Divergent Roles of CYP26B1 and Endogenous Retinoic Acid in Mouse Fetal Gonads

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Plasmids
2.3. Cell Lines and Plasmid Transfection
2.4. Electroporation of Fetal Gonads
2.5. Organotypic Culture
2.6. Measure of Testosterone Production
2.7. Measure of Sertoli Cell Proliferation
2.8. RNA Extraction and RT-qPCR
2.9. Purification of Fetal Germ and Somatic Cells
2.10. Histology and Immunohistochemistry
3. Results
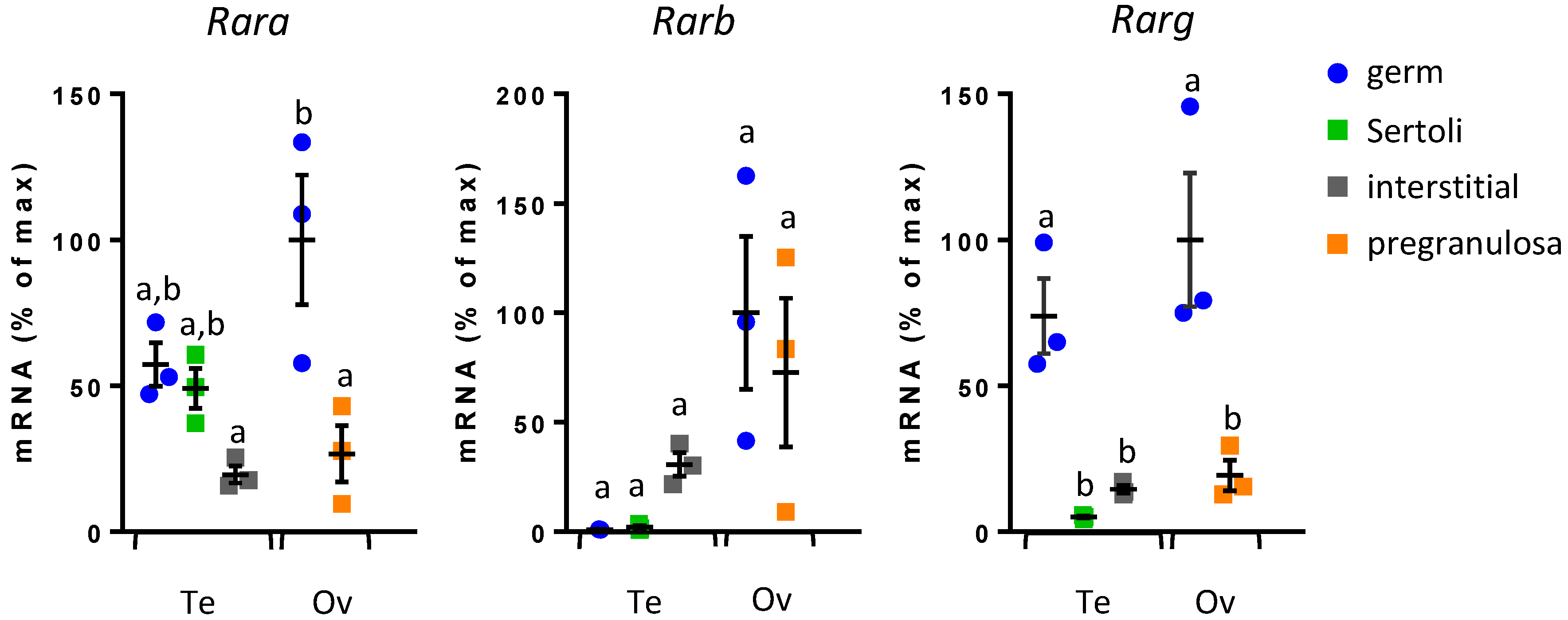
3.1. Rar Expression in Fetal Gonadal Cells
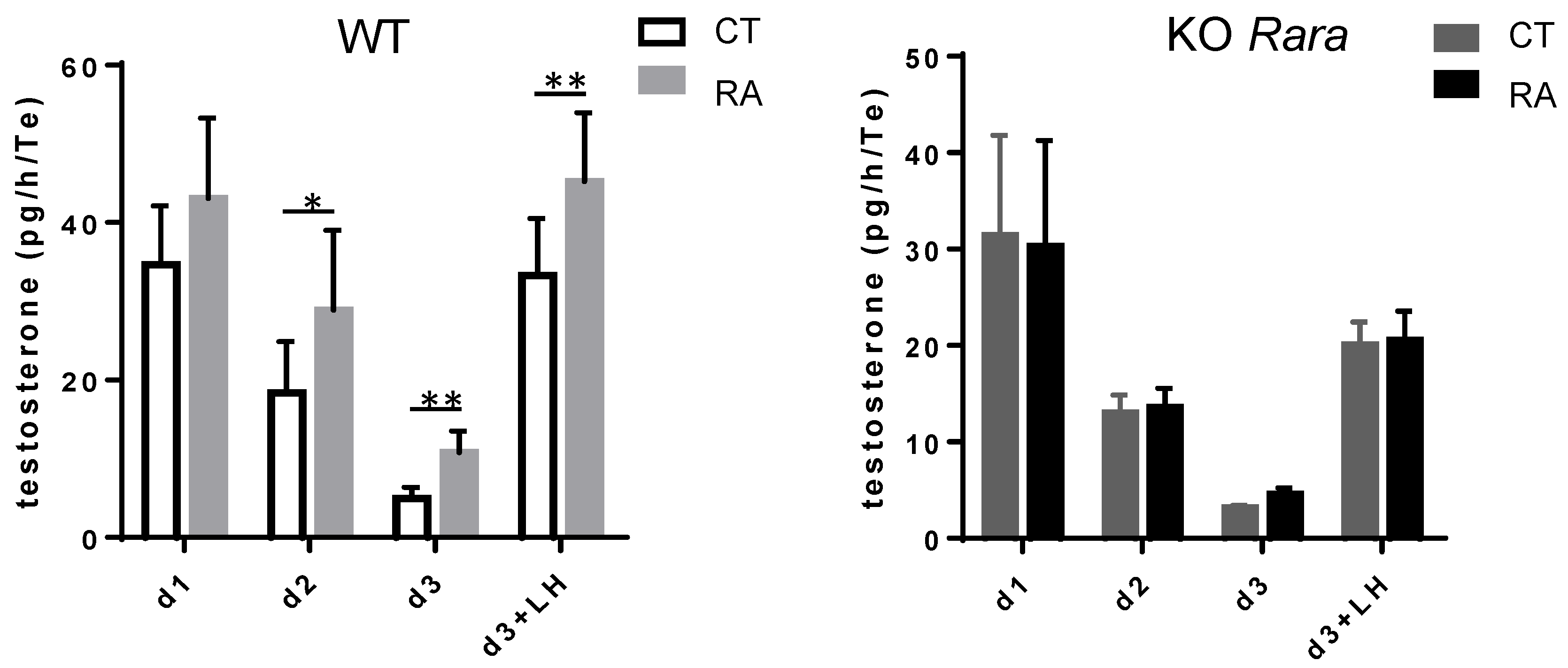
3.2. RA Increases Mouse Fetal Testosterone Production
3.3. RA Inhibits Sertoli Cell Proliferation
3.4. Cyp26b1 Invalidation Does Not Inhibit Sertoli Cell Proliferation
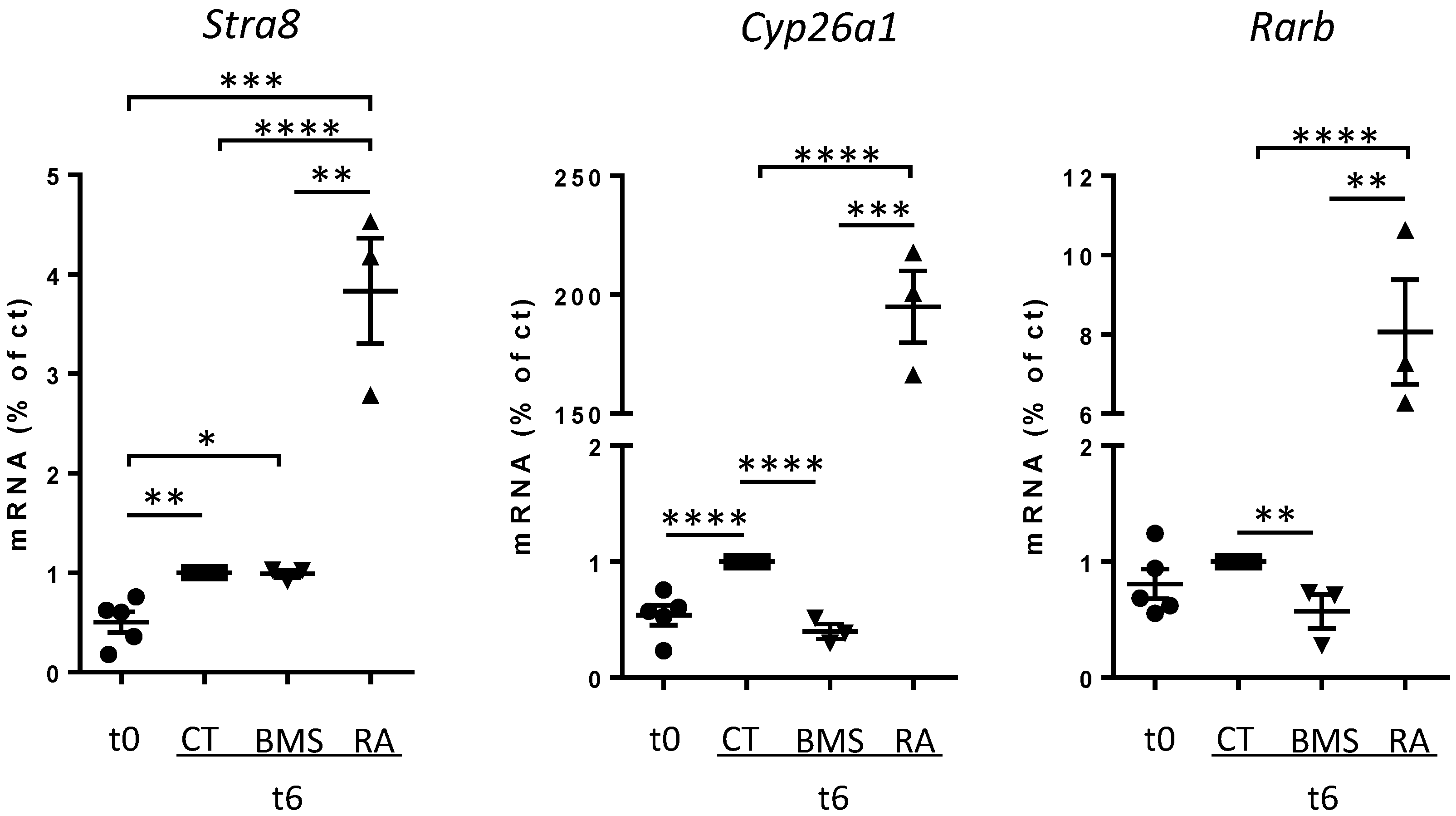
3.5. Short-Term Inhibition of RA Signaling Does Not Affect Stra8 Expression
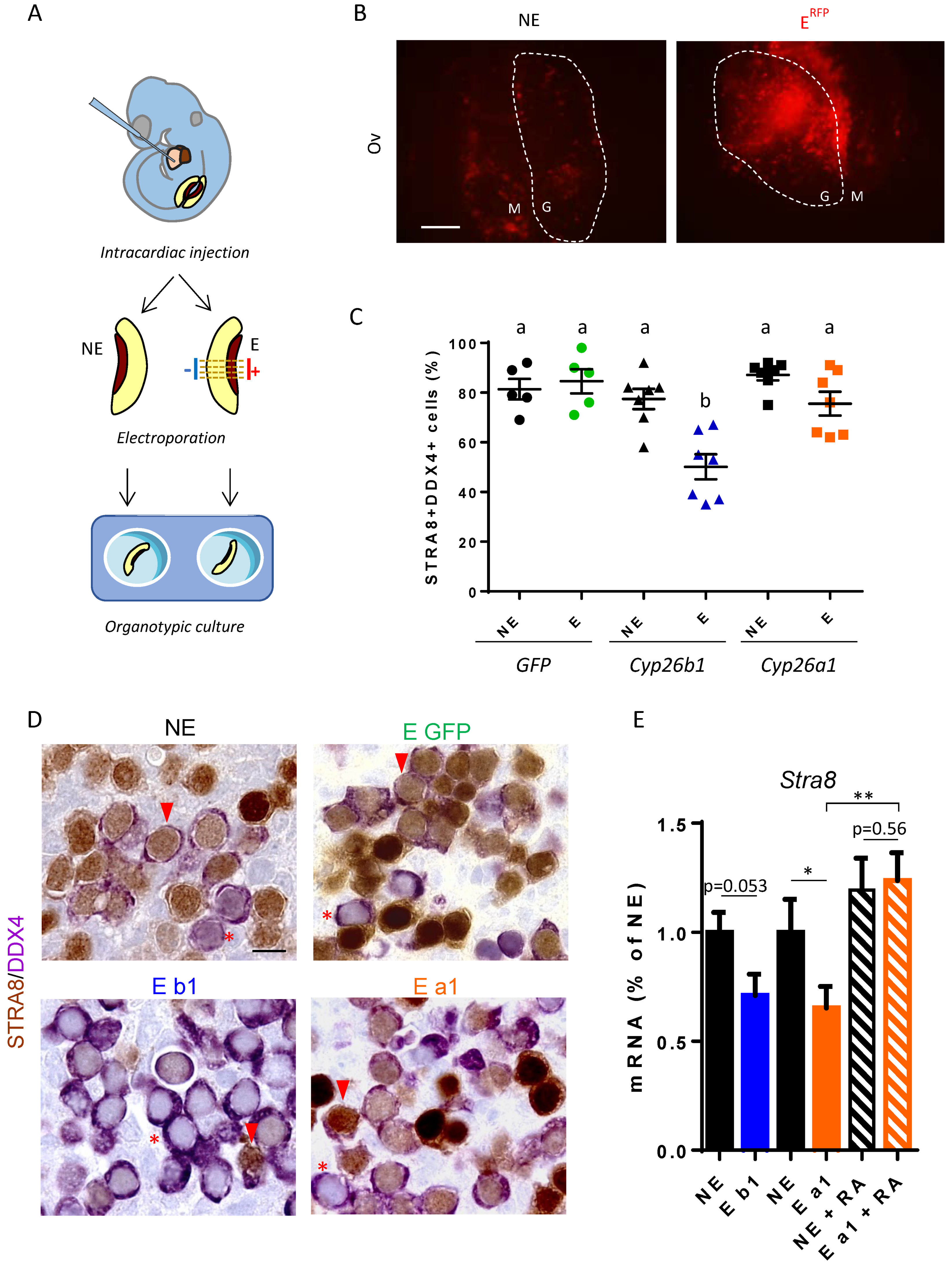
3.6. Ectopic Expression of CYP26B1 or CYP26A1 Differentially Regulates STRA8 in Fetal Ovaries
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Feng, C.-W.; Bowles, J.; Koopman, P. Control of mammalian germ cell entry into meiosis. Mol. Cell. Endocrinol. 2014, 382, 488–497. [Google Scholar] [CrossRef]
- Guerquin, M.-J.; Duquenne, C.; Lahaye, J.-B.; Tourpin, S.; Habert, R.; Livera, G. New testicular mechanisms involved in the prevention of fetal meiotic initiation in mice. Dev. Boil. 2010, 346, 320–330. [Google Scholar] [CrossRef] [Green Version]
- McLaren, A. Primordial germ cells in the mouse. Dev. Biol. 2003, 262, 1–15. [Google Scholar] [Green Version]
- Koubova, J.; Menke, D.B.; Zhou, Q.; Capel, B.; Griswold, M.D.; Page, D.C. Retinoic acid regulates sex-specific timing of meiotic initiation in mice. PNAS 2006, 103, 2474–2479. [Google Scholar] [CrossRef] [Green Version]
- E Baltus, A.; Menke, D.B.; Hu, Y.-C.; Goodheart, M.L.; E Carpenter, A.; De Rooij, D.G.; Page, D.C. In germ cells of mouse embryonic ovaries, the decision to enter meiosis precedes premeiotic DNA replication. Nat. Genet. 2006, 38, 1430–1434. [Google Scholar] [CrossRef]
- Anderson, E.L.; Baltus, A.E.; Roepers-Gajadien, H.L.; Hassold, T.J.; De Rooij, D.G.; Van Pelt, A.M.M.; Page, D.C. Stra8 and its inducer, retinoic acid, regulate meiotic initiation in both spermatogenesis and oogenesis in mice. PNAS 2008, 105, 14976–14980. [Google Scholar] [CrossRef]
- Mark, M.; Jacobs, H.; Oulad-Abdelghani, M.; Dennefeld, C.; Feret, B.; Vernet, N.; Codreanu, C.-A.; Chambon, P.; Ghyselinck, N. STRA8-deficient spermatocytes initiate, but fail to complete, meiosis and undergo premature chromosome condensation. J. Cell Sci. 2008, 121, 3233–3242. [Google Scholar] [CrossRef] [Green Version]
- Oulad-Abdelghani, M.; Bouillet, P.; Décimo, D.; Gansmuller, A.; Heyberger, S.; Dollé, P.; Bronner, S.; Lutz, Y.; Chambon, P. Characterization of a premeiotic germ cell-specific cytoplasmic protein encoded by Stra8, a novel retinoic acid-responsive gene. J. Cell Biol. 1996, 135, 469–477. [Google Scholar] [CrossRef]
- Bowles, J. Retinoid Signaling Determines Germ Cell Fate in Mice. Science 2006, 312, 596–600. [Google Scholar] [CrossRef] [Green Version]
- Trautmann, E.; Guerquin, M.-J.; Duquenne, C.; Lahaye, J.-B.; Habert, R.; Livera, G. Retinoic acid prevents germ cell mitotic arrest in mouse fetal testes. Cell Cycle 2008, 7, 656–664. [Google Scholar] [CrossRef] [Green Version]
- Niederreither, K.; Vermot, J.; Fraulob, V.; Chambon, P.; Dollé, P. Retinaldehyde dehydrogenase 2 (RALDH2)- independent patterns of retinoic acid synthesis in the mouse embryo. PNAS 2002, 99, 16111–16116. [Google Scholar] [CrossRef] [Green Version]
- Bowles, J.; Feng, C.-W.; Miles, K.; Ineson, J.; Spiller, C.; Koopman, P. ALDH1A1 provides a source of meiosis-inducing retinoic acid in mouse fetal ovaries. Nat. Commun. 2016, 7, 10845. [Google Scholar] [CrossRef] [Green Version]
- Saba, R.; Wu, Q.; Saga, Y. CYP26B1 promotes male germ cell differentiation by suppressing STRA8-dependent meiotic and STRA8-independent mitotic pathways. Dev. Boil. 2014, 389, 173–181. [Google Scholar] [CrossRef] [Green Version]
- MacLean, G.; Li, H.; Metzger, D.; Chambon, P.; Petkovich, M. Apoptotic Extinction of Germ Cells in Testes of Cyp26b1 Knockout Mice. Endocrinology 2007, 148, 4560–4567. [Google Scholar] [CrossRef] [Green Version]
- Bowles, J.; Feng, C.-W.; Spiller, C.; Davidson, T.-L.; Jackson, A.; Koopman, P. FGF9 Suppresses Meiosis and Promotes Male Germ Cell Fate in Mice. Dev. Cell 2010, 19, 440–449. [Google Scholar] [CrossRef] [Green Version]
- Pasquier, J.; Cabau, C.; Nguyen, T.; Jouanno, E.; Severac, D.; Braasch, I.; Journot, L.; Pontarotti, P.; Klopp, C.; Postlethwait, J.H.; et al. Gene evolution and gene expression after whole genome duplication in fish: The PhyloFish database. BMC Genom. 2016, 17, 13698. [Google Scholar] [CrossRef]
- Frydman, N.A.; Poulain, M.; Arkoun, B.; Duquenne, C.; Tourpin, S.; Messiaen, S.; Habert, R.; Rouiller-Fabre, V.; Benachi, A.; Livera, G. Human foetal ovary shares meiotic preventing factors with the developing testis. Hum. Reprod. 2017, 32, 631–642. [Google Scholar] [CrossRef]
- Le Bouffant, R.; Souquet, B.; Duval, N.; Duquenne, C.; Hervé, R.; Frydman, N.A.; Robert, B.; Habert, R.; Livera, G. Msx1 and Msx2 promote meiosis initiation. Development 2011, 138, 5393–5402. [Google Scholar] [CrossRef]
- Le Bouffant, R.; Guerquin, M.J.; Duquenne, C.; Frydman, N.A.; Coffigny, H.; Rouiller-Fabre, V.; Frydman, R.; Habert, R.; Livera, G. Meiosis initiation in the human ovary requires intrinsic retinoic acid synthesis. Hum. Reprod. 2010, 25, 2579–2590. [Google Scholar] [CrossRef] [Green Version]
- Jorgensen, A.; Nielsen, J.; Perlman, S.; Lundvall, L.; Mitchell, R.; Juul, A.; Meyts, E.R.-D. Ex vivo culture of human fetal gonads: Manipulation of meiosis signalling by retinoic acid treatment disrupts testis development. Hum. Reprod. 2015, 30, 2351–2363. [Google Scholar] [CrossRef]
- Li, H.; Kim, K.H. Retinoic Acid Inhibits Rat XY Gonad Development by Blocking Mesonephric Cell Migration and Decreasing the Number of Gonocytes. Boil. Reprod. 2004, 70, 687–693. [Google Scholar] [CrossRef]
- Livera, G.; Rouiller-Fabre, V.; Durand, P.; Habert, R. Multiple Effects of Retinoids on the Development of Sertoli, Germ, and Leydig Cells of Fetal and Neonatal Rat Testis in Culture1. Boil. Reprod. 2000, 62, 1303–1314. [Google Scholar] [CrossRef] [Green Version]
- Best, D.; Sahlender, D.A.; Walther, N.; Peden, A.A.; Adams, I.R. Sdmg1 is a conserved transmembrane protein associated with germ cell sex determination and germline-soma interactions in mice. Development 2008, 135, 1415–1425. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Chatzi, C.; Brade, T.; Cunningham, T.J.; Zhao, X.; Duester, G. Sex-specific timing of meiotic initiation is regulated by Cyp26b1 independent of retinoic acid signalling. Nat. Commun. 2011, 2, 151. [Google Scholar] [CrossRef]
- Dollé, P.; Ruberte, E.; Leroy, P.; Morriss-Kay, G.; Chambon, P. Retinoic acid receptors and cellular retinoid binding proteins. I. A systematic study of their differential pattern of transcription during mouse organogenesis. Development 1990, 110, 1133–1151. [Google Scholar]
- Ruberte, E.; Dollé, P.; Krust, A.; Zelent, A.; Morriss-Kay, G.; Chambon, P. Specific spatial and temporal distribution of retinoic acid receptor gamma transcripts during mouse embryogenesis. Development 1990, 108, 213–222. [Google Scholar]
- Ohbo, K.; Yoshida, S.; Ohmura, M.; Ohneda, O.; Ogawa, T.; Tsuchiya, H.; Kuwana, T.; Kehler, J.; Abe, K.; Schöler, H.R.; et al. Identification and characterization of stem cells in prepubertal spermatogenesis in mice. Dev. Boil. 2003, 258, 209–225. [Google Scholar] [CrossRef] [Green Version]
- Wojtasz, L.; Daniel, K.; Tóth, A. Fluorescence activated cell sorting of live female germ cells and somatic cells of the mouse fetal gonad based on forward and side scattering. Cytom. Part A 2009, 75, 547–553. [Google Scholar] [CrossRef]
- Yoshimizu, T.; Sugiyama, N.; Yeom, Y.I.; Masuko, K.; De Felice, M.; Ohbo, K.; Obinata, M.; Abe, K.; Schöler, H.R.; Matsui, Y. Germline-specific expression of the Oct-4/green fluorescent protein (GFP) transgene in mice. Dev. Growth Differ. 1999, 41, 675–684. [Google Scholar] [CrossRef]
- Lufkin, T.; Lohnes, D.; Mark, M.; Dierich, A.; Gorry, P.; Gaub, M.P.; LeMeur, M.; Chambon, P. High postnatal lethality and testis degeneration in retinoic acid receptor alpha mutant mice. PNAS 1993, 90, 7225–7229. [Google Scholar] [CrossRef]
- Livera, G.; Delbès, G.; Pairault, C.; Rouiller-Fabre, V.; Habert, R. Organotypic culture, a powerful model for studying rat and mouse fetal testis development. Cell Tissue Res. 2006, 324, 507–521. [Google Scholar] [CrossRef]
- Habert, R.; Picon, R. Testosterone, dihydrotestosterone and estradiol-17β levels in maternal and fetal plasma and in fetal testes in the rat. J. Steroid Biochem. 1984, 21, 193–198. [Google Scholar] [CrossRef]
- Lambrot, R.; Pairault, C.; Donnadieu, A.-C.; Rouiller-Fabre, V.; Coffigny, H.; Frydman, R.; Habert, R. Use of Organ Culture to Study the Human Fetal Testis Development: Effect of Retinoic Acid. J. Clin. Endocrinol. Metab. 2006, 91, 2696–2703. [Google Scholar] [CrossRef] [Green Version]
- Livera, G.; Rouiller-Fabre, V.; Habert, R. Retinoid Receptors Involved in the Effects of Retinoic Acid on Rat Testis Development1. Boil. Reprod. 2001, 64, 1307–1314. [Google Scholar] [CrossRef]
- Bowles, J.; Feng, C.-W.; Ineson, J.; Miles, K.; Spiller, C.M.; Harley, V.R.; Sinclair, A.H.; Koopman, P. Retinoic Acid Antagonizes Testis Development in Mice. Cell Rep. 2018, 24, 1330–1341. [Google Scholar] [CrossRef] [Green Version]
- Topletz, A.R.; Thatcher, J.E.; Zelter, A.; Lutz, J.D.; Tay, S.; Nelson, W.L.; Isoherranen, N. Comparison of the Function and Expression of CYP26A1 and CYP26B1, the two Retnoic Acid Hydroxylases. Biochem. Pharmacol. 2013, 83, 149–163. [Google Scholar] [CrossRef]
- MacLean, G.; Abu-Abed, S.; Dollé, P.; Tahayato, A.; Chambon, P.; Petkovich, M. Cloning of a novel retinoic-acid metabolizing cytochrome P450, Cyp26B1, and comparative expression analysis with Cyp26A1 during early murine development. Mech. Dev. 2001, 107, 195–201. [Google Scholar] [CrossRef]
- Foti, R.S.; Diaz, P.; Douguet, D. Comparison of the ligand binding site of CYP2C8 with CYP26A1 and CYP26B1: A structural basis for the identification of new inhibitors of the retinoic acid hydroxylases. J. Enzym. Inhib. Med. Chem. 2016, 31, 148–161. [Google Scholar] [CrossRef]
- Vernet, N.; Mark, M.; Condrea, D.; Feret, B.; Klopfenstein, M.; Alunni, V.; Teletin, M.; Ghyselinck, N.B. Meiosis Initiates In The Fetal Ovary Of Mice Lacking All Retinoic Acid Receptor Isotypes. bioRxiv 2019. Available online: https://www.biorxiv.org/content/10.1101/716498v1 (accessed on 26 July 2019).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Species | Applications | Forward | Reverse |
|---|---|---|---|---|
| Β-Actin | mouse | RT-qPCR | 5′-GCCCTGAGGCTCTTTTCCAG-3′ | 5′-TGCCACAGGATTCCATACCC-3′ |
| Amh | mouse | RT-qPCR | 5′-TTTGGTGCTAACCGTGGACTTC-3′ | 5′-GAGCCAAATAGAAAGGCTTGCA-3′ |
| Cyp26a1 | mouse | RT-qPCR | 5′-CTCGCACAAGCAGCGAAAG-3′ | 5′-GATCACGGGCACGTAGCACT-3′ |
| Cyp26b1 | mouse | RT-qPCR | 5′-TGGACTGTGTCATCAAGGAGGT-3′ | 5′-GTCGTGAGTGTCTCGGATGCTA-3′ |
| Ddx4 | mouse | RT-qPCR | 5′-GAAGAAATCCAGAGGTTGGC-3′ | 5′-GAAGGATCGTCTGTCTGAACA-3′ |
| HSD3b | mouse | RT-qPCR | 5′-TGGTGACAGGAGCAGGA-3′ | 5′-AGGAAGCTCACAGTTTCCA-3′ |
| Rara | mouse | RT-qPCR | 5′-TGTTTCGACGTGGGCATGT-3′ | 5′-TTTGTTTCGATCGTTTCGCA-3′ |
| Rarb | mouse | RT-qPCR | 5′-TTTAATCTGTGGAGACCGCCA-3′ | 5′-TTGTCTACTTTTGTTGGTTCCTCAAG-3′ |
| Rarg | mouse | RT-qPCR | 5′-GATGGATGACACCGAGACTGG-3′ | 5′-CCACAGATGAGGCAGATAGCAC-3′ |
| Stra8 | mouse | RT-qPCR | 5′-TGAAGCTCAAAGCATCCTTCAA-3′ | 5′-CTAAGCTGTTGGGATTCCCATC-3′ |
| Name | Company | Host Species | Mono/poly | Concentration | Applications |
|---|---|---|---|---|---|
| AMH | Santa Cruz | Goat | Poly | 1/200 | IHC |
| DDX4 | Abcam | Mouse | Mono | 1/200 | IHC |
| DDX4 | Abcam | Rabbit | Poly | 1/200 | IHC |
| RFP | Abcam | Rabbit | Poly | 1/200 | IHC |
| STRA8 | Abcam | Rabbit | Poly | 1/1000 | IHC |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bellutti, L.; Abby, E.; Tourpin, S.; Messiaen, S.; Moison, D.; Trautmann, E.; Guerquin, M.-J.; Rouiller-Fabre, V.; Habert, R.; Livera, G. Divergent Roles of CYP26B1 and Endogenous Retinoic Acid in Mouse Fetal Gonads. Biomolecules 2019, 9, 536. https://doi.org/10.3390/biom9100536
Bellutti L, Abby E, Tourpin S, Messiaen S, Moison D, Trautmann E, Guerquin M-J, Rouiller-Fabre V, Habert R, Livera G. Divergent Roles of CYP26B1 and Endogenous Retinoic Acid in Mouse Fetal Gonads. Biomolecules. 2019; 9(10):536. https://doi.org/10.3390/biom9100536
Chicago/Turabian StyleBellutti, Laura, Emilie Abby, Sophie Tourpin, Sébastien Messiaen, Delphine Moison, Emilie Trautmann, Marie-Justine Guerquin, Virginie Rouiller-Fabre, René Habert, and Gabriel Livera. 2019. "Divergent Roles of CYP26B1 and Endogenous Retinoic Acid in Mouse Fetal Gonads" Biomolecules 9, no. 10: 536. https://doi.org/10.3390/biom9100536