

Propentofylline Improves Thiol-Based Antioxidant Defenses and Limits Lipid Peroxidation following Gliotoxic Injury in the Rat Brainstem

 ,
,
Abstract
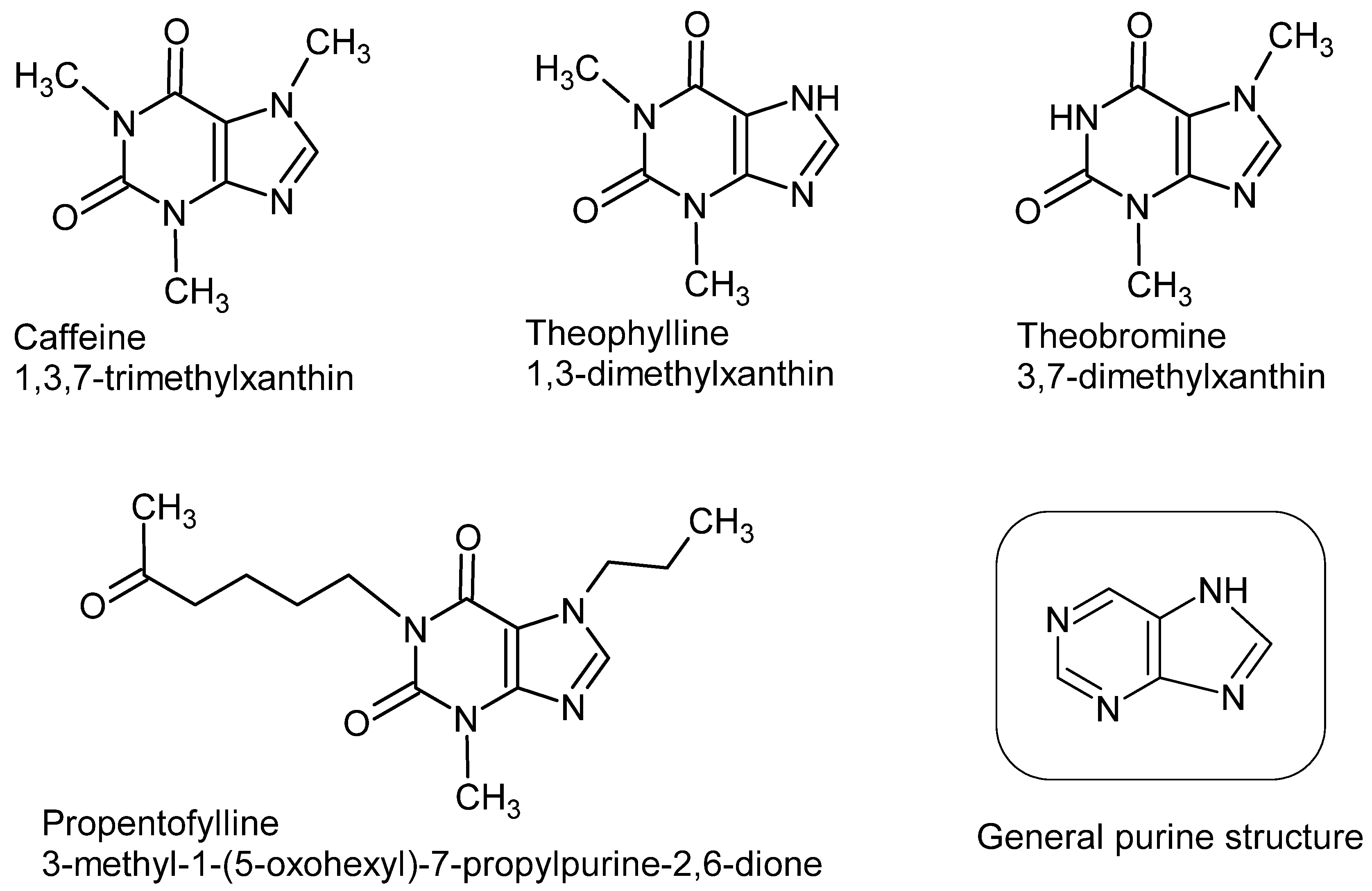
:1. Introduction
2. Materials and Methods
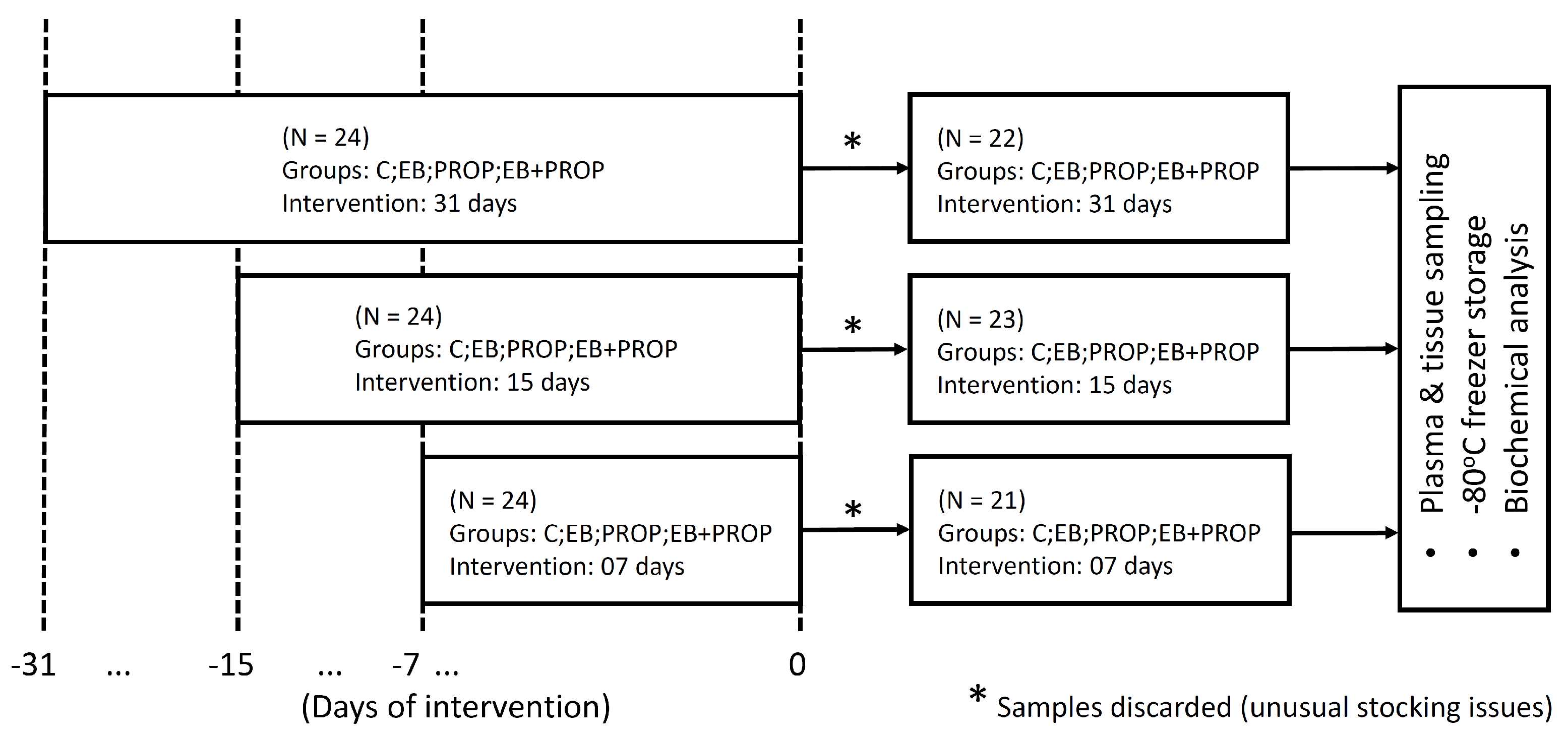
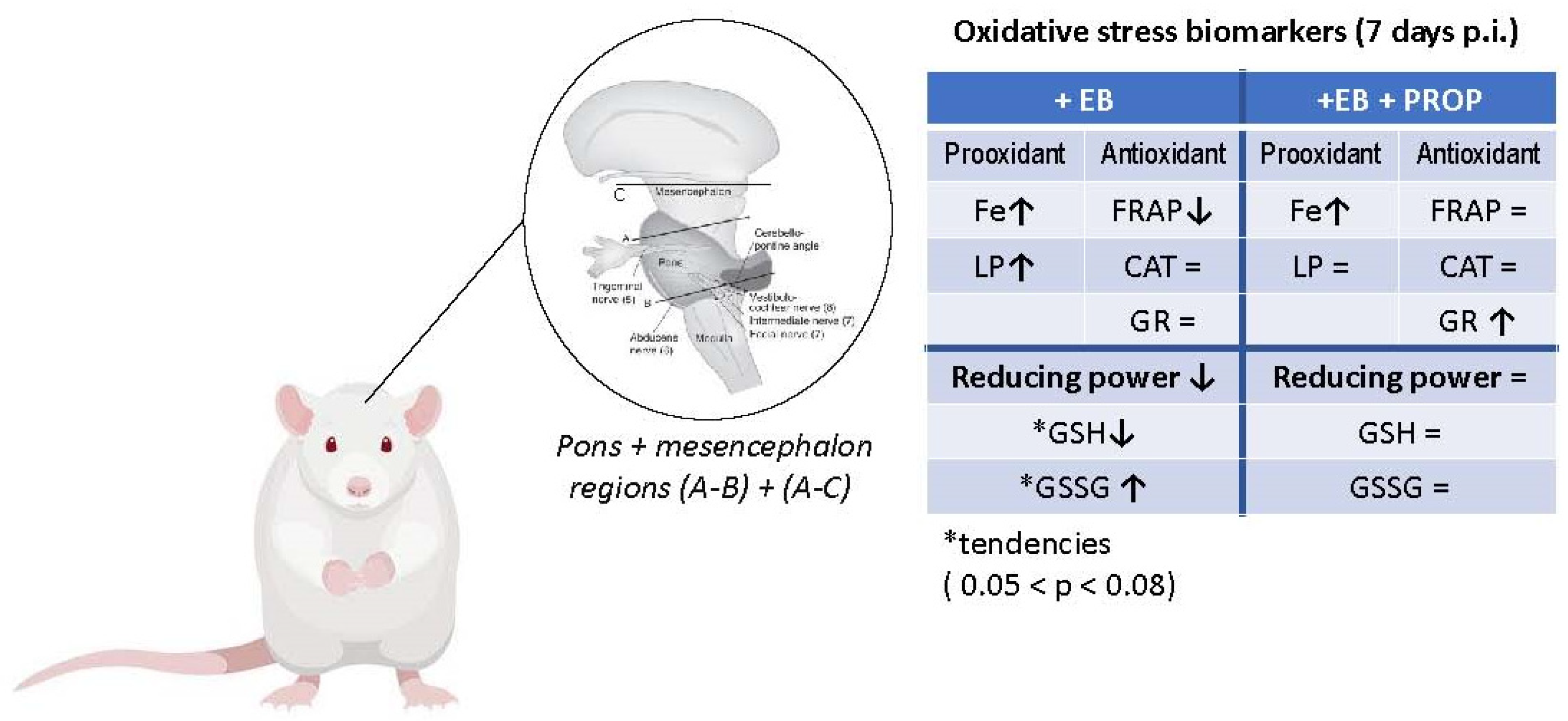
2.1. Animals
2.2. Plasma and Tissue Samples
2.3. Biochemical Analyses
2.4. Statistical Analyses
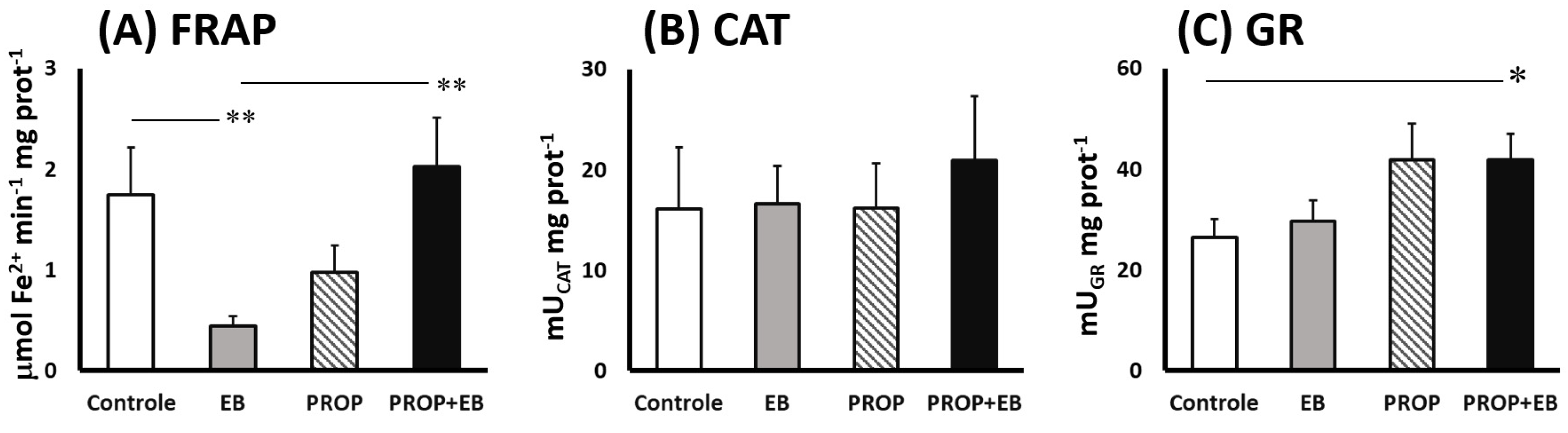
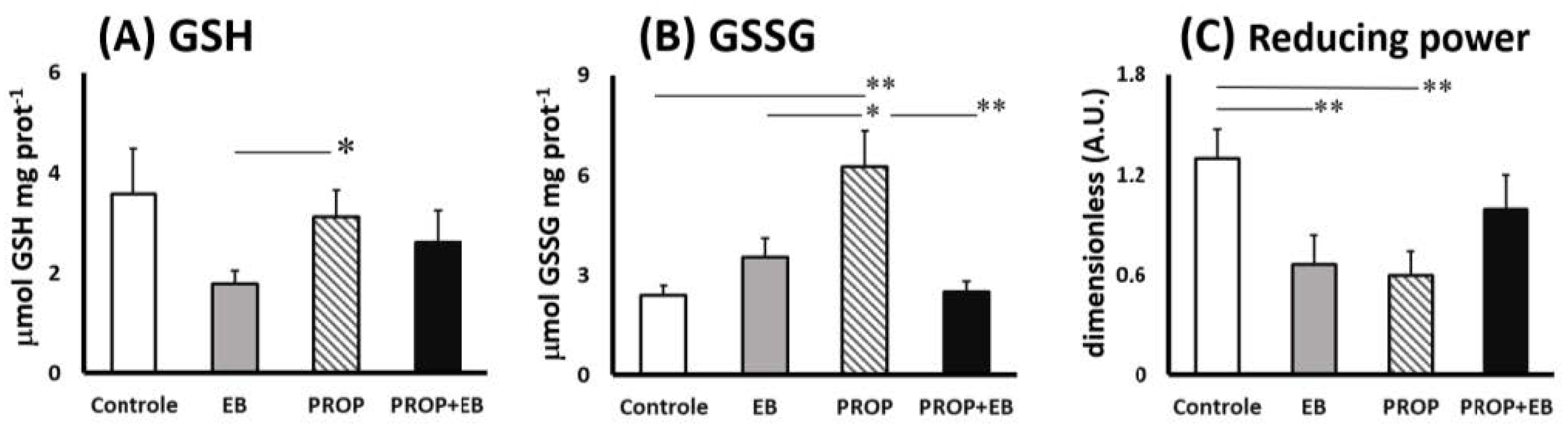
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Franco, R.; Oñatibia-Astibia, A.; Martínez-Pinilla, E. Health benefits of methylxanthines in cacao and chocolate. Nutrients 2013, 5, 4159–4173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goya, L.; Kongor, J.E.; de Pascual-Teresa, S. From Cocoa to Chocolate: Effect of Processing on Flavanols and Methylxanthines and Their Mechanisms of Action. Int. J. Mol. Sci. 2022, 23, 14365. [Google Scholar] [CrossRef] [PubMed]
- Janitschke, D.; Lauer, A.A.; Bachmann, C.M.; Winkler, J.; Griebsch, L.V.; Pilz, S.M.; Theiss, E.L.; Grimm, H.S.; Hartmann, T.; Grimm, M.O.W. Methylxanthines Induce a Change in the AD/Neurodegeneration-Linked Lipid Profile in Neuroblastoma Cells. Int. J. Mol. Sci. 2022, 23, 2295. [Google Scholar] [CrossRef] [PubMed]
- Noschang, C.G.; Krolow, R.; Pettenuzzo, L.F.; Avila, M.C.; Fachin, A.; Arcego, D.; von Pozzer Toigo, E.; Crema, L.M.; Diehl, L.A.; Vendite, D.; et al. Interactions between chronic stress and chronic consumption of caffeine on the enzymatic antioxidant system. Neurochem. Res. 2009, 34, 1568–1574. [Google Scholar] [CrossRef]
- Gołembiowska, K.; Wardas, J.; Noworyta-Sokołowska, K.; Kamińska, K.; Górska, A. Effects of adenosine receptor antagonists on the in vivo LPS-induced inflammation model of Parkinson’s disease. Neurotox. Res. 2013, 24, 29–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sigel, A.; Sigel, H.; Sigel, R.K.O. Coordination Chemistry of Nucleotides and Antivirally Active Acyclic Nucleoside Phosphonates, including Mechanistic Considerations. Molecules 2022, 27, 2625. [Google Scholar] [CrossRef]
- Ichiyama, T.; Hasegawa, S.; Matsubara, T.; Hayashi, T.; Furukawa, S. Theophylline inhibits NF-kappa B activation and I kappa B alpha degradation in human pulmonary epithelial cells. Naunyn Schmiedebergs Arch. Pharmacol. 2001, 364, 558–561. [Google Scholar] [CrossRef]
- Salim, S. Oxidative Stress and the Central Nervous System. J. Pharmacol. Exp. Ther. 2017, 360, 201–205. [Google Scholar] [CrossRef] [Green Version]
- Cobley, J.N.; Fiorello, M.L.; Bailey, D.M. 13 reasons why the brain is susceptible to oxidative stress. Redox Biol. 2018, 15, 490–503. [Google Scholar] [CrossRef] [PubMed]
- Andersen, H.H.; Johnsen, K.B.; Moos, T. Iron deposits in the chronically inflamed central nervous system and contributes to neurodegeneration. Cell. Mol. Life Sci. 2014, 71, 1607–1622. [Google Scholar] [CrossRef] [Green Version]
- Grimm, A.; Eckert, A. Brain aging and neurodegeneration: From a mitochondrial point of view. J. Neurochem. 2017, 143, 418–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bondan, E.F.; Martins, M.F.; Dossa, P.D.; Viebig, L.B.; Cardoso, C.V.; Martins, J.L., Jr.; Bernardi, M.M. Propentofylline reduces glial scar development following gliotoxic damage in the rat brainstem. Arq. Neuropsiquiatr. 2016, 74, 730–736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bondan, E.F.; Martins, M.F.; Baliellas, M.D.E.; Gimenez, M.C.F.; Poppe, C.S.; Bernardi, M.M. Effects of propentofylline on CNS remyelination in the rat brainstem. Microsc. Res. Tech. 2014, 77, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Bondan, E.F.; Martins, M.F.; Bernardi, M.M. Propentofylline reverses delayed remyelination in streptozotocin-induced diabetic rats. Arch. Endocrinol. Metab. 2015, 59, 47–53. [Google Scholar] [CrossRef] [Green Version]
- Bacchieri, A.; Cioppa, G.D. Fundamentals of Clinical Research: Bridging Medicine, Statistics and Operations; Springer: Milano, Italy, 2007. [Google Scholar]
- Allen, G.V.; Hopkins, D.A. Topography and synaptology of mamillary body projections to the mesencephalon and pons in the rat. J. Comp. Neurol. 1990, 301, 214–231. [Google Scholar] [CrossRef]
- Christensen, S.D.; Mikkelsen, L.F.; Fels, J.J.; Bodvarsdóttir, T.B.; Hansen, A.K. Quality of plasma sampled by different methods for multiple blood sampling in mice. Lab. Animals. 2009, 43, 65–71. [Google Scholar] [CrossRef] [Green Version]
- Aebi, H. Catalase in vitro. Method. Enzymol. 1984, 105, 121–126. [Google Scholar]
- Carlberg, I.; Mannervik, B. Glutathione reductase. Methods Enzymol. 1985, 113, 484–490. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [Green Version]
- Brewer, K.J.; Murphy, W.R., Jr.; Petersen, J.D. Synthesis and characterization of monometallic and bimetallic mixed-ligand complexes of iron(II) containing 2,2′-bipyrimidine or 2,3-bis(2-pyridyl)pyrazine. Inorg. Chem. 1987, 26, 3376–3379. [Google Scholar] [CrossRef]
- Goodwin, J.F.; Murphy, B. The colorimetric determination of iron in biological material with reference to its measurement during chelation therapy. Clin. Chem. 1966, 12, 58–69. [Google Scholar] [CrossRef]
- Rahman, I.; Kode, A.; Biswas, S.K. Assay for quantitative determination of glutathione and glutathione disulfide levels using enzymatic recycling method. Nat. Protoc. 2006, 1, 3159–3165. [Google Scholar] [CrossRef]
- Fraga, C.G.; Leibovitz, B.E.; Tappel, A.L. Lipid peroxidation measured as thiobarbituric acid-reactive substances in tissue slices: Characterization and comparison with homogenates and microsomes. Free Radic. Biol. Med. 1988, 4, 155–161. [Google Scholar] [CrossRef]
- Hoaglin, D.C.; Iglewicz, B. Fine-tuning some resistant rules for outlier labeling. J. Am. Stat. Assoc. 1987, 82, 1147–1149. [Google Scholar] [CrossRef]
- Dixon, W.J. Efficient analysis of experimental observations. Annu. Rev. Pharmacol. Toxicol. 1980, 20, 441–446. [Google Scholar] [CrossRef]
- Janitschke, D.; Lauer, A.A.; Bachmann, C.M.; Grimm, H.S.; Hartmann, T.; Grimm, M.O.W. Methylxanthines and Neurodegenerative Diseases: An Update. Nutrients 2021, 13, 803. [Google Scholar] [CrossRef] [PubMed]
- Polotow, T.G.; Poppe, S.C.; Vardaris, C.V.; Ganini, D.; Guariroba, M.; Mattei, R.; Hatanaka, E.; Martins, M.F.; Bondan, E.F.; Barros, M.P. Redox Status and Neuro Inflammation Indexes in Cerebellum and Motor Cortex of Wistar Rats Supplemented with Natural Sources of Omega-3 Fatty Acids and Astaxanthin: Fish Oil, Krill Oil, and Algal Biomass. Mar. Drugs. 2015, 13, 6117–6137. [Google Scholar] [CrossRef] [Green Version]
- Quincozes-Santos, A.; Santos, C.L.; de Souza Almeida, R.R.; da Silva, A.; Thomaz, N.K.; Costa, N.L.F.; Weber, F.B.; Schmitz, I.; Medeiros, L.S.; Medeiros, L.; et al. Gliotoxicity and Glioprotection: The Dual Role of Glial Cells. Mol. Neurobiol. 2021, 58, 6577–6592. [Google Scholar] [CrossRef]
- Ju, Q.; Li, X.; Zhang, H.; Yan, S.; Li, Y.; Zhao, Y. NFE2L2 Is a Potential Prognostic Biomarker and Is Correlated with Immune Infiltration in Brain Lower Grade Glioma: A Pan-Cancer Analysis. Oxid. Med. Cell. Longev. 2020, 2020, 3580719. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Haldar, S.; Tripathi, A.K.; Horback, K.; Wong, J.; Sharma, D.; Beserra, A.; Suda, S.; Anbalagan, C.; Dev, S.; et al. Brain iron homeostasis: From molecular mechanisms to clinical significance and therapeutic opportunities. Antioxid. Redox Signal. 2014, 20, 1324–1363. [Google Scholar] [CrossRef] [Green Version]
- Zhang, N.; Yu, X.; Xie, J.; Xu, H. New Insights into the Role of Ferritin in Iron Homeostasis and Neurodegenerative Diseases. Mol. Neurobiol. 2021, 58, 2812–2823. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Jankovic, J.; Le, W. Iron chelation and neuroprotection in neurodegenerative diseases. J. Neural Transm. 2011, 118, 473–477. [Google Scholar] [CrossRef] [PubMed]
- Aoyama, K. Glutathione in the Brain. Int. J. Mol. Sci. 2021, 22, 5010. [Google Scholar] [CrossRef] [PubMed]
- Shukla, V.; Gude, R.P. Potentiation of antimetastatic activity of pentoxifylline in B16F10 and B16F1 melanoma cells through inhibition of glutathione content. Cancer Biother. Radiopharm. 2003, 18, 559–564. [Google Scholar] [CrossRef] [PubMed]
- Bhat, J.A.; Gupta, S.; Kumar, M. Neuroprotective effects of theobromine in transient global cerebral ischemia-reperfusion rat model. Biochem. Biophys. Res. Commun. 2021, 571, 74–80. [Google Scholar] [CrossRef]
- Shukla, V.; Gude, R.P. Amelioration of B16F10 melanoma cells induced oxidative stress in DBA/2 mice by pentoxifylline. J. Exp. Clin. Cancer Res. 2003, 22, 407–410. [Google Scholar]
- Kalthoff, S.; Ehmer, U.; Freiberg, N.; Manns, M.P.; Strassburg, C.P. Coffee induces expression of glucuronosyltransferases by the aryl hydrocarbon receptor and Nrf2 in liver and stomach. Gastroenterology 2010, 139, 1699–1710.e2. [Google Scholar] [CrossRef]
- Sugimoto, N.; Miwa, S.; Hitomi, Y.; Nakamura, H.; Tsuchiya, H.; Yachie, A. Theobromine, the primary methylxanthine found in Theobroma cacao, prevents malignant glioblastoma proliferation by negatively regulating phosphodiesterase-4, extracellular signal-regulated kinase, Akt/mammalian target of rapamycin kinase, and nuclear factor-kappa B. Nutr. Cancer 2014, 66, 419–423. [Google Scholar] [CrossRef]
- Lu, S.C. Glutathione synthesis. Biochim. Biophys. Acta 2013, 1830, 3143–3153. [Google Scholar] [CrossRef] [Green Version]
- Brodal, P. Encyclopedia of the Neurological Sciences, 2nd ed.; Aminoff, M.J., Daroff, R.B., Eds.; Academic Press: Cambridge, MA, USA, 2014; pp. 936–937. ISBN 9780123851581. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | EB | PROP | PROP + EB | |
|---|---|---|---|---|
| (i) Plasma | ||||
| TBARS (μmol·mL−1) | ||||
| 7 d | 13.1 ± 2.4 | 20.2 ± 2.4 (*) | 11.8 ± 0.9 | 15.4 ± 1.5 |
| 15 d | n.d. | 21.0 ± 2.9 (*) | 15.1 ± 2.6 | 19.7 ± 6.4 (*) |
| 31 d | n.d. | 10.8 ± 1.1 | 12.9 ± 2.6 | 12.4 ± 2.1 |
| (ii) Brainstem tissue | ||||
| TBARS (μmol·mg prot−1) | ||||
| 7 d | 0.64 ± 0.04 | 1.28 ± 0.27 (**) | 0.54 ± 0.06 | 0.72 ± 0.09 |
| 15 d | n.d. | 1.55 ± 0.28 (**) | 0.63 ± 0.05 | 0.88 ± 0.05 (*) |
| 31 d | n.d. | 1.06 ± 0.13 (**) | 0.52 ± 0.05 | 0.72 ± 0.07 |
| Iron content (μg·mg prot−1) | ||||
| 7 d | 88.2 ± 10.5 | 279.0 ± 60.6 (**) | 75.8 ± 24.7 | 214.5 ± 52.4 (**) |
| 15 d | n.d. | 413.4 ± 31.6 (**) | 433.3 ± 69.2 (**) | 409.7 ± 41.8 (**) |
| 31 d | n.d. | 285.3 ± 50.8 (**) | 261.0 ± 35.2 (**) | 279.6± 30.8 (**) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baliellas, D.E.M.; Barros, M.P.; Vardaris, C.V.; Guariroba, M.; Poppe, S.C.; Martins, M.F.; Pereira, Á.A.F.; Bondan, E.F. Propentofylline Improves Thiol-Based Antioxidant Defenses and Limits Lipid Peroxidation following Gliotoxic Injury in the Rat Brainstem. Biomedicines 2023, 11, 1652. https://doi.org/10.3390/biomedicines11061652
Baliellas DEM, Barros MP, Vardaris CV, Guariroba M, Poppe SC, Martins MF, Pereira ÁAF, Bondan EF. Propentofylline Improves Thiol-Based Antioxidant Defenses and Limits Lipid Peroxidation following Gliotoxic Injury in the Rat Brainstem. Biomedicines. 2023; 11(6):1652. https://doi.org/10.3390/biomedicines11061652
Chicago/Turabian StyleBaliellas, Deborah E. M., Marcelo P. Barros, Cristina V. Vardaris, Maísa Guariroba, Sandra C. Poppe, Maria F. Martins, Álvaro A. F. Pereira, and Eduardo F. Bondan. 2023. "Propentofylline Improves Thiol-Based Antioxidant Defenses and Limits Lipid Peroxidation following Gliotoxic Injury in the Rat Brainstem" Biomedicines 11, no. 6: 1652. https://doi.org/10.3390/biomedicines11061652