A Preliminary Investigation towards the Risk Stratification of Allogeneic Stem Cell Recipients with Respect to the Potential for Development of GVHD via Their Pre-Transplant Plasma Lipid and Metabolic Signature

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients
2.2. Lipid and Metabolite Extraction for LC-MS/MS Analyses
2.3. Metabolomics: GC-MS Metabolite Extraction
2.4. Lipids: LC-MC/MC Conditions
2.5. Metabolites: GC-MS Conditions
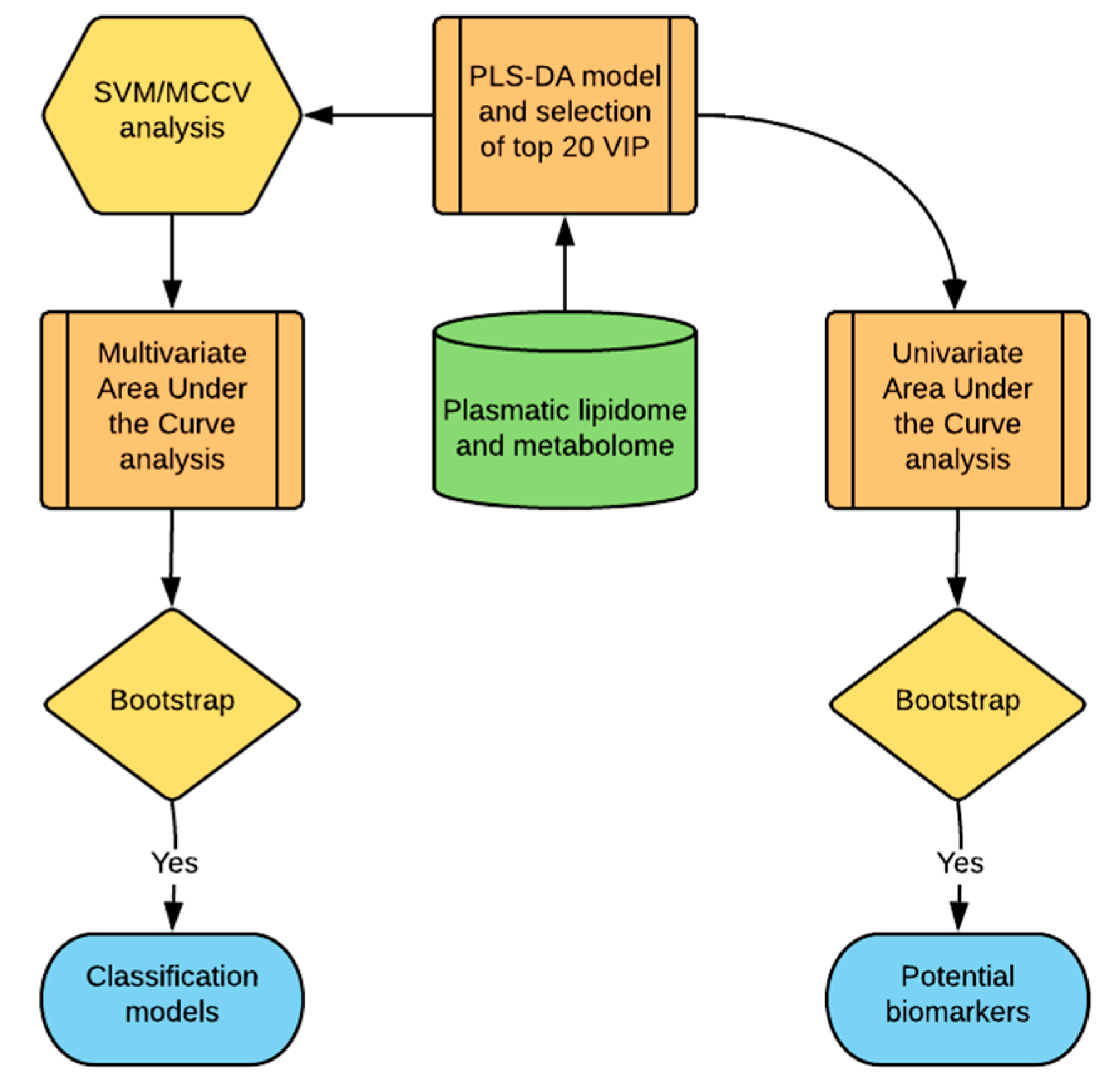
2.6. Statistical Analysis
3. Results
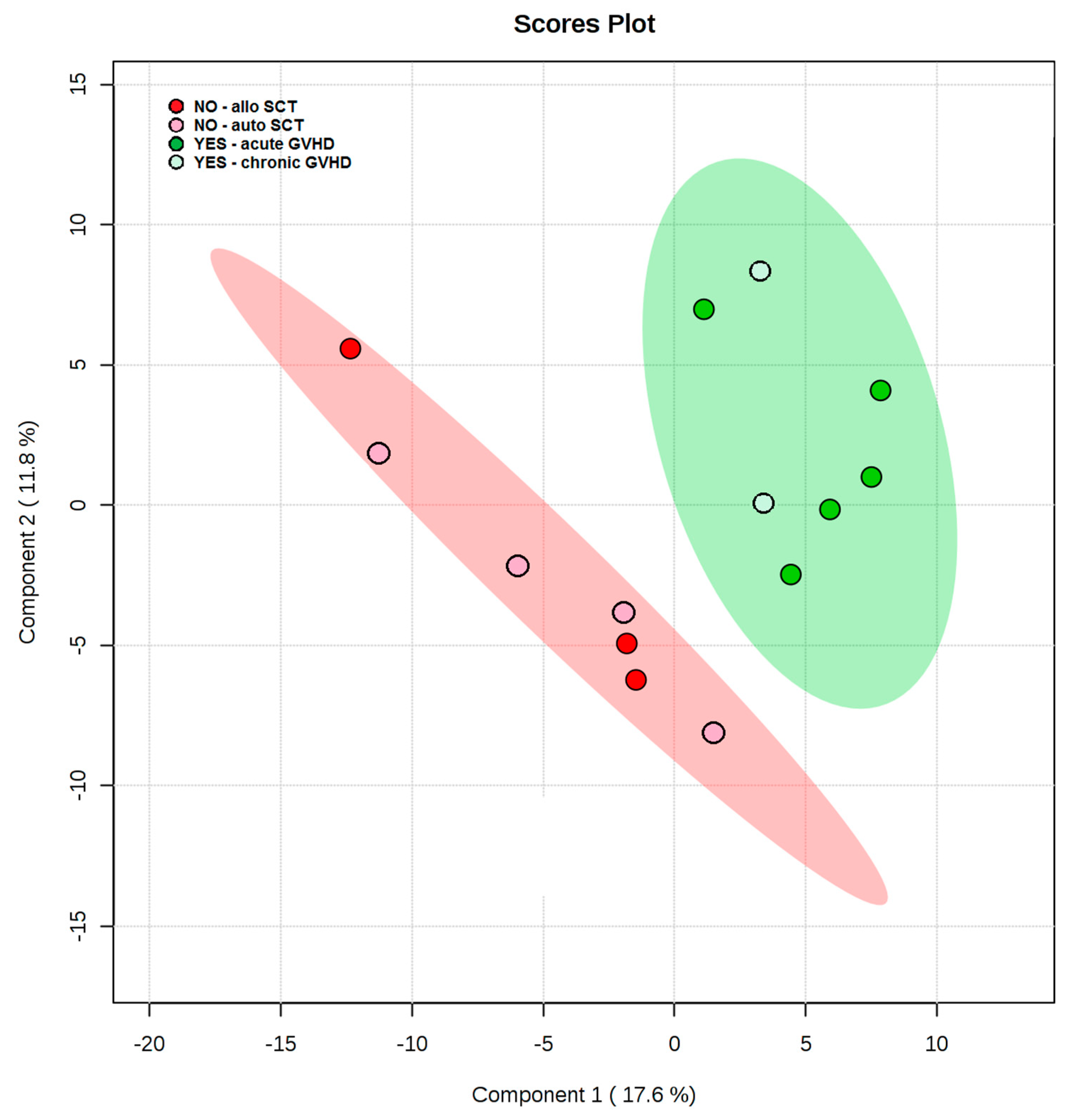
3.1. Pre-Transplant Plasma Lipid and Metabolite Profiles Reveals Class Separation between Those Patients Who Ultimately Developed GVHD and Those Who Did Not
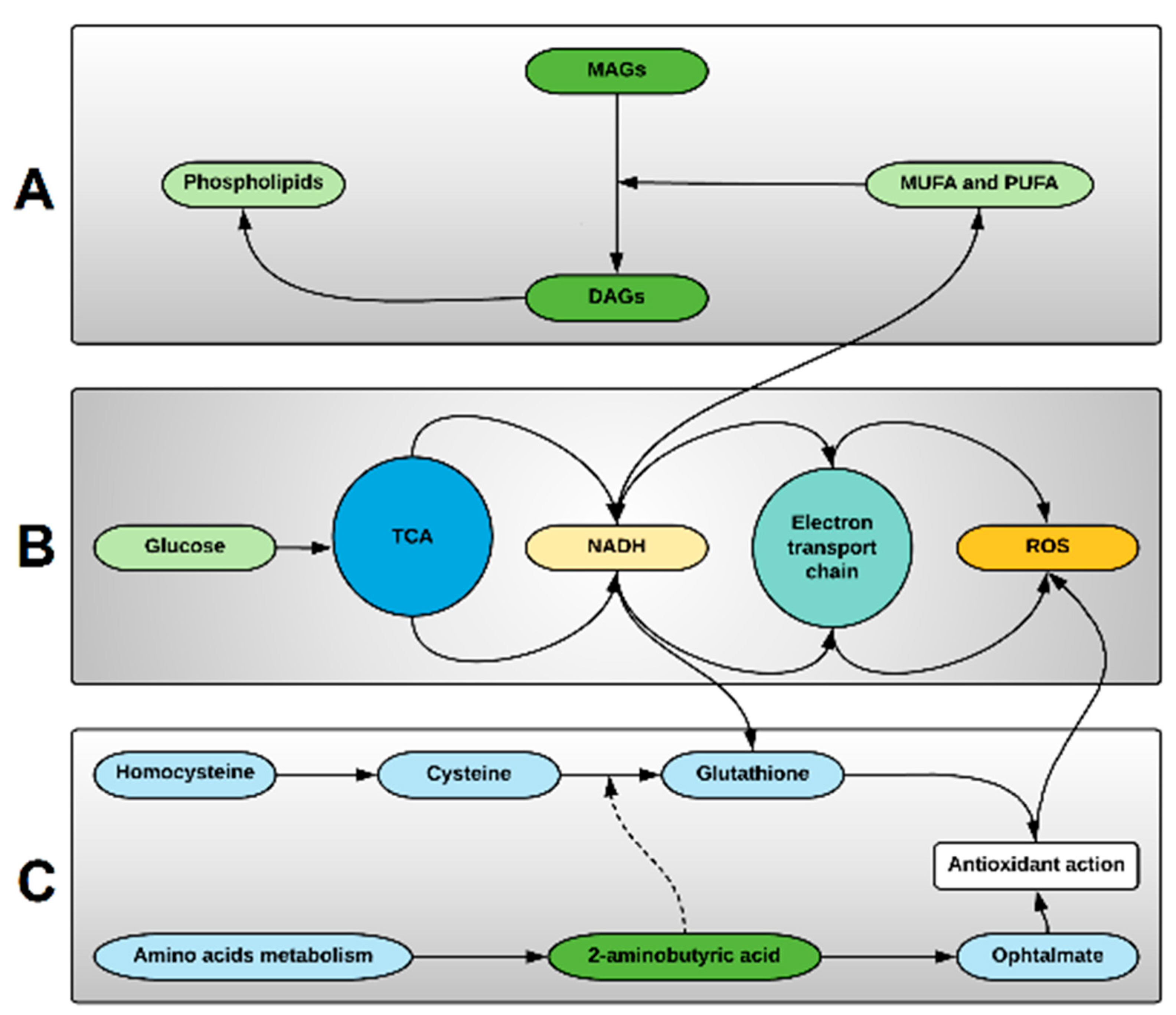
3.2. The More Important Variables for Class Separation Suggest Metabolic Pathway Tendencies Predispoising to Alloreactivity
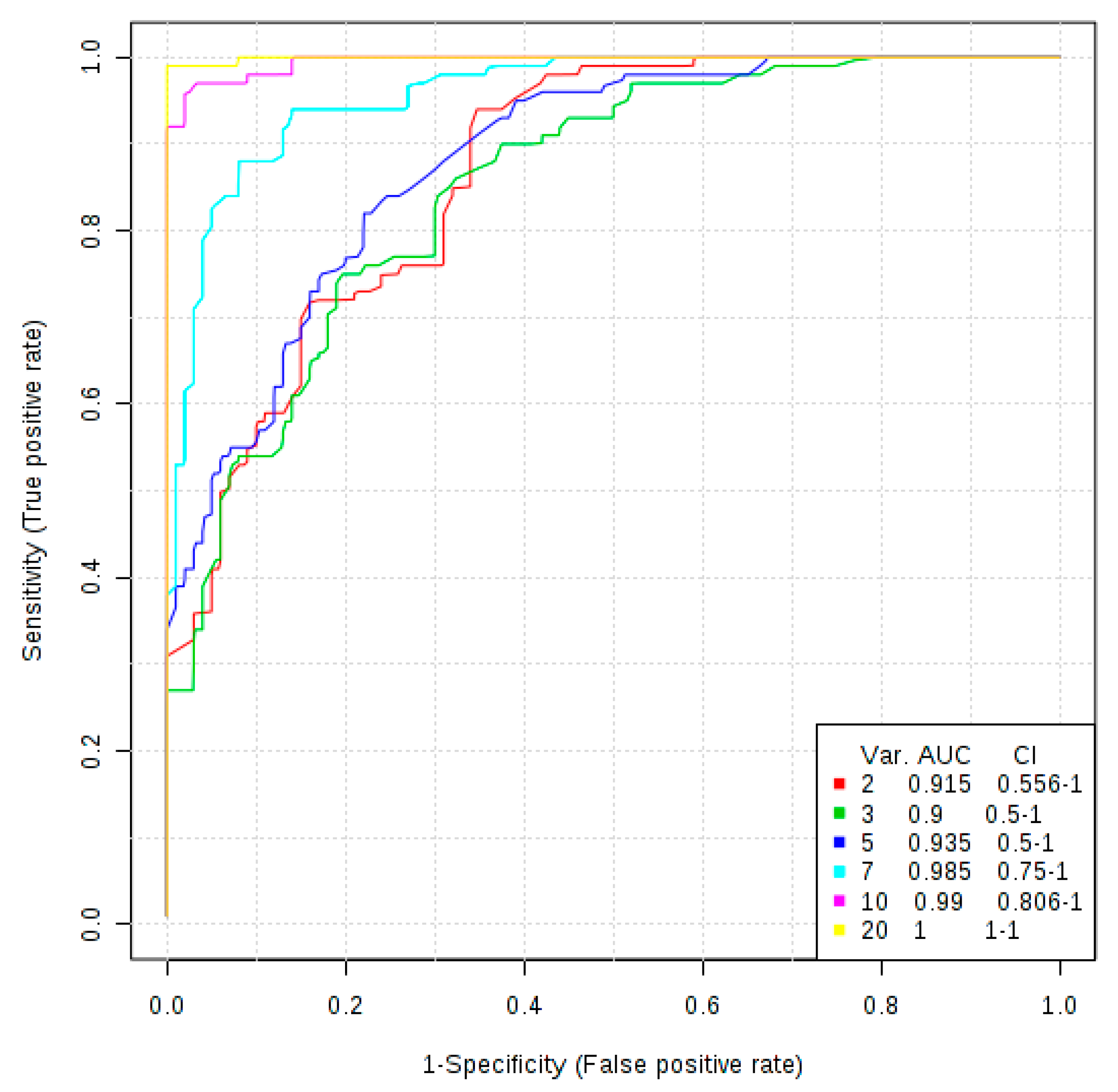
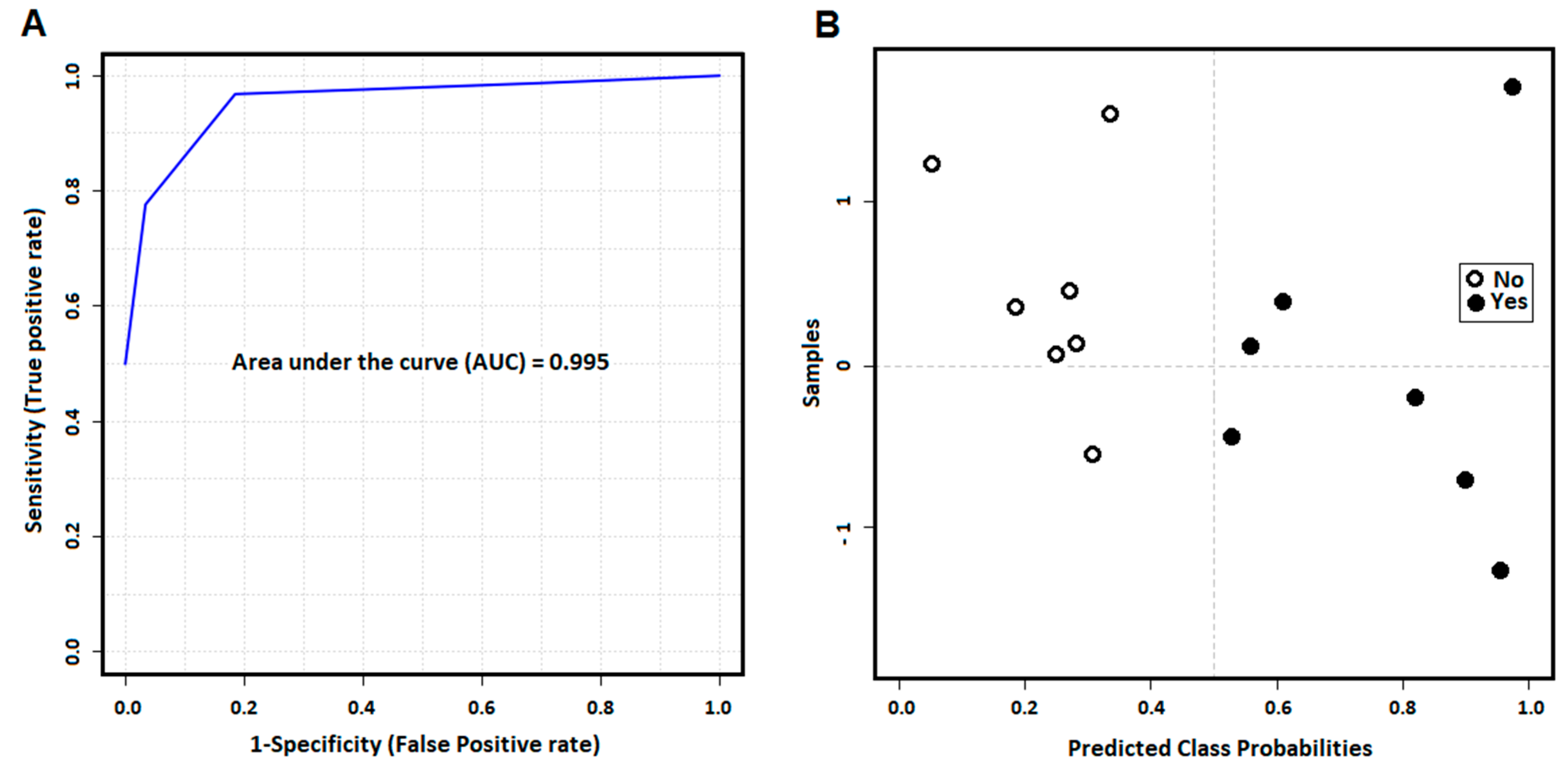
3.3. The More Important Variables for Class Separation Can Be Used to Build Models for GVHD Association
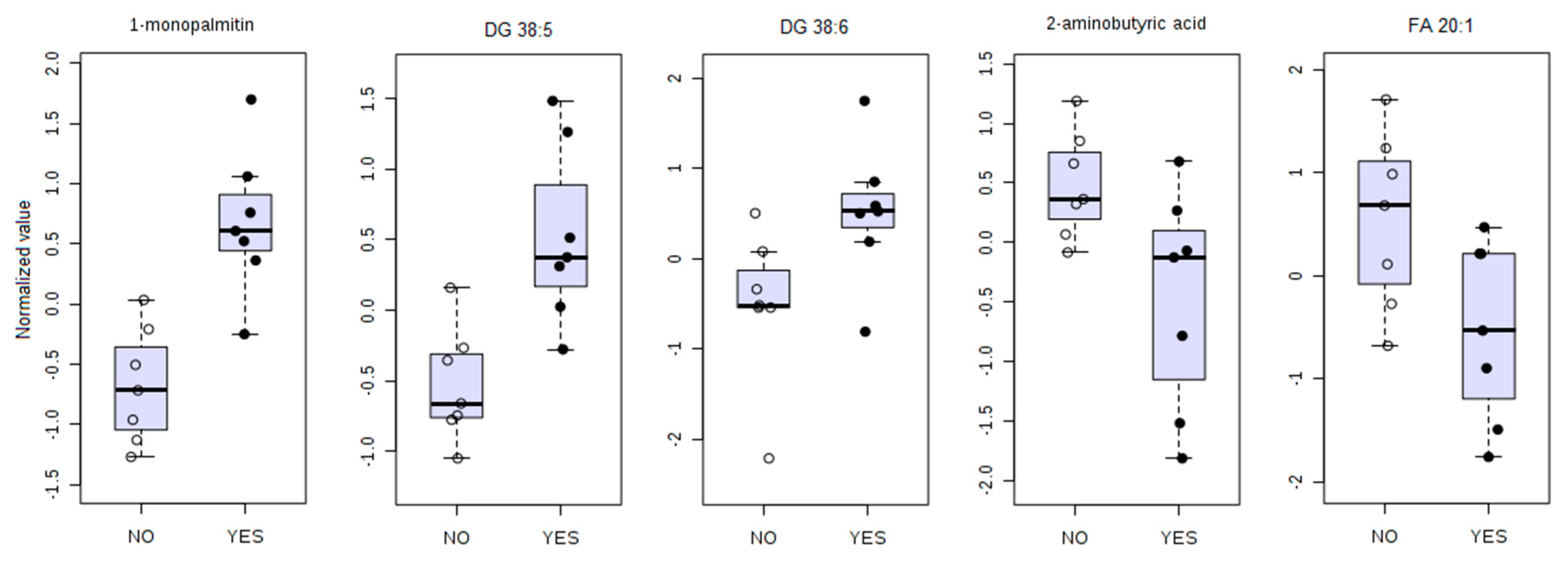
3.4. Univariate ROC Curve Analysis Finds Potential Biomarkers of GVHD With Plasmatic Data Pre-Transplant
4. Discussion
5. Limitations
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jones, N.; Cronin, J.G.; Dolton, G.; Panetti, S.; Schauenburg, A.J.; Galloway, S.A.E.; Sewell, A.K.; Cole, D.K.; Thornton, C.; Francis, N.J. Metabolic Adaptation of Human CD4+ and CD8+ T-Cells to T-Cell Receptor-Mediated Stimulation. Front. Immunol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Cretenet, G.; Clerc, I.; Matias, M.; Loisel, S.; Craveiro, M.; Oburoglu, L.; Kinet, S.; Mongellaz, C.; Dardalhon, V.; Taylor, N. Cell surface Glut1 levels distinguish human CD4 and CD8 T lymphocyte subsets with distinct effector functions. Sci. Rep. 2016, 6, 24129. [Google Scholar] [CrossRef] [PubMed]
- Martínez, C.; Millán, O.; Rovira, M.P.; Fernández-Avilés, F.; López, A.; Suárez-Lledó, M.; Carreras, E.; Urbano-Ispízua, Á.; Brunet, M. Harmacodynamics of T cell function for monitoring pharmacologic immunosuppression after allogeneic hematopoietic stem cell transplantation. Int. J. Hematol. 2017, 105, 497–505. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.D.; Chatterjee, S.; Haarberg, K.M.; Wu, Y.; Bastian, D.; Heinrichs, J.; Fu, J.; Daenthanasanmak, A.; Schutt, S.; Shrestha, S.; et al. Metabolic reprogramming of alloantigen-activated T cells after hematopoietic cell transplantation. J. Clin. Investig. 2016, 126, 1337–1352. [Google Scholar] [CrossRef] [PubMed]
- Shinjo, Y.; Makide, K.; Satoh, K.; Fukami, F.; Inoue, A.; Kano, K.; Otani, Y.; Ohwada, T.; Aoki, J. Lysophosphatidylserine suppresses IL-2 production in CD4 T cells through LPS3/GPR174. Biochem. Biophys. Res. Commun. 2017, 494, 332–338. [Google Scholar] [CrossRef] [PubMed]
- Bai, A.; Guo, Y. Acid sphingomyelinase mediates human CD4+ T-cell signaling: Potential roles in T-cell responses and diseases. Cell Death Dis. 2017, 8, e2963. [Google Scholar] [CrossRef]
- Rotolo, J.A.; Stancevic, B.; Lu, S.X.; Zhang, J.; Suh, D.; King, C.G.; Kappel, L.W.; Murphy, G.F.; Liu, C.; Fuks, Z.; et al. Cytolytic T cells induce ceramide-rich platforms in target cell membranes to initiate graft-versus-host disease. Blood 2009, 114, 3693–3706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lund, S.J.; Portillo, A.; Cavagnero, K.; Baum, R.E.; Naji, L.H.; Badrani, J.H.; Mehta, A.; Croft, M.; Broide, D.H.; Doherty, T.A. Leukotriene C4 Potentiates IL-33–Induced Group 2 Innate Lymphoid Cell Activation and Lung Inflammation. J. Immunol. 2017, 199, 1096–1104. [Google Scholar] [CrossRef] [PubMed]
- Raha, S.; Raud, B.; Oberdörfer, L.; Castro, C.N.; Schreder, A.; Freitag, J.; Longerich, T.; Lochner, M.; Sparwasser, T.; Berod, L.; et al. Disruption of de novo fatty acid synthesis via acetyl-CoA carboxylase 1 inhibition prevents acute graft-versus-host disease. Eur. J. Immunol. 2016, 46, 2233–2238. [Google Scholar] [CrossRef]
- Chiaranunt, P.; Ferrara, J.L.; Byersdorfer, C.A. Rethinking the paradigm: How comparative studies on fatty acid oxidation inform our understanding of T cell metabolism. Mol. Immunol. 2015, 68, 564–574. [Google Scholar] [CrossRef]
- Byersdorfer, C.A.; Tkachev, V.; Opipari, A.W.; Goodell, S.; Swanson, J.; Sandquist, S.; Glick, G.D.; Ferrara, J.L.M. Effector T cells require fatty acid metabolism during murine graft-versus-host disease. Blood 2013, 122, 3230–3237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Najar, M.; Raicevic, G.; Boufker, H.I.; Kazan, H.F.; De Bruyn, C.; Meuleman, N.; Bron, D.; Toungouz, M.; Lagneaux, L. Mesenchymal stromal cells use PGE2 to modulate activation and proliferation of lymphocyte subsets: Combined comparison of adipose tissue, Wharton’s Jelly and bone marrow sources. Cell. Immunol. 2010, 264, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Highfill, S.L.; Kelly, R.M.; O’Shaughnessy, M.J.; Zhou, Q.; Xia, L.; Panoskaltsis-Mortari, A.; Taylor, P.A.; Tolar, J.; Blazar, B.R. Multipotent adult progenitor cells can suppress graft-versus-host disease via prostaglandin E2 synthesis and only if localized to sites of allopriming. Blood 2009, 114, 693–701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Sun, X.; Kuang, X.; Liao, Y.; Li, H.; Luo, D. Mesenchymal stem cells suppress CD8+T cell-mediated activation by suppressing natural killer group 2, member D protein receptor expression and secretion of prostaglandin E2, indoleamine 2, 3-dioxygenase and transforming growth factor-β. Clin. Exp. Immunol. 2014, 178, 516–524. [Google Scholar] [CrossRef] [PubMed]
- Or, R.; Gesundheit, B.; Resnick, I.; Bitan, M.; Avraham, A.; Avgil, M.; Sacks, Z.; Shapira, M.Y. Sparing Effect by Montelukast Treatment for Chronic Graft Versus Host Disease: A Pilot Study. Transplantation 2007, 83, 577–581. [Google Scholar] [CrossRef] [PubMed]
- Bolanos-Meade, J.; Vogelsang, G.B. Montelukast in Chronic Graft-Versus-Host Disease: A Breath of Fresh Air? Transplantation 2007, 83, 527–528. [Google Scholar] [CrossRef] [PubMed]
- Reading, J.L.; Vaes, B.; Hull, C.; Sabbah, S.; Hayday, T.; Wang, N.S.; DiPiero, A.; A Lehman, N.; Taggart, J.M.; Carty, F.; et al. Suppression of IL-7-dependent Effector T-cell Expansion by Multipotent Adult Progenitor Cells and PGE2. Mol. Ther. 2015, 23, 1783–1793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takatsuka, H.; Takemoto, Y.; Yamada, S.; Wakae, T.; Mori, A.; Okada, M.; Iwata, N.; Okamoto, T.; Kanamaru, A.; Kakishita, E. Oral eicosapentaenoic acid for acute colonic graft-versus-host disease after bone marrow transplantation. Drugs Under Exp. Clin. Res. 2002, 28, 121–125. [Google Scholar]
- Takatsuka, H.; Takemoto, Y.; Iwata, N.; Suehiro, A.; Hamano, T.; Okamoto, T.; Kanamaru, A.; Kakishita, E. Oral eicosapentaenoic acid for complications of bone marrow transplantation. Bone Marrow Transplant. 2001, 28, 769–774. [Google Scholar] [CrossRef] [Green Version]
- Cutler, C.S.; Shoemaker, D.; Ballen, K.K.; Robbins, D.; Desponts, C.; Kao, G.S.; Chen, Y.B.A.; Dey, B.R.; McAfee, S.L.; Alyea, E.P.; et al. FT1050 (16-dimethyl Prostaglandin E2)-Enhanced Umbilical Cord Blood Accelerates Hematopoietic Engraftment After Reduced Intensity Conditioning and Double Umbilical Cord Blood Transplantation. Blood 2011, 118, 653. [Google Scholar]
- Guerrettaz, L.; Sahaf, N.; Mitchell, L.; Lynn, C.; Raynel, S.; Le, T.; Foster, H.; Truong, C.; Rezner, B.; Robbins, D.; et al. Ex Vivo Modulation of Donor Cells Results in Enhanced Survival and Reduced Gvhd Mortality. Blood 2015, 126, 1884. [Google Scholar]
- Kumar, N.G.; Contaifer, D.; Baker, P.R.; Ekroos, K.; Jefferson, K.K.; Wijesinghe, D.S. Untargeted lipidomic analysis to broadly characterize the effects of pathogenic and non-pathogenic staphylococci on mammalian lipids. PLoS ONE 2018, 13, e0206606. [Google Scholar]
- Contaifer, D.; Carl, D.E.; Warncke, U.O.; Martin, E.J.; Mohammed, B.M.; Van Tassell, B.; Brophy, D.F.; Chalfant, C.E.; Wijesinghe, D.S. Unsupervised analysis of combined lipid and coagulation data reveal coagulopathy subtypes among dialysis patients. J. Lipid Res. 2016, 58, 586–599. [Google Scholar] [CrossRef] [PubMed]
- Jayaraman, S.P.; Anand, R.J.; De Antonio, J.H.; Mangino, M.; Aboutanos, M.B.; Kasirajan, V.; Ivatury, R.R.; Valadka, A.B.; Glushakova, O.; Hayes, R.L.; et al. Metabolomics and Precision Medicine in Trauma: The State of the Field. Shock Augusta Ga 2017, 50, 5–13. [Google Scholar] [CrossRef] [PubMed]
- Hoeferlin, L.A.; Huynh, Q.K.; Mietla, J.A.; Sell, S.A.; Tucker, J.; Chalfant, C.E.; Wijesinghe, D.S. The Lipid Portion of Activated Platelet-Rich Plasma Significantly Contributes to Its Wound Healing Properties. Adv. Wound Care 2015, 4, 100–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wijesinghe, D.S.; Chalfant, C.E. Systems-Level Lipid Analysis Methodologies for Qualitative and Quantitative Investigation of Lipid Signaling Events During Wound Healing. Adv. Wound Care 2013, 2, 538–548. [Google Scholar] [CrossRef]
- Xia, J.; Broadhurst, D.I.; Wilson, M.; Wishart, D.S. Translational biomarker discovery in clinical metabolomics: An introductory tutorial. Metabolomics 2013, 9, 280–299. [Google Scholar] [CrossRef]
- Ali, A.M.; DiPersio, J.F.; Schroeder, M.A. The role of biomarkers in the diagnosis and risk stratification of Acute Graft vs. Host Disease (aGvHD): A systematic review. Biol. Blood Marrow Transplant. 2016, 22, 1552–1564. [Google Scholar] [CrossRef]
- Stikvoort, A.; Chen, Y.; Rådestad, E.; Törlén, J.; Lakshmikanth, T.; Björklund, A.; Mikes, J.; Achour, A.; Gertow, J.; Sundberg, B.; et al. Combining Flow and Mass Cytometry in the Search for Biomarkers in Chronic Graft-versus-Host Disease. Front. Immunol. 2017. [Google Scholar] [CrossRef]
- Paczesny, S.; Levine, J.E.; Braun, T.M.; Ferrara, J.L.M. Plasma Biomarkers in Graft-versus-Host Disease: A New Era? Biol. Blood Marrow Transplant. 2009, 15, 33–38. [Google Scholar] [CrossRef] [Green Version]
- Contaifer, D., Jr.; Buckley, L.F.; Wohlford, G.; Kumar, N.G.; Morriss, J.M.; Ranasinghe, A.D.; Carbone, S.; Canada, J.M.; Trankle, C.; Abbate, A.; et al. Metabolic modulation predicts heart failure tests performance. PLoS ONE 2019, 14, e0218153. [Google Scholar]
- Alborghetti, M.R.; Correa, M.E.P.; Whangbo, J.; Shi, X.; Aricetti, J.A.; Da Silva, A.A.; Miranda, E.C.M.; Sforca, M.L.; Caldana, C.; Gerszten, R.E.; et al. Clinical Metabolomics Identifies Blood Serum Branched Chain Amino Acids as Potential Predictive Biomarkers for Chronic Graft vs. Host Disease. Front. Oncol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Reikvam, H.; Hatfield, K.; Bruserud, Ø. The pretransplant systemic metabolic profile reflects a risk of acute graft versus host disease after allogeneic stem cell transplantation. Metabolomics 2016, 12, 12. [Google Scholar] [CrossRef] [PubMed]
- Soga, T.; Baran, R.; Suematsu, M.; Ueno, Y.; Ikeda, S.; Sakurakawa, T.; Kakazu, Y.; Ishikawa, T.; Robert, M.; Nishioka, T.; et al. Differential Metabolomics Reveals Ophthalmic Acid as an Oxidative Stress Biomarker Indicating Hepatic Glutathione Consumption. J. Biol. Chem. 2006, 281, 16768–16776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irino, Y.; Toh, R.; Nagao, M.; Mori, T.; Honjo, T.; Shinohara, M.; Tsuda, S.; Nakajima, H.; Satomi-Kobayashi, S.; Shinke, T.; et al. 2-Aminobutyric acid modulates glutathione homeostasis in the myocardium. Sci. Rep. 2016, 6, 36749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frischer, H.; Ahmad, T. Severe generalized glutathione reductase deficiency after antitumor chemotherapy with BCNU [1,3-bis(chloroethyl)-1-nitrosourea]. J. Lab. Clin. Med. 1977, 89, 1080–1091. [Google Scholar] [PubMed]
- Babson, J.R.; Reed, D.J. Inactivation of glutathione reductase by 2-chloroethyl nitrosourea-derived isocyanates. Biochem. Biophys. Res. Commun. 1978, 83, 754–762. [Google Scholar] [CrossRef]
- Buck, M.D.; Pearce, E.L.; O’Sullivan, D.; O’Sullivan, D. T cell metabolism drives immunity. J. Exp. Med. 2015, 212, 1345–1360. [Google Scholar] [CrossRef] [Green Version]
- Mak, T.W.; Grusdat, M.; Duncan, G.S.; Dostert, C.; Nonnenmacher, Y.; Cox, M.; Binsfeld, C.; Hao, Z.; Brüstle, A.; Itsumi, M.; et al. Glutathione Primes T Cell Metabolism for Inflammation. Immunity 2017, 46, 675–689. [Google Scholar] [CrossRef] [Green Version]
- Leslie, K.A.; Rasheed, M.; Sabo, R.T.; Roberts, C.C.; Toor, A.A.; Reed, J. Reconstituting donor T cells increase their biomass following hematopoietic stem cell transplantation. Analyst 2018, 143, 2479–2485. [Google Scholar] [CrossRef]
- Calder, P.C. Omega-3 polyunsaturated fatty acids and inflammatory processes: Nutrition or pharmacology? Br. J. Clin. Pharmacol. 2013, 75, 645–662. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C. Omega-3 fatty acids and inflammatory processes: From molecules to man. Biochem. Soc. Trans. 2017, 45, 1105–1115. [Google Scholar] [CrossRef] [PubMed]
- Ntambi, J.M. Regulation of stearoyl-CoA desaturase by polyunsaturated fatty acids and cholesterol. J. Lipid Res. 1999, 40, 1549–1558. [Google Scholar] [PubMed]
- Di Meo, S.; Reed, T.T.; Venditti, P.; Victor, V.M. Role of ROS and RNS Sources in Physiological and Pathological Conditions. Oxidative Med. Cell. Longev. 2016, 2016, 1–44. [Google Scholar] [CrossRef] [PubMed]
- Sevastou, I.; Kaffe, E.; Mouratis, M.A.; Aidinis, V. Lysoglycerophospholipids in chronic inflammatory disorders: The PLA2/LPC and ATX/LPA axes. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2013, 1831, 42–60. [Google Scholar] [CrossRef] [PubMed]
- Chiurchiù, V.; Leuti, A.; Maccarrone, M. Bioactive Lipids and Chronic Inflammation: Managing the Fire Within. Front. Immunol. 2018. [Google Scholar] [CrossRef]
- Murray, M.; Hraiki, A.; Bebawy, M.; Pazderka, C.; Rawling, T. Anti-tumor activities of lipids and lipid analogues and their development as potential anticancer drugs. Pharmacol. Ther. 2015, 150, 109–128. [Google Scholar] [CrossRef]
- Beloribi-Djefaflia, S.; Vasseur, S.; Guillaumond, F. Lipid metabolic reprogramming in cancer cells. Oncogenesis 2016, 5, e189. [Google Scholar] [CrossRef]
- Wang, Q.J. PKD at the crossroads of DAG and PKC signaling. Trends Pharmacol. Sci. 2006, 27, 317–323. [Google Scholar] [CrossRef]
- Storz, P.; Toker, A. Protein kinase D mediates a stress-induced NF-κB activation and survival pathway. EMBO J. 2003, 22, 109–120. [Google Scholar] [CrossRef]
- The Serine Hydrolase ABHD6 Is a Critical Regulator of the Metabolic Syndrome: Cell Reports. Available online: http://www.cell.com/cell-reports/fulltext/S2211-1247(13)00507-X?_returnURL=http%3A%2F%2Flinkinghub.elsevier.com%2Fretrieve%2Fpii%2FS221112471300507X%3Fshowall%3Dtrue (accessed on 6 February 2018).
- Alhouayek, M.; Masquelier, J.; Cani, P.D.; Lambert, D.M.; Muccioli, G.G. Implication of the anti-inflammatory bioactive lipid prostaglandin D2-glycerol ester in the control of macrophage activation and inflammation by ABHD6. Proc. Natl. Acad. Sci. 2013, 110, 17558–17563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, F.; Fei, X.; Xu, J.; Ji, C. An unannotated α/β hydrolase superfamily member, ABHD6 differentially expressed among cancer cell lines. Mol. Biol. Rep. 2009, 36, 691–696. [Google Scholar] [CrossRef] [PubMed]
- Grüner, B.M.; Schulze, C.J.; Yang, D.; Ogasawara, D.; Dix, M.M.; Rogers, Z.N.; Chuang, C.H.; McFarland, C.D.; Chiou, S.H.; Brown, J.M.; et al. An in vivo multiplexed small-molecule screening platform. Nat. Methods 2016, 13, 883–889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poursharifi, P.; Madiraju, S.R.M.; Prentki, M. Monoacylglycerol signalling and ABHD6 in health and disease. Diabetes Obes. Metab. 2017, 19, 76–89. [Google Scholar] [CrossRef] [Green Version]
- Chiurchiù, V.; Battistini, L.; Maccarrone, M. Endocannabinoid signalling in innate and adaptive immunity. Immunology 2015, 144, 352–364. [Google Scholar] [CrossRef] [PubMed]
- Cabral, G.A.; Griffin-Thomas, L. Emerging Role of the CB2 Cannabinoid Receptor in Immune Regulation and Therapeutic Prospects. Expert Rev. Mol. Med. 2009, 11, e3. [Google Scholar] [CrossRef]
- Bacchetti, P.; Deeks, S.G.; McCune, J.M. Breaking Free of Sample Size Dogma to Perform Innovative Translational Research. Sci. Transl. Med. 2011, 3, 87ps24. [Google Scholar] [CrossRef] [PubMed]
- Xi, B.; Gu, H.; Baniasadi, H.; Raftery, D. Statistical Analysis and Modeling of Mass Spectrometry-Based Metabolomics Data. Breast Cancer 2014, 1198, 333–353. [Google Scholar] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Donor Type | Age | Gender | Race | BMI | Prep Regimen; GVHD Prophylaxis | Disease | GVHD (Grade/Severity) | GVHD Organs/System |
|---|---|---|---|---|---|---|---|---|
| MRD | 60 | Female | C | 21.41 | Bu/Cy; Cyls/MTX | MDS | Acute (1) | Skin, liver, GI |
| URD | 50 | Female | C | 24.42 | Bu/Cy; Tac/MMF | AML * | Acute (4) | Skin, GI |
| URD | 31 | Female | C | 34.99 | Bu/Cy; Tac/MTX | AML | Chronic (mod) | Skin, GI |
| URD | 40 | Female | C | 42.08 | TBI/Cy; Tac/MTX | ALL | Acute (2) | GI |
| URD | 57 | Male | C | 22.09 | Bu/Flu; Tac/MMF | MDS | Acute (2) | Skin |
| URD | 50 | Male | C | 26.13 | Flu/Mel; Tac/MTX | ALL | Acute (1) | Skin |
| MRD | 59 | Male | C | 21.80 | Bu/Flu; Cyls/MMF | MDS | Chronic (mod) | Oral, GI |
| URD | 50 | Female | C | 28.85 | Bu/Cy; Tac/MTX | CML & | no | - |
| MRD | 63 | Female | C | 27.81 | Bu/Flu; Cyls/MTX | ET # | no | - |
| URD | 66 | Male | C | 29.45 | Flu/Mel; Tac/MMF | MCL | no | - |
| auto | 48 | Female | C | 17.60 | BEAM; n/a | PTCL | n/a | - |
| auto | 35 | Female | AA | 25.04 | Mel 200; n/a | MM | n/a | - |
| auto | 44 | Male | AA | 45.22 | BEAM; n/a | NHL | n/a | - |
| auto | 40 | Male | AA | 51.22 | BEAM; n/a | ALL | n/a | - |
| Class | Predictors | VIP | GVHD | |
|---|---|---|---|---|
| NO | YES | |||
| Alpha-amino acid | 2-aminobutyric acid | 1.80 | ||
| Monosaccharide | Hexose | 2.67 | ||
| Monoacylglycerol | 1-monoolein | 2.11 | ||
| 1-monopalmitin | 2.54 | |||
| Diacylglycerol | DG 38:5 | 1.97 | ||
| DG 38:6 | 1.91 | |||
| Fatty acid | FA 14:1 | 1.90 | ||
| FA 16:1 | 2.08 | |||
| FA 18:1 | 1.92 | |||
| FA 19:1 | 2.09 | |||
| FA 20:1 | 2.01 | |||
| FA 20:3 | 2.07 | |||
| Lysophosphatidylcholine | LPC 14:0 | 2.79 | ||
| LPC 20:0 | 2.46 | |||
| Phosphatidylcholine | PC 28:0 | 2.29 | ||
| PC 14:0/16:1 | 1.95 | |||
| PC 16:0/18:3 | 1.95 | |||
| Phosphatidylethanolamine | PE 16:0/18:1 | 2.02 | ||
| PE 18:0/22:5 | 2.23 | |||
| Plasmenyl-ethanolamine | PE (p-34:1) or PE (o-34:2) | 1.75 | ||
| Predictor | AUC | p-Value | Sensitivity | Specificity | LR+ | LR− |
|---|---|---|---|---|---|---|
| 1-monopalmitin | 0.96 (0.82–1.00) | 0.0005 | 0.86 (0.71–1.00) | 1.00 (1.00–1.00) | infinity | 0.14 |
| DG 38:5 | 0.96 (0.80–1.00) | 0.003 | 0.86 (0.57–1.00) | 0.86 (0.64–1.00) | 6.0 | 0.17 |
| DG 38:6 | 0.86 (0.57–1.00) | 0.035 | 0.86 (0.43–1.00) | 0.86 (0.64–1.00) | 6.0 | 0.17 |
| 2-aminobutyric acid | 0.86 (0.61–1.00) | 0.029 | 0.86 (0.57–1.00) | 0.71 (0.29–1.00) | 3.0 | 0.20 |
| FA 20:1 | 0.82 (0.49–0.97) | 0.039 | 1.00 (1.00–1.00) | 0.57 (0.29–0.86) | 2.3 | 0.00 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Contaifer, D., Jr.; Roberts, C.H.; Kumar, N.G.; Natarajan, R.; Fisher, B.J.; Leslie, K.; Reed, J.; Toor, A.A.; Wijesinghe, D.S. A Preliminary Investigation towards the Risk Stratification of Allogeneic Stem Cell Recipients with Respect to the Potential for Development of GVHD via Their Pre-Transplant Plasma Lipid and Metabolic Signature. Cancers 2019, 11, 1051. https://doi.org/10.3390/cancers11081051
Contaifer D Jr., Roberts CH, Kumar NG, Natarajan R, Fisher BJ, Leslie K, Reed J, Toor AA, Wijesinghe DS. A Preliminary Investigation towards the Risk Stratification of Allogeneic Stem Cell Recipients with Respect to the Potential for Development of GVHD via Their Pre-Transplant Plasma Lipid and Metabolic Signature. Cancers. 2019; 11(8):1051. https://doi.org/10.3390/cancers11081051
Chicago/Turabian StyleContaifer, Daniel, Jr., Catherine H. Roberts, Naren Gajenthra Kumar, Ramesh Natarajan, Bernard J. Fisher, Kevin Leslie, Jason Reed, Amir A. Toor, and Dayanjan S. Wijesinghe. 2019. "A Preliminary Investigation towards the Risk Stratification of Allogeneic Stem Cell Recipients with Respect to the Potential for Development of GVHD via Their Pre-Transplant Plasma Lipid and Metabolic Signature" Cancers 11, no. 8: 1051. https://doi.org/10.3390/cancers11081051