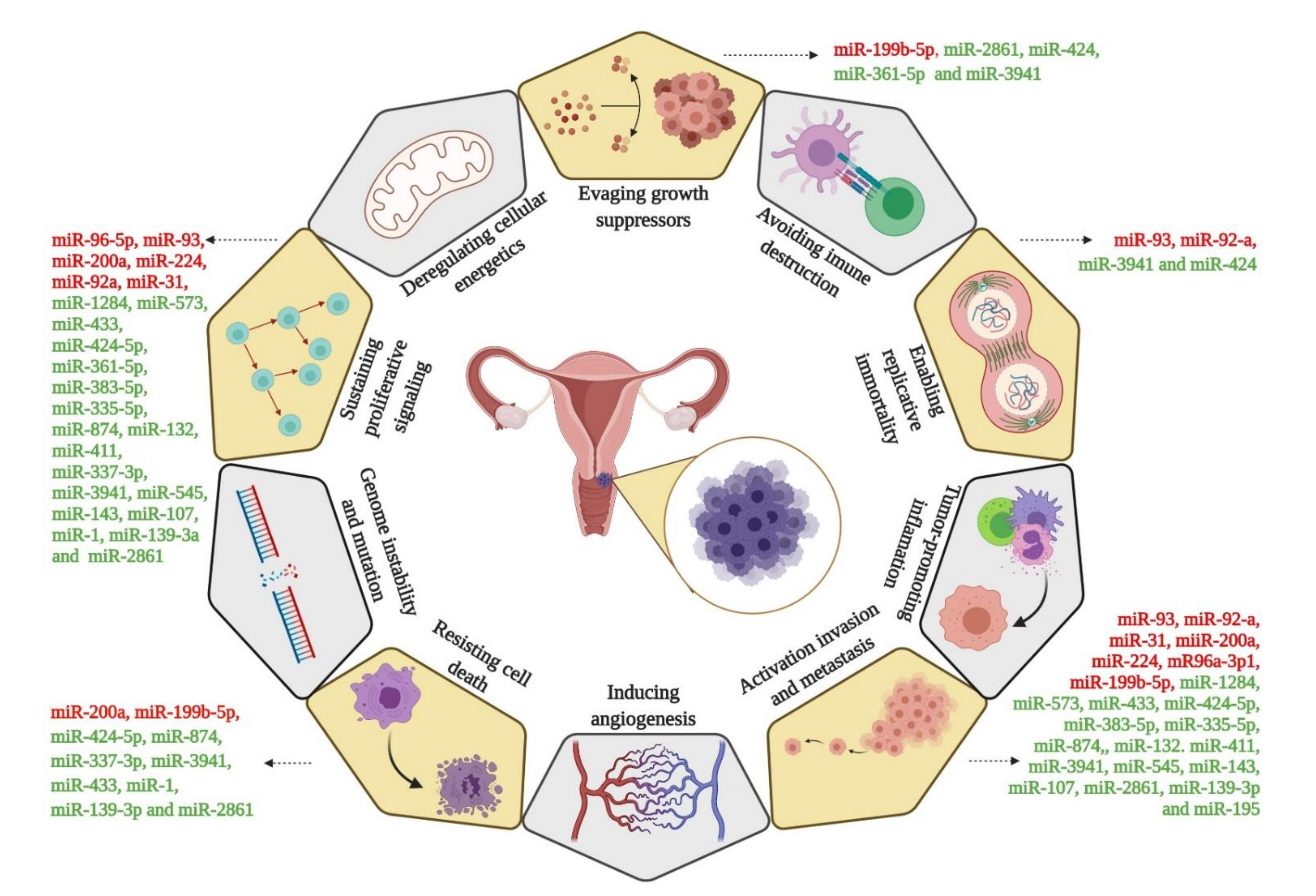
3.3. Targets of miRNAs Involved in the Cervical Cancer Progression
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Crosbie, E.J.; Einstein, M.H.; Franceschi, S.; Kitchener, H.C. Human Papillomavirus and Cervical Cancer. Lancet 2013, 382, 889–899. [Google Scholar] [CrossRef]
- Rosa, M.I.d.; Medeiros, L.R.; Rosa, D.D.; Bozzeti, M.C.; Silva, F.R.; Silva, B.R. Human Papillomavirus and Cervical Neoplasia. Cadernos Saúde Púb. 2009, 25, 953–964. [Google Scholar] [CrossRef] [Green Version]
- Ferlay, J.; Colombet, M.; Soerjomataram, I.; Mathers, C.; Parkin, D.M.; Piñeros, M.; Znaor, A.; Bray, F. Estimating the Global Cancer Incidence and Mortality in 2018: GLOBOCAN Sources and Methods. Int. J. Cancer 2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferlay, J.; Shin, H.-R.; Bray, F.; Forman, D.; Mathers, C.; Parkin, D.M. Estimates of Worldwide Burden of Cancer in 2008: GLOBOCAN 2008. Int. J. Cancer 2010, 127, 2893–2917. [Google Scholar] [CrossRef] [PubMed]
- INCA-Instituto Nacional de Câncer-Estimativa 2018. Available online: http://www1.inca.gov.br/estimativa/2018/ (accessed on 18 November 2019).
- Parkin, D.M.; Bray, F. The Burden of HPV-Related Cancers. Vaccine 2006, 24, S11–S25. [Google Scholar] [CrossRef]
- Sepúlveda, C.; Prado, R. Effective Cervical Cytology Screening Programmes in Middle-Income Countries: The Chilean Experience. Cancer Detection Prevention 2005, 29, 405–411. [Google Scholar] [CrossRef] [PubMed]
- Hull, R.; Mbele, M.; Makhafola, T.; Hicks, C.; Wang, S.-M.; Reis, R.M.; Mehrotra, R.; Mkhize‑Kwitshana, Z.; Kibiki, G.; Bates, D.O.; et al. Cervical Cancer in Low and Middle‑income Countries (Review). Oncol. Letters 2020, 20, 2058–2074. [Google Scholar] [CrossRef]
- Arbyn, M.; Weiderpass, E.; Bruni, L.; de Sanjosé, S.; Saraiya, M.; Ferlay, J.; Bray, F. Estimates of Incidence and Mortality of Cervical Cancer in 2018: A Worldwide Analysis. Lancet Global Health 2020, 8, e191–e203. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.-B.; Wang, J.-H.; Chen, X.-X.; Wu, Y.-Z.; Wu, Q. Values of Three Different Preoperative Regimens in Comprehensive Treatment for Young Patients with Stage Ib2 Cervical Cancer. Asian Pac. J. Cancer Prev. 2012, 13, 1487–1489. [Google Scholar] [CrossRef] [Green Version]
- Burki, T.K. Cervical Cancer: Screening and Risk with Age. Lancet Oncol. 2014, 15, e107. [Google Scholar] [CrossRef]
- Naga Ch, P.; Gurram, L.; Chopra, S.; Mahantshetty, U. The Management of Locally Advanced Cervical Cancer. Curr. Opin. Oncol. 2018. [Google Scholar] [CrossRef]
- Cohen, P.A.; Jhingran, A.; Oaknin, A.; Denny, L. Cervical Cancer. Lancet 2019, 393, 169–182. [Google Scholar] [CrossRef]
- Green, J.A.; Kirwan, J.M.; Tierney, J.F.; Symonds, P.; Fresco, L.; Collingwood, M.; Williams, C.J. Survival and Recurrence after Concomitant Chemotherapy and Radiotherapy for Cancer of the Uterine Cervix: A Systematic Review and Meta-Analysis. Lancet 2001, 358, 781–786. [Google Scholar] [CrossRef]
- De Castro, I. Análise da Expressão de Micrornas em Murganhos Transgénicos K14-HPV16: Implicações na Carcinogénesis. Master’s Thesis, University of Porto, Porto, Portugal, September 2011. [Google Scholar]
- Burd, E.M. Human Papillomavirus and Cervical Cancer. Clin. Microbiol. Rev. 2003, 16, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooper, D.B.; McCathran, C.E. Cervical, Dysplasia. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2017. [Google Scholar]
- Smalley Rumfield, C.; Roller, N.; Pellom, S.T.; Schlom, J.; Jochems, C. Therapeutic Vaccines for HPV-Associated Malignancies. Immunotargets Ther. 2020, 9, 167–200. [Google Scholar] [CrossRef] [PubMed]
- Wentzensen, N.; Vinokurova, S.; von Knebel Doeberitz, M. Systematic Review of Genomic Integration Sites of Human Papillomavirus Genomes in Epithelial Dysplasia and Invasive Cancer of the Female Lower Genital Tract. Cancer Res. 2004, 64, 3878–3884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosa, M.N.; Evangelista, A.F.; Leal, L.F.; De Oliveira, C.M.; Silva, V.A.O.; Munari, C.C.; Munari, F.F.; Matsushita, G.D.M.; Dos Reis, R.; Andrade, C.E.; et al. Establishment, Molecular and Biological Characterization of HCB-514: A Novel Human Cervical Cancer Cell Line. Sci. Rep. 2019, 9, 1913. [Google Scholar] [CrossRef] [Green Version]
- Calin, G.A.; Sevignani, C.; Dumitru, C.D.; Hyslop, T.; Noch, E.; Yendamuri, S.; Shimizu, M.; Rattan, S.; Bullrich, F.; Negrini, M.; et al. Human MicroRNA Genes Are Frequently Located at Fragile Sites and Genomic Regions Involved in Cancers. Proc. Natl. Acad. Sci. USA 2004, 101, 2999–3004. [Google Scholar] [CrossRef] [Green Version]
- He, L.; Hannon, G.J. MicroRNAs: Small RNAs with a Big Role in Gene Regulation. Nat. Rev. Genet. 2004, 5, 522–531. [Google Scholar] [CrossRef]
- Ferracin, M.; Negrini, M. Micromarkers 2.0: An Update on the Role of MicroRNAs in Cancer Diagnosis and Prognosis. Expert Rev. Mol. Diagn. 2015, 15, 1369–1381. [Google Scholar] [CrossRef]
- Iorio, M.V.; Croce, C.M. Causes and Consequences of MicroRNA Dysregulation. Cancer J. 2012, 18, 215–222. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Lin, J.; Ding, Y.; Liu, G.; Luo, Y.; Huang, M.; Xu, C.; Kim, T.K.; Etheridge, A.; Lin, M.; et al. A Systematic Study on Dysregulated MicroRNAs in Cervical Cancer Development. Int. J. Cancer 2016, 138, 1312–1327. [Google Scholar] [CrossRef] [Green Version]
- Almeida, M.I.; Reis, R.M.; Calin, G.A. MicroRNA History: Discovery, Recent Applications, and next Frontiers. Mutat. Res. 2011, 717, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ha, M.; Kim, V.N. Regulation of MicroRNA Biogenesis. Nat. Rev. Mol. Cell Biol. 2014, 15, 509–524. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Gregory, R.I. MicroRNA Biogenesis Pathways in Cancer. Nat. Rev. Cancer 2015, 15, 321–333. [Google Scholar] [CrossRef] [PubMed]
- Pal, A.; Kundu, R. Human Papillomavirus E6 and E7: The Cervical Cancer Hallmarks and Targets for Therapy. Front Microbiol. 2020, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Razavi, Z.S.; Tajiknia, V.; Majidi, S.; Ghandali, M.; Mirzaei, H.R.; Rahimian, N.; Hamblin, M.R.; Mirzaei, H. Gynecologic Cancers and Non-Coding RNAs: Epigenetic Regulators with Emerging Roles. Crit. Rev. Oncol. Hematol. 2021, 157, 103192. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.S.; Lu, J.; Mercer, K.L.; Golub, T.R.; Jacks, T. Impaired MicroRNA Processing Enhances Cellular Transformation and Tumorigenesis. Nat. Genet. 2007, 39, 673–677. [Google Scholar] [CrossRef] [PubMed]
- Grzywa, T.M.; Klicka, K.; Włodarski, P.K. Regulators at Every Step—How MicroRNAs Drive Tumor Cell Invasiveness and Metastasis. Cancers 2020, 12, 3709. [Google Scholar] [CrossRef]
- Svoronos, A.A.; Engelman, D.M.; Slack, F.J. OncomiR or Tumor Suppressor? The Duplicity of MicroRNAs in Cancer. Cancer Res. 2016, 76, 3666–3670. [Google Scholar] [CrossRef] [Green Version]
- Zheng, T.; Wang, J.; Chen, X.; Liu, L. Role of MicroRNA in Anticancer Drug Resistance. Int. J. Cancer 2010, 126, 2–10. [Google Scholar] [CrossRef]
- Parasramka, M.A.; Ho, E.; Williams, D.E.; Dashwood, R.H. MicroRNAs, Diet, and Cancer: New Mechanistic Insights on the Epigenetic Actions of Phytochemicals. Mol. Carcinogenesis 2012, 51, 213–230. [Google Scholar] [CrossRef] [Green Version]
- Kota, S.K.; Balasubramanian, S. Cancer Therapy via Modulation of Micro RNA Levels: A Promising Future. Drug Discovery Today 2010, 15, 733–740. [Google Scholar] [CrossRef]
- Hayes, J.; Peruzzi, P.P.; Lawler, S. MicroRNAs in Cancer: Biomarkers, Functions and Therapy. Trends Mol. Med. 2014, 20, 460–469. [Google Scholar] [CrossRef]
- Liberati, A.; Altman, D.G.; Tetzlaff, J.; Mulrow, C.; Gøtzsche, P.C.; Ioannidis, J.P.A.; Clarke, M.; Devereaux, P.J.; Kleijnen, J.; Moher, D. The PRISMA Statement for Reporting Systematic Reviews and Meta-Analyses of Studies That Evaluate Health Care Interventions: Explanation and Elaboration. PLoS Med. 2009, 6, e1000100. [Google Scholar] [CrossRef]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G.; Group, T.P. Preferred Reporting Items for Systematic Reviews and Meta-Analyses: The PRISMA Statement. PLoS Med. 2009, 6, e1000097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Methley, A.M.; Campbell, S.; Chew-Graham, C.; McNally, R.; Cheraghi-Sohi, S. PICO, PICOS and SPIDER: A Comparison Study of Specificity and Sensitivity in Three Search Tools for Qualitative Systematic Reviews. BMC Health Serv. Res. 2014, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Li, G. MiR-1284 Enhances Sensitivity of Cervical Cancer Cells to Cisplatin via Downregulating HMGB1. Biomed. Pharmacother. 2018, 107, 997–1003. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Ma, C.; Zhang, W.; Chen, Z.; Ma, L. Down Regulation of MiR-143 Is Related with Tumor Size, Lymph Node Metastasis and HPV16 Infection in Cervical Squamous Cancer. Diagn. Pathol. 2014, 9, 88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, L.; Zhang, H. Circ-ATP8A2 Promotes Cell Proliferation and Invasion as a CeRNA to Target EGFR by Sponging MiR-433 in Cervical Cancer. Gene 2019, 705, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.; Wang, Q.; Li, L.; Xiao-Jin, Z. Upregulation of Long Non-Coding RNA Small Nucleolar RNA Host Gene 12 Contributes to Cell Growth and Invasion in Cervical Cancer by Acting as a Sponge for MiR-424-5p. Cell. Physiol. Biochem. 2018, 45, 2086–2094. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Feng, J.; Yao, H.; Li, Y.; Xi, J.; Yang, J. LncRNA SBF2-AS1 Promotes the Progression of Cervical Cancer by Regulating MiR-361-5p/FOXM1 Axis. Artif Cells Nanomed. Biotechnol. 2019, 47, 776–782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Y.; Ma, Y.; Liu, J.; Cai, Y.; Zhang, M.; Fang, X. LINC01128 Expedites Cervical Cancer Progression by Regulating MiR-383-5p/SFN Axis. BMC Cancer 2019, 19. [Google Scholar] [CrossRef]
- Liang, H.; Zhang, C.; Guan, H.; Liu, J.; Cui, Y. LncRNA DANCR Promotes Cervical Cancer Progression by Upregulating ROCK1 via Sponging MiR-335-5p. J. Cell. Physiol. 2019, 234, 7266–7278. [Google Scholar] [CrossRef]
- Liao, H.; Pan, Y.; Pan, Y.; Shen, J.; Qi, Q.; Zhong, L.; Han, W.; Wang, Q.; Jiang, Y. MicroRNA‑874 Is Downregulated in Cervical Cancer and Inhibits Cancer Progression by Directly Targeting ETS1. Oncol. Rep. 2018, 40, 2389–2398. [Google Scholar] [CrossRef]
- Ou, R.; Zhu, L.; Zhao, L.; Li, W.; Tao, F.; Lu, Y.; He, Q.; Li, J.; Ren, Y.; Xu, Y. HPV16 E7-Induced Upregulation of KDM2A Promotes Cervical Cancer Progression by Regulating MiR-132-Radixin Pathway. J. Cell. Physiol. 2019, 234, 2659–2671. [Google Scholar] [CrossRef]
- Shan, D.; Shang, Y.; Hu, T. MicroRNA-411 Inhibits Cervical Cancer Progression by Directly Targeting STAT3. Oncol. Res. 2019, 27, 349–358. [Google Scholar] [CrossRef]
- Shao, S.; Wang, C.; Wang, S.; Zhang, H.; Zhang, Y. LncRNA STXBP5-AS1 Suppressed Cervical Cancer Progression via Targeting MiR-96-5p/PTEN Axis. Biomed. Pharmacother. 2019, 117, 109082. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, L.; Wang, W.; Guo, X. Overexpression of Circular RNA Hsa_circ_0001038 Promotes Cervical Cancer Cell Progression by Acting as a CeRNA for MiR-337-3p to Regulate Cyclin-M3 and Metastasis-Associated in Colon Cancer 1 Expression. Gene 2020, 733, 144273. [Google Scholar] [CrossRef]
- Xu, L.-J.; Duan, Y.; Wang, P.; Yin, H.-Q. MiR-199b-5p Promotes Tumor Growth and Metastasis in Cervical Cancer by down-Regulating KLK10. Biochem. Biophys. Res. Commun. 2018, 503, 556–563. [Google Scholar] [CrossRef]
- Xu, Y.; Zhou, W.; Zhang, C.; Liu, X.; Lv, J.; Li, X.; Zhao, L.; Li, W.; Li, J.; Ren, Y.; et al. Long Non-Coding RNA RP11-552M11.4 Favors Tumorigenesis and Development of Cervical Cancer via Modulating MiR-3941/ATF1 Signaling. Int. J. Biol. Macromol. 2019, 130, 24–33. [Google Scholar] [CrossRef]
- Zhang, X.; Li, F.; Zhu, L. Clinical Significance and Functions of MicroRNA-93/CDKN1A Axis in Human Cervical Cancer. Life Sci. 2018, 209, 242–248. [Google Scholar] [CrossRef]
- Hu, C.; Wang, Y.; Li, A.; Zhang, J.; Xue, F.; Zhu, L. Overexpressed Circ_0067934 Acts as an Oncogene to Facilitate Cervical Cancer Progression via the MiR-545/EIF3C Axis. J. Cell. Physiol. 2019, 234, 9225–9232. [Google Scholar] [CrossRef]
- Zhu, H.; Zheng, T.; Yu, J.; Zhou, L.; Wang, L. LncRNA XIST Accelerates Cervical Cancer Progression via Upregulating Fus through Competitively Binding with MiR-200a. Biomed. Pharmacother. 2018, 105, 789–797. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.; Ding, J.; Yang, Y.; Deng, L.; Li, X. MicroRNA-433 Inhibits Cervical Cancer Progression by Directly Targeting Metadherin to Regulate the AKT and β-Catenin Signalling Pathways. Oncol. Rep. 2017, 38, 3639–3649. [Google Scholar] [CrossRef]
- Dong, P.; Xiong, Y.; Hanley, S.J.B.; Yue, J.; Watari, H. Musashi-2, a Novel Oncoprotein Promoting Cervical Cancer Cell Growth and Invasion, Is Negatively Regulated by P53-Induced MiR-143 and MiR-107 Activation. J. Exp. Clin. Cancer Res. 2017, 36, 150. [Google Scholar] [CrossRef] [PubMed]
- Hu, T.; Chang, Y.-F.; Xiao, Z.; Mao, R.; Tong, J.; Chen, B.; Liu, G.-C.; Hong, Y.; Chen, H.-L.; Kong, S.-Y.; et al. MiR-1 Inhibits Progression of High-Risk Papillomavirus-Associated Human Cervical Cancer by Targeting G6PD. Oncotarget 2016, 7, 86103–86116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, P.; Xi, J.; Liu, S. MiR-139-3p Induces Cell Apoptosis and Inhibits Metastasis of Cervical Cancer by Targeting NOB1. Biomed. Pharmacother. 2016, 83, 850–856. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Li, Y.; Wang, F.F.; Lv, W.; Xie, X.; Cheng, X. Over-Expressed MiR-224 Promotes the Progression of Cervical Cancer via Targeting RASSF8. PLoS ONE 2016, 11, e0162378. [Google Scholar] [CrossRef]
- Su, Z.; Yang, H.; Zhao, M.; Wang, Y.; Deng, G.; Chen, R. MicroRNA-92a Promotes Cell Proliferation in Cervical Cancer via Inhibiting P21 Expression and Promoting Cell Cycle Progression. Oncol. Res. 2017, 25, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Han, L.R.; Zhou, Y.X.; Li, Y. MiR-195 Suppresses Cervical Cancer Migration and Invasion Through Targeting Smad3. Int. J. Gynecol. Cancer 2016, 26, 817–824. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Zhou, Y.; Zheng, L.; Li, H. MiR-31 Is an Independent Prognostic Factor and Functions as an Oncomir in Cervical Cancer via Targeting ARID1A. Gynecol. Oncol. 2014, 134, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Li, Y.; Wang, F.; Wang, X.; Cheng, B.; Ye, F.; Xie, X.; Zhou, C.; Lu, W. Suppressed MiR-424 Expression via Upregulation of Target Gene Chk1 Contributes to the Progression of Cervical Cancer. Oncogene 2013, 32, 976–987. [Google Scholar] [CrossRef]
- Xu, J.; Wan, X.; Chen, X.; Fang, Y.; Cheng, X.; Xie, X.; Lu, W. MiR-2861 Acts as a Tumor Suppressor via Targeting EGFR/AKT2/CCND1 Pathway in Cervical Cancer Induced by Human Papillomavirus Virus 16 E6. Sci. Rep. 2016, 6, 28968. [Google Scholar] [CrossRef] [Green Version]
- Hou, X.W.; Sun, X.; Yu, Y.; Zhao, H.M.; Yang, Z.J.; Wang, X.; Cao, X.C. MiR-361-5p Suppresses Lung Cancer Cell Lines Progression by Targeting FOXM1. Neoplasma 2017, 64, 526–534. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Zhao, Z.; Xie, L.; Zhu, J. MiR-361-5p Suppresses Chemoresistance of Gastric Cancer Cells by Targeting FOXM1 via the PI3K/Akt/MTOR Pathway. Oncotarget 2018, 9, 4886–4896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Xu, Z. MiR-93 Enhances Cell Proliferation by Directly Targeting CDKN1A in Nasopharyngeal Carcinoma. Oncol. Letters 2018, 15, 1723–1727. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Yu, B. MicroRNA‑93 Promotes Cell Proliferation by Directly Targeting P21 in Osteosarcoma Cells. Exp. Ther. Med. 2017, 13, 2003–2011. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Lyu, J.; Meng, Q.H. MiR-93 Promotes Tumorigenesis and Metastasis of Non-Small Cell Lung Cancer Cells by Activating the PI3K/Akt Pathway via Inhibition of LKB1/PTEN/CDKN1A. J. Cancer 2017, 8, 870–879. [Google Scholar] [CrossRef] [Green Version]
- Ohta, K.; Hoshino, H.; Wang, J.; Ono, S.; Iida, Y.; Hata, K.; Huang, S.K.; Colquhoun, S.; Hoon, D.S.B. MicroRNA-93 Activates c-Met/PI3K/Akt Pathway Activity in Hepatocellular Carcinoma by Directly Inhibiting PTEN and CDKN1A. Oncotarget 2015, 6, 3211–3224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.-J.; Cao, Y.-F.; Yang, Z.-Q.; Jiang, Z.-Y.; Cai, B.; Guo, J.; Zhang, S.; Zhang, X.-L.; Gao, F. MicroRNA-93-5p Increases Multidrug Resistance in Human Colorectal Carcinoma Cells by Downregulating Cyclin Dependent Kinase Inhibitor 1A Gene Expression. Oncol. Lett. 2017, 13, 722–730. [Google Scholar] [CrossRef] [PubMed]
- Su, X.; Wang, H.; Ge, W.; Yang, M.; Hou, J.; Chen, T.; Li, N.; Cao, X. An In Vivo Method to Identify MicroRNA Targets Not Predicted by Computation Algorithms: P21 Targeting by MiR-92a in Cancer. Cancer Res. 2015, 75, 2875–2885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanahan, D.; Weinberg, R.A. Hallmarks of Cancer: The next Generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Creasman, W.T.; Odicino, F.; Maisonneuve, P.; Quinn, M.A.; Beller, U.; Benedet, J.L.; Heintz, A.P.M.; Ngan, H.Y.S.; Pecorelli, S. Carcinoma of the Corpus Uteri. FIGO 26th Annual Report on the Results of Treatment in Gynecological Cancer. Int. J. Gynaecol. Obstet 2006, 95 Suppl 1, S105–S143. [Google Scholar] [CrossRef]
- Kong, Y.W.; Ferland-McCollough, D.; Jackson, T.J.; Bushell, M. MicroRNAs in Cancer Management. Lancet Oncol. 2012, 13, e249–e258. [Google Scholar] [CrossRef]
- González-Quintana, V.; Palma-Berré, L.; Campos-Parra, A.D.; López-Urrutia, E.; Peralta-Zaragoza, O.; Vazquez-Romo, R.; Pérez-Plasencia, C. MicroRNAs Are Involved in Cervical Cancer Development, Progression, Clinical Outcome and Improvement Treatment Response (Review). Oncol. Rep. 2016, 35, 3–12. [Google Scholar] [CrossRef]
- Chen, P.; Wang, R.; Yue, Q.; Hao, M. Long Non-Coding RNA TTN-AS1 Promotes Cell Growth and Metastasis in Cervical Cancer via MiR-573/E2F3. Biochem. Biophys. Res. Commun. 2018, 503, 2956–2962. [Google Scholar] [CrossRef]
- Jin, X.-J.; Chen, X.-J.; Zhang, Z.-F.; Hu, W.-S.; Ou, R.-Y.; Li, S.; Xue, J.-S.; Chen, L.-L.; Hu, Y.; Zhu, H. Long Noncoding RNA SNHG12 Promotes the Progression of Cervical Cancer via Modulating MiR-125b/STAT3 Axis. J. Cell. Physiol. 2019, 234, 6624–6632. [Google Scholar] [CrossRef]
- Grau, J.F.; Farinas-Madrid, L.; Oaknin, A. A Randomized Phase III Trial of Platinum Chemotherapy plus Paclitaxel with Bevacizumab and Atezolizumab versus Platinum Chemotherapy plus Paclitaxel and Bevacizumab in Metastatic (Stage IVB), Persistent, or Recurrent Carcinoma of the Cervix: The BEATcc Study (ENGOT-Cx10/GEICO 68-C/JGOG1084/GOG-3030). Int. J. Gynecol. Cancer 2020, 30, 139–143. [Google Scholar] [CrossRef] [PubMed]
- Forterre, A.; Komuro, H.; Aminova, S.; Harada, M. A Comprehensive Review of Cancer MicroRNA Therapeutic Delivery Strategies. Cancers 2020, 12, 1852. [Google Scholar] [CrossRef] [PubMed]
- Hanna, J.; Hossain, G.S.; Kocerha, J. The Potential for MicroRNA Therapeutics and Clinical Research. Front Genet. 2019, 10, 478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Condrat, C.E.; Thompson, D.C.; Barbu, M.G.; Bugnar, O.L.; Boboc, A.; Cretoiu, D.; Suciu, N.; Cretoiu, S.M.; Voinea, S.C. MiRNAs as Biomarkers in Disease: Latest Findings Regarding Their Role in Diagnosis and Prognosis. Cells 2020, 9, 276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, D.-D.; Lv, J.; Wei, K.-L.; Feng, Z.-B.; Chen, J.-T.; Liu, K.-C.; Chen, G.; Luo, D.-Z. A Nine-MiRNA Signature as a Potential Diagnostic Marker for Breast Carcinoma: An Integrated Study of 1,110 Cases. Oncol. Rep. 2017, 37, 3297–3304. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Domain | Inclusion Criteria |
|---|---|
| Patients (P) | Patients with cervical cancer or cervical intraepithelial neoplasia grade 2/3 |
| Interventions (I) | Differentially expressed miRNAs |
| Comparators (C) | Non-neoplastic control tissue or cervical cancer FIGO stage I |
| Outcomes (O) | Differentially expressed miRNAs indicate a statistically significant difference in the overall survival of patients |
| Author, Year | Country | Cell Lines | Sample | N | Non-Neoplastic Control Tissue (n) | Cervical Cancer Samples FIGO Stage (n) |
|---|---|---|---|---|---|---|
| Chen and Li, 2018 [41] | China | MS751, HeLa, SiHa e C33 A and ECS | Tissue | 82 | NAT (82) | I (28) II (54) |
| Chen et al., 2018 [42] | China | SiHa, HeLa, C-33A, Me180, Ms751 and Ect1/E6E7 | Frozen Tissue | 45 | NAT (45) | I-II (26) III-IV (19) |
| Ding and Zhang, 2019 [43] | China | HeLa, SiHa, C-33A, SW75 and HcerEpic | FrozenTissue | 46 | NAT (46) | I (31) II (15) |
| Dong et al., 2018 [44] | China | C-33A, ME-180, CaSki, HeLa, SiHa and NC104 | Tissue | 81 | NAT (81) | I (46) II (35) |
| Gao et al., 2019 [45] | China | SiHa, HeLa, C-33A, Me180 and Ms751 and Ect1/E6E7 | Frozen Tissue | 66 | NAT (66) | I-II (29) III-IV (37) |
| Hu et al., 2019 [46] | China | HeLa, SiHa, C-33A and Ect1-E6E7 | Frozen Tissue | 33 | NAT (33) | I-II (13) III-IV (20) |
| Liang et al., 2019 [47] | China | CaSki, SW756, SiHa, C-33A, HeLa, ME-180 and Ect1-E6E7 | Frozen Tissue | 65 | NAT (65) | I-II (37) III-IV (28) |
| Liao et al., 2018 [48] | China | SiHa, HeLa, C-33A, CaSki and Ect1/E6E7 | Frozen Tissue | 49 | NAT (49) | I-II (23) III-IV (26) |
| Ou et al., 2019 [49] | China | C-33A, SiHa, and CaSk | Tissue | 81 | NAT (81) | Ib-IIa (37) IIb-IIIa (44) |
| Shan et al., 2019 [50] | China | HeLa, SiHa, CaSki, C-33A and Ect1/E6E7 | Frozen Tissue | 45 | NAT (45) | I-II (17) III-IV (28) |
| Shao et al., 2019 [51] | China | C-33A, MS751, SiHa, HeLa, ME-180, CaSki and NC104 | Tissue | 37 | NAT (37) | I-II (19) III-IV (18) |
| Wang et al., 2020 [52] | China | HeLa, SiHa, C-33A, SW756 and HcerEpic | Tissue | 55 | NAT (55) | I (31) II (24) |
| Xu et al., 2018 [53] | China | HeLa, CaSki, SiHa, ME-180, MS-751, C-33 A, Ect1/E6E7 and HcerEpic | Frozen Tissue | 70 | NAT (70) | I-II (36) III-IV (34) |
| Xu et al., 2019 [54] | China | C-33A, SiHa, ME-180, HeLa, CaSki and NC104 | Tissue | 92 | NAT (92) | Ib-IIa (46) IIb-IIIa (46) |
| Zhang et al., 2018 [55] | China | HeLa | Tissue | 100 | NAT (100) | I (56) II (32) III (8) IV (4) |
| Hu et al., 2019 [56] | China | SiHa, CaSki, HeLa, C4-1, and NC104 | Frozen Tissue | 21 | NAT (21) | I (34) II (27) |
| Zhu et al., 2018 [57] | China | HeLa, CaSki, C4-1, SiHa, and GH329 | Tissue | 52 | NAT (52) | I-II (30) III-IV (23) |
| Liang et al., 2017 [58] | China | HeLa, C-33A, SiHa, CaSki, and Hct1/E6E7 | Frozen Tissue | 65 | NAT (65) | I-II (24) II-IV (41) |
| Dong et al., 2017 [59] | Japan | HeLa, SiHa and H8 | Frozen Tissue | 58 | NAT (58) | I-II (38) III-IV (20) |
| Hu et al., 2016 [60] | China | HeLa, SiHa, C-33A and H8 | Frozen Tissue | 57 | NAT (57) | I (28) II (14) III (9) IV (3) |
| Huang et al., 2016 [61] | China | HeLa, SiHa, CaSki, C-33A, and HaCaT | Frozen Tissue | 40 | NAT (40) | I-II (40) |
| Huang et al, 2016 [62] | China | SiHa and CaSki | Frozen Tissue | 190 | HT (64) | I (86) II (40) |
| Su et al., 2017 [63] | China | HeLa | Frozen Tissue | 74 | I-II (39) III-IV (35) | |
| Zhou et al., 2016 [64] | China | HeLa, SiHa, CaSki, ME-180, C-33A and HaCaT | Tissue | 50 | IB (27) IC-IV (23) | |
| Wang et al., 2014 [65] | China | HeLa, SiHa, CaSki, ME-180, C-33A and HaCaT | Tissue | 27 | I (20) II (7) | |
| Xu et al., 2013 [66] | China | SiHa and CaSki | Frozen Tissue | 147 | I (108) II (39) | |
| Xu et al., 2016 [67] | China | SiHa, CaSki, HEK293T and HaCaT | Frozen Tissue | 57 | NAT (57) | I (35) II (22) |
| miRNA | Target Interation | Target Prediction Tool | Target Validated | REF |
|---|---|---|---|---|
| miR-1284 | HMGB1 | miRanda | Luciferase | [41] |
| miR-573 | E2F3 | Targetscan | Luciferase | [42] |
| miR-433 | circ-ATP8A2/EGFR | Targetscan | Luciferase | [43,58] |
| miR-424-5p | SNHG12 | StarBase v2.0 | Luciferase | [44,66] |
| miR-361-5p | SBF2-AS/FOXM1 | Previous studies [68,69] | Luciferase | [45] |
| miR-383-5p | LINC01128/SFN | StarBase v2.0 | Luciferase | [46] |
| miR-335-5p | DANCR/ROCK1 | DIANA-LncBase V.2; Targetscan Human 7.2 | Luciferase | [47] |
| miR-874 | ETS1 | TargetScan7.1 and microRNA.org | Luciferase | |
| miR-132 | KDM2A/RDX | PicTar algorithm | Luciferase | [49] |
| miR-411 | STAT3 | TargetScan and miRanda | Luciferase | [50] |
| miR-96-5p | STXBP5-AS1/PTEN | TargetScan and miRanda | Luciferase | [51] |
| miR-337-3p | hsa_circ_0001038, CNNM3/MACC1 | Circular RNA Interactome and TargetScan | Luciferase | [52] |
| miR-199b-5p | KLK10 | TargetScan and miRanda | Luciferase | [53] |
| miR-3941 | lncRNA RP11-552M11.4/ATF1 | DIANA tool LncBase v.2 and TargetScan | Luciferase | [54] |
| miR-93 | CDKN1A | Previous studies [70,71,72,73,74] | Luciferase [70,71,72,73] | [55] |
| miR-545 | circ_0067934/EIF3C | Circular RNA Interactome and TargetScan | Luciferase | [56] |
| miR-200a | XIST/Fus | Starbase | Luciferase | [57] |
| miR-143 | MSI-2/c-FOS | RNA-IP | Luciferase | [59] |
| miR-107 | MSI-2/c-FOS | RNA-IP | Luciferase | [59] |
| miR-1 | G6PD | RNA-IP | Luciferase | [60] |
| miR-139-3p | NOB1 | TargetScan, miRanda, and Diana microT computational algorithms | RIP-Chip and Luciferase | [61] |
| miR-224 | RASSF8 | TargetScan Human 7.0 | Luciferase | [62] |
| miR-92a | p21 | TargetScan, PicTar and microbase | Luciferase | [63] |
| miR-195 | SMAD3 | Previous study [75] | Luciferase [75] | [64] |
| miR-31 | ARID1A | TargetScan, pictar, and miRanda | Luciferase | [65] |
| miR-2861 | EGFR/ AKT2/CCND1 | TargetScan, pictar, miRanda and Microcosm Targets | Luciferase | [67] |
| miRNA | Status | Methodology | Outcome | REF |
|---|---|---|---|---|
| miR-1284 | D | RT-qPCR | Poor prognosis | [41] |
| miR-573 | D | RT-qPCR | Poor prognosis | [42] |
| miR-433 | D | RT-qPCR | Poor prognosis | [43,58] |
| miR-424-5p | D | RT-qPCR | Poor prognosis | [44,66] |
| miR-361-5p | D | RT-qPCR | Poor prognosis | [45] |
| miR-383-5p | D | RT-qPCR | Poor prognosis | [46] |
| miR-335-5p | D | RT-qPCR | Poor prognosis | [47] |
| miR-874 | D | RT-qPCR | Poor prognosis | |
| miR-132 | D | RT-qPCR | Poor prognosis | [49] |
| miR-411 | D | RT-qPCR | Poor prognosis | [50] |
| miR-96-5p | U | RT-qPCR | Poor prognosis | [51] |
| miR-337-3p | D | RT-qPCR | Poor prognosis | [52] |
| miR-199b-5p | U | RT-qPCR | Poor prognosis | [53] |
| miR-3941 | D | RT-qPCR | Poor prognosis | [54] |
| miR-93 | U | RT-qPCR | Poor prognosis | [55] |
| miR-545 | D | RT-qPCR | Poor prognosis | [56] |
| miR-200a | U | RT-qPCR | Poor prognosis | [57] |
| miR-143 | D | RT-qPCR | Poor prognosis | [59] |
| miR-107 | D | RT-qPCR | Poor prognosis | [59] |
| miR-1 | D | RT-qPCR | Poor prognosis | [60] |
| miR-139-3p | D | RT-qPCR | Poor prognosis | [61] |
| miR-224 | U | RT-qPCR | Poor prognosis | [62] |
| miR-92a | U | RT-qPCR | Poor prognosis | [63] |
| miR-195 | D | RT-qPCR | Poor prognosis | [64] |
| miR-31 | U | RT-qPCR | Poor prognosis | [65] |
| miR-2861 | D | Microarray and RT-qPCR | Poor prognosis | [67] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Causin, R.L.; Freitas, A.J.A.d.; Trovo Hidalgo Filho, C.M.; Reis, R.d.; Reis, R.M.; Marques, M.M.C. A Systematic Review of MicroRNAs Involved in Cervical Cancer Progression. Cells 2021, 10, 668. https://doi.org/10.3390/cells10030668
Causin RL, Freitas AJAd, Trovo Hidalgo Filho CM, Reis Rd, Reis RM, Marques MMC. A Systematic Review of MicroRNAs Involved in Cervical Cancer Progression. Cells. 2021; 10(3):668. https://doi.org/10.3390/cells10030668
Chicago/Turabian StyleCausin, Rhafaela Lima, Ana Julia Aguiar de Freitas, Cassio Murilo Trovo Hidalgo Filho, Ricardo dos Reis, Rui Manuel Reis, and Márcia Maria Chiquitelli Marques. 2021. "A Systematic Review of MicroRNAs Involved in Cervical Cancer Progression" Cells 10, no. 3: 668. https://doi.org/10.3390/cells10030668