
YAP1 Is a Potential Predictive Molecular Biomarker for Response to SMO Inhibitor in Medulloblastoma Cells

 , , , and
, , , and 
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
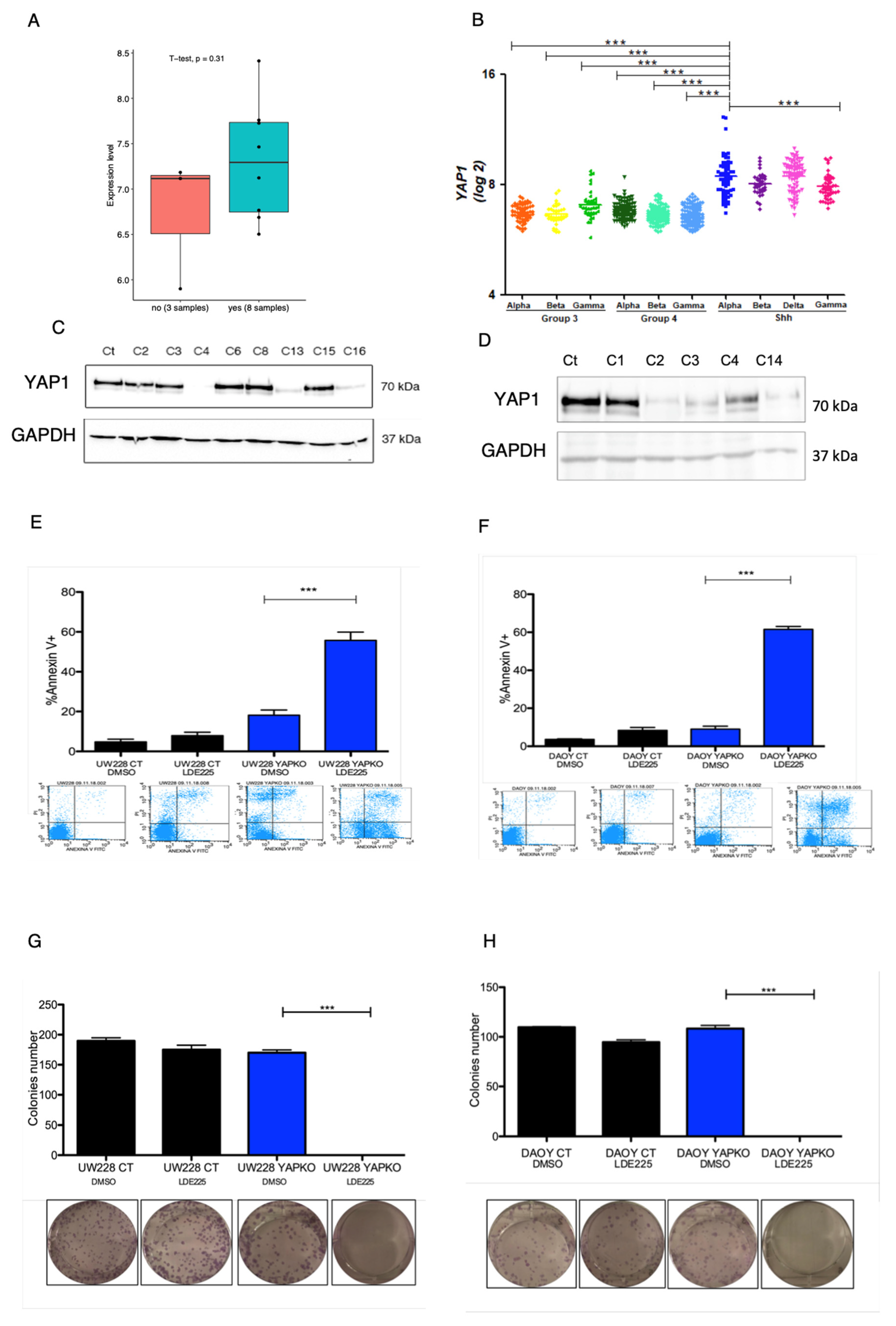
The Context of YAP1 in SHH Medulloblastoma
2. Methods
2.1. Cell Culture
2.2. Generation of CRISPR/Cas9 Single-Clones YAP1 Depleted Cells
2.3. Immunoblotting
2.4. CCK8 Proliferation Assay and Doubling Time
2.5. Retrospective Analysis of Microarray Data
2.6. Colony Formation Assay
2.7. Annexin V/Propidium Iodide Staining to Detect Apoptotic and Necrotic Cells
2.8. Statistical Analysis
2.9. Single-Cell RNA-Seq Analysis
3. Results
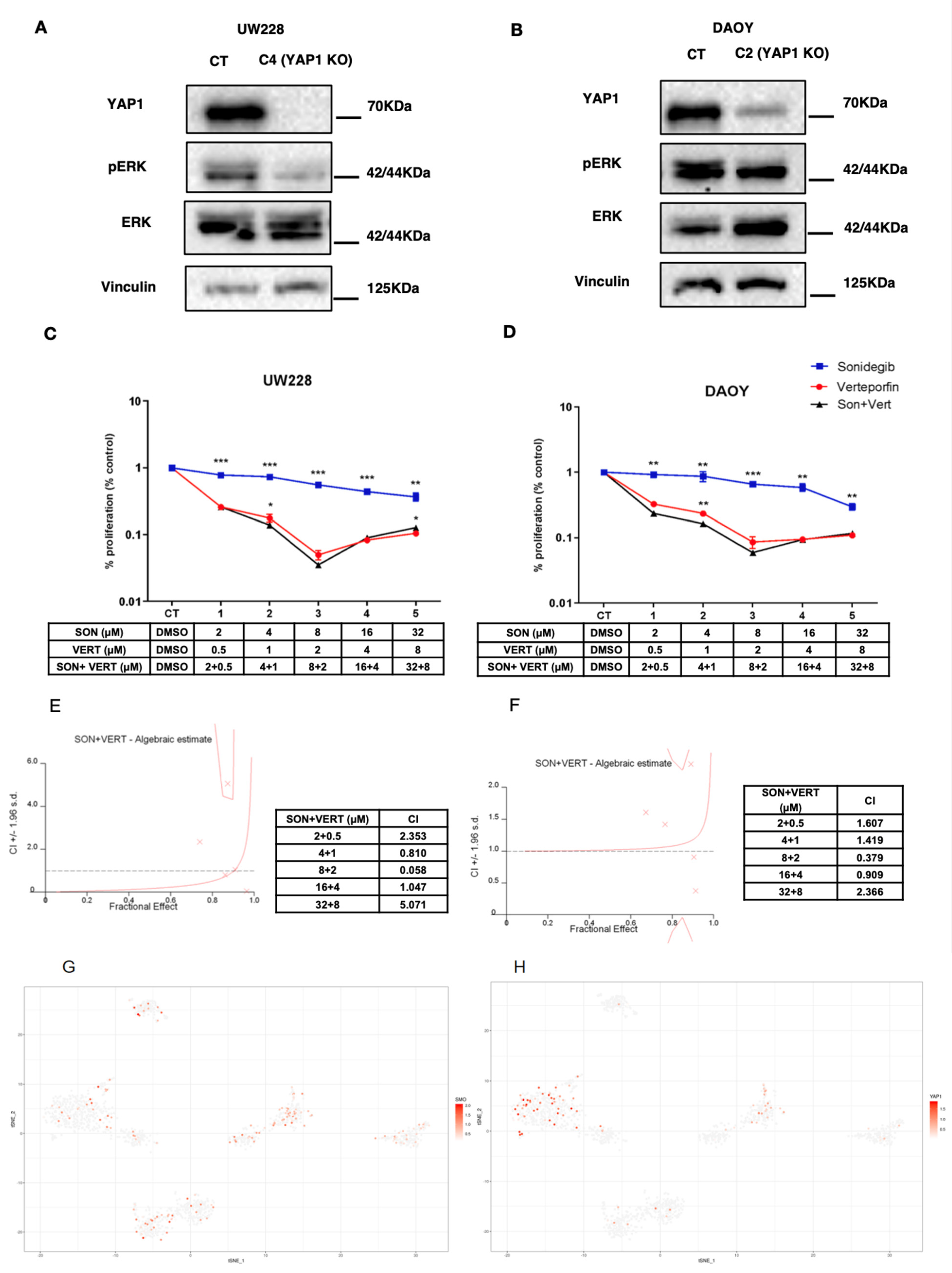
3.1. YAP1 Knockout in MB Cell Lines Bearing TP53 Mutation Leads to Anti-Proliferative and Pro-Apoptotic Response to Sonidegib
3.2. Sonidegib and Verteporfin in Specific Concentrations Show a Synergic Effect in DAOY and UW228 Cell Lines
3.3. Single-cell RNAseq Analysis Reveals That SMO, a Target of Sonidegib, and YAP1 Is Expressed in Distinct Subpopulations of MB Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cavalli, F.M.; Remke, M.; Rampasek, L.; Peacock, J.; Shih, D.J.H.; Luu, B.; Garzia, L.; Torchia, J.; Nor, C.; Morrissy, S.; et al. Intertumoral Heterogeneity within Medulloblastoma Subgroups. Cancer Cell 2017, 31, 737–754. [Google Scholar] [CrossRef] [Green Version]
- Northcott, P.A.; Robinson, G.W.; Kratz, C.P.; Mabbott, D.J.; Pomeroy, S.L.; Clifford, S.C.; Rutkowski, S.; Ellison, D.W.; Malkin, D.; Taylor, M.; et al. Medulloblastoma. Nat. Rev. Dis. Prim. 2019, 5, 11. [Google Scholar] [CrossRef] [PubMed]
- Ramaswamy, V.; Remke, M.; Bouffet, E.; Faria, C.C.; Perreault, S.; Cho, Y.-J.; Shih, D.J.; Luu, B.; Dubuc, A.M.; A Northcott, P.; et al. Recurrence patterns across medulloblastoma subgroups: An integrated clinical and molecular analysis. Lancet Oncol. 2013, 14, 1200–1207. [Google Scholar] [CrossRef] [Green Version]
- Kool, M.; Jones, D.T.W.; Jaeger, N.; Northcott, P.A.; Pugh, T.J.; Hovestadt, V.; Piro, R.M.; Esparza, L.A.; Markant, S.L.; Remke, M.; et al. Genome Sequencing of SHH Medulloblastoma Predicts Genotype-Related Response to Smoothened Inhibition. Cancer Cell 2014, 25, 393–405. [Google Scholar] [CrossRef] [Green Version]
- Schwalbe, E.; Lindsey, J.C.; Nakjang, S.; Crosier, S.; Smith, A.J.; Hicks, D.; Rafiee, G.; Hill, R.M.; Iliasova, A.; Stone, T.; et al. Novel molecular subgroups for clinical classification and outcome prediction in childhood medulloblastoma: A cohort study. Lancet Oncol. 2017, 18, 958–971. [Google Scholar] [CrossRef] [Green Version]
- Cruzeiro, G.A.V.; Lira, R.; de Almeida Magalhães, T.; Scrideli, C.; Valera, E.T.; Baumgartner, M.; Tone, L.G. CTGF expression is indicative of better survival rates in patients with medulloblastoma. Cancer Gene Ther. 2020, 27, 378–382. [Google Scholar] [CrossRef]
- Robinson, M.H.; Maximov, V.; Lallani, S.; Farooq, H.; Taylor, M.D.; Read, R.D.; Kenney, A.M. Upregulation of the chromatin remodeler HELLS is mediated by YAP1 in Sonic Hedgehog Medulloblastoma. Sci. Rep. 2019, 9, 13611–13615. [Google Scholar] [CrossRef] [PubMed]
- Huang, A.; Garraway, L.A.; Ashworth, A.; Weber, B. Synthetic lethality as an engine for cancer drug target discovery. Nat. Rev. Drug Discov. 2019, 19, 23–38. [Google Scholar] [CrossRef] [PubMed]
- Cruzeiro, G.A.V.; Salomão, K.B.; de Biagi, C.A.O., Jr.; Baumgartner, M.; Sturm, D.; Lira, R.; Magalhães, T.D.A.; Milan, M.B.; Silveira, V.D.S.; Saggioro, F.P.; et al. A simplified approach using Taqman low-density array for medulloblastoma subgrouping. Acta Neuropathol. Commun. 2019, 7, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Triscott, J.; Lee, C.; Foster, C.; Manoranjan, B.; Pambid, M.R.; Berns, R.; Fotovati, A.; Venugopal, C.; O’Halloran, K.; Narendran, A.; et al. Personalizing the Treatment of Pediatric Medulloblastoma: Polo-like Kinase 1 as a Molecular Target in High-Risk Children. Cancer Res. 2013, 73, 6734–6744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivanov, D.P.; Coyle, B.; Walker, D.A.; Grabowska, A.M. In vitro models of medulloblastoma: Choosing the right tool for the job. J. Biotechnol. 2016, 236, 10–25. [Google Scholar] [CrossRef]
- Greenfield, E.A. Single-Cell Cloning of Hybridoma Cells by Limiting Dilution. Cold Spring Harb. Protoc. 2019, 2019, pdb-prot103192. [Google Scholar] [CrossRef]
- De Almeida Magalhães, T.; Cruzeiro, G.A.V.; de Sousa, G.R.; Da Silva, K.R.; Lira, R.C.P.; Scrideli, C.; Tone, L.G.; Valera, E.T.; Borges, K.S. Notch pathway in ependymoma RELA-fused subgroup: Upregulation and association with cancer stem cells markers expression. Cancer Gene Ther. 2019, 27, 509–512. [Google Scholar] [CrossRef] [Green Version]
- Klinger, P.H.D.S.; Delsin, L.E.A.; Cruzeiro, G.A.V.; Andrade, A.F.; Lira, R.C.P.; De Andrade, P.V.; Das Chagas, P.F.; Queiroz, R.G.D.P.; Trevisan, F.A.; De Oliveira, R.S.; et al. Arsenic Trioxide exerts cytotoxic and radiosensitizing effects in pediatric Medulloblastoma cell lines of SHH Subgroup. Sci. Rep. 2020, 10, 6836. [Google Scholar] [CrossRef]
- Kieran, M.W.; Chisholm, J.; Casanova, M.; Brandes, A.A.; Aerts, I.; Bouffet, E.; Bailey, S.; Leary, S.; Macdonald, T.J.; Mechinaud, F.; et al. Phase I study of oral sonidegib (LDE225) in pediatric brain and solid tumors and a phase II study in children and adults with relapsed medulloblastoma. Neuro-Oncology 2017, 19, 1542–1552. [Google Scholar] [CrossRef]
- Butler, A.; Hoffman, P.; Smibert, P.; Papalexi, E.; Satija, R. Integrating single-cell transcriptomic data across different conditions, technologies, and species. Nat. Biotechnol. 2018, 36, 411–420. [Google Scholar] [CrossRef]
- Stuart, T.; Butler, A.; Hoffman, P.; Hafemeister, C.; Papalexi, E.; Mauck, W.M., III; Hao, Y.; Stoeckius, M.; Smibert, P.; Satija, R. Comprehensive Integration of Single-Cell Data. Cell 2019, 177, 1888–1902.E21. [Google Scholar] [CrossRef] [PubMed]
- Hovestadt, V.; Smith, K.S.; Bihannic, L.; Filbin, M.G.; Shaw, M.L.; Baumgartner, A.; DeWitt, J.C.; Groves, A.; Mayr, L.; Weisman, H.R.; et al. Resolving medulloblastoma cellular architecture by single-cell genomics. Nat. Cell Biol. 2019, 572, 74–79. [Google Scholar] [CrossRef] [PubMed]
- Keren-Paz, A.; Emmanuel, R.; Samuels, Y. YAP and the drug resistance highway. Nat. Genet. 2015, 47, 193–194. [Google Scholar] [CrossRef] [Green Version]
- Fernandez-L, A.; Northcott, P.A.; Dalton, J.; Fraga, C.; Ellison, D.; Angers, S.; Taylor, M.D.; Kenney, A.M. YAP1 is amplified and up-regulated in hedgehog-associated medulloblastomas and mediates Sonic hedgehog-driven neural precursor proliferation. Genes Dev. 2009, 23, 2729–2741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chou, T.C. Drug Combination Studies and Their Synergy Quantification Using the Chou-Talalay Method. Cancer Res. 2010, 70, 440–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Zhu, X.; Feng, W.; Yu, Y.; Jeong, K.; Guo, W.; Lu, Y.; Mills, G.B. Verteporfin inhibits YAP function through up-regulating 14-3-3σ sequestering YAP in the cytoplasm. Am. J. Cancer Res. 2015, 6, 27–37. [Google Scholar] [PubMed]
- Zhang, L.; He, X.; Liu, X.; Zhang, F.; Huang, L.; Potter, A.S.; Xu, L.; Zhou, W.; Zheng, T.; Luo, Z.; et al. Single-Cell Transcriptomics in Medulloblastoma Reveals Tumor-Initiating Progenitors and Oncogenic Cascades during Tumorigenesis and Relapse. Cancer Cell 2019, 36, 302–318. [Google Scholar] [CrossRef] [PubMed]
- Raj, N.; Bam, R. Reciprocal Crosstalk between YAP1/Hippo Pathway and the p53 Family Proteins: Mechanisms and Outcomes in Cancer. Front. Cell Dev. Biol. 2019, 7, 159. [Google Scholar] [CrossRef] [Green Version]
- Sorokin, M.; Borisov, N.; Kuzmin, D.; Gudkov, A.; Zolotovskaia, M.; Garazha, A.; Buzdin, A. Algorithmic Annotation of Functional Roles for Components of 3,044 Human Molecular Pathways. Front. Genet. 2021, 12, 617059. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alencastro Veiga Cruzeiro, G.; de Almeida Magalhães, T.; Ribeiro de Sousa, G.; Bonfim Silva, R.; Alberto Oliveira de Biagi Junior, C.; Ferreira das Chagas, P.; Gomes de Paula Queiroz, R.; Alberto Scrideli, C.; Gonzaga Tone, L.; Terci Valera, E. YAP1 Is a Potential Predictive Molecular Biomarker for Response to SMO Inhibitor in Medulloblastoma Cells. Cancers 2021, 13, 6249. https://doi.org/10.3390/cancers13246249
Alencastro Veiga Cruzeiro G, de Almeida Magalhães T, Ribeiro de Sousa G, Bonfim Silva R, Alberto Oliveira de Biagi Junior C, Ferreira das Chagas P, Gomes de Paula Queiroz R, Alberto Scrideli C, Gonzaga Tone L, Terci Valera E. YAP1 Is a Potential Predictive Molecular Biomarker for Response to SMO Inhibitor in Medulloblastoma Cells. Cancers. 2021; 13(24):6249. https://doi.org/10.3390/cancers13246249
Chicago/Turabian StyleAlencastro Veiga Cruzeiro, Gustavo, Taciani de Almeida Magalhães, Graziella Ribeiro de Sousa, Ricardo Bonfim Silva, Carlos Alberto Oliveira de Biagi Junior, Pablo Ferreira das Chagas, Rosane Gomes de Paula Queiroz, Carlos Alberto Scrideli, Luiz Gonzaga Tone, and Elvis Terci Valera. 2021. "YAP1 Is a Potential Predictive Molecular Biomarker for Response to SMO Inhibitor in Medulloblastoma Cells" Cancers 13, no. 24: 6249. https://doi.org/10.3390/cancers13246249