
Soybean Meal-Dependent Intestinal Inflammation Induces Different Patterns of Bone-Loss in Adult Zebrafish Scale

 , ,
, ,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Ethic Statement
2.2. Animals
2.3. Diets
2.4. Samples Collection
2.5. Histological Analysis and Morphometric Measures
2.6. Myeloperoxidase Staining and Cell Count
2.7. Intestinal Gene Expression Analysis and Inflammatory Status Estimation
2.8. Bone Matrix Vital Staining
2.9. Histological TRAP and ALP Assay in Scales
2.10. Biochemical TRAP and ALP Scale Assays
2.11. Statistics
3. Results
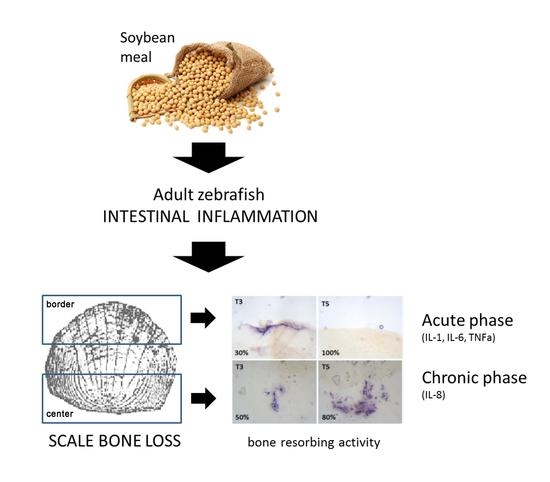
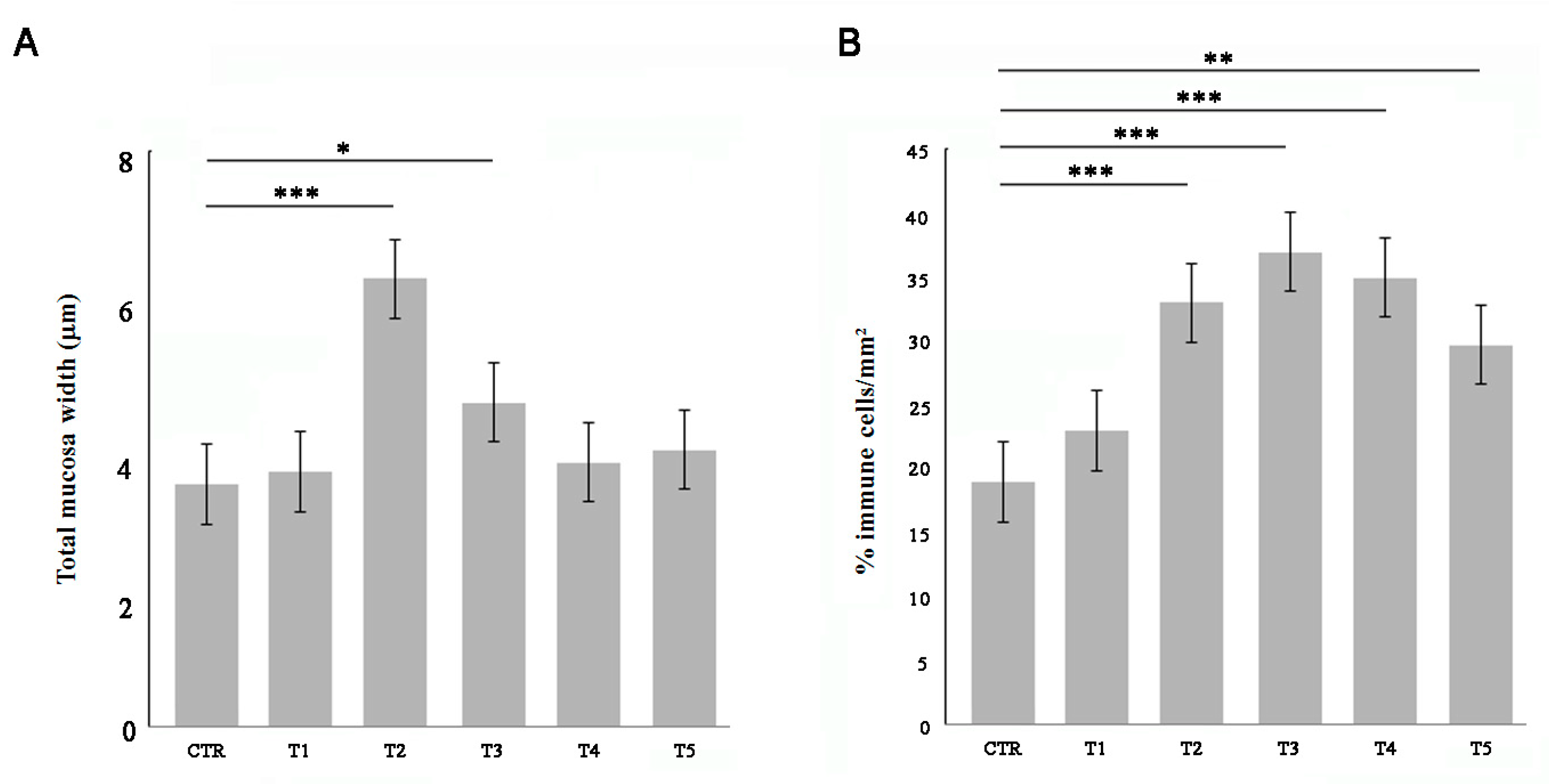
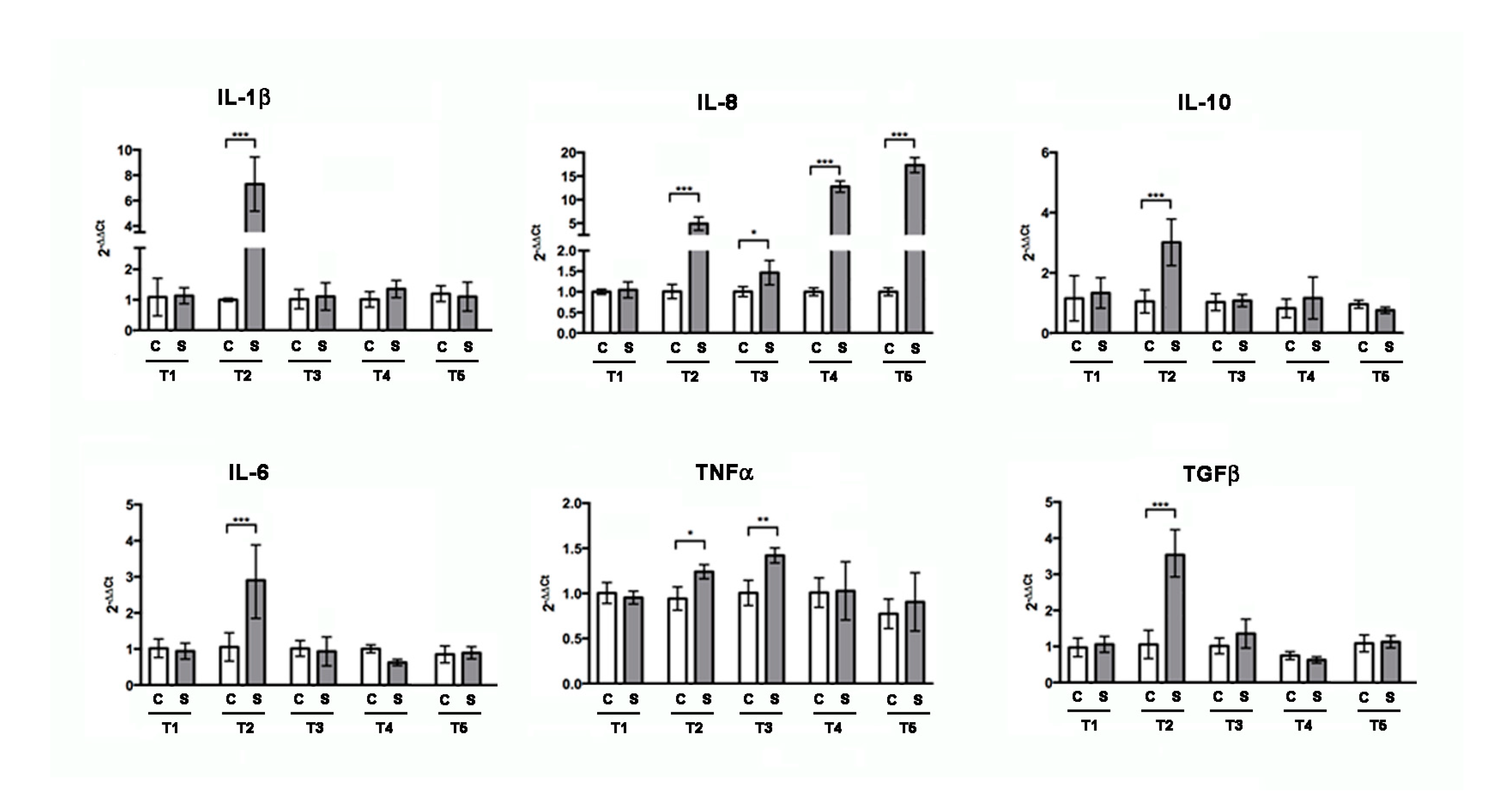
3.1. Soybean Meal Induces Intestinal Inflammation in Adult Zebrafish
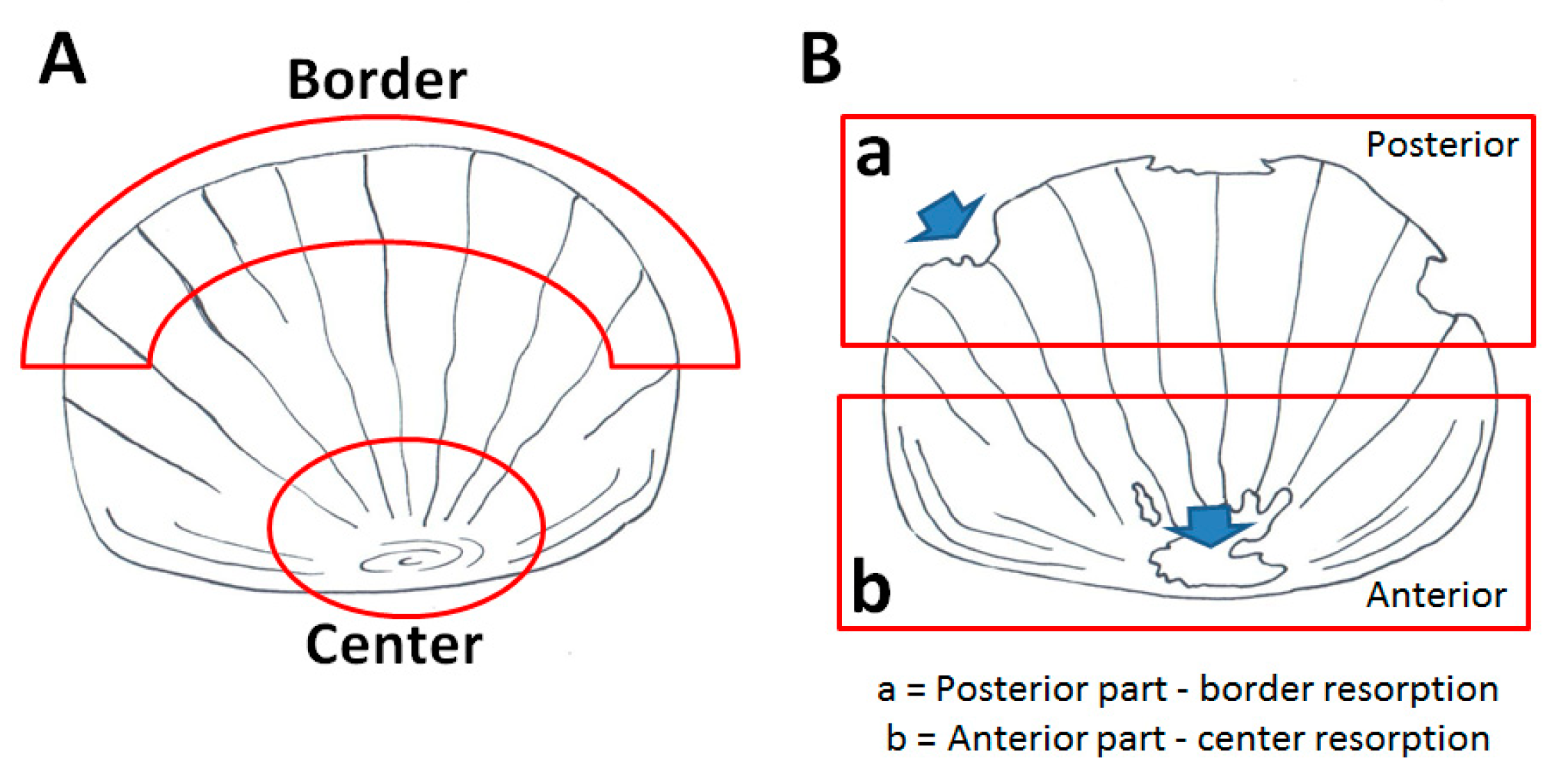
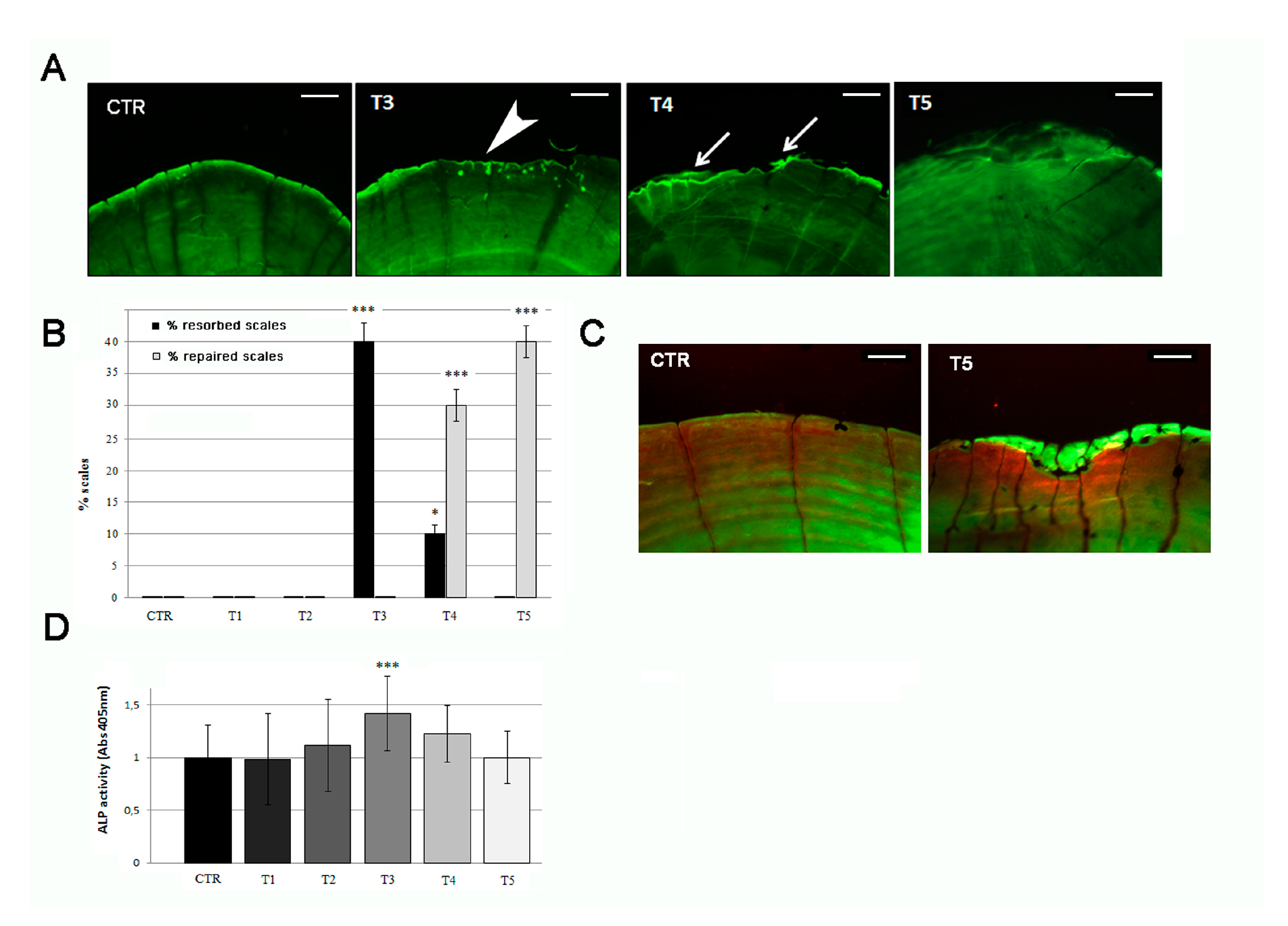
3.2. Soybean-Meal Induces a Transient Bone Resorption in the Scale Border
3.3. Anabolic Stimuli Rescued Osteoporotic Phenotype in the Scale Border
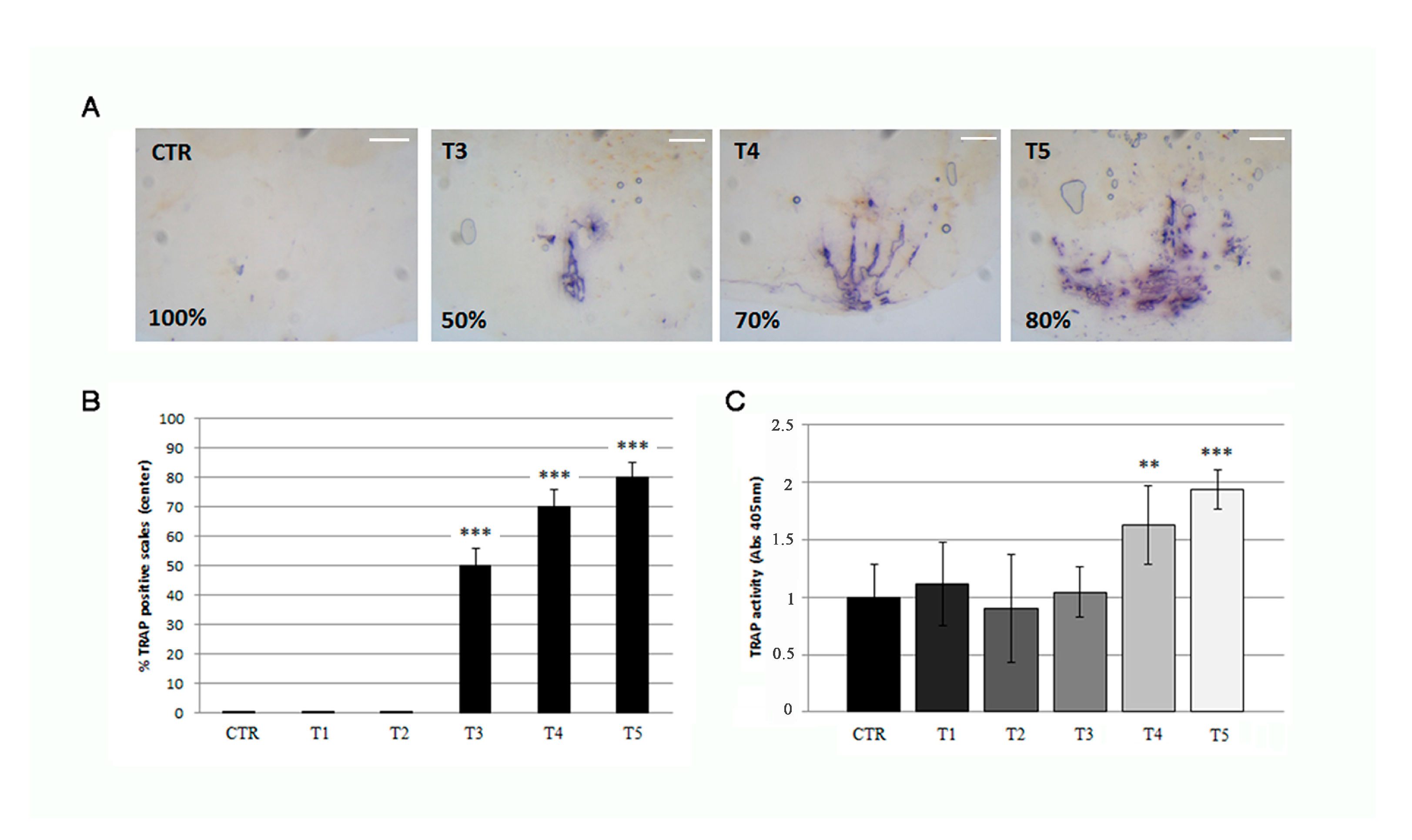
3.4. Soybean-Meal Induces a Permanent Bone Resorption in the Scale Center
3.5. Osteoporotic Phenotype in the Scale Center Is Not Rescued by Anabolic Stimuli
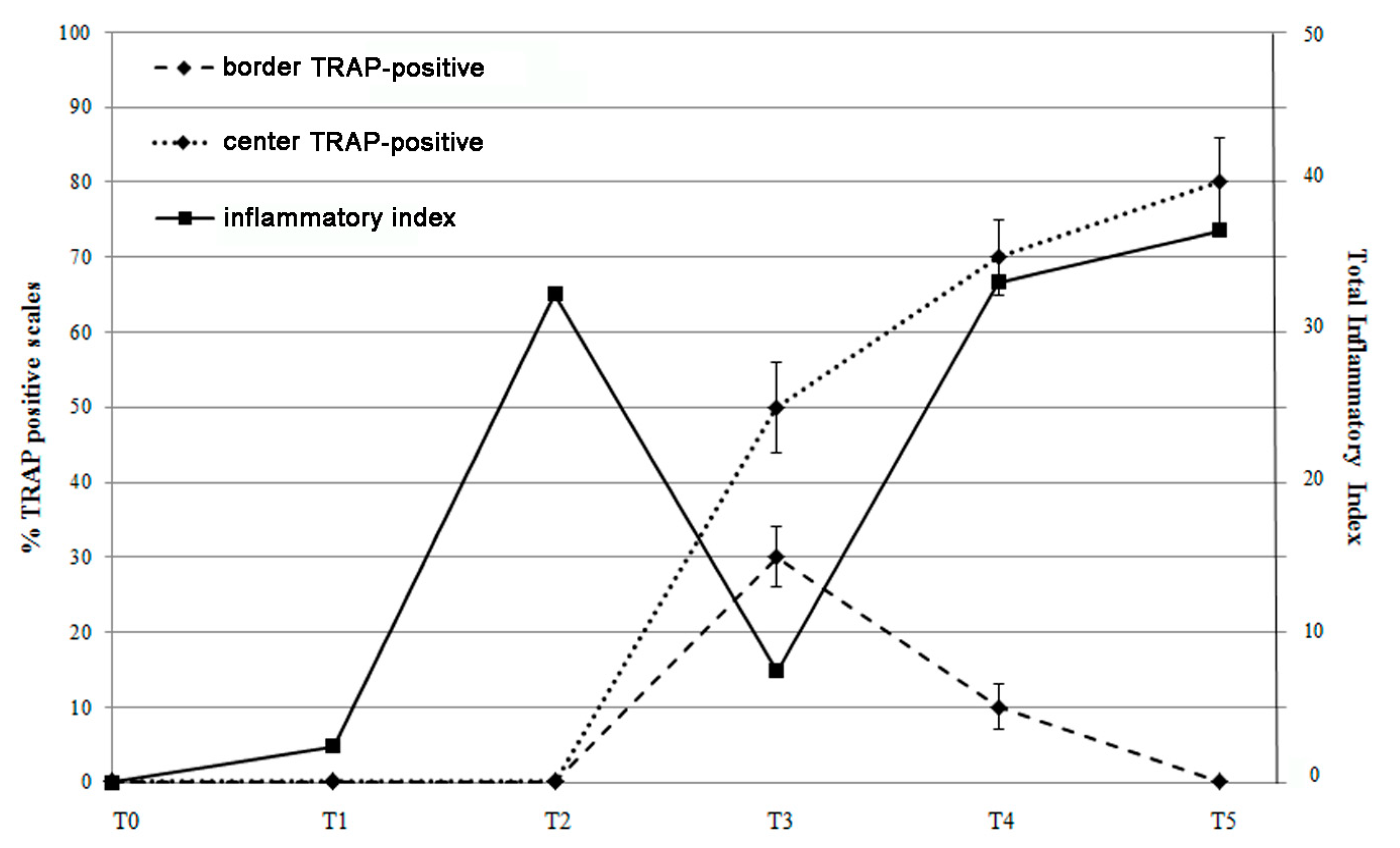
3.6. Different Inflammatory Stimuli Causes Different Bone Loss Patterns in Zebrafish Scales
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Bravenboer, N.; Oostlander, A.E.; van Bodegraven, A.A. Bone loss in patients with inflammatory bowel disease: Cause, detection and treatment. Curr. Opin. Gastroenterol. 2021, 37, 128–134. [Google Scholar] [CrossRef]
- Mariotti, M.; Carnovali, M.; Banfi, G. Danio rerio: The Janus of the bone from embryo to scale. Clin. Cases Miner. Bone Metab. 2015, 12, 188–194. [Google Scholar] [CrossRef]
- Yang, Y.; Tomkovich, S.; Jobin, C. Could a Swimming Creature Inform Us on Intestinal Diseases? Lessons from Zebrafish. Inflamm. Bowel Dis. 2014, 20, 956–966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallace, K.N.; Akhter, S.; Smith, E.M.; Lorent, K.; Pack, M. Intestinal growth and differentiation in zebrafish. Mech. Dev. 2005, 122, 157–173. [Google Scholar] [CrossRef] [PubMed]
- Hanyang, L.; Xuanzhe, L.; Xuyang, C.; Yujia, Q.; Jiarong, F.; Jun, S.; Zhihua, R. Application of Zebrafish Models in Inflammatory Bowel Disease. Front. Immunol. 2017, 8, 501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldsmith, J.R.; Jobin, C. Think small: Zebrafish as a model system of human pathology. J. Biomed. Biotechnol. 2012, 2012, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Marjoram, L.; Alvers, A.; Deerhake, M.E.; Bagwell, J.; Mankiewicz, J.; Cocchiaro, J.L.; Beerman, R.W.; Willer, J.; Sumigray, K.D.; Katsanis, N.; et al. Epigenetic control of intestinal barrier function and inflammation in zebrafish. Proc. Natl. Acad. Sci. USA 2015, 112, 2770–2775. [Google Scholar] [CrossRef] [Green Version]
- Baeverfjord, G.; Krogdahl, A. Development and regression of soybean meal induced enteritis in Atlantic salmon, Salmo salar L., distal intestine: A comparison with the intestines of fasted fish. J. Fish Dis. 1996, 19, 375–387. [Google Scholar] [CrossRef]
- Bakke-McKellep, A.M.; Penn, M.H.; Salas, P.M.; Refstie, S.; Sperstad, S.; Landsverk, T.; Ringø, E.; Krogdahl, A. Effects of dietary soyabean meal, inulin and oxytetracycline on intestinal microbiota and epithelial cell stress, apoptosis and proliferation in the teleost Atlantic salmon (Salmo salar L.). Br. J. Nutr. 2007, 97, 699–713. [Google Scholar] [CrossRef] [Green Version]
- Bakke-Mckellep, M.; Mcl Press, C.; Baeverfjord, G.; Krogdahl, Å.; Landsverk, T. Changes in immune and enzyme histochemical phenotypes of cell in the intestinal mucosa of Atlantic salmon, Salmo salar L., with soybean meal-induced enteritis. J. Fish Dis. 2000, 23, 115–127. [Google Scholar] [CrossRef]
- Nordrum, S.; Bakke-McKellep, A.; Khrogdahl, A.; Buddington, R.K. Effects of soybean meal and salinity on intestinal transport of nutrients in Atlantic salmon (Salmo salar L.) and rainbow trout (Oncorhynchus mykiss). Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2000, 125, 317–335. [Google Scholar] [CrossRef]
- Uran, P.; Schrama, J.; Rombout, J.; Obach, A.; Jensen, L.; Koppe, W.; Verreth, J.A.J. Soybean meal induced enteritis in Atlantic salmon (Salmo salar) at different temperatures. Aquacult. Nutr. 2008, 14, 325–330. [Google Scholar]
- Krogdahl, Å.; Bakke-Mckellep, A.; Baeverfjiord, G. Effects of graded levels of standard soybean meal on intestinal structure, mucosal enzyme activities, and pancreatic response in Atlantic salmon (Salmon salar L.). Aquacult. Nutr. 2003, 9, 361–371. [Google Scholar] [CrossRef]
- Urán, P.A.; Gonçalves, A.A.; Taverne-Thiele, J.J.; Schrama, J.W.; Verreth, J.A.; Rombout, J.H. Soybean meal induces intestinal inflammation in common carp (Cyprinus carpio L.). Fish Shellfish Immunol. 2008, 25, 751–760. [Google Scholar] [CrossRef]
- Gu, M.; Jia, Q.; Zhang, Z.; Bai, N.; Xu, X.; Xu, B. Soya-saponins induce intestinal inflammation and barrier dysfunction in juvenile turbot (Scophthalmus maximus). Fish Shellfish Immunol. 2018, 77, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Hedrera, M.I.; Galdames, J.A.; Jimenez-Reyes, M.F.; Reyes, A.E.; Avendaño-Herrera, R.; Romero, J.; Feijóo, C.G. Soybean meal induces intestinal inflammation in zebrafish larvae. PLoS ONE 2013, 8, e69983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Francis, G.; Kerem, Z.; Makkar, H.P.S.; Becker, K. The biological action of saponins in animal systems: A review. Br. J. Nutr. 2002, 88, 587–605. [Google Scholar] [CrossRef]
- Knudsen, D.; Jutfelt, F.; Sundh, H.; Sundell, K.; Koppe, W.; Frøkiær, H. Dietary soya saponins increase gut permeability and play a key role in the onset of soyabean-induced enteritis in Atlantic salmon (Salmo salar L.). Br. J. Nutr. 2008, 100, 120–129. [Google Scholar] [CrossRef] [Green Version]
- Fuentes-Appelgren, P.; Opazo, R.; Barros, L.; Feijoó, C.G.; Urzúa, V.; Romero, J. Effect of the dietary inclusion of soybean components on the innate immune system in zebrafish. Zebrafish 2014, 11, 41–49. [Google Scholar] [CrossRef] [Green Version]
- Kiron, V.; Park, Y.; Siriyappagouder, P.; Dahle, D.; Vasanth, G.K.; Dias, J.; Fernandes, J.M.O.; Sørensen, M.; Trichet, V.V. Intestinal Transcriptome Analysis Reveals Soy Derivative-Linked Changes in Atlantic Salmon. Front. Immunol. 2020, 11, 596514. [Google Scholar] [CrossRef]
- Kortner, T.M.; Skugor, S.; Penn, M.H.; Mydland, L.T.; Djordjevic, B.; Hillestad, M.; Krasnov, A.; Krogdahl, A. Dietary soyasaponin supplementation to pea protein concentrate reveals nutrigenomic interactions underlying enteropathy in Atlantic salmon (Salmo salar). BMC Vet. Res. 2012, 8, 101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adamopoulos, I.E. Inflammation in bone physiology and pathology. Curr. Opin. Rheumatol. 2018, 30, 59–64. [Google Scholar] [CrossRef]
- Ali, T.; Lam, D.; Bronze, M.S.; Humphrey, M.B. Osteoporosis in inflammatory bowel disease. Am. J. Med. 2009, 122, 599–604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westerfield, M. The zebrafish book. In A Guide for the Laboratory Use of Zebrafish (Danio Rerio), 5th ed.; University of Oregon Press: Eugene, OR, USA, 2007. [Google Scholar]
- Matthews, M.; Varga, Z.M. Anesthesia and euthanasia in zebrafish. ILAR J. 2012, 53, 192–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, X.; Huang, X.; Zhou, Z.; Lin, X. An improvement of the 2ˆ(–delta delta CT) method for quantitative real-time polymerase chain reaction data analysis. Biostat. Bioinf. Biomath. 2013, 3, 71–85. [Google Scholar]
- Bond, D.J.; Andreazza, A.C.; Hughes, J.; Dhanoa, T.; Torres, I.J.; Kozicky, J.-M.; Young, L.; Lam, R.W.; Yatham, L.N. Association of peripheral inflammation with body mass index and depressive relapse in bipolar disorder. Psychoneuroendocrinology 2016, 65, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Kimmel, C.B.; DeLaurier, A.; Ullmann, B.; Dowd, J.; McFadden, M. Modes of developmental outgrowth and shaping of a craniofacial bone in zebrafish. PLoS ONE 2010, 5, e9475. [Google Scholar] [CrossRef] [Green Version]
- Pasqualetti, S.; Banfi, G.; Mariotti, M. Osteoblast and osteoclast behavior in zebrafish cultured scales. Cell Tissue Res. 2012, 350, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Perrson, P.; Takagi, Y.; Björnsson, B.T. Tartrate resistant acid phosphatases as a marker for scale resorption in rainbow trout, Oncorhynchus mykiss: Effects of estradiol-17β treatment and refeeding. Fish Physiol. Biochem. 1995, 14, 329–339. [Google Scholar] [CrossRef]
- Schulte, C.M.; Dignass, A.U.; Goebell, H.; Röher, H.D.; Schulte, K.M. Genetic factors determine extent of bone loss in inflammatory bowel disease. Gastroenterology 2000, 119, m909–m920. [Google Scholar] [CrossRef]
- Straub, R.H.; Cutolo, M.; Pacifici, R. Evolutionary medicine and bone loss in chronic inflammatory diseases: A theory of inflammation-related osteopenia. Semin. Arthritis Rheum. 2015, 45, 220–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tobias, J.H.; Sasi, M.R.; Greenwood, R.; Probert, C.S. Rapid hip bone loss in active Crohn’s disease patients receiving short-term corticosteroid therapy. Aliment. Pharmacol. Ther. 2004, 20, 951–957. [Google Scholar] [CrossRef] [PubMed]
- Bendre, M.S.; Montague, D.C.; Peery, T.; Akel, N.S.; Gaddy, D.; Suva, L.J. Interleukin-8 stimulation of osteoclastogenesis and bone resorption is a mechanism for the increased osteolysis of metastatic bone disease. Bone 2003, 33, 28–37. [Google Scholar] [CrossRef]
- Kopesky, P.; Tiedemann, K.; Alkekhia, D.; Zechner, C.; Millard, B.; Schoeberl, B.; Komarova, S.V. Autocrine signaling is a key regulatory element during osteoclastogenesis. Biol. Open 2014, 3, 767–776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Krishnamurthy, A.; Hensvold, A.; Joshua, V.; Sun, M.; Engstrom, M.; Wähämaa, H.; Malmström, V.; Jopling, L.; Rethi, B.; et al. AB0078 Role of IL-8 and its receptor in anti-citrullinated protein antibody mediated osteoclastogenesis in RA. Ann. Rheum. Dis. 2016, 75, 923. [Google Scholar] [CrossRef]
- Seitz, M.; Dewald, B.; Gerber, N.; Baggiolini, M. Enhanced Production of Neutrophil-Activating peptide-1/interleukin-8 in Rheumatoid Arthritis. J. Clin. Investig. 1991, 8, 463–469. [Google Scholar] [CrossRef]
- Anttila, H.S.; Reitamo, S.; Erkko, P.; Ceska, M.; Moser, B.; Baggiolini, M. Interleukin-8 immunoreactivity in the skin of healthy subjects and patients with palmoplantar pustulosis and psoriasis. J. Investig. Dermatol. 1992, 98, 96–101. [Google Scholar] [CrossRef] [Green Version]
- Grimm, M.C.; Elsbury, S.K.; Pavli, P.; Doe, W.F. Interleukin 8: Cells of origin in inflammatory bowel disease. Gut 1996, 38, 90–98. [Google Scholar] [CrossRef] [Green Version]
- Brandt, E.; Colombel, J.; Ectors, N.; Gambiez, L.; Emilie, D.; Geboes, K.; Capron, M.; Desreumaux, P. Enhanced production of IL-8 in chronic but not in early ileal lesions of Crohn’s disease (CD). Clin. Exp. Immunol. 2000, 122, 180–185. [Google Scholar] [CrossRef]
- Goodman, S.B.; Pajarinen, J.; Yao, Z.; Lin, T. Inflammation and Bone Repair: From Particle Disease to Tissue Regeneration. Front. Bioeng. Biotechnol. 2019, 19, 230. [Google Scholar] [CrossRef]
- Roldán, J.F.; Del Rincón, I.; Escalante, A. Loss of Cortical Bone from the Metacarpal Diaphysis in Patients with Rheumatoid Arthritis: Independent Effects of Systemic Inflammation and Glucocorticoids. J. Rheumatol. 2006, 33, 508–516. [Google Scholar] [PubMed]
- Cascio, V.L.; Bonucci, E.; Imbimbo, B.; Ballanti, P.; Adami, S.; Milani, S.; Tartarotti, D.; Della Rocca, C. Bone loss in response to long-term glucocorticoid therapy. Bone Miner. 1990, 8, 39–51. [Google Scholar]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carnovali, M.; Valli, R.; Banfi, G.; Porta, G.; Mariotti, M. Soybean Meal-Dependent Intestinal Inflammation Induces Different Patterns of Bone-Loss in Adult Zebrafish Scale. Biomedicines 2021, 9, 393. https://doi.org/10.3390/biomedicines9040393
Carnovali M, Valli R, Banfi G, Porta G, Mariotti M. Soybean Meal-Dependent Intestinal Inflammation Induces Different Patterns of Bone-Loss in Adult Zebrafish Scale. Biomedicines. 2021; 9(4):393. https://doi.org/10.3390/biomedicines9040393
Chicago/Turabian StyleCarnovali, Marta, Roberto Valli, Giuseppe Banfi, Giovanni Porta, and Massimo Mariotti. 2021. "Soybean Meal-Dependent Intestinal Inflammation Induces Different Patterns of Bone-Loss in Adult Zebrafish Scale" Biomedicines 9, no. 4: 393. https://doi.org/10.3390/biomedicines9040393