
Into the Deep: New Data on the Lipid and Fatty Acid Profile of Redfish Sebastes mentella Inhabiting Different Depths in the Irminger Sea

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
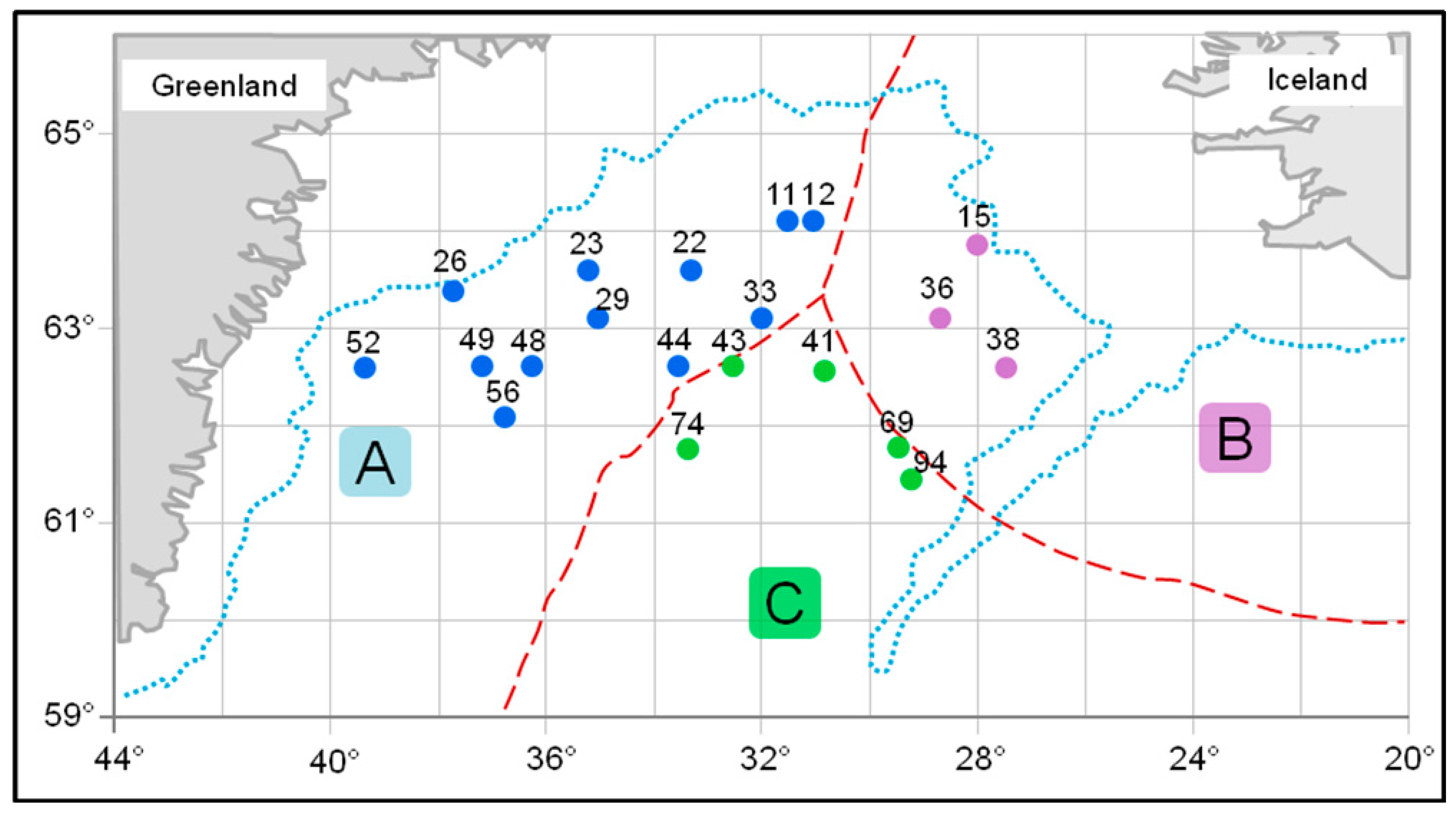
2.1. Sample Collection
2.2. Lipid Extraction and Analysis
2.2.1. Neutral Lipids Analysis
2.2.2. Polar Lipids Analysis
2.2.3. Fatty Acids Analysis
2.3. Statistical Analysis
3. Results
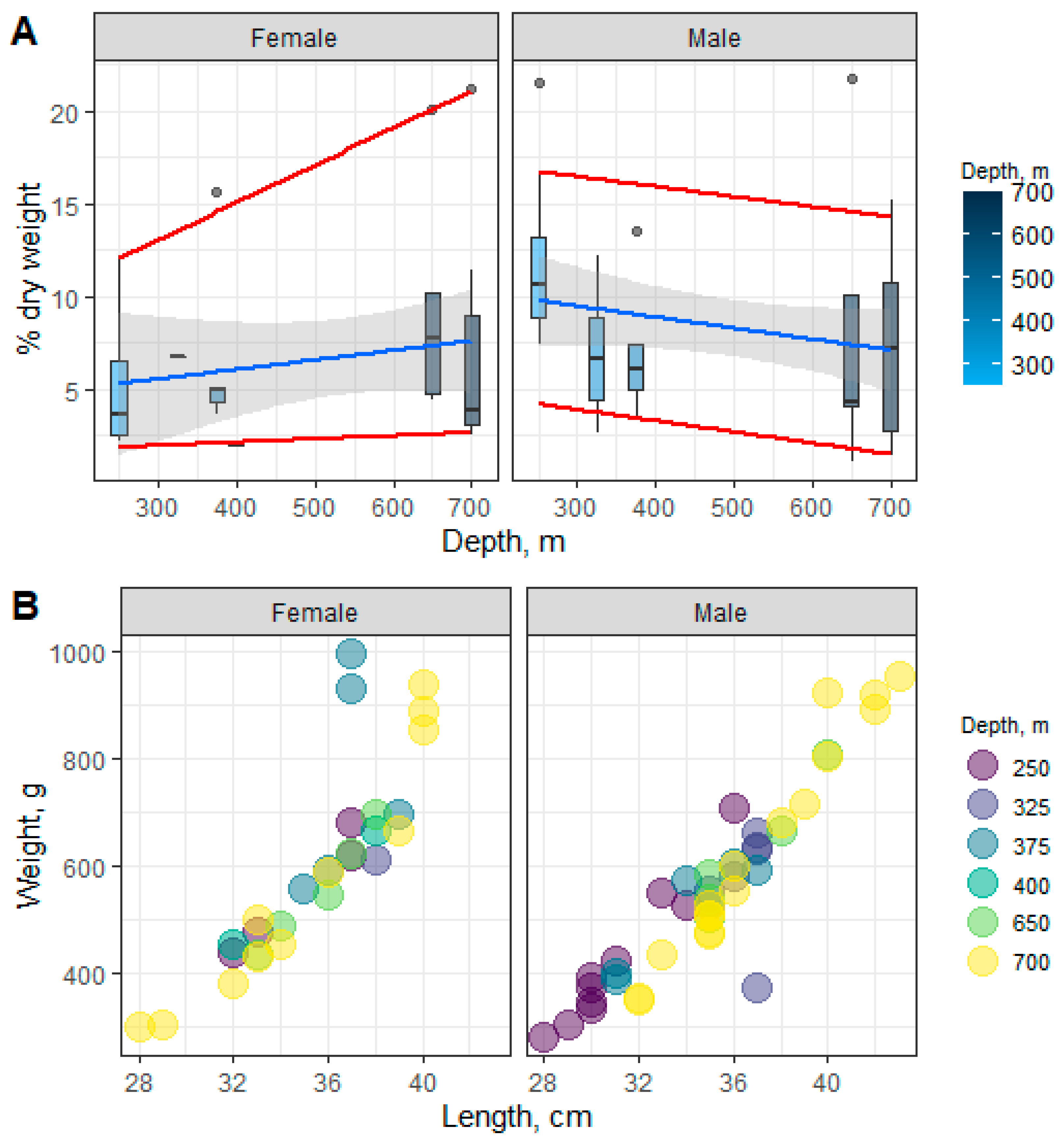
3.1. Species-Specific Characteristics of the Total Lipids (TLs) in Beaked Redfish, Their Sex Specificity, and Change Depth-Wise
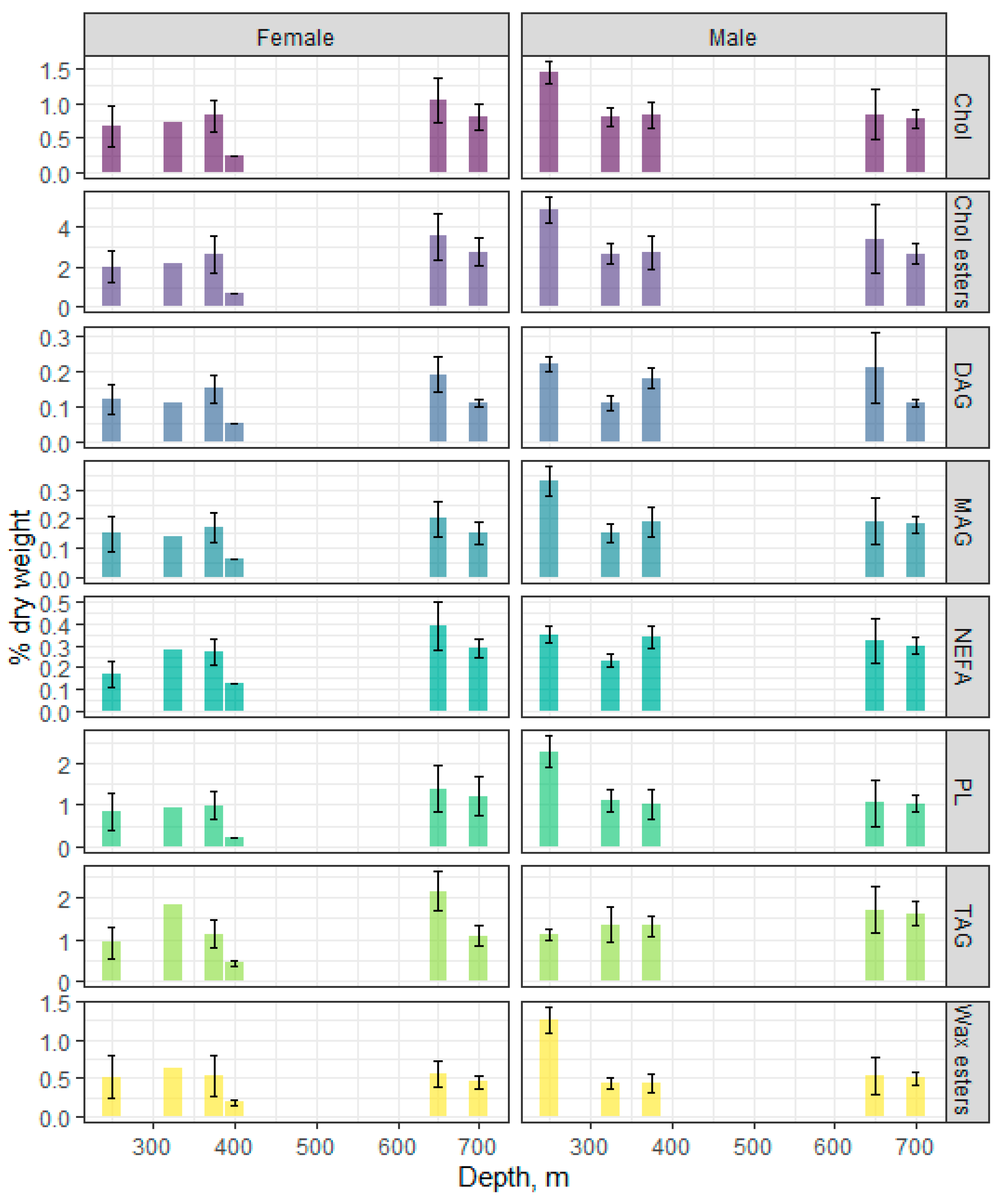
3.2. Species-Specific Characteristics of Neutral Lipids in Beaked Redfish, Their Sex Specificity and Change Depth-Wise
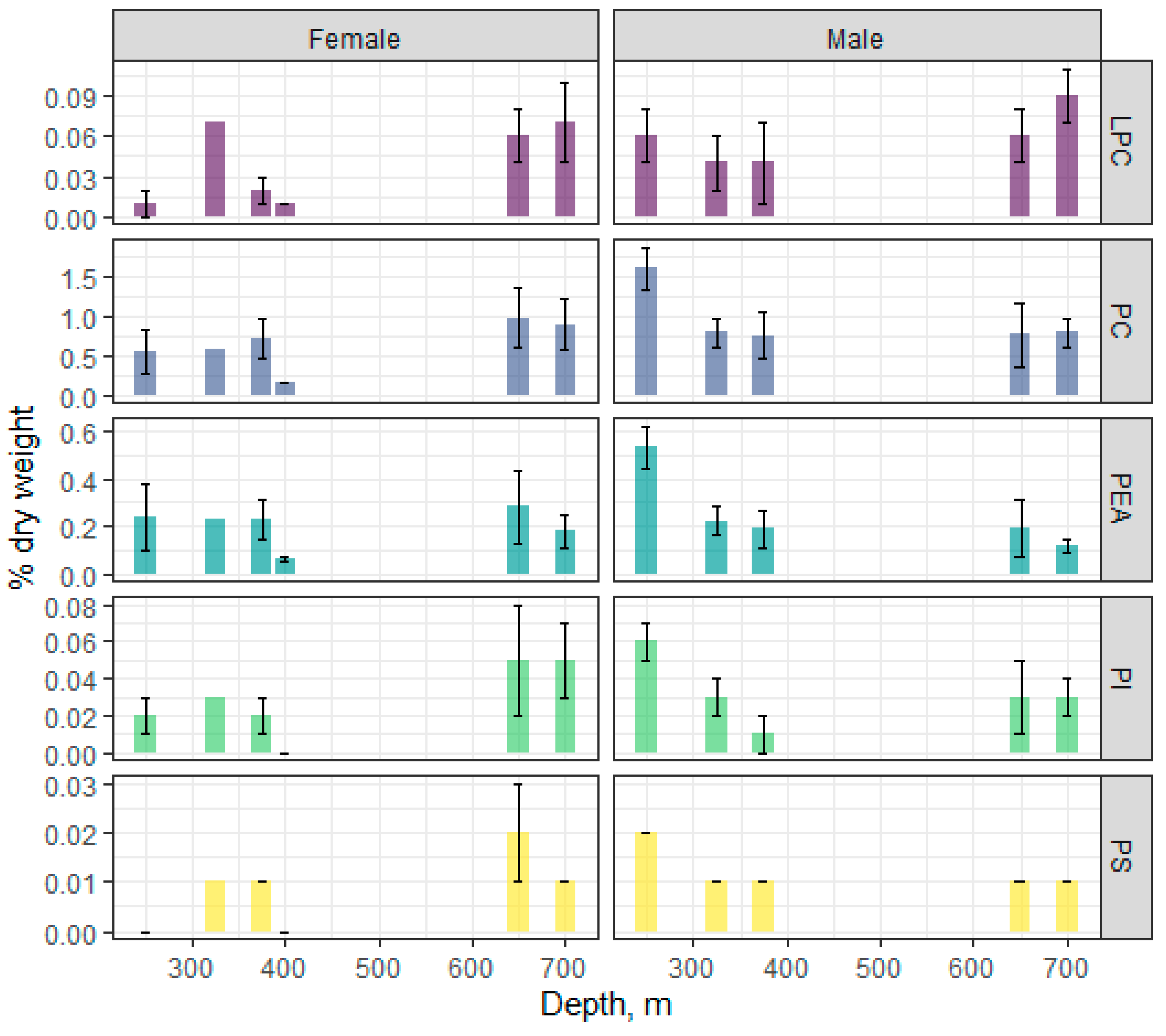
3.3. Species-Specific Characteristics of Polar Lipids in Beaked Redfish, Their Sex Specificity, and Change in the Depth Gradient
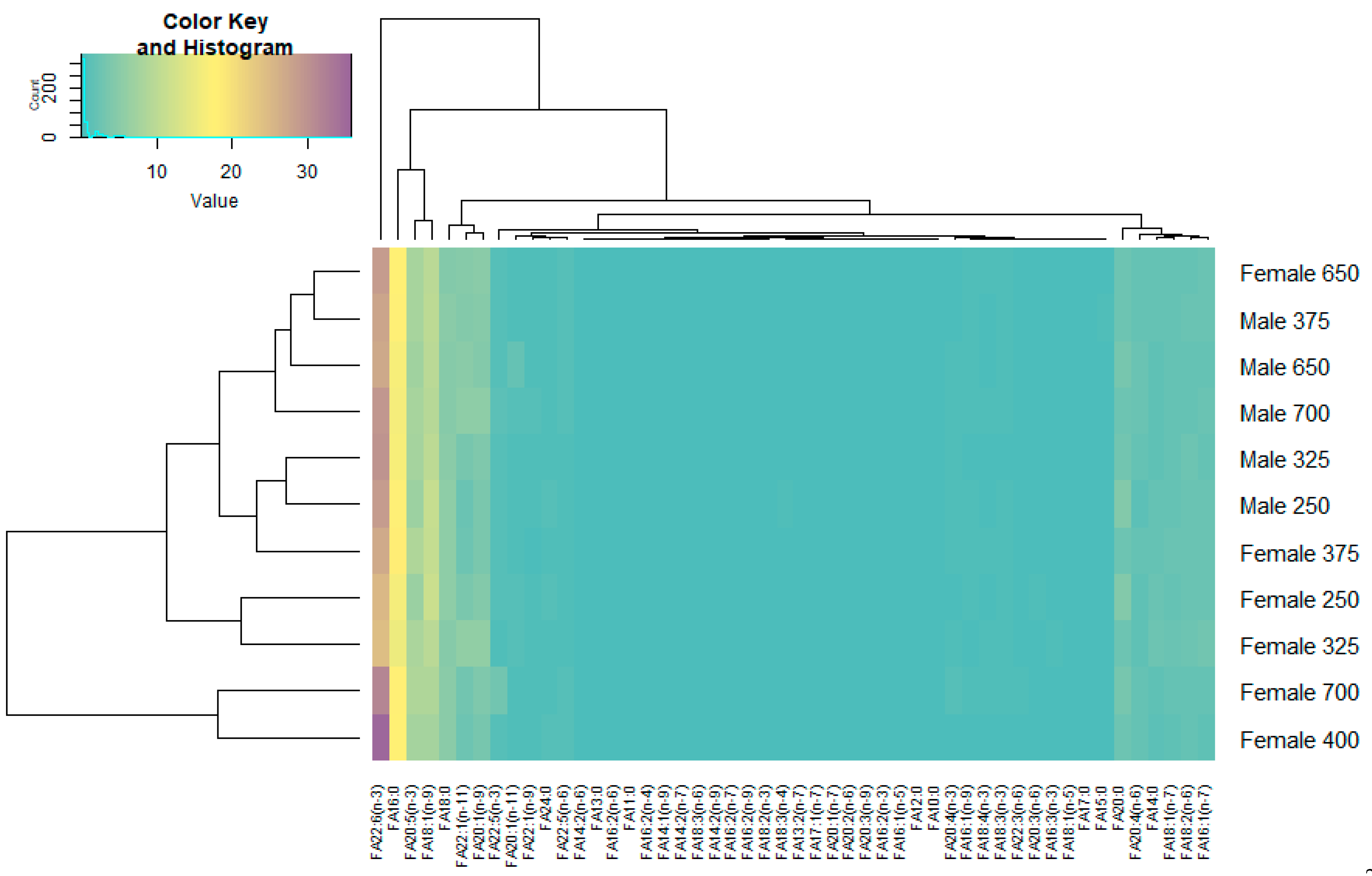
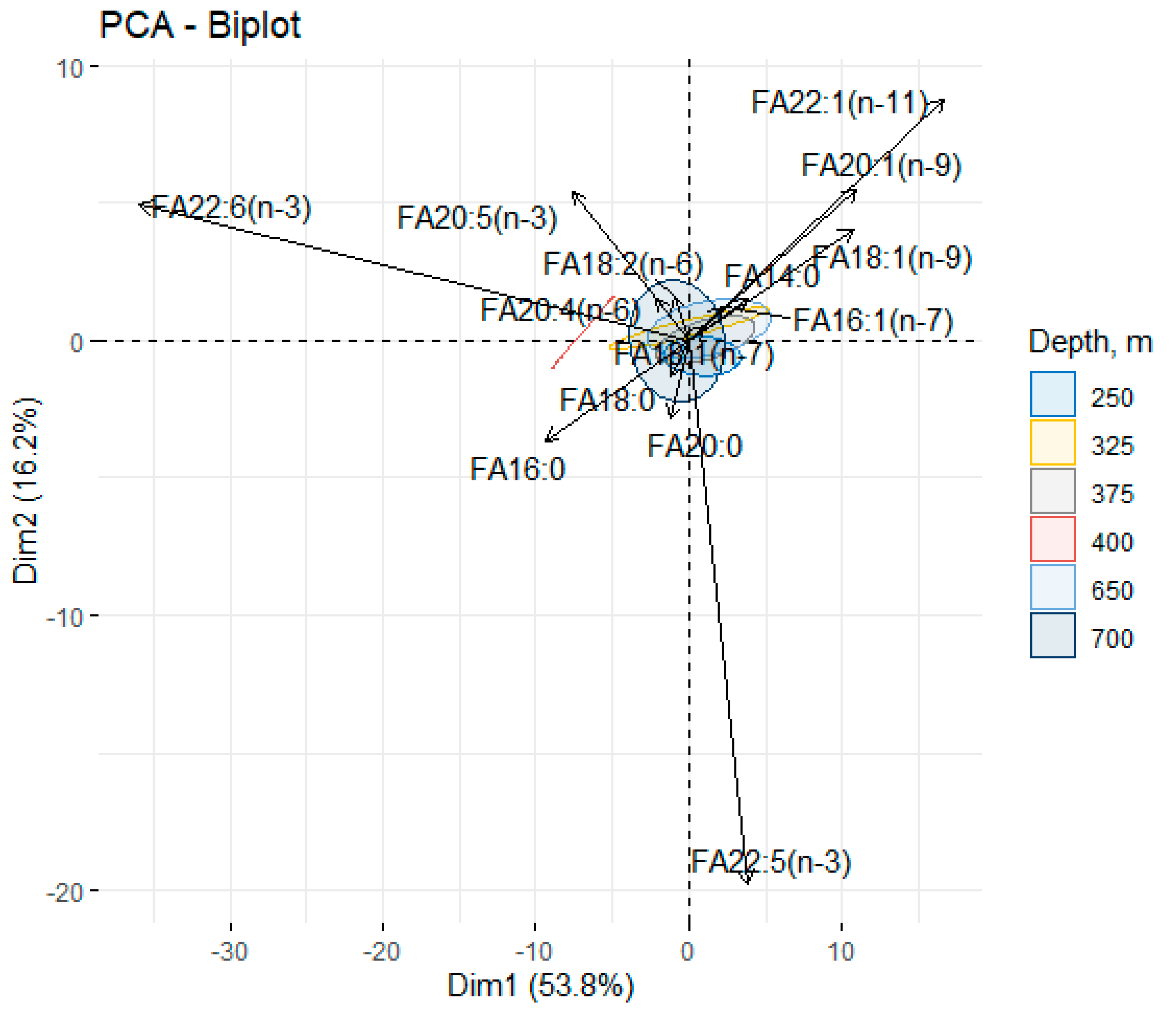
3.4. Species-Specific Characteristics of the Fatty Acid Spectrum in Beaked Redfish, Its Sex Specificity, and Change Depth-Wise
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kreps, E.M. Lipids of Cellular Membranes: Evolution of Brain Lipids: Adaptive Function of Lipids; Nauka: St. Petersburg, Russia, 1981; p. 339. [Google Scholar]
- Sidorov, V.S. Ecological Biochemistry of Fish; Nauka: St. Petersburg, Russia, 1983; p. 240. [Google Scholar]
- Henderson, R.J. Fatty acid metabolism in freshwater fish with particular reference to polyunsaturated fatty acid. Arch. Anim. Nutr. 1996, 49, 5–22. [Google Scholar] [CrossRef] [PubMed]
- Tocher, D.R.; Bell, J.G.; Dick, J.R.; Henderson, R.J.; McGhee, F.; Michell, D.; Morris, P.C. Polyunsaturated fatty acid metabolism in Atlantic salmon (Salmo salar) undergoing parr-smolt transformation and the effects of dietary linseed and rapeseed oils. Fish Physiol. Biochem. 2000, 23, 59–73. [Google Scholar] [CrossRef]
- Hochachka, P.W.; Somero, G.N. Biochemical Adaptation: Mechanism and Process in Physiological Evolution; Oxford University Press: Oxford, UK, 2002; p. 466. [Google Scholar]
- Arts, M.T.; Kohler, C.C. Health and conditions in fish: The influence of lipids on membrane competency and immune response. In Lipids in Aquatic Ecosystems; Arts, M.T., Brett, M.T., Kainz, M.J., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 237–257. [Google Scholar]
- Murzina, S.A.; Nefedova, Z.A.; Nemova, N.N. Effects of fatty acids (biomarkers of food sources) on the mechanisms of fish adaptation in high latitudes. Trans. KarRC RAS 2012, 2, 18–25. [Google Scholar]
- Pavlov, D.S.; Nemova, N.N.; Kirillova, E.A.; Kirillov, P.I.; Nefedova, Z.A.; Murzina, S.A. Lipid Content in the Young of the Year Sockeye Salmon Oncorhynchus nerka during Feeding Migration (the Ozernaya River, Western Kamchatka). Dokl. Biol. Sci. 2012, 445, 235–238. [Google Scholar] [CrossRef] [PubMed]
- Nemova, N.N.; Nefyodova, Z.A.; Murzina, S.A. Lipid patterns early in Atlantic salmon, Salmo salar L., ontogeny. Trans. KarRC RAS 2014, 5, 44–52. [Google Scholar]
- Berge, J.; Renaud, P.E.; Darnis, G.; Cottier, F.; Last, K.S.; Gabrielsen, T.M.; Johnsen, G.; Seuthe, L.; Wes1awski, J.M.; Leu, E.; et al. In the dark: A review of ecosystem processes during the Arctic polar night. Prog. Oceanogr. 2015, 139, 258–271. [Google Scholar] [CrossRef] [Green Version]
- Murzina, S.A.; Pekkoeva, S.N.; Kondakova, E.A.; Nefedova, Z.A.; Filippova, K.A.; Nemova, N.N.; Orlov, A.M.; Berge, J.; Falk-Petersen, S. Tiny but Fatty: Lipids and Fatty Acids in the Daubed Shanny (Leptoclinus maculatus), a Small Fish in Svalbard Waters. Biomolecules 2020, 10, 368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Somero, G.N. Adaptation to high hydrostatic pressure. Annu. Rev. Physiol. 1992, 54, 557–577. [Google Scholar] [CrossRef]
- Gerringer, M.E.; Drazen, J.C.; Yancey, P.H. Metabolic enzyme activities of abyssal and hadal fishes: Pressure effects and a re-evaluation of depth-related changes. Deep-Sea Res. Part 1 2017, 125, 135–146. [Google Scholar] [CrossRef]
- Neighbors, M.A. Triacylglycerols and wax esters in the lipids of deep midwater teleost fishes of the Southern California Bright. Mar. Biol. 1988, 98, 15–22. [Google Scholar] [CrossRef]
- Phleger, C.F.; Nelson, M.M.; Mooney, B.D.; Nichols, P.D. Wax esters versus triacylglycerols in myctophid fishes from the Southern Ocean. Antarct. Sci. 1999, 11, 436–444. [Google Scholar] [CrossRef]
- Tocher, D.R. Metabolism and functions of lipids and fatty acids in teleost fish. Rev. Fish. Sci. 2003, 11, 107–184. [Google Scholar] [CrossRef]
- Shulman, G.E.; Jakovleva, K.K. Hexaenoic Acid and Natural Mobility of Fish. Biol. Bull. Rev. 1983, 44, 529–540. [Google Scholar]
- Zabelinscii, S.A.; Chebotareva, M.A.; Brovcina, N.B.; Krivchenko, A.I. On “adaptive signaling” of the composition of conformational states of fatty acids in membrane lipids of fish gills. J. Evol. Biochem. Physiol. 1995, 31, 29–37. [Google Scholar]
- Lapin, V.I.; Shatunovskii, M.I. Features of the composition, physiological and ecological significance of fish lipids. Biol. Bull. Rev. 1981, 92, 380–394. [Google Scholar]
- Velansky, P.V.; Kostetsky, E.Y. Lipids of marine cold-water fishes. Russ. J. Mar. Biol. 2008, 34, 51–56. [Google Scholar] [CrossRef]
- Joensen, H.; Grahl-Nielsen, O. Discrimination of Sebastes viviparus, Sebastes marinus and Sebastes mentella from Faroe Islands by chemometry of the fatty acid profile in heart and gill tissues and in the skull oil. Comp. Biochem. Physiol. Part B 2000, 126, 69–79. [Google Scholar] [CrossRef]
- Joensen, H.; Grahl-Nielsen, O. The redfish species Sebastes viviparus, Sebastes marinus and Sebastes mentella have different composition of their tissue fatty acids. Comp. Biochem. Physiol. Part B 2001, 129, 73–85. [Google Scholar] [CrossRef]
- Joensen, H.; Grahl-Nielsen, O. Stock structure of Sebastes mentella in the North Atlantic revealed by chemometry of the fatty acid profile in heart tissue. ICES J. Mar. Sci. 2004, 61, 113–126. [Google Scholar] [CrossRef] [Green Version]
- Petursdottir, H.; Gislason, A.; Falk-Petersen, S. Lipid classes and fatty acid composition of muscle, liver and skull oil in deep-sea redfish Sebastes mentella over the Reykjanes Ridge. J. Fish Biol. 2008, 73, 2485–2496. [Google Scholar] [CrossRef]
- Nefedova, Z.A.; Murzina, S.A.; Veselov, A.E.; Ripatti, P.O.; Nemova, N.N. Heterogeneity of Lipids and Fatty Acids of Fingerlings of the Atlantic Salmon Salmo Salar L. Different in Weight and Size. Contemp. Probl. Ecol. 2014, 4, 639–645. [Google Scholar] [CrossRef]
- Nefedova, Z.A.; Murzina, S.A.; Pekkoeva, S.N.; Nemova, N.N. Comparative Characteristic of Fatty Acids Profiles of Smolts of Brown Trout Salmo salar L. and Atlantic Salmon Salmo salar During Smoltification (Indera River, White Sea Basin). Biol. Bull 2018, 2, 144–149. [Google Scholar]
- Murzina, S.A.; Nefedova, Z.A.; Ripatti, P.O.; Nemova, N.N.; Pekkoeva, S.N. Lipid and fatty acid content of the White Sea herring (Clupea pallasi marisalbi Berg) in relation to geographical distribution and environment in the White Sea (Northern Karelia, Russia). Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2012, 163. [Google Scholar] [CrossRef]
- Murzina, S.A.; Nefedova, Z.A.; Pekkoeva, S.N.; Ruokolainen, T.R.; Ripatti, P.O.; Nemova, N.N.; Semushin, A.V. Lipids and fatty acids of the White Sea herring Clupea pallasi marisalbi Berg (Clupeiformes, Clupeidae) from different habitats of the White Sea. Fishes 2016, 1, 65. [Google Scholar] [CrossRef] [Green Version]
- Nemova, N.N.; Murzina, S.A.; Nefedova, Z.A.; Pekkoeva, S.N.; Ropatti, P.O. Lipid status of larvae and adults of the White Sea herring Clupea pallasii marisalbi Berg (Clupeiformes, Clupeidae). Dokl. Biochem. Biophys. 2015, 460, 37–41. [Google Scholar] [CrossRef]
- Johnson, L.; Zhu, J.; Scott, E.R.; Wineinger, N.E. An Examination of the Relationship between Lipid Levels and Associated Genetic Markers across Racial/Ethnic Populations in the Multi-Ethnic Study of Atherosclerosis. PLoS ONE 2015, 10, e0126361. [Google Scholar]
- Zubair, N.; Graff, M.; Luis Ambite, J.; Bush, W.S.; Kichaev, G.; Lu, Y.; Manichaikul, A.; Sheu, W.H.; Absher, D.; Assimes, T.L.; et al. Fine-mapping of lipid regions in global populations discovers ethnic-specific signals and refines previosly identified lipid loci. Hum. Mol. Genet. 2016, 25, 5500–5512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sidore, C.; Busonero, F.; Maschio, A.; Porcu, E.; Naitza, S.; Zoledziewska, M.; Mulas, A.; Pistis, G.; Steri, M.; Danjou, F.; et al. Genome sequencing elucidates Sardinian genetic architecture and augments association analyses for lipid and blood inflammatory markers. Nat. Genet. 2015, 47, 1272–1281. [Google Scholar] [CrossRef] [Green Version]
- Reis-Santos, P.; Tanner, S.E.; Aboim, M.A.; Vasconcelos, R.P.; Laroche, J.; Charrier, G.; Perez, M.; Presa, P.; Gillanders, B.M.; Cabral, H.N. Reconciling differences in natural tags to infer demographic and genetic connectivity in marine fish populations. Sci. Rep. 2018, 8, 10343. [Google Scholar] [CrossRef]
- Xu, S.; Song, N.; Zhao, L.; Cai, S.; Han, Z.; Gao, T. Genomic evidence for local adaptation in the ovoviviparous marine fish Sebasticus marmoratus with a background of population homogeneity. Sci. Rep. 2017, 7, 1562. [Google Scholar] [CrossRef] [Green Version]
- Saborido-Rey, F.; Garabana, D.; Stransky, C.; Melnikov, S.; Shibanov, V. Review of the population structure and ecology of S. mentella in the Irminger sea and adjacent waters. Rev. Fish Biol. Fish. 2004, 14, 455–479. [Google Scholar] [CrossRef]
- Pedchenco, A.P. The role of interannual environmental variations in the geographic range of spawning and feeding concentrations of redfish Sebastes mentella in the Irminger Sea. ICES J. Mar. Sci. 2005, 62, 1501–1510. [Google Scholar] [CrossRef] [Green Version]
- Sigurŏsson, T.; Kristinsson, K.; Ratz, H.J.; Nedreaas, K.H.; Melnikov, S.P.; Reinert, J. The fishery for pelagic redfish (Sebastes mentella) in the Irminger Sea and adjacent waters. ICES J. Mar. Sci. 2006, 63, 725–736. [Google Scholar] [CrossRef]
- Planque, B.; Kristinsson, K.; Astakhov, A.; Bernreuther, M.; Bethke, E.; Drevetnyak, K.; Nedreaas, K.; Reinert, J.; Rolskiy, A.; Sigurdsson, T.; et al. Monitoring beaked redfish (Sebastes mentella) in the North Atlantic, current challenges and future prospects. Aquat. Living Resour. 2013, 26, 293–306. [Google Scholar] [CrossRef] [Green Version]
- Krovnin, A.S.; Melnikov, S.P.; Kivva, K.K.; Artemenkov, D.V.; Moury, G.P. Influence of variability of oceanological conditions on redfish in the North Atlantic pelagial. Trudy VNIRO 2017, 169, 51–63. [Google Scholar]
- Travin, V.I. New species of the sea bass in Barents sea (Sebastes mentella Travin sp. nov.). Dokl. Akad. Nauk. SSSR 1951, 77, 741–744. [Google Scholar] [PubMed]
- Templeman, W. Redfish distribution in the North Atlantic. Bull. Fish. Res. Board Can. 1959, 120, 120–173. [Google Scholar]
- Hureau, J.-C.; Litvinenko, N.I. Scorpaenidae. In Fishes of the North-Eastern Atlantic and the Mediterranean (FNAM); Whitehead, P.J.P., Bauchot, M.-L., Hureau, J.-C., Nielsen, J., Tortonese, E., Eds.; UNESCO: Paris, France, 1986; Volume 3, pp. 1211–1229. [Google Scholar]
- Barsukov, V.V. Annotated and Illustrated Check-List of Rockfishes of the World; Proceedings of Zoological Institute; Zoological Institute Russian Academy of Sciences: St. Petersburg, Russia, 2003; Volume 295, p. 320. [Google Scholar]
- Zakharov, G.P. Redfish Above the Ocean Depths. ICNAF Res. Bull. 1964, 1, 39–42. [Google Scholar]
- Pavlov, A.; Mamylov, V.; Noskov, A. Results of the USSR Investigations of Sebastes Mentella Travin in 1981–1988 (ICES Subareas XII, XIV); ICES: Copenhagen, Denmark, 1989; p. 25. [Google Scholar]
- Magnússon, J.; Magnússon, J.V. Oceanic redfish (Sebastes mentella) in the Irminger Sea and adjacent waters. Scentia Mar. 1995, 59, 241–254. [Google Scholar]
- Barsukov, V.V.; Litvinenko, N.I.; Serebryakov, V.P. Manual for the identification of redfish species of the North Atlantic and adjacent areas. AtlantNIRO. Can. Transl. Fish. Aquat. Sci. 1984, 5168, 25. [Google Scholar]
- Pavlov, A.I. Distribution and Behavior of Redfish (Sebastes mentella Travin) on the Reykjanes Ridge as Observed from the Underwater Vehicle “Sever-2”; “Biological Resources of the Thalassobathial Zone of the World Ocean”; Theses of the Reports of the All-Union Meeting; VNIRO Press: Moscow, Russia, 1988; pp. 54–55. [Google Scholar]
- Melnikov, S.P. North Atlantic Ocean Deep-Sea Redfish: Biology and Fisheries; PINRO Press: Murmansk, Russia, 2006; p. 112. [Google Scholar]
- Bakay, Y.I.; Melnikov, S.P. Biological and ecological characteristics of deepwater redfish Sebastes mentella (Scorpaenidae) at different depths in the pelagial of the Irminger Sea. J. Ichthyol. 2008, 48, 68–80. [Google Scholar] [CrossRef]
- Stefansson, M.O.; Sigurdsson, T.; Pampoulie, C.; Danielsdottir, A.K.; Thorgilsson, B.; Ragnarsdottir, A.; Gislason, D.; Coughlan, J.; Cross, T.F.; Bernatchez, L. Pleistocene genetic legacy suggests incipient species of Sebastes mentella in the Irminger Sea. Heredity 2009, 102, 514–524. [Google Scholar] [CrossRef] [Green Version]
- Filina, E.A.; Rolskiy, A.Y.; Bakay, Y.I.; Popov, V.I.; Makeenko, G.A. Features of the reproductive cycle in females of the beaked redfish Sebastes mentella (Sebastidae). J. Ichthyol. 2017, 57, 129–135. [Google Scholar] [CrossRef]
- Petursdottir, H.; Gislason, A.; Falk-Petersen, S.; Hop, H.; Svavarsson, J. Trophic interaction of the pelagic ecosystem over the Reykjanes Ridge as evaluated by fatty acid and stable isotope analyses. Deep-Sea Res. Part II 2008, 55, 83–93. [Google Scholar] [CrossRef] [Green Version]
- Barsukov, V.V.; Litvinenko, N.I.; Serebryakov, V.P. Methods of Determination of Redfish Species of the Northern Part of the Atlantic Ocean and Adjacent Sees; AtlantNIRO: Kaliningrad, Russia, 1984; p. 28. [Google Scholar]
- Manual for the International Deep Pelagic Ecosystem Survey in the Irminger Sea and Adjacent Waters; ICES Series of ICES Survey Protocol SISP 11—IDEEPS VI; ICES: Copenhagen, Denmark, 2015; p. 49.
- Folch, J.; Lees, M.; Sloan-Syanley, G.H. A simple method for the isolation and purification of total lipids from animal tissue (for brain, liver and muscle). J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Olsen, R.E.; Henderson, R.J. The rapid analysis of neutral and polar marine lipids using double development HPTLC and scanning densitometry. J. Exp. Mar. Biol. Ecol. 1989, 129, 189–197. [Google Scholar] [CrossRef]
- Hellwig, J. Defining Parameters for A Reproducible TLC-Separation of Phospholipids Using ADC 2. Ph.D. Thesis, University of Applied Sciences Northwestern Switzerland (FHNW), Windisch, Switzerland, 2005. [Google Scholar]
- Kabakoff, R. R in Action: Data Analysis and Graphics with R; Volkova, P.A., Translator; DMK Press: Moscow, Russia, 2014; p. 588. [Google Scholar]
- Bruce, A.; Bruce, P. Practical Statistic for Data Scientists; Bruce, P.; Bruce, A., Translators; BHV-Peterburg: St. Peterburg, Russia, 2018; p. 304. [Google Scholar]
- Karl, H.; Numata, J.; Lahrssen-Wiederholt, M. Variability of fat, water and protein content in the flesh of beaked redfish (Sebastes mentella) and Greenland halibut (Reinhardtius hippoglossoides) from arctic fishing grounds. J. Consum. Prot. Food Saf. 2018, 13, 383–389. [Google Scholar] [CrossRef]
- Nemova, N.N. (Ed.) Ecological and Biochemical Status of Juvenile Atlantic Salmon Salmo Salar L. from Some Rivers of the White Sea Basin; RIO KarRC RAS: Petrozavodsk, Russia, 2016; p. 205. [Google Scholar]
- Melnikov, S.P. Intraspecies structure of beaked redfish Sebastes mentella of the Atlantic and Arctic oceans. J. Ichthyol. 2016, 56, 52–71. [Google Scholar] [CrossRef]
- Lee, R.F.; Nevenzel, J.C.; Paffenhöfer, G.-A. Importance of wax esters and other lipids in the marine food chain: Phytoplankton and copepods. Mar Biol. 1971, 9, 99–108. [Google Scholar] [CrossRef]
- Wang, F.; Wu, Y.; Chen, Z.; Zhang, G.; Zhang, J.; Zheng, S.; Katter, G. Trophic interactions of mesopelagic fishes in the South China Sea illustrated by stable isotopes and fatty acids. Front. Mar. Sci. 2019, 5, 522. [Google Scholar] [CrossRef]
- Russ, T.S.; Linberg, G.U. Modern understanding of the natural system of living fish. J. Ichthyol. 1971, 11, 380–407. [Google Scholar]
- Perevozchikov, A.P. Sterols and their transport in animal development. Russ. J. Dev. Biol. 2008, 39, 131–150. [Google Scholar] [CrossRef]
- Ozdemir, N.S.; Parrish, C.C.; Parzanini, C.; Mercier, A. Neutral and polar lipid fatty acids in five families of demersal and pelagic fish from the deep Northwest Atlantic. ICES J. Mar. Sci. 2019, 76, 1807–1815. [Google Scholar] [CrossRef]
- Sargent, J.R. Marine wax esters. Sci. Progr. 1978, 65, 437–458. [Google Scholar] [CrossRef]
- Shchepkina, A.M.; Trusevich, V.V.; Pavlovskaya, T.Y. Peculiarities of lipid composition in some representatives of the mass species of tropical zooplancton from the Atlantic and Indian Ocean. Ecol. Sea 1991, 38, 84–88. [Google Scholar]
- Dalsgaard, J.; St. John, M.; Kattner, G.; Muller-Navarra, D.; Hagen, W. Fatty acid trophic markers in the pelagic marine environment. Adv. Mar. Biol. 2003, 46, 225–340. [Google Scholar] [PubMed]
- Gislason, A. Life-cycle strategies and seasonal migrations of oceanic copepods in the Irminger Sea. Hydrobiologia 2003, 503, 195–203. [Google Scholar] [CrossRef]
- Anderson, T.R.; Martin, A.P.; Lampitt, R.S.; Trueman, C.N.; Henson, S.A.; Mayor, D. Quantifying carbon fluxes from primary production to mesopelagic fish using a simple food web model. ICES J. Mar. Sci. 2019, 76, 690–701. [Google Scholar] [CrossRef]
- Berge, J.; Geoffroy, M.; Daase, M.; Cottier, F.; Priou, P.; Cohen, J.H.; Johnsen, G.; McKee, D.; Kostakis, I.; Renaud, P.E.; et al. Artificial light during the polar night disrupts Arctic fish and zooplankton behaviour down to 200 m depth. Commun. Biol. 2020, 3, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Gennis, R. Biomembranes: Molecular Structure and Function; Mir: Moscow, Russia, 1997; p. 624. [Google Scholar]
- Boldyrev, A.A.; Kyayvaryainen, E.I.; Ilyukha, V.A. Biomembranology: A Textbook; KarRC RAS: Petrozavodsk, Russia, 2006; p. 226. [Google Scholar]
- Makarova, I.I.; Golovko, M.Y. Asymmetry of the source of secondary messengers—Phosphatidylinositol of the cerebral cortex of rats with an increase in geomagnetic activity. In Actual Problems of Functional Interhemispheric Asymmetry: Conference 13–14 December 2001, Moskow; Institute of Brain RAMS: Moscow, Russia, 2001; pp. 103–104. [Google Scholar]
- Colman, J.; Rem, K.-G. Visual Biochemistry, 3rd ed.; Per. with German Language; Mir, BIONOM, Laboratory of Knowledge: Moscow, Russia, 2009; p. 469. [Google Scholar]
- Sandel, E.; Nixon, O.; Lutzky, S.; Ginsbourg, B.; Tandler, A.; Uni, Z.; Koven, W. The effect of dietary phosphatidylcholine/phosphatidylinositol ratio on malformation in larvae and juvenile gilthead sea bream (Sparus aurata). Aquaculture 2010, 304, 42–48. [Google Scholar] [CrossRef]
- Kim, Y.J.; Guzman-Hernandez, M.L.; Balla, T. Inositol lipid regulation of lipid transfer in specialized membrane domains. Trends Cell Biol. 2013, 23, 270–278. [Google Scholar] [CrossRef] [Green Version]
- Aarsman, A.J.; van den Bosch, H. Does de novo synthesis of lysophosphatidylcholine occur in rat lung microsome? Biochim. Biophys. Acta 1980, 620, 410–417. [Google Scholar] [CrossRef]
- Dobrynina, V.I. Biological Chemistry; Medicine: Moskow, Russia, 1976; p. 503. [Google Scholar]
- Osadchaya, L.M.; Galkina, O.V.; Eshchenko, N.D. Effect of Corazole on the Activity of Na+ -K+ ATP—The Basics and the Intensity of Lipid Peroxidation in Neurons and Neuroglia: Biochemical and Molecular-Biological Foundations of Physiological Functions; Publishing House of St. Petersburg State University: St. Petersburg, Russia, 2004; Volume 37, pp. 220–226. [Google Scholar]
- Berdichevets, I.N.; Tyazhelova, T.V.; Shimshilashvili, K.R.; Rogaev, E.I. Lysophosphatidic acid is a lipid mediator with wide range of biological activities. Biosynthetic pathways and mechanism of action. Biochemistry 2010, 75, 1088–1097. [Google Scholar] [CrossRef]
- Tocher, D.R.; Fonseca-Madrigal, J.; Bell, J.G.; Dick, J.R.; Henderson, R.J.; Sargent, J.R. Effect of diets containing linseed oil on fatty acid desaturation and oxidation in hepatocytes and intestinal enterocytes in Atlantic salmon (Salmo salar). Fish Physiol. Biochem. 2002, 26, 157–170. [Google Scholar] [CrossRef]
- Shulman, G.E.; Yuneva, T.V. Role of docosahexaenoic acid in adaptations fishes (review). Hydrobiol. J. 1990, 26, 43–51. [Google Scholar]
- Kanazawa, A. Effects of docosahexaenoic acid and phospholipids on stress tolerance of fish. Aquaculture 1997, 155, 129–134. [Google Scholar] [CrossRef]
- Murzina, S.A. The Role of Lipids and Their Fatty Acid Components in Biochemical Adaptations of the Spotted Lumpen Leptoclinus Maculatus F. Spitsbergen. Ph.D. Thesis, Karelian State Pedagogical Academy, Petrozavodsk, Russia, 2010; p. 184. [Google Scholar]
- Rabinovich, A.L.; Ripatti, P.O. Polyunsaturated carbon chins of lipids: Structure, properties, functions. Biol. Bull. Rev. 1994, 114, 581–594. [Google Scholar]
- Rabinovich, A.L.; Ripatti, P.O.; Balabaev, N.K. Molecular parameters of hydrated bilayers of unsaturated phosphatidylcholines. Russ. J. Phys. Chem. A 2004, 78, 1004–1009. [Google Scholar]
- Rabinovich, A.L. Chain molecules as components of membrane systems: Computer modeling. In Computer Simulation Methods for Researching Polymers and Biopolymers; Ivanov, V.A., Rabinovich, A.L., Hohlov, A.R., Eds.; Limited Liability Company “Book House ”LIBROKOM”: Moscow, Russia, 2009; pp. 409–454. [Google Scholar]
- Ponomarenco, A.I.; Tyrtyshnaia, A.A.; Pislyagin, E.A.; Dyuizen, I.V.; Sultanov, R.M.; Manzhulo, I.V. N-docosahexaenoylethanolamine reduces neuroinflammation and cognitive impairment after mild traumatic brain injury in rats. Sci. Rep. 2021, 11, 756. [Google Scholar] [CrossRef] [PubMed]
- Suhr, S.B.; Pond, D.W.; Gooday, A.J.; Smith, C.R. Selective feeding by benthic foraminifera on phytodetritus on the western Antarctic Peninsula shelf: Evidence from fatty acid biomarker analysis. Mar. Ecol. Prog. Ser. 2003, 262, 153–162. [Google Scholar] [CrossRef]
- Hudson, I.R.; Pond, D.W.; Billett, D.S.M.; Tyler, P.A.; Lampitt, R.S.; Wolff, G.A. Temporal variations in fatty acid composition of deep-sea holothurians: Evidence of bentho-pelagic coupling. Mar. Ecol. Prog. Ser. 2004, 281, 109–120. [Google Scholar] [CrossRef]
- Gershanovich, A.D. Lipid mobilization during early development of turgeons. In Proceedings of the First International Symposium Sturgeon, Bordeaux, France, 3–6 October 1989; pp. 41–52. [Google Scholar]
- Sargent, J.R.; McEvoy, L.A.; Estevez, A.; Bell, G.; Bell, M.; Henderson, J.; Tocher, D. Requirements, presentation and sources of polyunsaturated fatty acids in marine fish larval feeds. Aquaculture 1987, 155, 117–127. [Google Scholar] [CrossRef]
- Falk-Petersen, S.; Sargent, J.R.; Henderson, J.; Hagseth, E.N.; Hop, H.; Okolodkov, Y.B. Lipids and fatty acids in ice algae and phytoplankton from the Marginal Ice Zone in the Barents Sea. Polar. Biol. 1998, 20, 41–47. [Google Scholar] [CrossRef]
- Nelson, M.M.; Mooney, B.; Nichols, J.D.; Phleger, C.F. Lipids of Antarctic Ocean amphipods: Food chain interactions and the occurrence of novel biomarkers. Mar. Chem. 2001, 73, 53–64. [Google Scholar] [CrossRef]
- Lee, R.F. Lipid composition of the copepod Calanus hyperboreas from the Arctic Ocean. Changes with depth and season. Mar. Biol. 1974, 26, 313–318. [Google Scholar] [CrossRef]
- Hagen, W.; Auel, H. Seasonal adaptations and the role of lipids in oceanic zooplankton. Zoology 2001, 104, 313–326. [Google Scholar] [CrossRef]
- Sargent, J.R.; Falk-Petersen, S. The lipid biochemistry of calanoid copepods. Hydrobiologia 1988, 47, 101–114. [Google Scholar] [CrossRef]
- Scott, C.L.; Kwasniewski, S.; Falk-Petersen, S.; Sargent, J.R. Species differences, origins and functions of fatty alcohols and fatty acids in the wax esters and phospholipids of Calanus hyperboreus, C. glacialis and C. finmarchicus from Arctic waters. Mar. Ecol. Prog. Ser. 2002, 235, 127–134. [Google Scholar] [CrossRef]
- Dolgov, V.A.; Rolsky, A. Yu.; Popov, V.I. Feeding of redfish Sebastes mentella in the Irminger Sea—What do the data on feeding show? In Proceedings of the ICES Annual Science Conference, Gdansk, Poland, 19–23 September 2011. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sex | Quantity of Individuals | Body Length Min–Max, sm | Body Weight Min–Max, g | Date of Catch | № Trawl | Latitude, N | Longitude, W | Depth, m |
|---|---|---|---|---|---|---|---|---|
| Females | 1 | 39 | 696 | June 17 | 12 | 64°6′ | 29°37′ | 375 |
| 1 | 40 | 854 | June 20 | 26 | 63°24′ | 37°34′ | 700 | |
| 1 | 38 | 614 | June 21 | 29 | 63°5′ | 34°56′ | 325 | |
| 2 | 37–37 | 932–996 | June 22 | 33 | 63°5′ | 31°54′ | 375 | |
| 2 | 40–40 | 890–940 | June 23 | 36 | 63°5′ | 28°33′ | 700 | |
| 1 | 34 | 456 | June 24 | 38 | 63°35′ | 27°18′ | 700 | |
| 2 | 38–38 | 666–710 | June 25 | 41 | 62°31′ | 30°49′ | 400 | |
| 3 | 33–37 | 478–682 | June 26 | 43 | 62°35′ | 32°28′ | 250 | |
| 1 | 32 | 440 | June 27 | 48 | 62°35′ | 36°2′ | 250 | |
| 2 | 35–36 | 558–594 | June 27 | 49 | 62°35′ | 37°7′ | 375 | |
| 4 | 28–33 | 302–432 | June 28 | 52 | 62°36′ | 38°29′ | 700 | |
| 3 | 33–39 | 502–668 | June 29 | 56 | 62°5′ | 36°35′ | 700 | |
| 1 | 36 | 546 | July 03 | 69 | 61°45′ | 29°20′ | 650 | |
| 3 | 33–37 | 436–626 | July 04 | 74 | 61°45′ | 33°13′ | 650 | |
| 1 | 38 | 696 | July 12 | 94 | 61°26′ | 29°6′ | 650 | |
| All | 28 | 28–40 | 302–996 | June 17–July 12 | 12–94 | 61°26′–64°6′ | 27°18′–38°29′ | 250–700 |
| Males | 2 | 35–40 | 478–806 | June 16 | 11 | 64°6′ | 31°22′ | 700 |
| 1 | 36 | 604 | June 17 | 12 | 64°6′ | 29°37′ | 375 | |
| 1 | 37 | 630 | June 18 | 15 | 63°54′ | 27°57′ | 325 | |
| 2 | 32–35 | 352–476 | June 19 | 22 | 63°35′ | 33°10′ | 700 | |
| 1 | 37 | 376 | June 20 | 23 | 63°35′ | 35°5′ | 325 | |
| 5 | 35–37 | 528–664 | June 21 | 29 | 63°5′ | 34°56′ | 325 | |
| 1 | 42 | 918 | June 24 | 38 | 63°35′ | 27°18′ | 700 | |
| 1 | 36 | 708 | June 26 | 43 | 62°35′ | 32°28′ | 250 | |
| 3 | 39–41 | 686–786 | June 26 | 44 | 62°36′ | 33°24′ | 700 | |
| 9 | 28–34 | 282–550 | June 27 | 48 | 62°35′ | 36°2′ | 250 | |
| 1 | 34 | 574 | June 27 | 49 | 62°35′ | 37°7′ | 375 | |
| 5 | 32–36 | 354–556 | June 28 | 52 | 62°36′ | 38°29′ | 700 | |
| 7 | 35–43 | 508–954 | June 29 | 56 | 62°5′ | 36°35′ | 700 | |
| 1 | 35 | 538 | July 03 | 69 | 61°45′ | 29°20′ | 650 | |
| 4 | 35–40 | 506–810 | July 04 | 74 | 61°45′ | 33°13′ | 650 | |
| All | 44 | 28–43 | 282–954 | June 16–July 04 | 11–74 | 61°45′–64°6′ | 27°18′–38°29′ | 250–700 |
| Total | 72 | 28–43 | 282–996 | June 16–July 12 | 11–94 | 61°45′–64°6′ | 27°18′–38°29′ | 250–700 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Voronin, V.P.; Nemova, N.N.; Ruokolainen, T.R.; Artemenkov, D.V.; Rolskii, A.Y.; Orlov, A.M.; Murzina, S.A. Into the Deep: New Data on the Lipid and Fatty Acid Profile of Redfish Sebastes mentella Inhabiting Different Depths in the Irminger Sea. Biomolecules 2021, 11, 704. https://doi.org/10.3390/biom11050704
Voronin VP, Nemova NN, Ruokolainen TR, Artemenkov DV, Rolskii AY, Orlov AM, Murzina SA. Into the Deep: New Data on the Lipid and Fatty Acid Profile of Redfish Sebastes mentella Inhabiting Different Depths in the Irminger Sea. Biomolecules. 2021; 11(5):704. https://doi.org/10.3390/biom11050704
Chicago/Turabian StyleVoronin, Viktor P., Nina N. Nemova, Tatjana R. Ruokolainen, Dmitrii V. Artemenkov, Aleksei Y. Rolskii, Alexei M. Orlov, and Svetlana A. Murzina. 2021. "Into the Deep: New Data on the Lipid and Fatty Acid Profile of Redfish Sebastes mentella Inhabiting Different Depths in the Irminger Sea" Biomolecules 11, no. 5: 704. https://doi.org/10.3390/biom11050704