1. Introduction
Cancer vaccines based on plasmid DNA have been proved as good alternative to the more traditional subunit vaccines like recombinant proteins and peptides. Nevertheless, their limited potency is one of the “core” issues of DNA vaccination [
1].
To increase the efficacy of genetic vaccines, several studies are focused on delivery systems. They envisage compounds and technical strategies helping to deliver the DNA vaccines into the right cell compartment for obtaining maximum potency together with improved safety profile.
Hyaluronidases (Hyals) or hyaluronoglucosaminidases are a family of enzymes degrading hyaluronic acid (HA), also called hyaluronan, which is an important component of the extracellular matrix (ECM) of the connective and hepitelial tissues. Hyals hydrolyze the hexosaminidic β (1–4) linkages between
N-acetyl-
D-glucosamine and
D-glucuronic acid residues in HA molecule and release HA fragments [
2]. The vertebrate Hyals generate a various range of HA oligomers [
3]. Some bacteria, such as
Staphylococcus aureus,
Streptococcus pyogenes, and
Clostridium perfringens, produce Hyals as a virulence factor, that facilitates toxins diffusion and bacterial motility through the host’s tissues [
4,
5]. It is also well known that HA and HA-binding proteins play a role in inflammation, tissue injury and repair through regulating inflammatory cell recruitment, release of inflammatory cytokines and cell migration [
6].
Hyal is proven to greatly increase the dispersion and absorption of subcutaneously injected large molecules. It is used as an adjunct to nonpharmacological management of extravasation of selected antineoplastic agents, and in the management of extravasation contrast media [
7,
8]. Bacterial [
9], Bovine (Amphadase
TM, Hydase
TM) [
10,
11], ovine (Vitrase
TM) [
12], and human recombinant (Hylenex
TM) [
13] Hyal preparations have been developed over the years for wider uses in the clinical practice.
In the years 1940–1950, Hyal became popular in the vaccination field as a “spreading factor” used for a better diffusion of traditional vaccines at the site of injection [
14,
15,
16]. Recently, the positive effects of Hyal were shown in gene therapy targeting neurons [
17], and in pre-clinical protocols based on Gene Electro-Transfer (GET) [
18,
19,
20,
21].
GET is a method that allows the targeted transfer of plasmid DNA into cells in a tissue and promotes efficient gene expression. It results from the direct application of electric field pulses, avoiding the use of viral vectors. The application of controlled electric pulses induces a transient permeabilisation of the plasma membrane and the uptake of the negatively charged plasmid DNA inside the cell [
22,
23]. GET has been applied in vivo since 1998 [
24] in different tissues and organs [
25,
26,
27]. During the last decade, several clinical trials based on GET were initiated [
28]. Many of them are focused in developing anti-tumor vaccine therapy. In a phase I clinical trial on patients with metastatic melanoma, 10% of the patients treated by GET of a plasmid encoding the cytokine IL-12 exhibited a complete regression of all metastases (treated and untreated) while 42% displayed a stable disease or partial regression [
29].
The strength of GET is to induce a massive immune response. The transfected plasmid encoding specific antigens is uptaken by muscular cells and by professional antigen-presenting-cells (APC) present in the pulsed tissue. These APC migrate to the draining lymph nodes where they activate the adaptive immune response through stimulation of CD4+ and CD8+ T cells. Moreover, the delivered electric field pulse by itself induces a local inflammation characterized by the attraction of macrophages and dendritic cells amplifying the strength and the duration of the immune response [
30,
31,
32]. However, in cancer, because of the self-origin of the cancer cells, only few antigens associated to the tumors are efficiently immunogenic (i.e., able to trigger an effective immune response). Therefore, the development of a GET-based therapy able to improve the response of the immune system is still needed [
33].
It is known that the treatment of skeletal muscle with Hyal allows the application of lower levels of electric fields, thus reducing muscle fibres damage [
34]. Moreover, in a recent article on the effect of Hyal on mouse skeletal muscle, pretreatment with this enzyme was demonstrated to be a good enhancer of GET efficiency in gene vaccination protocols. In particular, when used in combination with pulsed electric fields, Hyal increased the release of pro-inflammatory cytokines (Interleukin-1β (IL-1β), tumor necrosis factor (TNF-α), Interleukin-6 (IL-6)) and favoured local APC infiltration in a defined time-window between day 4 and day 7 following cytokine induction [
21]. The final effect was an improved delivery of the DNA vaccine and a general enhancement in terms of treatment efficacy. These observations underline the relevance of appropriate technologies and molecules able to improve the drug delivery in GET protocols to be introduced into the clinical practice.
Bovine-derived Hyal (Amphadase
TM) [
10] and ovine-derived Hyal (Vitrase
TM) [
12] were approved by the Food and Drug Administration (FDA) in 2004; a second bovine-derived Hyal (Hydase
TM) [
11] was subsequently approved in 2005. Notwithstanding that animal-derived Hyals are regularly employed, the production of such formulations have been generally reported to be limited due to low purity, variable potency and uncertain safety. These preparations are often contaminated with proteases, immunoglobulins and other factors that increase capillary permeability and may induce IgE-mediated allergic reactions when repeatedly administrated, generally precluding their subsequent use. Importantly, many case reports of adverse reactions were published in medical journals connected with the use of animal-derived Hyals that were subsequently taken off the market [
35,
36,
37,
38]. Therefore, they appear to bring a risk for application in human patients and also preclinical protocols based on DNA vaccination by pretreatment with animal-derived Hyal and electroporation could be narrowly proposed in clinical trials.
A human recombinant Hyal (Hylenex
TM, Halozyme Therapeutics Inc., San Diego, CA, USA) has been produced by genetically engineered Chinese Hamster Ovary (CHO) cells containing a DNA plasmid encoding for a soluble fragment of human Hyal (PH20) [
13], and has been recently approved by the FDA for use as an adjuvant agent in subcutaneous fluid administration for achieving hydration, to increase the dispersion and absorption of other injected drugs and as an adjunct in subcutaneous urography for improving resorption of radiopaque agents [
39]. This product has been widely used in man, however no data were available describing its use for GET assays. However, a possible utilization of recombinant human PH20 Hyal (rHuPH20) in GET protocols, i.e., in the context of a high stimulation of the immune response, could have a limitation related to the eventual production of non-neutralizing antibodies against rHuPH20, with associated risk of cross-reactivity versus endogenous PH20 with potential long-term effects [
40,
41].
A novel Hyal, called rHyal-
sk (Fidia Farmaceutici S.p.A, Abano Terme, Padova, Italy) was produced as a recombinant protein in a non-pathogenic bacteria (GRAS product: Generally Regarded As Safe) and obtained by a new protocol of extraction [
9,
42]. rHyal-
sk is a non-glycosylated, cysteine disulfide bond free enzyme containing a novel bacterial catalytic domain with high enzymatic activity and it is characterized by a very low molecular weight (about 22 kDa), an excellent purity profile, high stability to proteolytic enzymes, at low/high pH and at high temperatures up to 70 °C. Additional properties include: long shelf-life, high performance at physiological pH and at body temperature, and it is not inhibited by human blood [
9,
42]. In addition, rHyal-
sk exhibits remarkable substrate specificity for HA [
43] and no risk of animal cross-infection.
In order to improve GET-based protocols of therapeutic plasmid injection and move rapidly to safe and efficient translational gene therapy protocols, we evaluated the efficacy and safety of this novel Hyal. Here we describe a pretreatment of murine skeletal muscle with rHyal-
sk followed by GET of plasmid DNA (coding for the fluorescent protein tdTomato) to improve the transfection efficiency of plasmids within the injected muscle. Results were compared with a pretreatment with bHyal, already known to bring a positive effect on electrotransfection [
18,
21,
44]. The evaluation of this new kind of hyaluronidase was performed both in terms of the overall levels of gene expression in the transfected muscle fibres by fluorescence imaging and morphological damage occurring in the muscle. We also investigated the potential to activate a local proinflammatory immune response in injected muscle, a crucial aspect that should be considered in the optimization of GET protocols against cancer.
3. Discussion
Several strategies have been proposed to potentiate DNA immunotherapy effectiveness in anti-cancer protocols, being plasmid DNA vaccines characterized by low immunogenicity. Chemical or physical devices have been exploited to increase DNA delivery into target organs, as the promising Gene-electro-transfer (GET) method.
Here we investigated a new kind of soluble recombinant bacterial Hyal, the rHyal-
sk, in order to assess the effect of this Hyal treatment at the cellular and biochemical levels, when injected on skeletal muscle as adjuvant of GET-based immunotherapy protocols. The rHyal-
sk is a recombinant Hyal that has a novel bacterial catalytic domain with high enzymatic activity [
42]. Compared to commercially available animal-derived hyaluronidase, rHyal-
sk is more thermostable [
9,
42] and presents higher proteolytic resistance, with activity over a broad pH spectrum [
9,
42]. In addition, it exhibits remarkable substrate specificity for HA [
43] and no risk of animal cross-infection.
In this study, we evaluated whether rHyal-sk treatment influenced the efficiency of gene uptake in the muscular fibres and how the inflammatory process was activated by the administration of GET after rHyal-sk pretreatment. We assessed the cellular events and the inflammation chemical mediators involved in the first two weeks after rHyal-sk administration with and without the combination with GET.
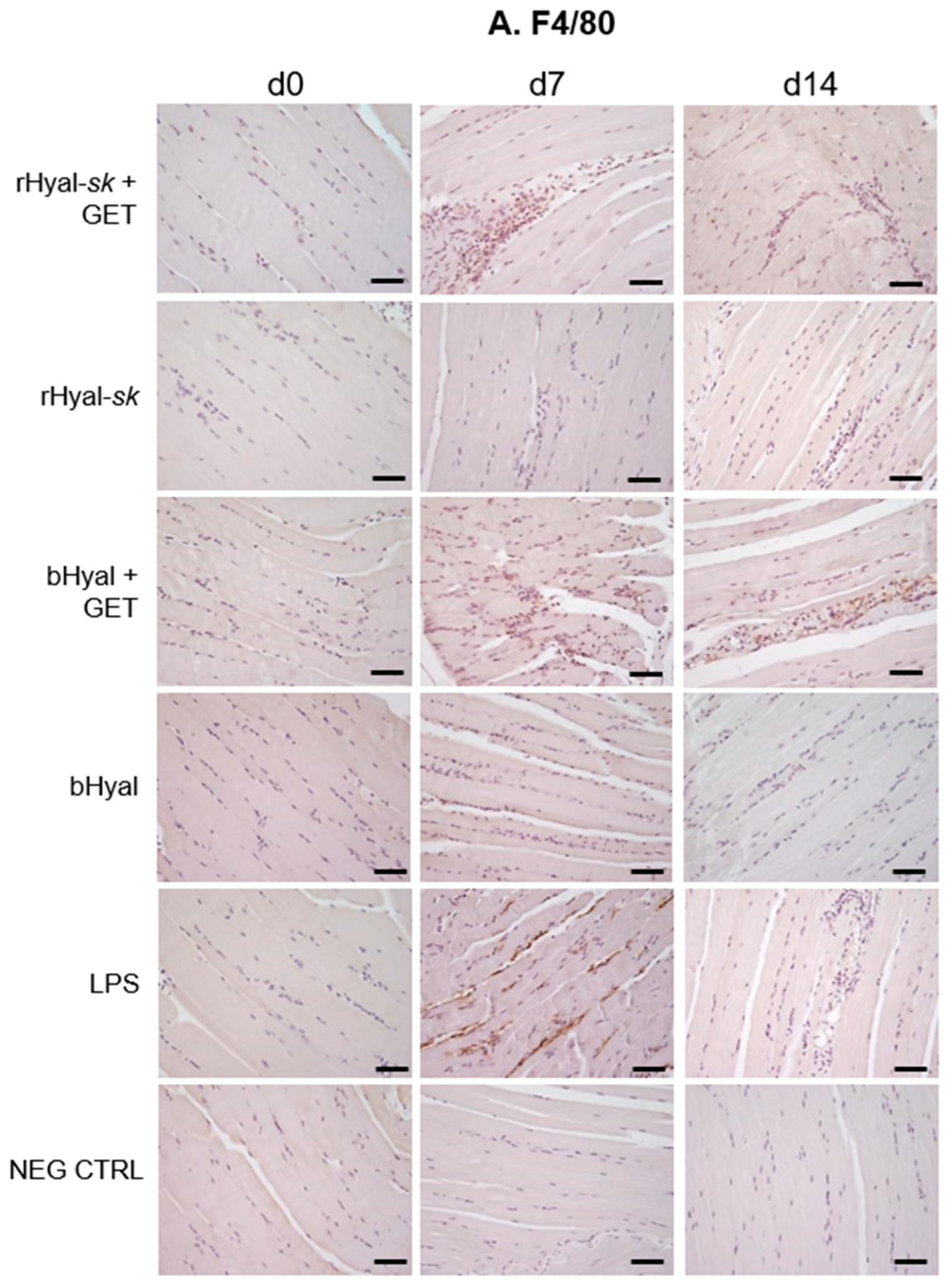
The comparative analysis of skeletal muscles morphology at three time points (3 h, 7 days, 14 days) revealed a slight tissue damage and recruitment of infiltrating monocytes and granulocytes in the early phase (7 days) after rHyal-
sk pretreatment plus GET (
Figure 2), which create a proinflammatory context useful in local antigen presentation in case of DNA vaccination protocols. The immunohistochemistry analysis (
Figure 3) demonstrated indeed that the combination of rHyal-
sk with GET favours the migration of high number of polymorphonuclear cells and APCs in comparison to rHyal-
sk alone administration. As expected, the rHyal-
sk did not induce any proinflammatory context in the absence of GET, similarly to the bHyal.
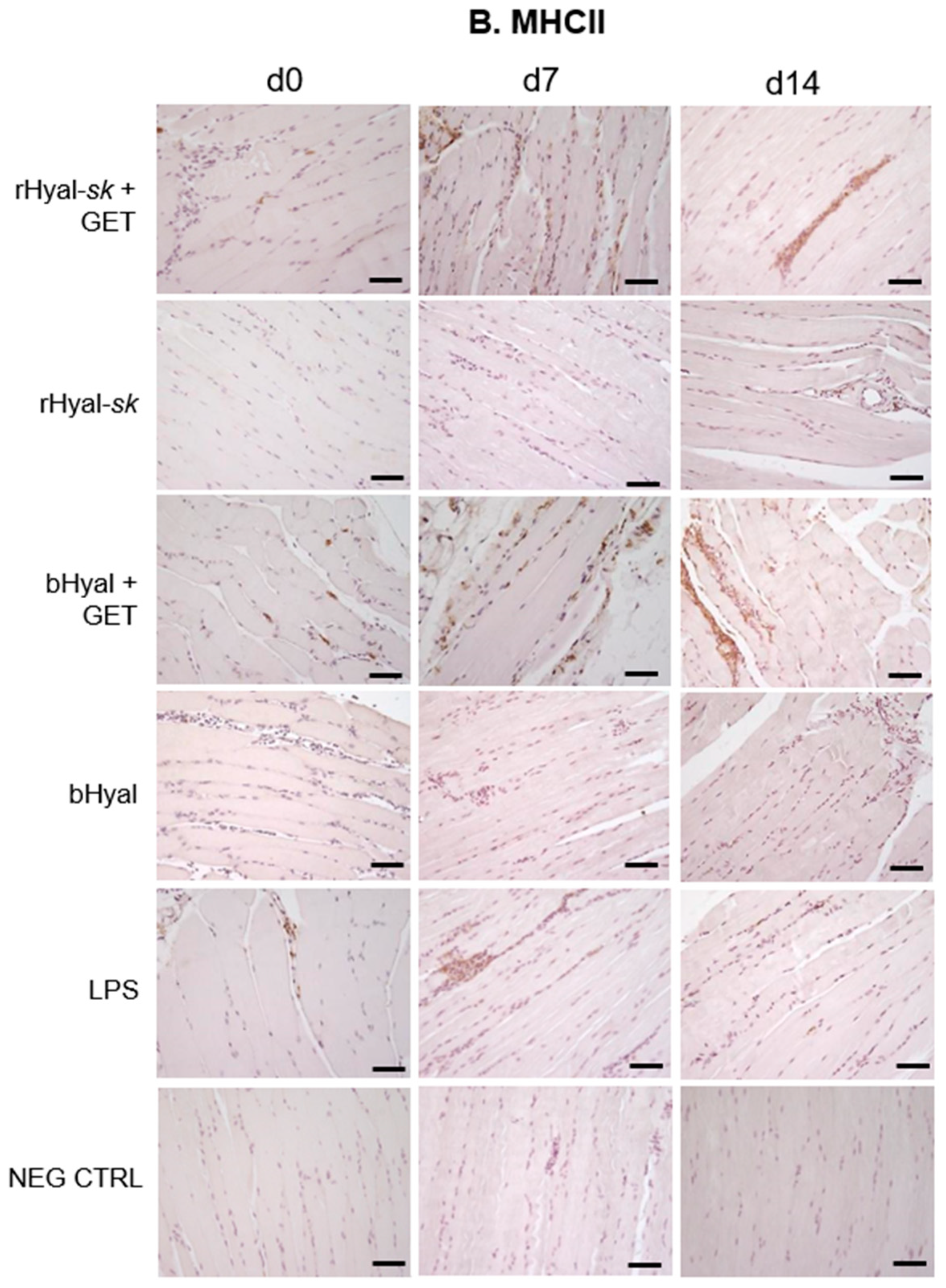
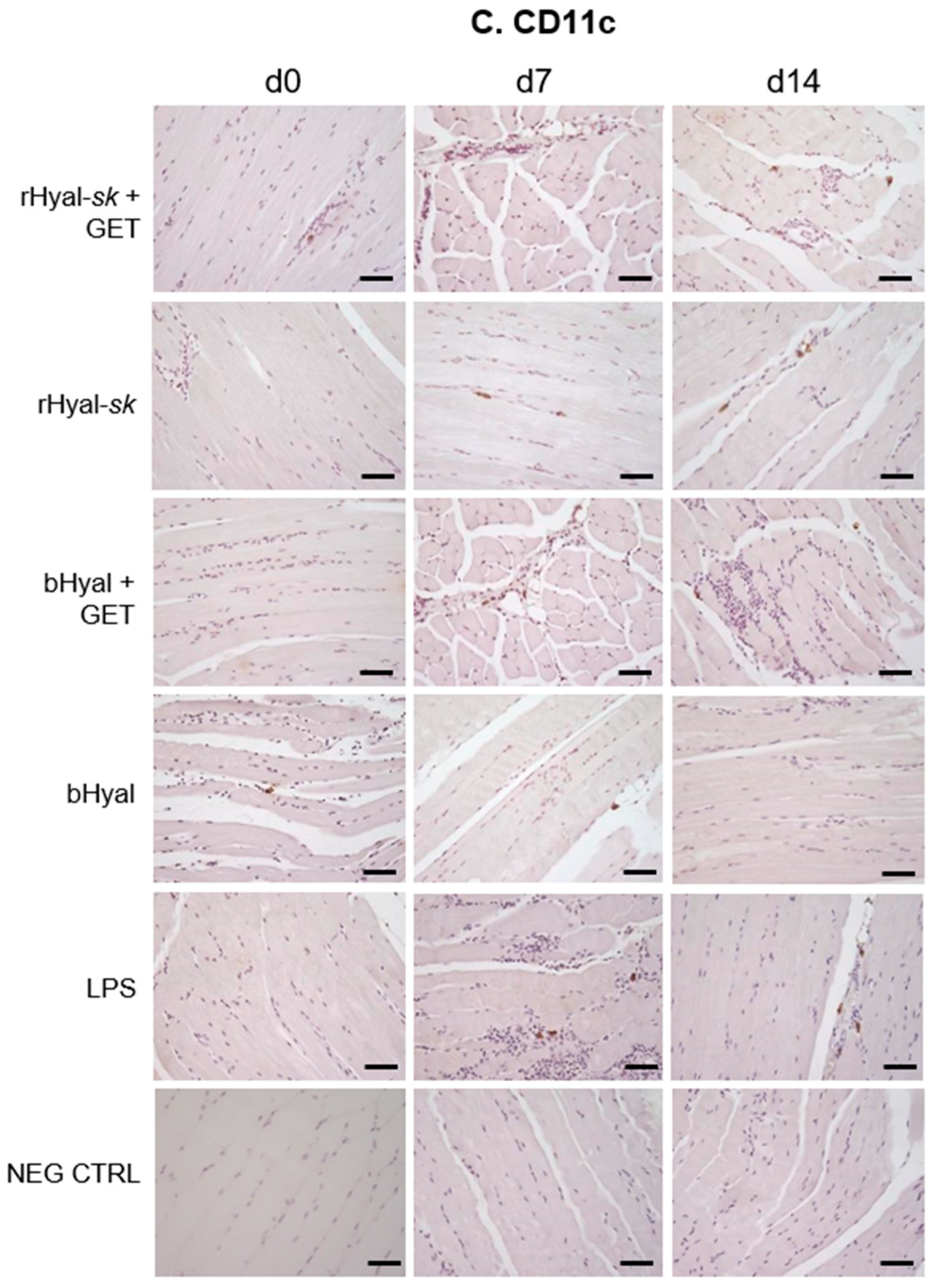
As shown in
Figure 3, GET in presence of rHyal-
sk pretreatment induced a visible infiltration of inflammatory cells 7 days after treatment. Additionally, we observed a clear cell recruitment into the muscle, mostly represented by F4/80+ macrophages and dendritic cells expressing MHC-II. A high number of such cell subpopulations into the damaged muscles was observed both at 7 and 14 days. This process was strictly linked to the substantial muscle regeneration observed at 14 days after the treatment with rHyal-
sk with GET (
Figure 2 and
Figure 3), characterized by cell proliferation and differentiation of new myogenic cells that fused to existing damaged fibres for repair. When fusion of myogenic cells was completed, the size of the newly formed myofibres increased and myonuclei moved to the periphery of the fibre [
44]. These data were in line with previous experimental observations obtained with a different type of Hyal of extraction origin [
21,
43,
47] and implied that rHyal-
sk could be used in optimized DNA immunotherapy protocols performed with GET using 14 days as the appropriate interval for DNA administration into skeletal muscle. This time is indeed required to create the proinflammatory environment necessary for APC recruitment and the myofibres regeneration fundamental for the uptake of DNA plasmid [
48].
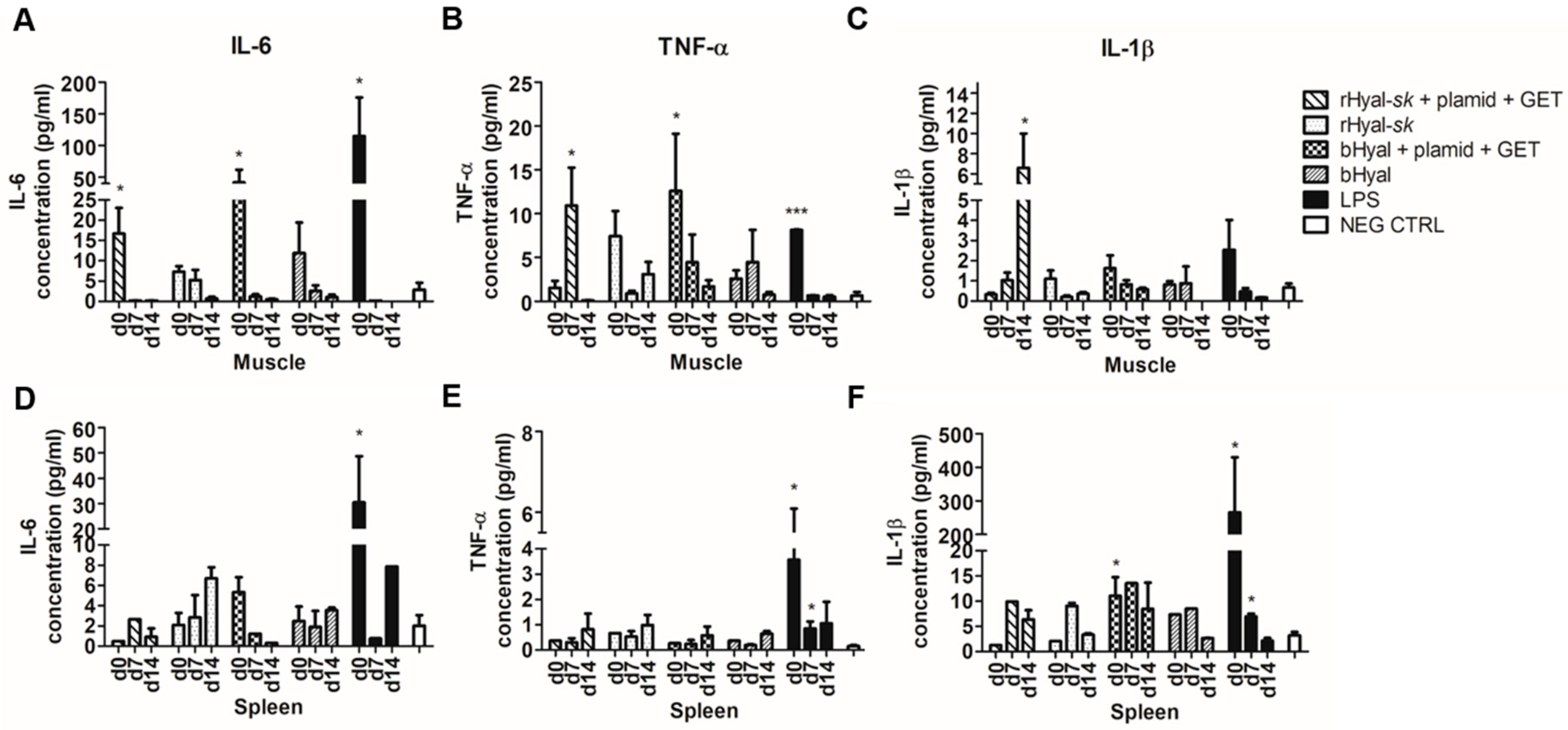
The analysis of the cytokine released upon treatment revealed an IL-6 production in muscles as early as 3 h after treatment with rHyal-
sk plus GET, demonstrating the ability of such treatment to stimulate an early cytokine production (
Figure 5). IL-6 production in muscles dropped down to the detection limit at day 7. No differences were observed between the use of rHyal-
sk or bHyal on the production of IL-6. Conversely, a different timing of TNF-α and IL-1β production was observed in muscles treated with rHyal-
sk and bHyal. TNF-α and IL-1β release in rHyal-
sk plus GET treated muscles peaked at day 7 and day 14, respectively, and 3 h after treatment on bHyal plus GET treated muscles suggesting a long lasting inflammatory response induction by the rHyal-
sk with respect to bHyal (
Figure 5). In addition, we observed that bHyal plus GET induced a rapid production of TNF-α in the first hours after treatment while rHyal-
sk plus GET induced a delayed pic of production 7 days after treatment. In view of these differences of TNF-α production kinetics, one could hypothesize that bHyal plus GET acts mainly on the innate immune response with a production mainly by macrophage and monocytes while the rHyal-
sk plus GET activates the adaptive immune response and trigger a TNF-α production from lymphocytes.
Moreover, TNF-α and IL-1β production patterns for rHyal-sk plus GET and bHyal plus GET were quite different in the muscle where and when the GET is delivered (local effect). For both cytokines, the expression was high at day 0 with the bHyal pretreatment and then decreases. A different pattern was observed with the rHyal-sk. A low level was detected at day 0 followed by a sharp increase in the following days. A final decrease is present at day 14 for the TNF-α. This is not the case with IL-1β where a high level was observed at day 14.
We should consider that the endotoxin is a major contributing factor in inflammation in Hyals partially purified from bovine testis (bHyal) [
49,
50]. In addition, these preparations showed the presence of other proteins, which also may contribute to their proinflammatory activity [
50]. Therefore, we could hypothesize that the only differences observed regard TNF-αand IL-1β production patterns for rHyal-
sk plus GET and bHyal plus GET, could be due to the presence of endotoxin and some other components in protein preparation of bHyal, which stimulated the inflammatory response in vivo. In contrast, rHyal-
sk is a hightly purified recombinant protein preparation, in fact it is free of pg amounts of endotoxin and other components contamination.
Apart from this, the treatment with rHyal-sk was able to activate an adequate response which could efficiently recruit APCs and macrophages in the site of injection for a suitable immunotherapy strategy. Importantly, cytokines release was not affected by rHyal-sk alone.
Since rHyal-
sk is a Hyal obtained by a recombinant pathway in the non-pathogenic microorganism S. koganeiensis, it has low homology with the human enzyme. Consequently, we evaluated the level of its immunogenicity in vivo. We found that when repeatedly administered subcutaneously in rats, the presence of nADA could be detected following the highest dose and remains measurable for at least one month afterward. We also investigated the immune response of rHyal-
sk when injected intramuscularly associated to GET and we did not observe any antibody titer against rHyal-
sk at day 7 (
Figure 4). More importantly, no particular toxicological findings were recorded in any of the treated animals. Therefore, while the induction of nADA could limit the use of rHyal-
sk in treatments requiring repeated administrations for long periods, this enzyme can find use as an adjuvant to potentiate the immune response of DNA electrotransfer, which needs a single injection, and in all the “single treatments” into the clinics.
Taken as a whole, our results indicate that rHyal-sk injection in skeletal muscles with the application of electric field pulses (1) induces a slight tissue damage and a recruitment of infiltrating monocytes and granulocytes, which produce a proinflammatory context useful for local antigen presentation in anti tumoral vaccination protocols; (2) is effective in terms of safety, in-take ability, immune response activation; (3) does not induce a significant production of nADA when injected once intramuscularly and associated to GET.
In view of these results, we support the adoption of rHyal-sk as a new constituent of immunotherapy protocols based on plasmid delivery, to potentiate the administration of DNA electrotransfer in the field of novel veterinary and human anti-cancer therapeutic strategies.
4. Materials and Methods
4.1. Production of rHyal-sk, a Soluble Recombinant Bacterial Hyal
rHyal-
sk (Fidia Farmaceutici, Local Unit Fidia Research Sud, Siracusa, Italy) was cloned through an expression system leading to the production of the bacterial Hyal soluble domain (aminoacids 31–247), which was identified and isolated from the
Streptomyces koganeiensis DNA. The expression system was based on a pET vector, carrying the kanamycin resistance as selectable marker, the T7 expression region, the f1 origin and containing a gene construct encoding the soluble form of bacterial Hyal. rHyal-
sk sequence data were submitted to GenBank under the accession number: KP313606. Protein 3D structure was deposited in EMBLEBI, Protein Data Bank (
http://www.ebi.ac.uk/pdbe/) under accession number: PDB ID 4UFQ.
The
Escherichia coli BL21 (DE3) competent cells were transformed with the selected vector. Following the fermentation process, the enzyme activity in the periplasmic extract from the bacterial culture was analyzed as briefly described: a pellet was produced from 100 μL of culture after centrifugation at 7500 rpm for 10 min. The cellular pellet was again suspended in 500 μL of B-PER Bacterial Protein Extraction Reagent (Pierce, Waltham, MA, USA), vortexed and centrifuged at 13,000 rpm for 10 min in order to separate the soluble proteins from the insoluble. By means of the turbidimetric assay [
51,
52], the collected supernatant was tested for the presence of Hyal activity.
Subsequently, for the production of a suitable quantity of the soluble rHyal-
sk destined specifically for this study, the recombinant bacterial Hyal was purified from the pellet via periplasmic extraction from osmotic shock and ultimately through three chromatographic steps: anion exchange (quaternary ammonium group, GE Healthcare, Chicago, IL, USA), cation exchange (carboxymethyl group, GE Healthcare), and hydrophobic interaction (phenyl sepharose, GE Healthcare). This purification sequence was followed by the analysis of purity and specificity through SDS-PAGE (12%) and enzymatic activity [
51,
52].
Contaminants of the
E. coli were analyzed by ELISA for the host protein (Cygnus Technologies, Inc., Southport, NC, USA), by Threshold System for the host DNA, and by Chromo-LAL Kinetic Chromogenic Endotoxin Testing for the endotoxin content. Enzyme potency in bulk preparations was determined by the turbidimetric assay [
51,
52] using Ph. Eur. bovine Hyal as the reference standard.
4.2. Toxicology Study and Evaluation of Anti-Drug Antibodies (ADA) Production
A 28-day toxicity study of rHyal-sk was performed in rats at Harlan Laboratories S.A. (Santa Perpetua de Mogoda, Barcelona, Spain). All animal procedures were executed in accordance with institutional guidelines for laboratory animal care, in adherence with ethical standards, and in compliance with good laboratory procedures (GLP). One hundred Wistar rats were distributed into four dose groups, each containing ten animals per sex, except for the control group (group 1) and the high dose group treated with the test item Hyal (rHyal-sk) at 10,500 U/kg (group 4), which contained a further five animals per sex for an additional 2-week treatment-free recovery period. The animals received the test item Hyal (rHyal-sk) subcutaneously every forty-eight hours for 4 weeks at the doses of 150 U/kg (Group 2), 1050 U/kg (Group 3) and 10,500 U/kg (Group 4). Control animals from Group 1 received the vehicle only, PBS, following the same regimen as the groups treated with rHyal-sk. At the end of the treatment period, all animals from the main study were necropsied and examined postmortem. The rest of the animals remained on-study, untreated, for additional 2 weeks and were necropsied and examined postmortem at the end of the recovery period. The injection sites and the following standard list of tissues were collected, processed for histology and evaluated by a veterinary pathologist: epididymides, esophagus, eyes with optic nerve, femur, heart, large and small intestine, kidneys, larynx, liver, lungs, lymph nodes (mandibular and mesenteric), lymph nodes that drain or are in contact with the injection sites (axillary and brachial), mammary gland area, ovaries, pancreas, peyer’s patches, pituitary gland, prostate and seminal vesicles, salivary glands, sciatic nerve, skeletal muscle, skin (abdominal), spinal cord, spleen, sternum (with bone marrow), stomach, testes, thymus, thyroid, tongue, trachea, urinary bladder, uterus, cervix, oviducts, vagina, and all gross lesions.
The following toxicological examination were performed in all the necroscopsied mice: mortality, histopathology, macroscopic and microscopic findings on the injection site and all the tissues listed above.
To check if there was a clear indication of an immunogenic potential of rHyal-sk, all samples which were confirmed positive for the presence of anti-drug antibodies (ADA) were further investigated in the neutralizing antibody (nAbs) assay. Samples that showed results above the respective run-specific screening Cut Point for nAbs were considered potentially positive and were further investigated in the confirmatory assay by immunocompetition for nAbs.
The determination of nAbs against rHyal-sk was conducted in 3 male and 3 female rats treated subcutaneously every forty-eight hours for a period of 4 weeks (for a total of 14 administrations): group 1 (Control)—0 U/kg (only vehicle); group 2—150 U/kg rHyal-sk; group 3—1050 U/kg rHyal-sk; group 4—10,500 U/kg rHyal-sk. ADAs were determined from samples before administration on day 1 (acclimatization), at the end of the treatment period (week 4) and two weeks after treatment (recovery). This analytical phase was conducted at Harlan Laboratories Ltd., Itingen, BL, Switzerland, under GLP-compliant conditions. The determination of nAbs against rHyal-sk in serum samples was carried out with an enzymatic assay based on the Hyaluronidase ELISA assay kit (K-6000, Echelon Biosciences, Salt Lake City, UT, USA) developed and validated by Harlan Laboratories Ltd., in order to quantify Hyal activity inhibition due to binding antibodies in rat serum (5% rat serum matrix). Quality Controls (QCs) at concentrations of 0.05 µg/mL Low Quality Controls (LQC) and 1 µg/mL High Quality Controls (HQC) were used. The acceptance criteria were fully met as optical density (OD) of HQC > LQC > Negative Controls (NC) and ODs ≥ Screening Cut Point (SCP) were obtained for the QCs of the screening assay. The used NCs met the acceptance criteria of OD < SCP. For the confirmatory assay, the immunocompetition of HQC and LQC was investigated. For all immunocompeted QCs, the immunocompetition was higher than the Confirmatory Cut Point (CCP) of 79.1% (determined during validation).
4.3. Animal Treatment with rHyal-sk Plus GET
All experiments involving 36 six-weeks-old female BALB/c mice (Charles River Laboratories International, Inc. Wilmington, MA, USA) were approved by the Ethical Committee at the University Cattolica Sacro Cuore-Roma, Italy, according to the decree Prot. pdc. CESA/A/61/2011; experimental Protocol 48 G. Mice were equally distributed in 6 groups: (1) rHyal-sk plus GET, (2) bHyal plus GET, (3) rHyal-sk alone, (4) bHyal alone, (5) LPS, (6) untreated.
In both groups 2 and 4 (reference groups) mice were injected with a bovine Hyal (bHyal) (H-4272, Sigma-Aldrich, St. Louis, MO, USA).
Ten units of rHyal-
sk (for groups 1 and 3) or bHyal (for groups 2 and 4) were suspended in 30 µL of 0.15 M sodium phosphate buffer and intramuscularly injected in the gastrocnemius muscle of both posterior hind limbs 2 h prior other treatments, as performed in the study of McMahon et al. [
18].
For GET treatment, mice were shaved after anaesthesia with a mixture of ketamine-medetomidine (Domitor) injected in the muscle of the anterior limb, just before the GET treatment. Mice previously treated with rHyal-
sk (6 mice) or bHyal (6 mice) were intramuscularly injected with pCMV-CpGfree-tdTomato (Invivogen, Toulouse, France), a 4400 bp plasmid encoding the fluorescent protein TdTomato used as a reporter of GET efficiency. An amount of 25 µg of plasmid suspended in a final volume of 25 µL of 0.15 M sodium phosphate buffer was injected in the gastrocnemius muscle of both posterior limbs. The GET was done using tweezer electrodes with terminal circular plates, applied on the gastrocnemius muscle after conductive gel application. The electrical parameters consisted of 10 square-wave pulses of 175 V to cm (voltage to electrode gap), duration 20 ms, delivered at a frequency of 1 Hz using an ECM 830 generator (Harvard Apparatus, BTX Instrument Division, Holliston, MA, USA). Electrical parameters have been decided based on previous experiments aimed to optimize the GET conditions [
21,
44,
53]. As a control of endotoxin induced inflammation, an additional group of 6 mice were injected intramuscularly with 25 µg of LPS (Sigma-Aldrich) in a volume of 25 µL. Negative controls were six untreated mice, injected only with PBS. Two mice per group were sacrificed by CO
2 asphyxiation at the following time points: 3 h (d0), 7 days (d7) and 14 days (d14) after treatment and muscles of both legs and spleens were collected and divided for tdTomato fluorescence quantification, histological examination and dosage of cytokines secretion by ELISA.
4.4. Handling of Tissue Specimens and Histological Examination
Skeletal muscles and spleens were harvested from animals treated with rHyal-sk or bHyal alone or in combination with GET, immediately frozen in dry ice and stored at −80 °C or fixed in 4% paraformaldehyde for 24 h and embedded in paraffin. Some of the frozen organs were embedded in cryostat embedding medium (Bioptica, Milan, Italy) for cryosectioning, and the remaining tissues were used for protein extraction. Histological sections of formalin-fixed paraffin-embedded (FFPE) samples were prepared using routine procedures for histopathological analyses.
Briefly, serial sections of 4 μm thickness were cut, stained with hematoxylin and eosin (Bio-Optica, Milano, Italy), dehydrated and mounted using Eukitt (O Kindler GmbH & Co., Freiburg, Germany) and air-dried before morphological analysis.
4.5. Immunohistochemistry
Immunohistochemistry was performed on 4-μm-thick FFPE tissue sections after antigen retrieval in sodium citrate buffer (10 mM sodium citrate, pH 6), for 40 min at 95 °C.
Sections were incubated in a humid chamber overnight at 4 °C with the following primary antibodies used at 1:50 dilution in blocking buffer: anti-mouse F4/80 clone BM8, anti-mouse MHC class II clone M5/114.15.2, and anti-mouse CD11c, clone N418 (eBioscience, San Diego, CA, USA). Next, slides were incubated with a biotin-labelled secondary antibody and HRP-conjugated avidin for 30 min at room temperature. Detection was achieved using a substrate/chromogen mixture (DAB) and haematoxylin counterstaining. Incubation with the primary antibody was omitted for the negative controls. The immunostained slides were observed under an optical microscope (Leitz DMR; Leica, Wetzlar, Germany), 20× magnification, and the image data were analysed using NIS FreeWare 2.10 software (Nikon, Minato, Tokyo, Japan).
4.6. tdTomato Fluorescence Analysis
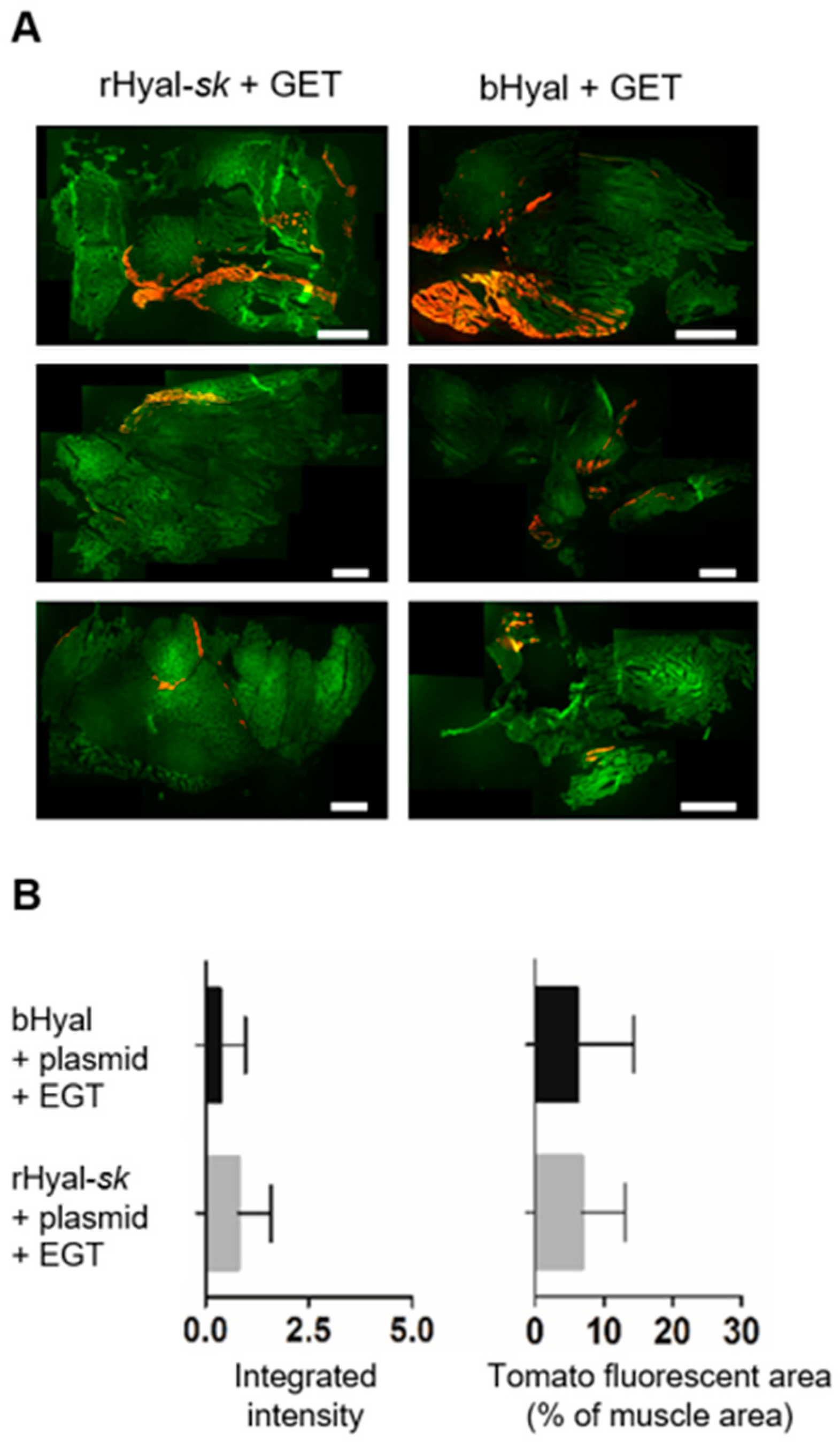
Skeletal muscles were fixed in 4% buffered formalin for 24 h and dehydrated with increasing scale of sucrose. Samples were embedded in cryostat-embedding medium (Bio-Optica, Milan, Italy) and frozen. Cryosections of 5 µm were cut with a cryostat (Leica), fixed with 4% paraformaldehyde, dried and rinsed in PBS for 10 min. Slides were stained with DAPI and mounted in Mowiol mounting medium. Microscopy was carried out using an upright Macroscopic-fluorescence microscope (Macroscope, Leica Microsystems SA, Rueil-Malmaison, France), equipped with a Cool Snap HQ Camera (Roper Scientific, Photometrics, Tucson, AZ, USA). Slides were imaged by fluorescence using appropriate filters (TdTomato: excitation filter, BP: 560/40 nm, emission filter 630/75 nm and autofluorescence: excitation filter, BP: 480/40 nm, emission filter, BP 527/30 nm). The files were stored with image acquisition software (Metavue, Metamorph, Molecular Devices, Sunnyvale, CA, USA). The files were stitched with the Adobe Photoshop software to obtain a single image of the whole muscle. Muscle area, mean and integrated fluorescence intensity were calculated with an analysis software (Image J) [
54]. After a background correction, by applying a suitable threshold on the TdTomato images, TdTomato positive area, mean and integrated fluorescence intensity were measured in the limit of this threshold. Similarly, the entire muscle area was measured in the limit of a suitable threshold defined on the autofluorescence image.
4.7. Protein Extraction
Skeletal muscles and spleens were frozen in dry ice and stored at −80 °C. Protein mass of samples was extracted as previously described [
53]. Briefly, organ samples were weighted, and 50 µL of lysis buffer (0.1% Igepal, 1 mM PMSF, 1 × Protease Cocktail inhibitor (Sigma-Aldrich)) were added for 10 mg of tissue. Muscle and spleen samples were homogenized using an electrical homogenizer (Ultraturrax IKA, Staufen Germany). After an incubation of 20 min on ice, suspensions were centrifuged 15 min at 20,000×
g at 4 °C. Supernatants were collected, aliquoted and stored at −20 °C. Protein concentration was determined by the Qubit according to the instructions of the manufacturer (Invitrogen/Life Technologies, Carlsbad, CA, USA).
4.8. Measurement of Antibody Response to rHyal-sk Plus GET
Serum samples were collected from the tail vein of mice treated with rHyal-
sk plus GET and mice treated with bHyal plus GET at 3 h (d0) and 7 days after the treatment. Serum samples were harvested from untreated mice as negative control. Nunc Maxisorp 96 well-plates (Nunc A/S, Roskilde, Denmark) were coated with rHyal-
sk or bHyal (8 μg/mL) in 0.1 M NaHCO
3 buffer, pH 9.6. Serum ELISA was performed as previously described [
21].
4.9. ELISA for Cytokines Quantification
The concentration of IL-1β, IL-6 and TNF-α in the protein extracts was quantified by ELISA according to the instructions of the manufacturers (Invitrogen/Life Technologies and eBioscience). Briefly, plates were coated overnight at 4 °C with a capture antibody and blocked with a blocking buffer (3% BSA in PBS or diluent buffer) for 1 h at room temperature (RT). Plates were then incubated 2 h at RT with 50 μg of protein extract per well diluted in diluent buffer. After three washes in PBS-Tween20-0.5%, plates were incubated 1 h at RT with biotinylated detection antibody, washed, and incubated 30 min at RT with Avidin-Horseradish Peroxidase (HRP). After the last wash, plates were developed with 3,3′,5,5′-tetremethylbenzidine (TMB) substrate for 15–20 min. The reaction was stopped by adding 2 N H2SO4. The optical density was measured at 450 nm.
4.10. Statistical Analysis
Statistical analysis was performed using Prism 5 statistical software (GraphPad Software Inc., San Diego, CA, USA). Results were reported as the mean ± (standard deviation) SD of quadruplicates. Comparison of the ELISA results obtained in the different experimental groups was performed with the unpaired Student’s t test. Statistical analysis of the TdTomato fluorescence evaluation was performed using a Mann Whitney test. Differences with p value < 0.05 were considered statistically significant.

 ,
, 







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}