
The Role of Interstitial Fluid Pressure in Cerebral Porous Biomaterial Integration


 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Biomaterial Manufacturing
2.2. Protein Covalent Immobilization: Matrigel and Laminin
2.3. Protein Covalent Immobilization: Collagen Type I
2.4. Bradford Protein Quantification Assay
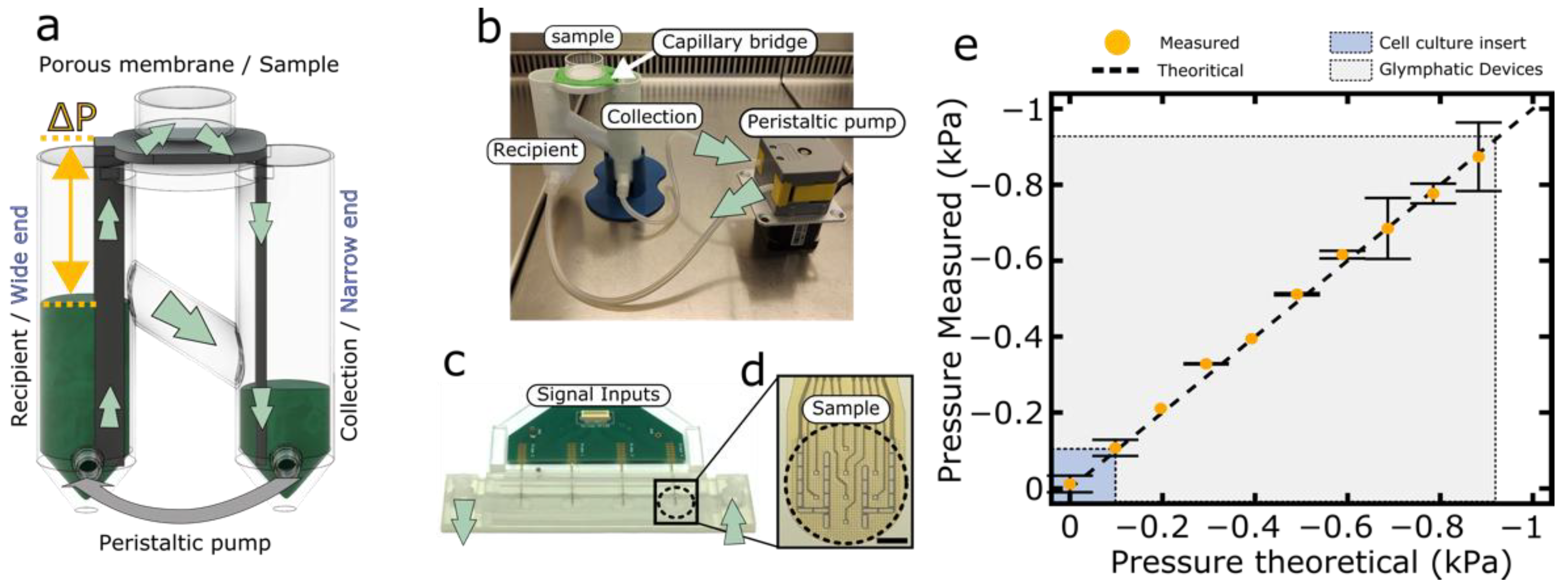
2.5. Glymphatic Brain-on-a-Chip Pressure Device
2.6. LUHMES-Based CellBrain Production
2.7. Biointegration as a Function of the Pressure
2.8. Histology
2.9. hiPSCs 2D and 3D Culture
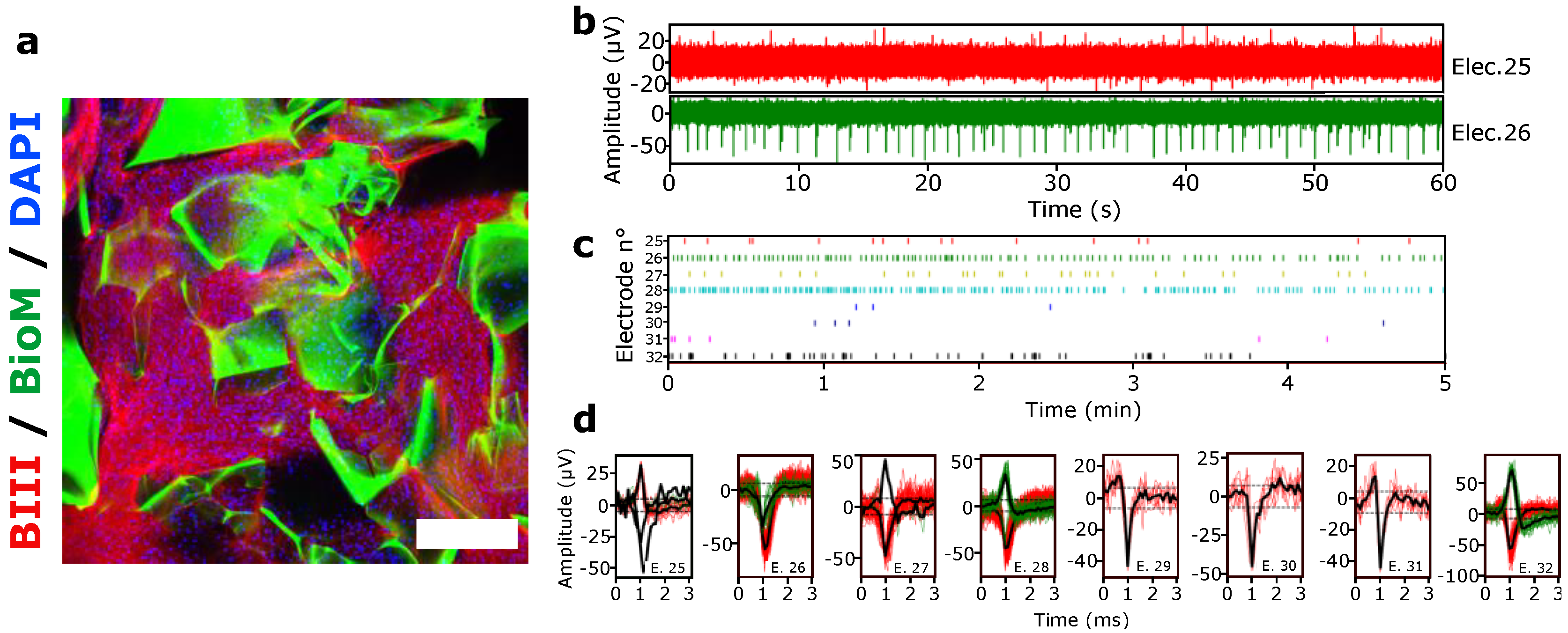
2.10. Electrophysiology
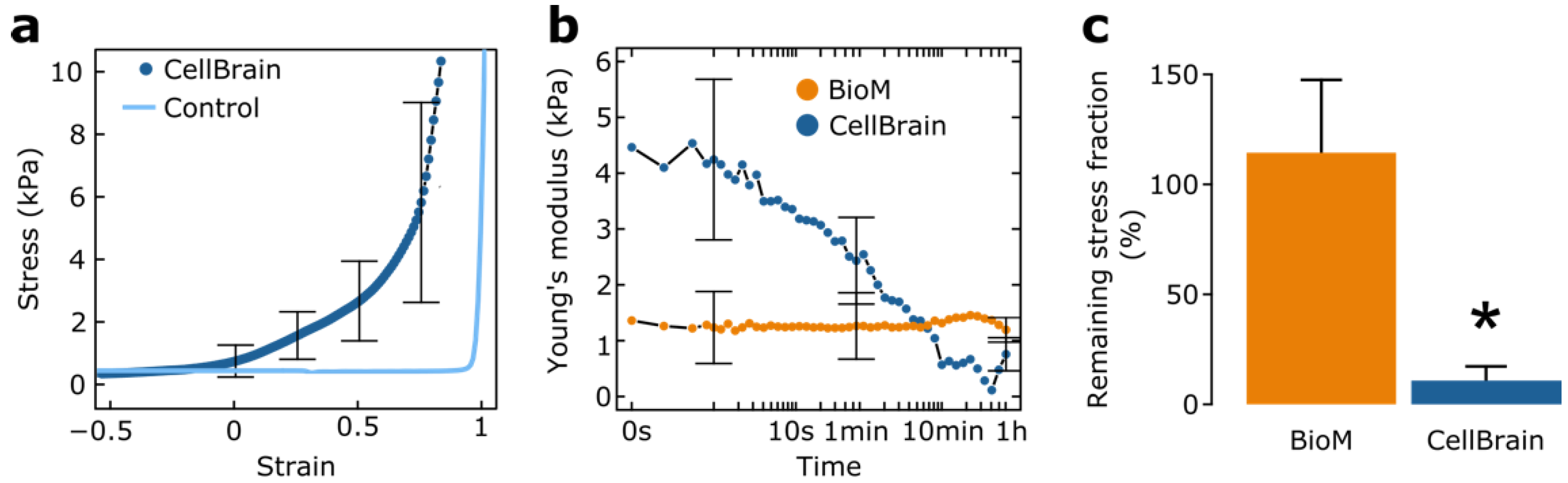
2.11. Mechanical Characterizations
3. Results
3.1. Results
3.1.1. Glymphatic Brain-on-a-Chip Principle and Characterization
3.1.2. Biomaterial Fabrication and Covalent Attachment of Cell-Adhesive Proteins
3.1.3. Interstitial Fluid Pressure-Dependent Biointegration by a Porous Biomaterial Scaffold
3.1.4. Spontaneous Electrical Activity of hiPSCs 3D Culture under Negative Pressure Conditions
3.1.5. Mechanical Characterization of the CellBrains
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gorelick, P.B. The Global Burden of Stroke: Persistent and Disabling. Lancet Neurol. 2019, 18, 417–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feigin, V.L.; Vos, T.; Nichols, E.; Owolabi, M.O.; Carroll, W.M.; Dichgans, M.; Deuschl, G.; Parmar, P.; Brainin, M.; Murray, C. The Global Burden of Neurological Disorders: Translating Evidence into Policy. Lancet Neurol. 2020, 19, 255–265. [Google Scholar] [CrossRef] [PubMed]
- Chin, J.H.; Vora, N. The Global Burden of Neurologic Diseases. Neurology 2014, 83, 349–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reynolds, B.A.; Weiss, S. Clonal and Population Analyses Demonstrate That an EGF-Responsive Mammalian Embryonic CNS Precursor Is a Stem Cell. Dev. Biol. 1996, 175, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Gage, F.H. Brain, Repair Yourself. Sci. Am. 2003, 289, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Fry, E.J. Central Nervous System Regeneration: Mission Impossible? Clin. Exp. Pharm. Physiol. 2001, 28, 253–258. [Google Scholar] [CrossRef]
- Bechmann, I. Failed Central Nervous System Regeneration: A Downside of Immune Privilege? NMM 2005, 7, 217–228. [Google Scholar] [CrossRef]
- Alexander, L.D.; Black, S.E.; Gao, F.; Szilagyi, G.; Danells, C.J.; McIlroy, W.E. Correlating Lesion Size and Location to Deficits after Ischemic Stroke: The Influence of Accounting for Altered Peri-Necrotic Tissue and Incidental Silent Infarcts. Behav. Brain Funct. 2010, 6, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, Y.-C.; Shyu, W.-C.; Lin, S.-Z.; Li, H. Regenerative Therapy for Stroke. Cell Transpl. 2007, 16, 171–181. [Google Scholar]
- Krause, M.; Phan, T.G.; Ma, H.; Sobey, C.G.; Lim, R. Cell-Based Therapies for Stroke: Are We There Yet? Front. Neurol. 2019, 10, 656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, J.P.; Burrell, J.C.; Struzyna, L.A.; Chen, H.I.; Serruya, M.D.; Wolf, J.A.; Duda, J.E.; Cullen, D.K. Emerging Regenerative Medicine and Tissue Engineering Strategies for Parkinson’s Disease. npj Parkinsons Dis. 2020, 6, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Díaz, M.L. Regenerative Medicine: Could Parkinson’s Be the First Neurodegenerative Disease to Be Cured? Future Sci. OA 2019, 5, FSO418. [Google Scholar] [CrossRef] [Green Version]
- Tuladhar, A.; Payne, S.L.; Shoichet, M.S. Harnessing the Potential of Biomaterials for Brain Repair after Stroke. Front. Mater. 2018, 5, 14. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.-S.; Harn, H.-J.; Chiou, T.-W. The Role of Biomaterials in Implantation for Central Nervous System Injury. Cell Transpl. 2018, 27, 407–422. [Google Scholar] [CrossRef]
- Ghuman, H.; Modo, M. Biomaterial Applications in Neural Therapy and Repair. Chin. Neurosurg. J. 2016, 2, 34. [Google Scholar] [CrossRef] [Green Version]
- Modo, M. Bioscaffold-Induced Brain Tissue Regeneration. Front. Neurosci. 2019, 13, 1156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, K.; Mao, X.; Xie, L.; Galvan, V.; Lai, B.; Wang, Y.; Gorostiza, O.; Wang, X.; Greenberg, D.A. Transplantation of Human Neural Precursor Cells in Matrigel Scaffolding Improves Outcome from Focal Cerebral Ischemia after Delayed Postischemic Treatment in Rats. J. Cereb. Blood Flow Metab. 2010, 30, 534–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boisserand, L.S.B.; Kodama, T.; Papassin, J.; Auzely, R.; Moisan, A.; Rome, C.; Detante, O. Biomaterial Applications in Cell-Based Therapy in Experimental Stroke. Stem Cells Int. 2016, 2016, 6810562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skop, N.B.; Calderon, F.; Cho, C.H.; Gandhi, C.D.; Levison, S.W. Improvements in Biomaterial Matrices for Neural Precursor Cell Transplantation. Mol. Cell 2014, 2, 19. [Google Scholar] [CrossRef] [Green Version]
- Jeong, K.J.; Kohane, D.S. Surface Modification and Drug Delivery for Biointegration. Ther. Deliv. 2011, 2, 737–752. [Google Scholar] [CrossRef]
- Yanez, M.; Blanchette, J.; Jabbarzadeh, E. Modulation of Inflammatory Response to Implanted Biomaterials Using Natural Compounds. CPD 2018, 23, 6347–6357. [Google Scholar] [CrossRef] [Green Version]
- Anderson, J.M.; Rodriguez, A.; Chang, D.T. Foreign Body Reaction to Biomaterials. Semin. Immunol. 2008, 20, 86–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdulghani, S.; Mitchell, G.R. Biomaterials for In Situ Tissue Regeneration: A Review. Biomolecules 2019, 9, 750. [Google Scholar] [CrossRef] [Green Version]
- Béduer, A.; Piacentini, N.; Aeberli, L.; Da Silva, A.; Verheyen, C.A.; Bonini, F.; Rochat, A.; Filippova, A.; Serex, L.; Renaud, P.; et al. Additive Manufacturing of Hierarchical Injectable Scaffolds for Tissue Engineering. Acta Biomater. 2018, 76, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Béduer, A.; Bonini, F.; Verheyen, C.A.; Genta, M.; Martins, M.; Brefie-Guth, J.; Tratwal, J.; Filippova, A.; Burch, P.; Naveiras, O.; et al. An Injectable Meta-Biomaterial: From Design and Simulation to In Vivo Shaping and Tissue Induction. Adv. Mater. 2021, 33, 2102350. [Google Scholar] [CrossRef]
- Watts, M.E.; Pocock, R.; Claudianos, C. Brain Energy and Oxygen Metabolism: Emerging Role in Normal Function and Disease. Front. Mol. Neurosci. 2018, 11, 216. [Google Scholar] [CrossRef] [PubMed]
- Junka, R.; Valmikinathan, C.M.; Kalyon, D.M.; Yu, X. Laminin Functionalized Biomimetic Nanofibers for Nerve Tissue Engineering. J. Biomat. Tissue Engng. 2013, 3, 494–502. [Google Scholar] [CrossRef] [Green Version]
- Parenteau-Bareil, R.; Gauvin, R.; Berthod, F. Collagen-Based Biomaterials for Tissue Engineering Applications. Materials 2010, 3, 1863–1887. [Google Scholar] [CrossRef] [Green Version]
- Bellis, S.L. Advantages of RGD Peptides for Directing Cell Association with Biomaterials. Biomaterials 2011, 32, 4205–4210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, R.; Santhosh, M.; Dash, J.K.; Karpoormath, R.; Jha, A.; Kwak, J.; Patel, M.; Kim, J.H. Ile-Lys-Val-Ala-Val (IKVAV) Peptide for Neuronal Tissue Engineering. Polym Adv. Technol 2019, 30, 4–12. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Mao, Z.; Tan, H.; Han, L.; Ren, T.; Gao, C. Gradient Biomaterials and Their Influences on Cell Migration. Interface Focus 2012, 2, 337–355. [Google Scholar] [CrossRef] [Green Version]
- Sharma, K.; Mujawar, M.A.; Kaushik, A. State-of-Art Functional Biomaterials for Tissue Engineering. Front. Mater. 2019, 6, 172. [Google Scholar] [CrossRef] [Green Version]
- Peplow, P.V.; Chatterjee, M.P. A Review of the Influence of Growth Factors and Cytokines in in Vitro Human Keratinocyte Migration. Cytokine 2013, 62, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Seo, J.; Shin, J.-Y.; Leijten, J.; Jeon, O.; Camci-Unal, G.; Dikina, A.D.; Brinegar, K.; Ghaemmaghami, A.M.; Alsberg, E.; Khademhosseini, A. High-Throughput Approaches for Screening and Analysis of Cell Behaviors. Biomaterials 2018, 153, 85–101. [Google Scholar] [CrossRef] [PubMed]
- Guyton, A.C.; Granger, H.J.; Taylor, A.E. Interstitial Fluid Pressure. Physiol. Rev. 1971, 51, 527–563. [Google Scholar] [CrossRef]
- Kang, P.H.; Kumar, S.; Schaffer, D.V. Novel Biomaterials to Study Neural Stem Cell Mechanobiology and Improve Cell-Replacement Therapies. Curr. Opin. Biomed. Eng. 2017, 4, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Lacour, S.P.; Courtine, G.; Guck, J. Materials and Technologies for Soft Implantable Neuroprostheses. Nat. Rev. Mater. 2016, 1, 16063. [Google Scholar] [CrossRef] [Green Version]
- Moshayedi, P.; Ng, G.; Kwok, J.C.F.; Yeo, G.S.H.; Bryant, C.E.; Fawcett, J.W.; Franze, K.; Guck, J. The Relationship between Glial Cell Mechanosensitivity and Foreign Body Reactions in the Central Nervous System. Biomaterials 2014, 35, 3919–3925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, J.K.; Park, D.J.; Skousen, J.L.; Hess-Dunning, A.E.; Tyler, D.J.; Rowan, S.J.; Weder, C.; Capadona, J.R. Mechanically-Compliant Intracortical Implants Reduce the Neuroinflammatory Response. J. Neural Eng. 2014, 11, 056014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Annabi, N.; Mithieux, S.M.; Camci-Unal, G.; Dokmeci, M.R.; Weiss, A.S.; Khademhosseini, A. Elastomeric Recombinant Protein-Based Biomaterials. Biochem. Eng. J. 2013, 77, 110–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wellman, S.M.; Eles, J.R.; Ludwig, K.A.; Seymour, J.P.; Michelson, N.J.; McFadden, W.E.; Vazquez, A.L.; Kozai, T.D.Y. A Materials Roadmap to Functional Neural Interface Design. Adv. Funct. Mater. 2018, 28, 1701269. [Google Scholar] [CrossRef] [PubMed]
- Gasik, M. Understanding Biomaterial-Tissue Interface Quality: Combined in Vitro Evaluation. Sci. Technol. Adv. Mater. 2017, 18, 550–562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, R.; Murphy, M.; Li, C.; Ting, K.; Soo, C.; Zheng, Z. Current Development of Biodegradable Polymeric Materials for Biomedical Applications. DDDT 2018, 12, 3117–3145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brodersen, P.; Højgaard, K.; Lassen, N.A. Measurement of “Interstitial Fluid” Pressure in the Brain in Dogs. In Intracranial Pressure; Brock, M., Dietz, H., Eds.; Springer: Berlin/Heidelberg, Germany, 1972; pp. 185–187. ISBN 978-3-642-65488-6. [Google Scholar]
- Jessen, N.A.; Munk, A.S.F.; Lundgaard, I.; Nedergaard, M. The Glymphatic System: A Beginner’s Guide. Neurochem. Res. 2015, 40, 2583–2599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iliff, J.J.; Wang, M.; Liao, Y.; Plogg, B.A.; Peng, W.; Gundersen, G.A.; Benveniste, H.; Vates, G.E.; Deane, R.; Goldman, S.A.; et al. A Paravascular Pathway Facilitates CSF Flow Through the Brain Parenchyma and the Clearance of Interstitial Solutes, Including Amyloid. Sci. Transl. Med. 2012, 4, 147ra111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nedergaard, M. Garbage Truck of the Brain. Science 2013, 340, 1529–1530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benveniste, H.; Liu, X.; Koundal, S.; Sanggaard, S.; Lee, H.; Wardlaw, J. The Glymphatic System and Waste Clearance with Brain Aging: A Review. Gerontology 2019, 65, 106–119. [Google Scholar] [CrossRef]
- Mestre, H.; Hablitz, L.M.; Xavier, A.L.; Feng, W.; Zou, W.; Pu, T.; Monai, H.; Murlidharan, G.; Castellanos Rivera, R.M.; Simon, M.J.; et al. Aquaporin-4-Dependent Glymphatic Solute Transport in the Rodent Brain. eLife 2018, 7, e40070. [Google Scholar] [CrossRef]
- Abaci, H.E.; Gledhill, K.; Guo, Z.; Christiano, A.M.; Shuler, M.L. Pumpless Microfluidic Platform for Drug Testing on Human Skin Equivalents. Lab. Chip 2015, 15, 882–888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sepúlveda, G.; Espíndola, M.; Maureira, M.; Sepúlveda, E.; Ignacio Fernández, J.; Oliva, C.; Sanhueza, A.; Vial, M.; Manterola, C. Negative-pressure wound therapy versus standard wound dressing in the treatment of diabetic foot amputation. A randomised controlled trial. Cir. Esp. 2009, 86, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Blalock, L. Use of Negative Pressure Wound Therapy With Instillation and a Novel Reticulated Open-Cell Foam Dressing With Through Holes at a Level 2 Trauma Center. Wounds 2019, 31, 55–58. [Google Scholar] [PubMed]
- Wilkes, R.P.; McNulty, A.K.; Feeley, T.D.; Schmidt, M.A.; Kieswetter, K. Bioreactor for Application of Subatmospheric Pressure to Three-Dimensional Cell Culture. Tissue Eng. 2007, 13, 3003–3010. [Google Scholar] [CrossRef]
- Giandomenico, S.L.; Mierau, S.B.; Gibbons, G.M.; Wenger, L.M.D.; Masullo, L.; Sit, T.; Sutcliffe, M.; Boulanger, J.; Tripodi, M.; Derivery, E.; et al. Cerebral Organoids at the Air–Liquid Interface Generate Diverse Nerve Tracts with Functional Output. Nat. Neurosci. 2019, 22, 669–679. [Google Scholar] [CrossRef] [PubMed]
- Béduer, A.; Braschler, T.; Peric, O.; Fantner, G.E.; Mosser, S.; Fraering, P.C.; Benchérif, S.; Mooney, D.J.; Renaud, P. A Compressible Scaffold for Minimally Invasive Delivery of Large Intact Neuronal Networks. Adv. Healthc. Mater. 2015, 4, 301–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carter, J.D.; LaBean, T.H. Coupling Strategies for the Synthesis of Peptide-Oligonucleotide Conjugates for Patterned Synthetic Biomineralization. J. Nucleic Acids 2011, 2011, 926595. [Google Scholar] [CrossRef] [Green Version]
- Filippova, A.; Bonini, F.; Efremova, L.; Preynat-Seauve, O.; Béduer, A.; Krause, K.-H.; Braschler, T. Neurothreads: Cryogel Carrier-Based Differentiation and Delivery of Mature Neurons in the Treatment of Parkinson’s Disease; Biomaterials: New York, NY, USA, 2020. [Google Scholar]
- Smirnova, L.; Harris, G.; Delp, J.; Valadares, M.; Pamies, D.; Hogberg, H.T.; Waldmann, T.; Leist, M.; Hartung, T. A LUHMES 3D Dopaminergic Neuronal Model for Neurotoxicity Testing Allowing Long-Term Exposure and Cellular Resilience Analysis. Arch. Toxicol. 2016, 90, 2725–2743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, J.I.; Lee, Y.-K. EVA-Enhanced Embedding Medium for Histological Analysis of 3D Porous Scaffold Material. Micron 2009, 40, 756–760. [Google Scholar] [CrossRef] [PubMed]
- Govindan, S.; Batti, L.; Osterop, S.F.; Stoppini, L.; Roux, A. Mass Generation, Neuron Labeling, and 3D Imaging of Minibrains. Front. Bioeng. Biotechnol. 2021, 8, 582650. [Google Scholar] [CrossRef]
- Wertenbroek, R.; Thoma, Y.; Mor, F.M.; Grassi, S.; Heuschkel, M.O.; Roux, A.; Stoppini, L. SpikeOnChip: A Custom Embedded Platform for Neuronal Activity Recording and Analysis. IEEE Trans. Biomed. Circ. Syst. 2021, 9, 155–165. [Google Scholar] [CrossRef]
- Abbott, N.J. Evidence for Bulk Flow of Brain Interstitial Fluid: Significance for Physiology and Pathology. Neurochem. Int. 2004, 45, 545–552. [Google Scholar] [CrossRef] [PubMed]
- Iliff, J.J.; Wang, M.; Zeppenfeld, D.M.; Venkataraman, A.; Plog, B.A.; Liao, Y.; Deane, R.; Nedergaard, M. Cerebral Arterial Pulsation Drives Paravascular CSF-Interstitial Fluid Exchange in the Murine Brain. J. Neurosci. 2013, 33, 18190–18199. [Google Scholar] [CrossRef] [Green Version]
- Starling, E.H. On the Absorption of Fluids from the Connective Tissue Spaces. J. Physiol 1896, 19, 312–326. [Google Scholar] [CrossRef] [PubMed]
- Chiu, Y.-C.; Cheng, M.-H.; Engel, H.; Kao, S.-W.; Larson, J.C.; Gupta, S.; Brey, E.M. The Role of Pore Size on Vascularization and Tissue Remodeling in PEG Hydrogels. Biomaterials 2011, 32, 6045–6051. [Google Scholar] [CrossRef]
- Del Monte, U. Does the Cell Number 109 Still Really Fit One Gram of Tumor Tissue? Cell Cycle 2009, 8, 505–506. [Google Scholar] [CrossRef] [Green Version]
- Budday, S.; Nay, R.; de Rooij, R.; Steinmann, P.; Wyrobek, T.; Ovaert, T.C.; Kuhl, E. Mechanical Properties of Gray and White Matter Brain Tissue by Indentation. J. Mech. Behav. Biomed. Mater. 2015, 46, 318–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estermann, S.-J.; Pahr, D.H.; Reisinger, A. Hyperelastic and Viscoelastic Characterization of Hepatic Tissue under Uniaxial Tension in Time and Frequency Domain. J. Mech. Behav. Biomed. Mater. 2020, 112, 104038. [Google Scholar] [CrossRef]
- Forgacs, G.; Foty, R.A.; Shafrir, Y.; Steinberg, M.S. Viscoelastic Properties of Living Embryonic Tissues: A Quantitative Study. Biophys. J. 1998, 74, 2227–2234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghashghaei, H.T.; Lai, C.; Anton, E.S. Neuronal Migration in the Adult Brain: Are We There Yet? Nat. Rev. Neurosci. 2007, 8, 141–151. [Google Scholar] [CrossRef]
- Kaneko, N.; Sawada, M.; Sawamoto, K. Mechanisms of Neuronal Migration in the Adult Brain. J. Neurochem. 2017, 141, 835–847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liesi, P. Do Neurons in the Vertebrate CNS Migrate on Laminin? EMBO J. 1985, 4, 1163–1170. [Google Scholar] [PubMed]
- Fujioka, T.; Kaneko, N.; Ajioka, I.; Nakaguchi, K.; Omata, T.; Ohba, H.; Fässler, R.; García-Verdugo, J.M.; Sekiguchi, K.; Matsukawa, N.; et al. Β1 Integrin Signaling Promotes Neuronal Migration along Vascular Scaffolds in the Post-Stroke Brain. EBioMedicine 2017, 16, 195–203. [Google Scholar] [CrossRef] [Green Version]
- Andorko, J.I.; Jewell, C.M. Designing Biomaterials with Immunomodulatory Properties for Tissue Engineering and Regenerative Medicine. Bioeng. Transl. Med. 2017, 2, 139–155. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.; Li, P.; Feng, H.; Zhang, X.; Chu, P.K. Engineering and Functionalization of Biomaterials via Surface Modification. J. Mater. Chem. B 2015, 3, 2024–2042. [Google Scholar] [CrossRef]
- Fowler, J.B.; De Jesus, O.; Mesfin, F.B. Ventriculoperitoneal Shunt. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Hofmann, M.; Pflanzer, R.; Zöller, N.N.; Bernd, A.; Kaufmann, R.; Thaci, D.; Bereiter-Hahn, J.; Hirohata, S.; Kippenberger, S. Vascular Endothelial Growth Factor C-Induced Lymphangiogenesis Decreases Tumor Interstitial Fluid Pressure and Tumor. Transl. Oncol. 2013, 6, 398–404. [Google Scholar] [CrossRef] [Green Version]
- Song, E.; Mao, T.; Dong, H.; Boisserand, L.S.B.; Antila, S.; Bosenberg, M.; Alitalo, K.; Thomas, J.-L.; Iwasaki, A. VEGF-C-Driven Lymphatic Drainage Enables Immunosurveillance of Brain Tumours. Nature 2020, 577, 689–694. [Google Scholar] [CrossRef]
- Guyton, A.C.; Frank, M.; Abernathy, B. A Concept of Negative Interstitial Pressure Based on Pressures in Implanted Perforated Capsules. Circ. Res. 1963, 12, 399–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Michniak-Kohn, B.B. Tissue Engineered Human Skin Equivalents. Pharmaceutics 2012, 4, 26–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bonini, F.; Mosser, S.; Mor, F.M.; Boutabla, A.; Burch, P.; Béduer, A.; Roux, A.; Braschler, T. The Role of Interstitial Fluid Pressure in Cerebral Porous Biomaterial Integration. Brain Sci. 2022, 12, 417. https://doi.org/10.3390/brainsci12040417
Bonini F, Mosser S, Mor FM, Boutabla A, Burch P, Béduer A, Roux A, Braschler T. The Role of Interstitial Fluid Pressure in Cerebral Porous Biomaterial Integration. Brain Sciences. 2022; 12(4):417. https://doi.org/10.3390/brainsci12040417
Chicago/Turabian StyleBonini, Fabien, Sébastien Mosser, Flavio Maurizio Mor, Anissa Boutabla, Patrick Burch, Amélie Béduer, Adrien Roux, and Thomas Braschler. 2022. "The Role of Interstitial Fluid Pressure in Cerebral Porous Biomaterial Integration" Brain Sciences 12, no. 4: 417. https://doi.org/10.3390/brainsci12040417