Prognostic Value and Function of KLF5 in Papillary Thyroid Cancer

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Results
2.1. KLF5 Expression in Normal Thyroid and PTC Tissues
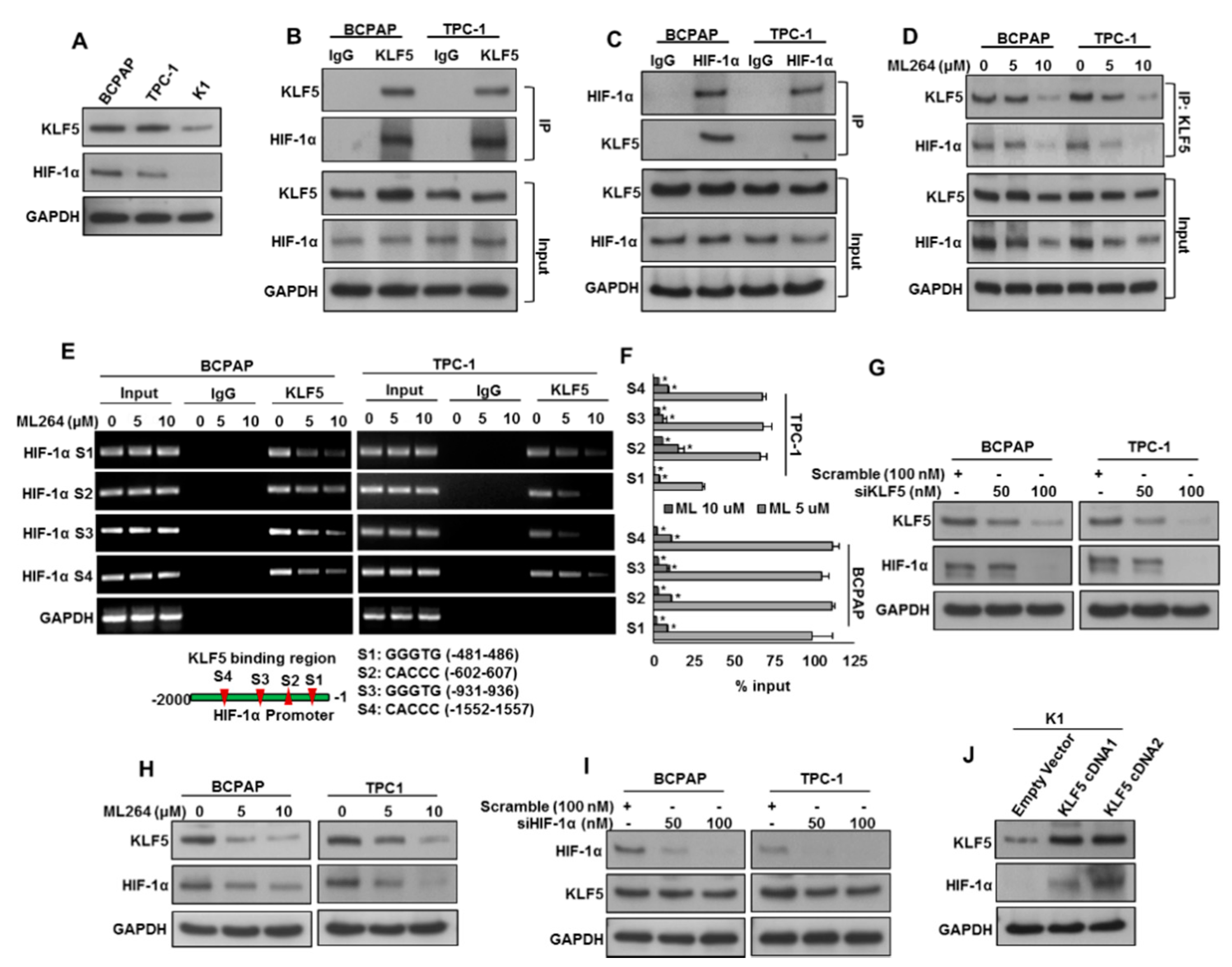
2.2. KLF5 Functions Upstream of HIF-1α.
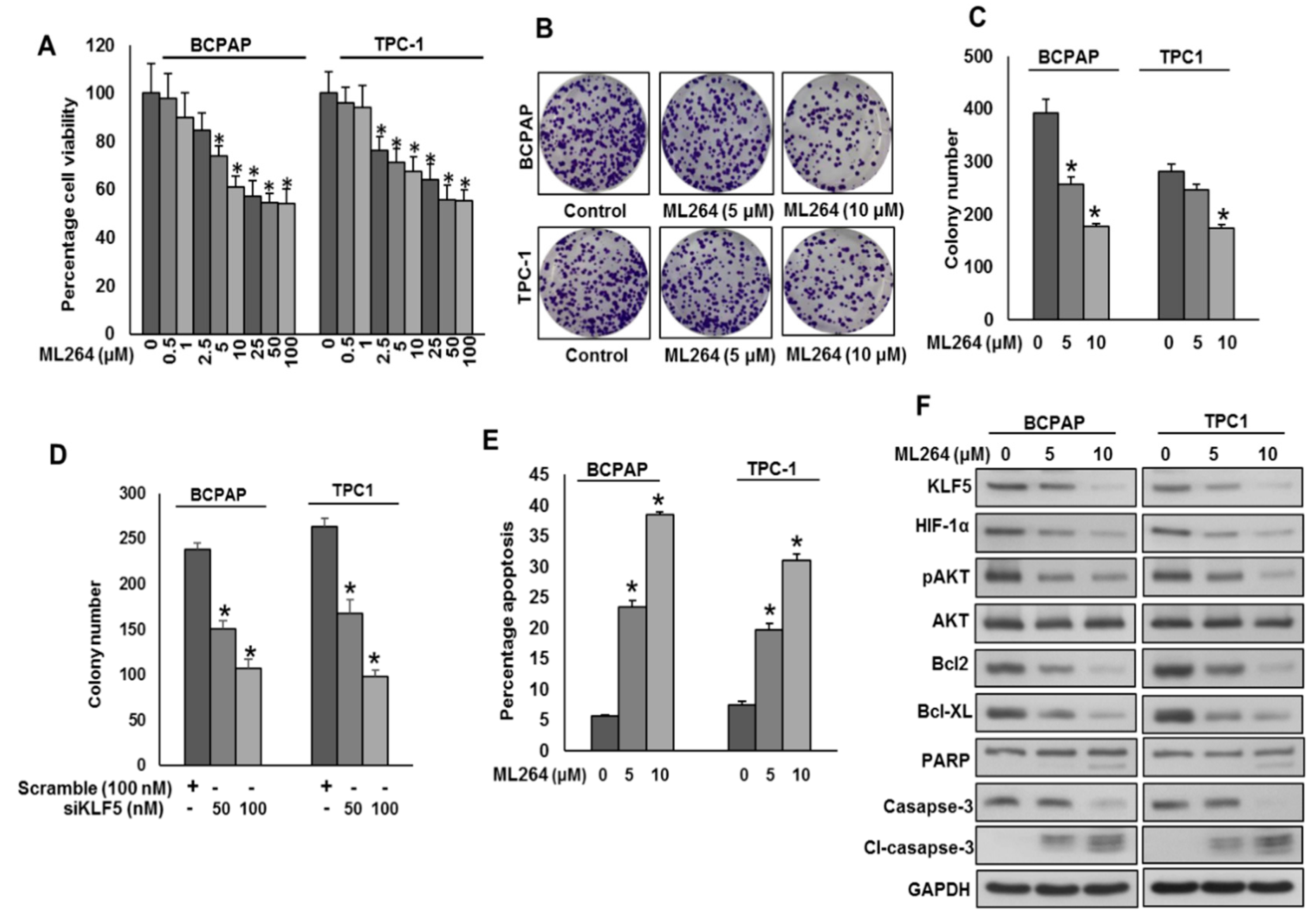
2.3. Downregulation of KLF5 Inhibits Tumor Cell Invasion, Migration, and Angiogenesis
2.4. Downregulation of KLF5 Inhibits PTC Cell Growth In Vitro
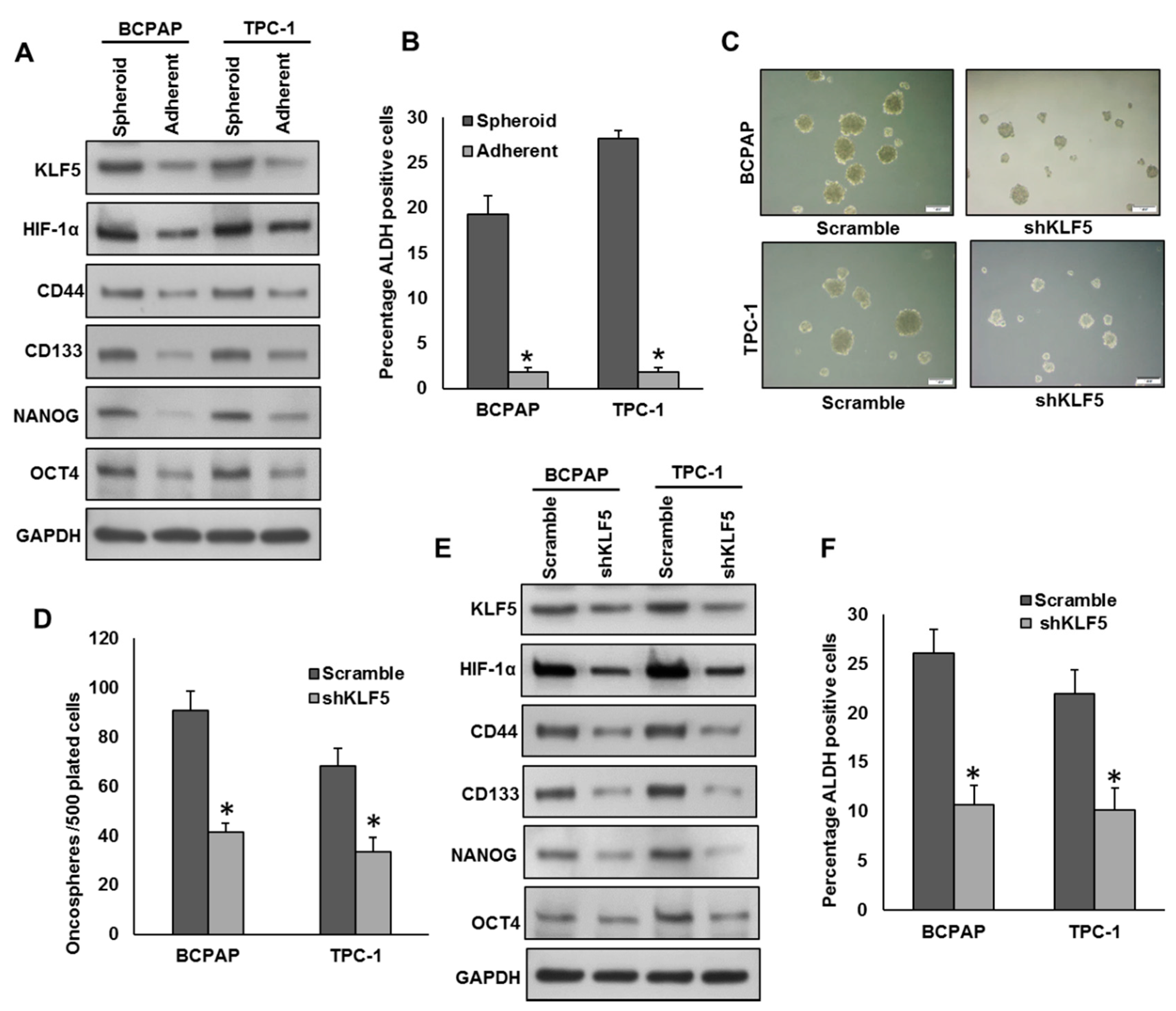
2.5. Inhibition of KLF5 Decreases the Self-Renewal Ability of Spheroids Generated from PTC Cells
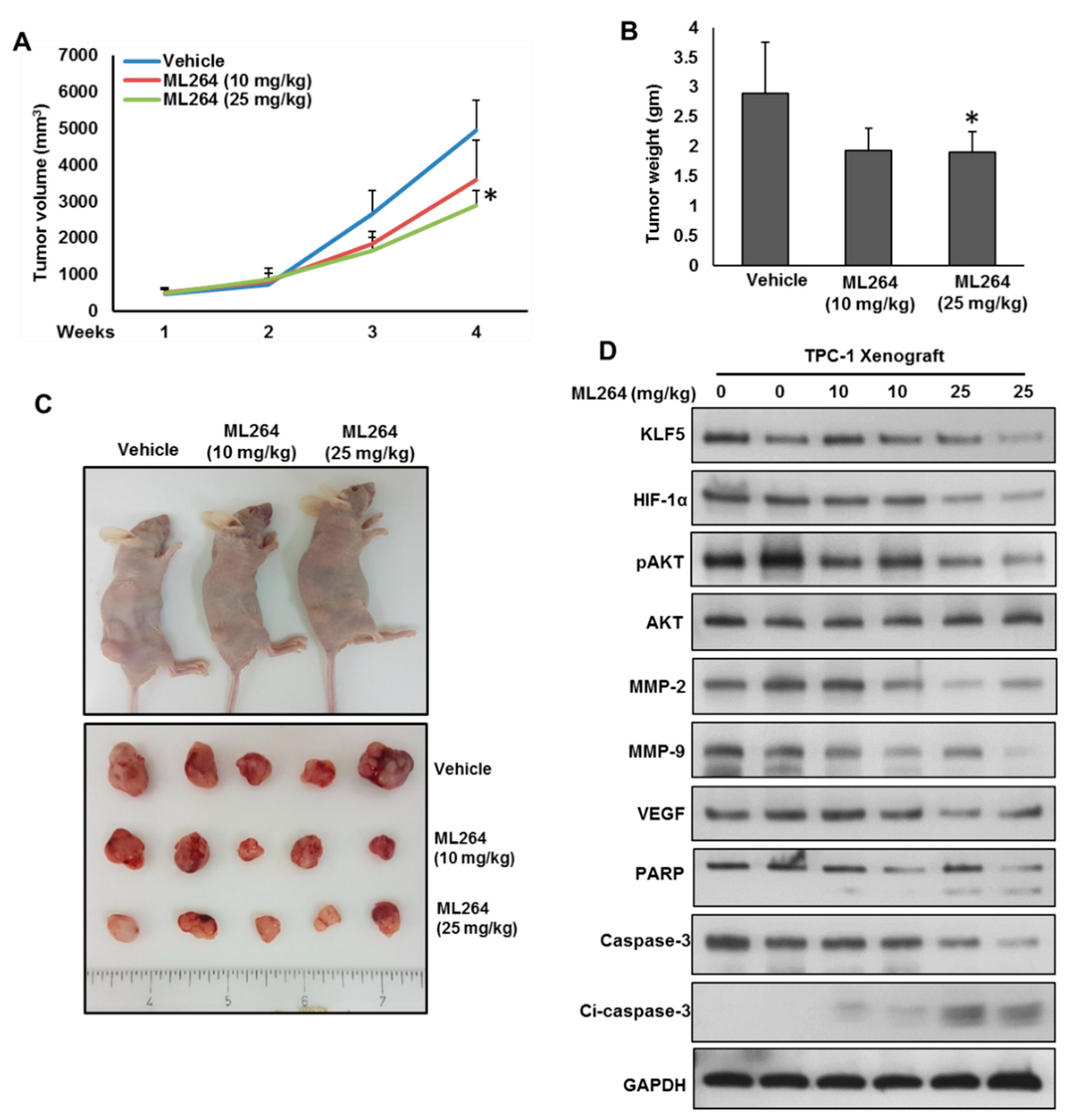
2.6. Downregulation of KLF5 Inhibits PTC Cell Growth In Vivo
3. Discussion
4. Materials and Methods
4.1. Sample Selection
4.2. Tissue Microarray Construction and Immunohistochemistry
4.3. Cell Culture
4.4. Reagents and Antibodies
4.5. MTT (3-(4,5-Dimethylthiazol-2-yl)-2,5-Diphenyltetrazolium Bromide) Assays
4.6. Gene Silencing Using siRNA
4.7. Plasmid and Transfection
4.8. Cell Invasion and Migration Assays
4.9. Sphere Forming Assay
4.10. The ALDH Activity
4.11. Chromatin Immunoprecipitation (ChIP) Assay
4.12. Animals and Xenografts Study
4.13. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Howlader, N.; Noone, A.; Krapcho, M.; Neyman, N.; Aminou, R.; Altekruse, S.; Kosary, C.; Ruhl, J.; Tatalovich, Z.; Cho, H. SEER Cancer Statistics Review, 1975–2009 (Vintage 2009 Populations); National Cancer Institute: Bethesda, MD, USA, 2012.
- Tumino, D.; Frasca, F.; Newbold, K. Updates on the management of advanced, metastatic, and radioiodine refractory differentiated thyroid cancer. Front. Endocrinol. (Lausanne) 2017, 8, 312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grant, C.S. Recurrence of papillary thyroid cancer after optimized surgery. Gland Surg. 2015, 4, 52. [Google Scholar] [PubMed]
- Lee, J.-W.; Bae, S.-H.; Jeong, J.-W.; Kim, S.-H.; Kim, K.-W. Hypoxia-inducible factor (HIF-1) α: Its protein stability and biological functions. Exp. Mol. Med. 2004, 36, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Mori, A.; Moser, C.; Lang, S.A.; Hackl, C.; Gottfried, E.; Kreutz, M.; Schlitt, H.J.; Geissler, E.K.; Stoeltzing, O. Up-regulation of Krüppel-like factor 5 in pancreatic cancer is promoted by interleukin-1β signaling and hypoxia-inducible factor-1α. Mol. Cancer Res. 2009, 7, 1390–1398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Kong, R.; Chen, H.; Zhao, Z.; Li, L.; Li, J.; Hu, J.; Zhang, G.; Pan, S.; Wang, Y. Overexpression of KLF5 is associated with poor survival and G1/S progression in pancreatic cancer. Aging (Albany NY) 2019, 11, 5035. [Google Scholar] [CrossRef]
- Ma, D.; Chang, L.-Y.; Zhao, S.; Zhao, J.-J.; Xiong, Y.-J.; Cao, F.-Y.; Yuan, L.; Zhang, Q.; Wang, X.-Y.; Geng, M.-L. KLF5 promotes cervical cancer proliferation, migration and invasion in a manner partly dependent on TNFRSF11a expression. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Wu, K.; Chen, Y.; Zhou, J.; Du, C.; Shi, Q.; Xu, S.; Jia, J.; Tang, X.; Li, F. Beyond proliferation: KLF5 promotes angiogenesis of bladder cancer through directly regulating VEGFA transcription. Oncotarget 2015, 6, 43791. [Google Scholar] [CrossRef]
- Maehara, O.; Sato, F.; Natsuizaka, M.; Asano, A.; Kubota, Y.; Itoh, J.; Tsunematsu, S.; Terashita, K.; Tsukuda, Y.; Nakai, M. A pivotal role of Krüppel-like factor 5 in regulation of cancer stem-like cells in hepatocellular carcinoma. Cancer Biol. Ther. 2015, 16, 1453–1461. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Wang, Q.; Liu, F.; Ma, X.; Wu, L.; Guo, F.; Zhao, S.; Huang, F.; Qin, G. KLF5 promotes the tumorigenesis and metastatic potential of thyroid cancer cells through the NF-κB signaling pathway. Oncol. Rep. 2018, 40, 2608–2618. [Google Scholar] [CrossRef]
- Takagi, K.; Miki, Y.; Onodera, Y.; Nakamura, Y.; Ishida, T.; Watanabe, M.; Inoue, S.; Sasano, H.; Suzuki, T. Krüppel-like factor 5 in human breast carcinoma: A potent prognostic factor induced by androgens. Endocr. Relat. Cancer 2012, 19, 741–750. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Dong, Z.; Zhou, F.; Cai, X.; Gao, Y.; Wang, L.-W. Krüppel-like factor 5 promotes lung tumorigenesis through upregulation of Sox4. Cell. Physiol. Biochem. 2014, 33, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, Z.; Yang, L.; Lai, D. KLF 5 strengthens drug resistance of ovarian cancer stem-like cells by regulating survivin expression. Cell Prolif. 2013, 46, 425–435. [Google Scholar] [CrossRef] [PubMed]
- Tong, D.; Czerwenka, K.; Heinze, G.; Ryffel, M.; Schuster, E.; Witt, A.; Leodolter, S.; Zeillinger, R. Expression of KLF5 is a prognostic factor for disease-free survival and overall survival in patients with breast cancer. Clin. Cancer Res. 2006, 12, 2442–2448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.-J.; No, Y.R.; Dang, D.T.; Dang, L.H.; Yang, V.W.; Shim, H.; Yun, C.C. Regulation of hypoxia-inducible factor 1α (HIF-1α) by lysophosphatidic acid is dependent on interplay between p53 and Krüppel-like factor 5. J. Biol. Chem. 2013, 288, 25244–25253. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Liu, X.; Xu, Y.; Liu, J.; Xie, M.; Ni, W.; Chen, S. KLF5 promotes hypoxia-induced survival and inhibits apoptosis in non-small cell lung cancer cells via HIF-1α. Int. J. Oncol. 2014, 45, 1507–1514. [Google Scholar] [CrossRef] [Green Version]
- Bu, R.; Siraj, A.K.; Divya, S.P.; Kong, Y.; Parvathareddy, S.K.; Al-Rasheed, M.; Al-Obaisi, K.A.; Victoria, I.G.; Al-Sobhi, S.S.; Al-Dawish, M. Telomerase reverse transcriptase mutations are independent predictor of disease-free survival in M iddle E astern papillary thyroid cancer. Int. J. Cancer 2018, 142, 2028–2039. [Google Scholar] [CrossRef] [Green Version]
- Soon, M.-S.; Hsu, L.-S.; Chen, C.-J.; Chu, P.-Y.; Liou, J.-H.; Lin, S.-H.; Hsu, J.-D.; Yeh, K.-T. Expression of Krűppel-like factor 5 in gastric cancer and its clinical correlation in Taiwan. Virchows Arch. 2011, 459, 161. [Google Scholar] [CrossRef]
- Zhang, H.; Shao, F.; Guo, W.; Gao, Y.; He, J. Knockdown of KLF5 promotes cisplatin-induced cell apoptosis via regulating DNA damage checkpoint proteins in non-small cell lung cancer. Thorac. Cancer 2019, 10, 1069–1077. [Google Scholar] [CrossRef] [Green Version]
- Bialkowska, A.; Crisp, M.; Madoux, F.; Spicer, T.; Knapinska, A.; Mercer, B.; Bannister, T.D.; He, Y.; Chowdhury, S.; Cameron, M. ML264: An antitumor agent that potently and selectively inhibits Krüppel-like factor five (KLF5) expression: A probe for studying Colon Cancer development and progression. In Probe Reports from the NIH Molecular Libraries Program [Internet]; National Center for Biotechnology Information (US): Bethesda, MD, USA, 2013. [Google Scholar]
- Gao, Y.; Ding, Y.; Chen, H.; Chen, H.; Zhou, J. Targeting Kruppel-like factor 5 (KLF5) for cancer therapy. Curr. Top. Med. Chem. 2015, 15, 699–713. [Google Scholar] [CrossRef]
- Huang, H.; Han, Y.; Chen, Z.; Pan, X.; Yuan, P.; Zhao, X.; Zhu, H.; Wang, J.; Sun, X.; Shi, P. ML264 inhibits osteosarcoma growth and metastasis via inhibition of JAK2/STAT3 and WNT/β-catenin signalling pathways. J. Cell. Mol. Med. 2020, 24, 5652–5664. [Google Scholar] [CrossRef] [Green Version]
- de Sabando, A.R.; Wang, C.; He, Y.; García-Barros, M.; Kim, J.; Shroyer, K.R.; Bannister, T.D.; Yang, V.W.; Bialkowska, A.B. ML264, a novel small-molecule compound that potently inhibits growth of colorectal cancer. Mol. Cancer Ther. 2016, 15, 72–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siraj, A.; Bavi, P.; Abubaker, J.; Jehan, Z.; Sultana, M.; Al-Dayel, F.; Al-Nuaim, A.; Alzahrani, A.; Ahmed, M.; Al-Sanea, O. Genome-wide expression analysis of Middle Eastern papillary thyroid cancer reveals c-MET as a novel target for cancer therapy. J. Pathol. 2007, 213, 190–199. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, Y.; Akaogi, K.; Suzuki, T.; Osakabe, A.; Yamaguchi, C.; Sunahara, N.; Ishida, J.; Kako, K.; Ogawa, S.; Fujimura, T. Estrogen regulates tumor growth through a nonclassical pathway that includes the transcription factors ERβ and KLF5. Sci. Signal. 2011, 4, ra22. [Google Scholar] [CrossRef]
- Qin, R.; Smyrk, T.C.; Reed, N.; Schmidt, R.; Schnelldorfer, T.; Chari, S.T.; Petersen, G.M.; Tang, A. Combining clinicopathological predictors and molecular biomarkers in the oncogenic K-RAS/Ki67/HIF-1α pathway to predict survival in resectable pancreatic cancer. Br. J. Cancer 2015, 112, 514–522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, M.; Uddin, S.; Hussain, A.R.; Alyan, A.; Jehan, Z.; Al-Dayel, F.; Al-Nuaim, A.; Al-Sobhi, S.; Amin, T.; Bavi, P. FoxM1 and its association with matrix metalloproteinases (MMP) signaling pathway in papillary thyroid carcinoma. J. Clin. Endocrinol. Metab. 2012, 97, E1–E13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, M.; Sano, D.; Pickering, C.R.; Jasser, S.A.; Henderson, Y.C.; Clayman, G.L.; Sturgis, E.M.; Ow, T.J.; Lotan, R.; Carey, T.E. Assembly and initial characterization of a panel of 85 genomically validated cell lines from diverse head and neck tumor sites. Clin. Cancer Res. 2011, 17, 7248–7264. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clinico-Pathological Characteristics | Total | KLF5 Over-Expression | KLF5 Low Expression | p-Value | |||
|---|---|---|---|---|---|---|---|
| No. | % | No. | % | No. | % | ||
| No. of patients | 1219 | 793 | 65.1 | 426 | 34.9 | ||
| Age (Y) | |||||||
| <45 | 774 | 64.1 | 490 | 63.3 | 284 | 36.7 | 0.0858 |
| ≥45 | 434 | 35.9 | 296 | 68.2 | 138 | 31.8 | |
| Sex | |||||||
| Female | 916 | 75.9 | 587 | 64.1 | 329 | 35.9 | 0.2151 |
| Male | 291 | 24.1 | 198 | 68.0 | 93 | 32.0 | |
| Histology Type | |||||||
| Classical Variant | 839 | 73.9 | 554 | 66.0 | 285 | 34.0 | <0.0001 |
| Follicular Variant | 191 | 16.8 | 102 | 53.4 | 89 | 46.6 | |
| Tall-Cell Variant | 105 | 9.3 | 83 | 79.0 | 22 | 21.0 | |
| Extrathyroidal extension | |||||||
| Absent | 523 | 49.4 | 314 | 60.0 | 209 | 40.0 | 0.0003 |
| Present | 536 | 50.6 | 379 | 70.7 | 157 | 29.3 | |
| pT | |||||||
| pT1 | 307 | 26.1 | 186 | 60.6 | 118 | 39.4 | 0.1009 |
| pT2 | 239 | 20.3 | 152 | 63.6 | 87 | 36.4 | |
| pT3 | 482 | 41.0 | 330 | 68.5 | 152 | 31.5 | |
| pT4 | 148 | 12.6 | 104 | 70.3 | 44 | 29.7 | |
| pN | |||||||
| pN0 | 553 | 46.8 | 313 | 56.6 | 240 | 43.4 | <0.0001 |
| pN1 | 628 | 53.2 | 453 | 72.1 | 175 | 27.9 | |
| pM | |||||||
| pM0 | 1149 | 95.5 | 747 | 65.0 | 402 | 35.0 | 0.8026 |
| pM1 | 54 | 4.5 | 36 | 66.7 | 16 | 33.3 | |
| Stage | |||||||
| I | 811 | 68.9 | 511 | 63.0 | 300 | 37.0 | 0.0003 |
| II | 94 | 8.0 | 54 | 57.4 | 40 | 42.6 | |
| III | 122 | 10.3 | 78 | 69.6 | 44 | 30.4 | |
| IVA | 114 | 9.7 | 95 | 83.3 | 19 | 16.7 | |
| IVB | 36 | 3.1 | 25 | 69.4 | 11 | 30.6 | |
| HIF-1α IHC | |||||||
| High | 796 | 68.7 | 530 | 66.6 | 266 | 33.4 | 0.0492 |
| Low | 363 | 31.3 | 220 | 60.6 | 143 | 39.4 | |
| pAKT IHC | |||||||
| High | 260 | 21.7 | 185 | 71.2 | 75 | 28.8 | 0.0279 |
| Low | 940 | 78.3 | 600 | 53.8 | 340 | 36.2 | |
| BRAF (V600E) mutation | |||||||
| Present | 498 | 59.5 | 368 | 73.9 | 130 | 26.1 | <0.0001 |
| Absent | 339 | 40.5 | 184 | 54.3 | 155 | 45.7 | |
| NRAS mutation | |||||||
| Present | 43 | 5.1 | 20 | 46.5 | 23 | 53.5 | 0.0073 |
| Absent | 794 | 94.9 | 532 | 67.0 | 262 | 33.0 | |
| HRAS mutation | |||||||
| Present | 24 | 2.9 | 16 | 66.7 | 8 | 33.3 | 0.9400 |
| Absent | 813 | 97.1 | 536 | 65.9 | 277 | 34.1 | |
| Disease Free Survival | |||||||
| 5 years | 72.3 | 80.4 | 0.0066 | ||||
| Clinicopathological Characteristics | n (%) |
|---|---|
| Age | |
| Median | 38.0 |
| Range (IQR) ^ | 29.0–51.0 |
| Gender | |
| Female | 925 (75.9) |
| Male | 294 (24.1) |
| Histopathology | |
| Classical Variant | 839 (68.8) |
| Follicular Variant | 191 (15.7) |
| Tall Cell Variant | 105 (8.6) |
| Other Variants | 84 (6.9) |
| Extra Thyroidal Extension | |
| Absent | 523 (42.9) |
| Present | 536 (44.0) |
| Unknown | 160 (13.1) |
| pT | |
| T1 | 307 (25.2) |
| T2 | 239 (19.6) |
| T3 | 482 (39.6) |
| T4 | 148 (12.1) |
| Unknown | 43 (3.5) |
| pN | |
| N0 | 553 (45.4) |
| N1 | 628 (51.5) |
| Unknown | 38 (3.1) |
| pM | |
| M0 | 1149 (94.3) |
| M1 | 54 (4.4) |
| Unknown | 16 (1.3) |
| Stage | |
| I | 811 (66.5) |
| II | 94 (7.8) |
| III | 122 (10.0) |
| IVA | 114 (9.4) |
| IVB | 36 (2.9) |
| Unknown | 42 (3.4) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pratheeshkumar, P.; Siraj, A.K.; Divya, S.P.; Parvathareddy, S.K.; Siraj, S.; Diaz, R.; Begum, R.; Al-Sobhi, S.S.; Al-Dayel, F.; Al-Kuraya, K.S. Prognostic Value and Function of KLF5 in Papillary Thyroid Cancer. Cancers 2021, 13, 185. https://doi.org/10.3390/cancers13020185
Pratheeshkumar P, Siraj AK, Divya SP, Parvathareddy SK, Siraj S, Diaz R, Begum R, Al-Sobhi SS, Al-Dayel F, Al-Kuraya KS. Prognostic Value and Function of KLF5 in Papillary Thyroid Cancer. Cancers. 2021; 13(2):185. https://doi.org/10.3390/cancers13020185
Chicago/Turabian StylePratheeshkumar, Poyil, Abdul K. Siraj, Sasidharan Padmaja Divya, Sandeep Kumar Parvathareddy, Sarah Siraj, Roxanne Diaz, Rafia Begum, Saif S. Al-Sobhi, Fouad Al-Dayel, and Khawla S. Al-Kuraya. 2021. "Prognostic Value and Function of KLF5 in Papillary Thyroid Cancer" Cancers 13, no. 2: 185. https://doi.org/10.3390/cancers13020185