
Filmy Ferns (Hymenophyllaceae) and Associated Spike-Mosses (Selaginellaceae) from the Mid-Cretaceous Kachin Amber, Myanmar

1
State Key Laboratory of Palaeobiology and Stratigraphy, Nanjing Institute of Geology and Palaeontology and Center for Excellence in Life and Paleoenvironment, Chinese Academy of Sciences, Nanjing 210008, China
2
Komarov Botanical Institute of the Russian Academy of Sciences, Prof. Popova Str. 2, 197376 Saint Petersburg, Russia
3
University of Chinese Academy of Sciences, Beijing 100049, China
*
Author to whom correspondence should be addressed.
Biology 2022, 11(11), 1629; https://doi.org/10.3390/biology11111629
Submission received: 28 September 2022
/
Revised: 2 November 2022
/
Accepted: 4 November 2022
/
Published: 7 November 2022
(This article belongs to the Special Issue Recent Advances in Palaeobotany)
Abstract
:Simple Summary
Three new species of filmy ferns are described in mid-Cretaceous Kachin amber, which represent the first fossil record of Hymenophyllaceae from tropical Asia. These filmy ferns and the syninclusions of spike-mosses greatly expand the diversity of the cryptogams in Kachin amber and provide additional evidence regarding the paleoenvironment. Together with other contemporaneous findings, the present fossils indicate that Hymenophyllaceae have already accumulated some notable diversity in the Cretaceous.
Abstract
Filmy ferns (Hymenophyllaceae) are the most diverse lineage of the early-diverging leptosporangiate ferns with ca. 430 species widely distributed around the world but with the highest diversity in the humid tropics. However, their fossil record is scarce because of the low preservation potential of the delicate, membranous laminae. So far, no Hymenophyllaceae fossils have been reported from tropical Asia. Here, we describe some fern remains and their syninclusions (spike-mosses) in four pieces of Kachin amber from the mid-Cretaceous of Hukawng Valley, Northern Myanmar, as Hymenophyllites angustus sp. nov., H. kachinensis sp. nov., H. setosus sp. nov. (Hymenophyllaceae) and Selaginella alata sp. nov. (Selaginellaceae), respectively. These fern remains are assigned to Hymenophyllaceae based on the filmy, one-cell thick, decompound pinnatifid laminae and dichotomous venation. They represent the first fossil record of Hymenophyllaceae in tropical Asia. The growth habits of these ferns and associated spike-mosses and their implication for paleoenvironment are discussed. Our study expands the diversity of the cryptogams in mid-Cretaceous Kachin amber. Together with other contemporaneous findings, the present fossils indicate that Hymenophyllaceae have already accumulated some notable diversity in the Cretaceous.
1. Introduction
Molecular phylogenetic analyses reveal that during the Cretaceous Terrestrial Revolution (KTR) period around 125–80 Ma, the radiations of many lineages of land plants, including polypod ferns, epiphytic leafy liverworts and mosses, coincide with the rapid rise of angiosperms [1,2,3,4]. Recent palaeobotanical findings indicate that the mid-Cretaceous Kachin amber deposit of Myanmar is an important source of Mesozoic cryptogams and angiosperms [5,6,7], which could provide direct evidence for the above hypothesis. Indeed, a large number of cryptogamic plant inclusions has already been described from Kachin amber, including liverworts [7,8,9,10,11,12,13,14,15,16,17,18,19], mosses [20,21,22], lycophytes [23,24,25] and ferns [26,27,28,29,30,31,32,33,34,35,36]. As far as ferns are concerned, nine genera and ten species have been reported, most of which could be assigned to the extant families, including Cystodiaceae, Pteridaceae, Dennstaedtiaceae, Marsileaceae, Lindsaeaceae and Thyrsopteridaceae. Here, we recognized Hymenophyllaceae for the first time in the mid-Cretaceous Kachin amber.
The filmy ferns (Hymenophyllaceae) are an early-diverging lineage of leptosporangiate ferns [37,38,39,40], including ca. 430 extant species [39] characterized by their thin filmy lamina usually one-cell thick between veins, pubescent rhizome and marginal indusiate sori [41,42,43,44,45]. Hymenophyllaceae occur predominantly in mossy forest in tropical mountains [45,46]. Traditionally, only two genera have been recognized in this family, viz., Hymenophyllum Sm. with bivalved involucres (indusia) and included receptacles and Trichomanes L. with tubular involucres and long and projecting receptacles [47,48]. Taking into account past studies, as well as recent investigations including molecular data, Ebihara et al. [42] proposed a modern scheme of classification for this family. They divide the Hymenophyllaceae into two subfamilies (clades): the subfamily Hymenophylloideae Burnett (hymenophylloids) with a single genus Hymenophyllum, and the subfamily Trichomanoideae C. Presl (trichomanoids) with eight genera, including Abrodictyum C. Presl, Callistopteris Copel., Cephalomanes C. Presl, Crepidomanes C. Presl, Didymoglossum Desv., Polyphlebium Copel., Trichomanes L. and Vandenboschia Copel. [39,42]. This scheme has been widely accepted by recent researchers.
Numerous Paleozoic and Mesozoic Hymenophyllaceae-like fossils were assigned to the extinct genera Hymenophyllites, Trichomanides and Trichomanites particularly by 19th century researchers, with references compiled in [49,50]. However, most of these fossils lack definite evidence of a membranous habit or a marginal indusiate soral morphology [51,52]. Among them, only two species were thought to be putative members of Hymenophyllaceae [53], namely Hymenophyllites quadridactylites (Gutbier) Kidston from the Late Carboniferous of France [54] and Trichomanides laxum Tenison-Woods from the Jurassic of Queensland, Australia [55]. However, there is no convincing evidence for the presence of a tubular or bivalved indusium in Hymenophyllites quadridactylites [56], although this species has clusters of sporangia at the ends of the pinnule lobes, resembling extant Hymenophyllum. The type of material of Trichomanides laxum was re-examined and determined to be actually a poorly preserved leaf of the corystospermalean seed fern Dicroidium [51].
The oldest unequivocal representative of the Hymenophyllaceae is Hopetedia praetermissa Axsmith et al. from the Late Triassic (Carnian) Pekin Formation of North Carolina, USA [51]. This species is represented by creeping rhizomes, fertile tripinnate membranaceous fronds, marginal funnel-shaped indusia (involucres) and sporangia borne on a short receptacle [51]. The funnel-shaped indusia resemble those of extant genera of the subfamily Trichomanoideae, while the short receptacles are similar to those of extant Hymenophyllum [51]. Other convincing Hymenophyllaceae fossils do not appear until the Early Cretaceous, including Hymenophyllites macrosporangiatus Vachrameev from the Albian of Kazakhstan [57], Eogonocormus cretaceum Deng and E. linearifolium (Deng) Deng from Houlinhe Basin, Inner Mongolia, China [58,59], and Hymenophyllum iwatsukii Herrera et al. from Mongolia [60]. Reinvestigation of the Early Cretaceous Acrostichopteris Fontaine previously assigned to the Schizaeaceae by Reed [61] indicates that it could be reassigned to the Hymenophyllaceae based on characteristics of the pinnules, sori, sporangia and spores [62]. Other Early Cretaceous fossils, such as Hymenophyllites amplus Zheng et Zhang and Hymenophyllites hailarense Ren et Chen from Inner Mongolia and Heilongjiang, China [63,64,65] and Hymenophyllum priscum Menendez from Chile [66], are less convincing [60,67]. Hymenophyllum axsmithii Pigg et al. from the early Eocene Okanogan Highlands, Washington, USA represents an unequivocal member of Hymenophyllaceae in the Cenozoic [67]. All other Cenozoic records of Hymenophyllaceae, such as Hymenophyllum confusum Lesquereux [68], are considered unconfirmed based upon a critical re-appraisal of these reported fossils [69].
Although it is clear that the filmy ferns are ancient lineages according to their phylogenetic position [3,37,38,39], the unequivocal fossil records of them are scarce [45,51,67], mainly because of the low preservation potential for the delicate, membranous lamina and the possibility that some specimens are misidentified to other ferns or even seed ferns [51,53]. In the present paper, we describe some fern remains in the mid-Cretaceous Kachin amber from Myanmar as members of Hymenophyllaceae, namely Hymenophyllites angustus sp. nov., Hymenophyllites kachinensis sp. nov. and Hymenophyllites setosus sp. nov., based on several exquisitely preserved lamina fragments. They represent the first fossil record of the family in tropical Asia. Associated spike-mosses are described as Selaginella alata sp. nov. (Selaginellaceae). The growth habits of these ferns and spike-mosses and their implication for paleoenvironments are discussed.
2. Materials and Methods
Burmese amber originates from several localities at Tanai in the Hukuang Valley, Kachin State (Kachin amber) [70,71], the Hti Lin (Tilin amber, uppermost Campanian ~72.1 Ma) in the Magwe Region [72] and the Pat-tar bum southeast of Khamti (Hkamti Amber, 109.7 ± 0.4 Ma), Sagain Region [73,74]. Kachin amber contains diverse plant and animal fossils [75], while no plant fossils have been reported from Hkamti amber and Tilin amber [72,73]. The age of Kachin amber is widely regarded as the late Albian–early Cenomanian, based on the evidence of the ammonite Puzosia Matsumoto [76] and palynomorphs [77]. U-Pb dating of zircons of Kachin amber suggests an earliest Cenomanian age (98.79 ± 0.62 Ma) for the amber-bearing horizon [78]. Concerning the recent conflicts in Myanmar, we declare that all Kachin amber pieces mentioned in this study were collected before the year 2017 and that we followed the recommendations by Haug et al. [79].
Four pieces of Kachin amber from Myanmar contain fourteen lamina fragments of Hymenophyllaceae. They were all housed at the Collection Department of Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences, with catalog numbers PB200744–200747. The amber inclusions were studied under a ZEISS Axio Zoom.V16 microscope equipped with a high-resolution digital camera (Axiocam 512 color). Incident and transmitted light were used simultaneously for photography. All images were digitally stacked photomicrographic composites of ca. 20–50 individual focal planes using the software package ZEN 2.3 pro for a better illustration of the three-dimensional inclusions. Terminology of the descriptions of the ferns follows Axsmith et al. [51] and Liu et al. [41].
3. Results
Order: Hymenophyllales A.B. Frank
Family: Hymenophyllaceae Mart.
Genus: Hymenophyllites H.R. Goeppert
Species: Hymenophyllites angustus Y. Li et Y.-D. Wang, sp. nov.
Etymology: The specific epithet refers to the narrow ultimate lobes
Holotype: PB200744a (Figure 1D)
Paratypes: PB200744b–h
Type locality: Amber mines southwest of the village of Tanai ca. 105 km north of Myitkyina in Kachin State, Northern Myanmar.
Age: Late Albian–early Cenomanian, mid-Cretaceous.
Repository: Collection Department of Nanjing Institute of Geology and Paleontology, Chinese Academy of Sciences, Nanjing, China.
Specific diagnosis: Lamina fragments tripinnatifid, glabrous. Pinnae closely spaced, up to 0.8 cm long. Pinnules up to 0.5 cm long, distinctly segmented into simple or forked ultimate segments. Venation of pinnules anadromous. Segments characterized by one to several lobes. Lobes sterile, flat, narrow, slightly elongate, ca. 0.1–0.4 mm wide, entire-margined. Veins dichotomous, false veinlets absent. Cell walls straight.
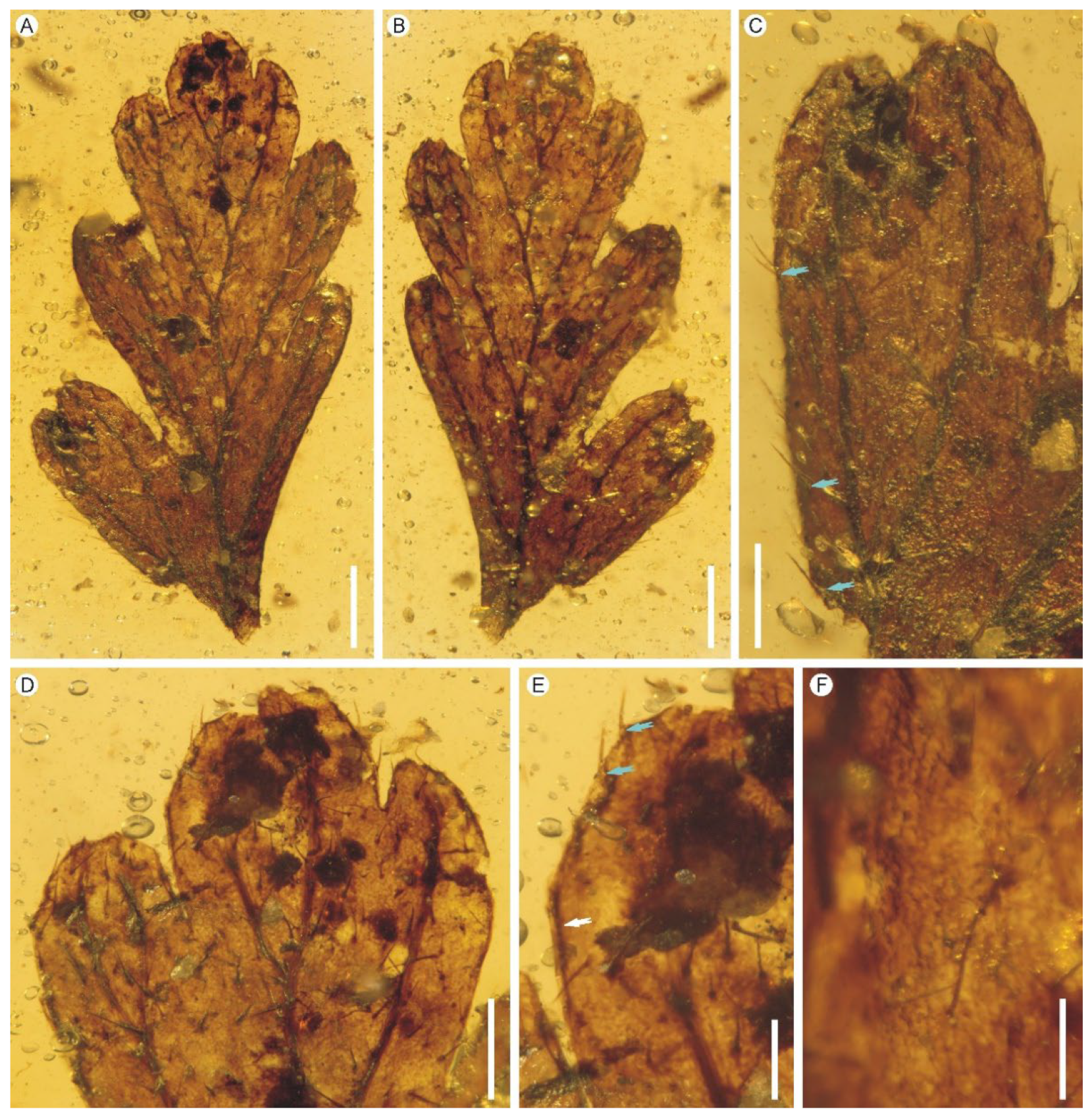
Description: Eight lamina fragments are preserved in one piece of amber (Figure 1A–C). They are, at most, tripinnatifid, up to ca. 1.1–1.2 cm long, membranous, one cell layer thick between the veins, glabrous (Figure 1D–H). The rachis is winged throughout, slightly zigzag or curved, with a flat to crisped wing (Figure 1D). The pinnae are closely spaced, alternate, sessile, up to 0.8 cm long (Figure 1D). The pinnules are closely spaced, up to 0.5 cm long, often with the most proximal ones overlapping the rachis of pinna (Figure 1D). The venation of pinnules is anadromous. The pinnules are distinctly segmented into simple or forked ultimate segments which are further divided into one to several lobes (Figure 1 D–H). The lobes are sterile, flat, narrow, slightly elongated, 0.1–0.4 mm wide, entire-margined, with acute, obtuse, truncate to retuse apices (Figure 1F,G). Each lobe is vascularized by a single veinlet. Veins are dichotomous, forming a zigzag costa, and false veinlets are absent. Lamina cells are polygonal, isodiametrical to slightly elongated, 25–70 μm long and 14–48 μm wide, with straight cell walls; stomata are absent (Figure 1I,J).
Remarks: The present fern remains display the most distinctive characteristic of the family Hymenophyllaceae, viz., membranaceous, translucent laminas which are only one cell layer thick between the veins. Due to the lack of material of fertile fronds with indusia, it is really difficult to nail down the systematic relationships with the subfamilies of the Hymenophyllaceae. However, some characters allow at least the exclusion of most genera of the subfamily Trichomanoideae. For example, the present fern remains differ from Abrodictyum in having more than three rows of cells between midribs and laminar margins; they differ from Cephalomanes in having tripinnatifid lamina; they have glabrous lamina, while Callistopteris and Vandenboschia have hairs on stipes and rachises; the absence of false veinlets in the present fern remains differentiate them from most species of Crepidomanes and Didymoglossum [41,80]. Additionally, the presence of anadromous venation of pinnules also makes our fern remains more consistent with Hymenophylloideae, because most species of extant Hymenophylloideae have anadromous venation of pinnules, whereas, in Trichomanoideae, both anadromous and catadromous venation is present [60]. Due to the lack of fertile materials, we tentatively place our fern remains in the fossil genus Hymenophyllites, which displays great resemblance to the extant Hymenophyllum but still lacks some characters of it. Hymenophyllites represents a putative member of Hymenophyllaceae reported from the Carboniferous to Cretaceous [54,57,63,64,65].
Species: Hymenophyllites kachinensis Y. Li et Y.-D. Wang, sp. nov.
Etymology: The specific epithet refers to amber locality in Kachin State, Myanmar.
Holotype: PB200745d (Figure 2C)
Paratypes: PB200745b,c, PB200746c,d
Type locality: Amber mines southwest of the village of Tanai ca. 105 km north of Myitkyina in Kachin State, northern Myanmar.
Age: Late Albian–early Cenomanian, mid-Cretaceous.
Repository: Collection Department of Nanjing Institute of Geology and Paleontology, Chinese Academy of Sciences, Nanjing, China.
Specific diagnosis: Pinnae or pinnules ca. 1.2–1.3 cm long, once pinnatifid, glabrous, distinctly segmented into simple or forked ultimate segments. Segments characterized by one to several lobes. Lobes sterile, flat, slightly elongate to oblong, 0.7–1.0 mm wide, entire-margined. Veins dichotomous, false veinlets absent. Cell walls straight to slightly wavy.
Description: Five lamina fragments are preserved as sterile pinnae or pinnules in two pieces of amber (Figure 2A,B). The pinnae or pinnules are ca. 1.2–1.3 cm long, once pinnatifid, membranous, one cell layer thick between the veins, glabrous, distinctly segmented into simple or forked ultimate segments which are characterized by one to several lobes (Figure 2C–F). The lobes are flat, slightly elongate to oblong, 0.7–1.0 mm wide, entire-margined with obtuse, truncate to retuse apices. Each lobe is vascularized by a single veinlet. Veins are dichotomous, forming zigzag or curved costules; false veinlets are absent. Lamina cells are polygonal, isodiametrical to elongated, 109–124 μm long and 40–76 μm wide, with straight to slightly wavy cell walls; stomata are absent (Figure 2G). A juvenile sorus is born at apex of ultimate lobe in leaf margin (Figure 2H).
Remarks: The present fern remains are similar to Hymenophyllites angustus sp. nov. in gross morphology, but slightly differs in having larger pinnae and/or pinnules, wider lobes of the ultimate segments and comparatively thinner veins. These characters are often used to distinguish extant species of Hymenophyllum [41] and thus support the present fossils as a new species of Hymenophyllites. The discovery of a juvenile sorus with a likely bivalved indusium born in our leaf margin indicate the present fossils may belong to the extant Hymenophyllum of the subfamily Hymenophylloideae. More fertile materials are still needed to finally assign them into the extant genus. Some sterile and fertile shoots of Selaginella (Figure 2A,B), as well as a fern sporangium (Figure 2D), are preserved as syninclusions with Hymenophyllites kachinensis sp. nov. The sporangium is ovate, sessile, 181 μm long and 146 μm wide, with an uninterrupted annulus (Figure 2I).
Species: Hymenophyllites setosus Y. Li et Y.-D. Wang, sp. nov.
Etymology: The specific epithet refers to the pinna covered with bristle-like hairs.
Holotype: PB200747 (Figure 3)
Type locality: Amber mines southwest of the village of Tanai ca. 105 km north of Myitkyina in Kachin State, northern Myanmar.
Age: Late Albian–early Cenomanian, mid-Cretaceous.
Repository: Collection Department of Nanjing Institute of Geology and Paleontology, Chinese Academy of Sciences, Nanjing, China.
Diagnosis: Pinna or pinnule once pinnatifid, membranous, probably one cell layer thick between the veins, both adaxial and abaxial surfaces sparsely covered with bristle-like hairs. Ultimate segments simple, characterized by single sterile lobes, entire-margined, 0.7–1.3 mm wide, sometimes with darkened submarginal lines. Submarginal false veinlets absent, internal false veinlets absent.
Description: The pinna or pinnule fragment is cuneate at base, once pinnatifid, 7.4 mm long, 3.2 mm wide, membranous, probably one cell layer thick between the veins, with unequally cuneate bases (Figure 3A,B). Both adaxial and abaxial surfaces are sparsely covered with bristle-like hairs (Figure 3C,D). The hairs are single, straight to curved, ca. 68–292 μm long, erect to leaning. The pinna or pinnule is distinctly segmented into 7 ultimate segments. Individual segments are simple, characterized by single sterile lobes, closely spaced, alternate, 0.7–1.3 mm wide; acroscopic and basiscopic parts are shallowly and deeply lobed, respectively; apices are obtuse, rounded to retuse; margin is generally entire but with some hairs located very close to or nearly on leaf margin (Figure 3C,E), and sometimes folded to form darkened submarginal lines (Figure 3E). Veins are dichotomous, forming a zigzag costule and seven lateral veins; lateral veins are simple or dichotomous, reaching high up to approach segment apex, ending just below apex; submarginal false veinlets are absent; internal false veinlets are absent. Lamina cells are polygonal; stomata are absent (Figure 3F).
Remarks: The membranaceous, translucent lamina fragment indicates that the present fern remain probably belongs to the family Hymenophyllaceae. Leaf surface sparsely covered with single setose hairs is characteristic of Hymenophyllum subgenus Sphaerocionium (C. Presl) C. Chr., which is cosmopolitan with ca. 70 species, highly diversified in the Neotropics [42]. In subgenus Sphaerocionium today, the majority of species have stellate hairs but there are also some species with exclusively unbranched single hairs [42]. The darkened submarginal lines in our fossil look like marginal false veinlets of the extant genera Crepidomanes and Didymoglossum in the subfamily Trichomanoideae of Hymenophyllaceae [41,42,80]. But these two extant genera have glabrous leaf surface except D. tahitense (Nadeaud) Ebihara et K. Iwatsuki which has many dark brown hairs along veins [41]. The darkened submarginal lines can also be interpreted as the folds of the differentiated segment margin. Some species of Hymenophyllum subg. Hymenophyllum, e.g., H. rolandi-principis Rosenst., have a marginal seam of smaller, rounded cells that may even be darkened/indurated. Thus, the fern remain studied here closely resembles extant lineages of Hymenophyllum in vegetative characters and could be tentatively assigned to the fossil genus Hymenophyllites due to the lack of fertile characters. The subfamily assignment also awaits future confirmation when fertile material is available. The presence of bristle-like hairs, darkened submarginal lines and slightly thicker leaf lamina make the present species very easy to distinguish from the previous two new species Hymenophyllites angustus and Hymenophyllites kachinensis (Figure 4).
Order: Selaginellales Prantl
Family: Selaginellaceae Willk.
Genus: Selaginella P. Beauv.
Species: Selaginella alata Y. Li et Y.-D. Wang, sp. nov.
Etymology: The specific epithet refers to the alary keel on the abaxial side of the midrib of sporophylls.
Holotype: PB200746b (Figure 5C)
Paratypes: PB200745a, PB200746a
Type locality: Amber mines southwest of the village of Tanai ca. 105 km north of Myitkyina in Kachin State, northern Myanmar.
Age: Late Albian–early Cenomanian, mid-Cretaceous.
Repository: Collection Department of Nanjing Institute of Geology and Paleontology, Chinese Academy of Sciences, Nanjing, China.
Diagnosis: Shoot anisotomously branched. Vegetative leaves dimorphic, arranged in four ranks, two ranks of smaller ascending median leaves and two other ranks of larger spreading lateral leaves. Median leaves non-carinate with aristate apex and denticulate to ciliolate margin. Lateral leaves non-carinate, lanceolate, with acute to acuminate apex and entire to serrulate to ciliolate margin. Strobili born at apex of the fertile shoots, compact, tetragonal. Sporophylls arranged in four ranks, monomorphic, with long acuminate apex, dentate margin, alary serrulate keel.
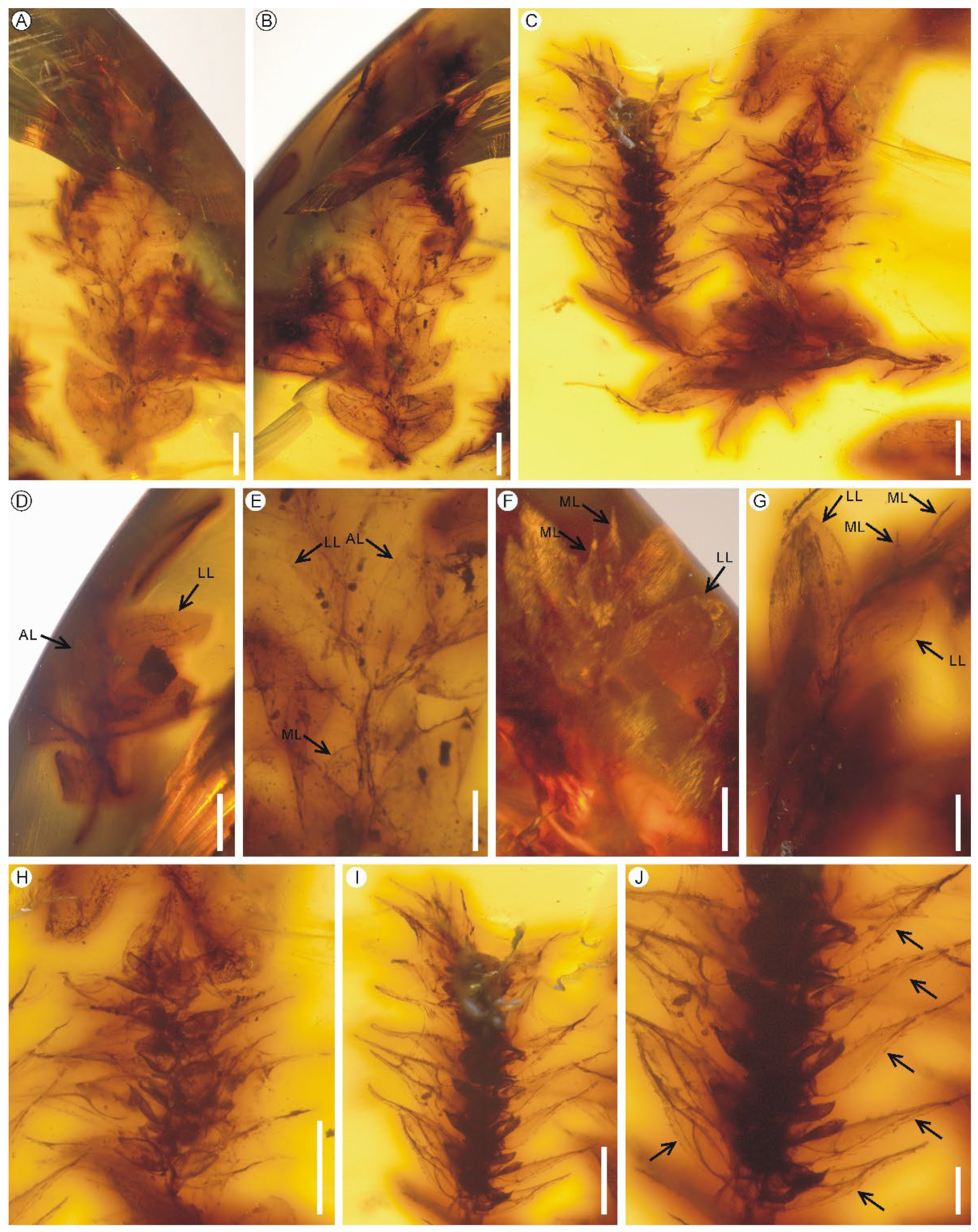
Description: The two fertile shoots are 1.0 cm and 0.5 cm long and bear four and three strobili respectively (Figure 5A–C). Stems are anisotomously branched, without articulations. Rhizophores are not observed. Leaves are dimorphic, arranged in four ranks, two ranks of median leaves on dorsal or upper side of stem and branch and two other ranks of lateral leaves on lateral or lower side (Figure 5D–G). Ligules are not observed. Axillary leaves are symmetrical, lanceolate, 1.6 mm long and 0.6 mm wide (measurement of one leaf), with non-auriculate bases and ciliolate margin (with 52–81 μm long cilia) (Figure 5D,E). Median leaves are smaller, ascending, non-carinate, ovate to obovate, 0.6–0.8 mm long and 0.3–0.4 mm wide (measurement of four leaves), with aristate apex and denticulate to ciliolate margin (with 21–64 μm long teeth or cilia) (Figure 5F,G). Lateral leaves are larger and spreading, non-carinate, lanceolate, 1.2–2.0 mm long and 0.6–0.8 mm wide (measurement of eight leaves), with acute to acuminate apex and entire serrulate to ciliolate margin (with 6–88-μm-long teeth or cilia that occur mainly at the acroscopic base) (Figure 5D–G). Strobili are solitary, born at apex of main stem or branch, compact, tetragonal, 1.7–4.6 mm long and 1.4–1.8 mm wide (Figure 5H,I). Sporophylls are arranged in four ranks, monomorphic, ovate-lanceolate, 0.8–1.1 mm long and 0.1–0.2 mm high, with long acuminate apex, dentate margin (with 21–47 μm long teeth) and alary serrulate keel (with 14–32 μm long teeth) (Figure 5J). Sporophyll-pteryx is not observed. Sporangia are single per sporophyll, mature, and deeply two-valved nearly to the base. Macrospores are not observed.
Remarks: Selaginella is the sole extant genus of Selaginellaceae and includes approximately 750 herbaceous species, widely distributed around the world but with its highest diversity in the tropics [81,82,83,84]. Selaginella is an ancient lineage with fossils dating back to the Carboniferous or even Devonian [53,85]. Schmidt et al. [23,25] discovered the presence of hyperdiverse Selaginella in the mid-Cretaceous Kachin amber, Myanmar, including ten species of Selaginella subgenus Stachygynandrum with anisophyllous (bilateral) strobili and eleven species with isophyllous strobili. All these fossil species are erected based on the combination of a series of key characters [23,25]. Furthermore, Li et al. [24] described a new species of Selaginella subgenus Stachygynandrum with anisophyllous strobili from Kachin amber. Compared with all these Selaginella fossils from Kachin amber, our fossils are clearly different from the eleven species with anisophyllous strobili by having isophyllous strobili, and distinct from the two species S. isophylla A.R. Schmidt et L. Regalado and S. wunderlichiana A.R. Schmidt et L. Regalado with monomorphic and decussately arranged vegetative leaves in having dimorphic vegetative leaves arranged in four rows [25]. Our fossils resemble the rest nine species in the aspects of dimorphic vegetative leaves and isophyllous strobili, but none of the nine species has alary keel on the abaxial side of the midrib of sporophyll [25]. Alary keel is also not documented in the extant species of Selaginella [81,84].
4. Discussion
4.1. Comparison with Hymenophyllaceae Fossils
Hymenophyllites angustus sp. nov. is distinctive for its very narrow ultimate lobes (ca. 0.1–0.4 mm wide), while the lobes are more than 0.5 mm wide in other Hymenophyllaceae fossils (Table 1). Hymenophyllites kachinensis sp. nov. are distinguished from the Triassic Hopetedia praetermissa by the presence of winged costae rather than naked ones [51]. Hymenophyllites kachinensis has pinnatifid lamina and pinnules, whereas most coeval fossils have either fan-like lamina in Eogonocormus cretaceum, E. linearifolium, and Hymenophyllites macrosporangiatus [57,58] or flabellate pinnules in Acrostichopteris alcainensis Sender, A. fimbriata Knowlton, A. interpinnula Meng et Chen and A. longipennis Fontaine [62,86,87,88] (Table 1). Sterile pinnules are strongly segmented and entire-margined in Hymenophyllites kachinensis, while they are weakly segmented with serrate margins in Hymenophyllum iwatsukii [60]. The laminae are at least twice pinnatifid in Hymenophyllites kachinensis, but they are only once pinnate-pinnatifid in the Eocene species Hymenophyllum axsmithii [67]. Hymenophyllites setosus sp. nov. is distinctive for its leaf surface covered with single setose hairs.
Although the subfamily Trichomanoideae are a high diversified lineage with eight genera and an estimated 184 species [39], unequivocal fossils of this subfamily are extremely rare, only with Eogonocormus as a possible member. The thalloid fan-like form of the frond, marginal sori borne on fanlike pinnule lobes, and in situ trilete papillate spores indicate that Eogonocormus is close to the extant genus Gonocormus Bosch [58]. At present, the genus Gonocormus has been merged into Crepidomanes within the subfamily Trichomanoideae, but some members of it were transferred into Hymenophyllum, such as Hymenophyllum nitidulum (Bosch) Ebihara et K. Iwatsuki [41]. These new taxonomic treatments document how misleading morphology alone can be in this group of plants.
4.2. Ecological and Putatively Evolutionary Implications
It is suggested based on ancestral state reconstructions of habit that filmy ferns (Hymenophyllaceae) were ancestrally terrestrial, with epiphytism having evolved several times independently during the Cretaceous [89]. The development of more humid climates in the increasingly closed canopy forests caused by the rapid rise of angiosperms in the Cretaceous may have provided optimal conditions for epiphytic filmy ferns to diversify [89]. It is likely that Hymenophyllites angustus sp. nov., Hymenophyllites kachinensis sp. nov. and Hymenophyllites setosus sp. nov. were epiphytic ferns, because the extant species of Hymenophyllum are almost uniformly epiphytic [89]. Hymenophyllites kachinensis was always found together with the spike-mosses (Selaginella), which are understory herbaceous plants, normally growing on open soil or exposed rocks but also occasionally epiphytic in moist forests [84]. These filmy ferns and associated spike-mosses probably lived on tree trunks and therefore could easily be incased in resin.
Due to the thin filmy laminas, the extant Hymenophyllaceae predominantly live in moist mossy forest [45,90], and the evolution of the filmy ferns shows a tendency towards progressive reduction in size, which may be considered as adaptive to the extremely moist mossy zone [45]. Additionally, the Kachin Selaginella fossils also possess several characters suggestive of humid conditions, such as the relatively thin leaves often with fungal colonization [25]. Therefore, the discovery of the filmy ferns and associated spike-mosses from Kachin amber provides additional evidence for the presence of a tropical humid forest located close to the seashore [70,76,91] and with a diverse fern flora in the mid-Cretaceous of Kachin area. Besides the present fossils, other members of this fern flora also include one aquatic fern Marsileaceaephyllum ciliatum S. Wang et al. (Marsileaceae) [35], two tree ferns Thyrsopteris cretacea C.X. Li et R.C. Moran and T. cyathindusia Shi et al. (Thyrsopteridaceae) [33,34] and eight polypod ferns as follows: Cladarastega burmanica G. Poinar Jr. (Dennstaedtiaceae), Cretacifilix fungiformis G.O. Poinar Jr. et R. Buckley (family incertae sedis), Cystodium sorbifolioides L. Regalado et al. (Cystodiaceae), Heinrichsia cheilanthoides L. Regalado et al. (Pteridaceae), Holttumopteris burmensis L. Regalado et al. (family incertae sedis), Krameropteris resinatus H. Schneid. et al. (Dennstaedtiaceae), Proodontosoria myanmarensis Li et Moran (Lindsaeaceae) and an indeterminate species of Lindsaeaceae [26,27,28,29,30,31,32,36].
Divergence time estimates based on molecular data suggest that the stem group of Hymenophyllaceae may have diverged from other leptosporangiate ferns as early as the Carboniferous to Triassic [3,37,38,89,92,93], with the divergence between Trichomanoideae and Hymenophylloideae occurring during the Middle Jurassic [89]. Based on the analysis of rbcL sequence data, the family Hymenophyllaceae probably arose and first diverged in the Paleotropics, possibly in Asia [90] and subsequently dispersed from there [45]. However, prior to this study, Hymenophyllaceae fossils have never been reported from modern tropical Asia. The rarity of fossils found in tropical regions today is because of the high rate of chemical weathering under present per humid tropical conditions that dissolves any calcium or carbon-based fossils in the rocks. Amber is chemically rather inert and thus the best chance for modern tropical fossils. Hymenophyllites angustus sp. nov., Hymenophyllites kachinensis sp. nov. and Hymenophyllites setosus sp. nov. represent the first fossil record of Hymenophyllaceae from tropical Asia. Although much of the extant diversity of Hymenophyllaceae appears to have accumulated later in the angiosperm-dominated forests of the Cenozoic [89,94], the present fossils, along with other Cretaceous fossil records [57,58,60], indicate that Hymenophyllaceae have already accumulated some notable diversity in the Cretaceous. The study of Dubuisson et al. [93] supported the Early Cretaceous as a critical time for early diversification in Hymenophyllum.
5. Conclusions
Here, we describe fern remains from the mid-Cretaceous Kachin amber, Myanmar as three new species of Hymenophyllites (Hymenophyllaceae), namely H. angustus sp. nov., H. kachinensis sp. nov. and H. setosus sp. nov., and also describe associated Selaginella alata sp. nov. (Selaginellaceae) embedded together with Hymenophyllites kachinensis. The presence of these filmy ferns and associated spike-mosses are suggestive of high humidity in the source forests of this amber. These fern remains of Hymenophyllaceae represent the first fossil record of the family in tropical Asia. Our results greatly expand the known diversity of the fern flora in Kachin amber, Myanmar. Together with other contemporaneous findings, the present fossils indicate that Hymenophyllaceae have already accumulated some notable diversity in the Cretaceous.
Author Contributions
Conceptualization, Y.-D.W.; methodology, Y.L.; software, Y.L.; validation, Y.L.; formal analysis, Y.L.; investigation, Y.L.; resources, Y.L. and N.N.; data curation, Y.L.; writing—original draft preparation, Y.L.; writing—review and editing, Y.L., Y.-D.W., N.N., N.L. and Y.-Y.X.; visualization, Y.L.; supervision, Y.-D.W.; project administration, Y.-D.W. and funding acquisition, Y.-D.W. and N.L. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the National Natural Science Foundation of China, grant number 41790454, the Strategic Priority Research Program of Chinese Academy of Sciences, grant number XDB26000000, and State Key Laboratory of Palaeobiology and Stratigraphy (Nanjing Institute of Geology and Palaeontology, CAS), grant number 213112.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data dealing with this study are reported in the paper.
Acknowledgments
The authors thank the academic editor and assistant editor for handling our manuscript and two anonymous reviewers for their constructive comments and helpful suggestions that greatly improved our manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Fiz-Palacios, O.; Schneider, H.; Heinrichs, J.; Savolainen, V. Diversification of land plants: Insights from a family-level phylogenetic analysis. BMC Evol. Biol. 2011, 11, 341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newton, A.E.; Wikström, N.; Bell, N.; Forrest, L.L.; Ignatov, M.S. Dating the diversification of the pleurocarpous mosses. In Pleurocarpous Mosses: Systematics and Evolution; Newton, A.E., Tangney, R., Eds.; CRC Press: Boca Raton, FL, USA, 2007; pp. 337–366. [Google Scholar]
- Schneider, H.; Schuettpelz, E.; Pryer, K.M.; Cranfill, R.; Magallón, S.; Lupia, R. Ferns diversified in the shadow of angiosperms. Nature 2004, 428, 553–557. [Google Scholar] [CrossRef] [PubMed]
- Feldberg, K.; Schneider, H.; Stadler, T.; Schäferverwimp, A.; Schmidt, A.R.; Heinrichs, J. Epiphytic leafy liverworts diversified in angiosperm-dominated forests. Sci. Rep. 2014, 4, 5974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poinar, G.O. Mid-Cretaceous angiosperm flowers in Myanmar amber. In Recent Advances in Plant Research; Welch, B., Wilkerson, M., Eds.; Nova Science Publishers: New York, NY, USA, 2018; pp. 187–218. [Google Scholar]
- Heinrichs, J.; Feldberg, K.; Bechteler, J.; Regaldo, L.; Renner, M.A.M.; Schäfer-Verwimp, A.; Groehn, C.; Müller, P.; Schneider, H.; Krings, M. Chapter 12—A comprehensive assessment of the fossil record of liverworts in amber. In Transformative Paleobotany: Papers to Commemorate the Life and Legacy of Thomas N. Taylor; Krings, M., Harper, C.J., Cuneo, N.R., Rothwell, G.W., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 213–252. [Google Scholar]
- Feldberg, K.; Schäfer-Verwimp, A.; Renner, M.A.M.; von Konrat, M.; Bechteler, J.; Müller, P.; Wang, Y.-D.; Schneider, H.; Schmidt, A.R. Liverworts from Cretaceous amber. Cretac. Res. 2021, 128, 104987. [Google Scholar] [CrossRef]
- Hentschel, J.; Schmidt, A.R.; Heinrichs, J. Frullania cretacea sp. nov. (Porellales, Jungermanniopsida), a leafy liverwort preserved in Cretaceous amber from Myanmar. Cryptogam. Bryol. 2009, 30, 323–328. [Google Scholar]
- Heinrichs, J.; Reiner-Drehwald, M.E.; Feldberg, K.; von Konrat, M.; Hentschel, J.; Váňa, J.; Grimaldi, D.A.; Nascimbene, P.C.; Schmidt, A.R. The leafy liverwort Frullania (Jungermanniopsida) in the Cretaceous amber forest of Myanmar. Rev. Palaeobot. Palynol. 2012, 169, 21–28. [Google Scholar] [CrossRef]
- Heinrichs, J.; Schäfer-Verwimp, A.; Feldberg, K.; Schmidt, A.R. The extant liverwort Gackstroemia (Lepidolaenaceae, Porellales) in Cretaceous amber from Myanmar. Rev. Palaeobot. Palynol. 2014, 203, 48–52. [Google Scholar] [CrossRef]
- Heinrichs, J.; Schäfer-Verwimp, A.; Hedenäs, L.; Ignatov, M.S.; Schmidt, A.R. An acrocarpous moss in Cretaceous amber from Myanmar. Cretac. Res. 2014, 51, 260–265. [Google Scholar] [CrossRef]
- Bechteler, J.; Schmidt, A.R.; Renner, M.A.M.; Wang, B.; Pérez-Escobar, O.A.; Schäfer-Verwimp, A.; Feldberg, K.; Heinrichs, J. A Burmese amber fossil of Radula (Porellales, Jungermanniopsida) provides insights into the Cretaceous evolution of epiphytic lineages of leafy liverworts. Foss. Rec. 2017, 20, 201–213. [Google Scholar] [CrossRef] [Green Version]
- Heinrichs, J.; Feldberg, K.; Bechteler, J.; Müller, P.; Renner, M.A.M.; Váňa, J.; Schäfer-Verwimp, A.; Schmidt, A.R. A fossil genus of the Frullaniaceae (Porellales, Jungermanniopsida) from the mid-Cretaceous of Myanmar. Cretac. Res. 2017, 74, 223–226. [Google Scholar] [CrossRef]
- Heinrichs, J.; Feldberg, K.; Müller, P.; Schäfer-Verwimp, A.; von Konrat, M.; Ilsemann, B.; Krings, M. Frullania pinnata spec. nov. (Frullaniaceae, Porellales), a new leafy liverwort in mid-Cretaceous Burmese amber from Myanmar. Cretac. Res. 2017, 78, 56–60. [Google Scholar] [CrossRef]
- Li, Y.; Wang, Y.-D.; Schneider, H.; Wu, P.-C. Frullania partita sp. nov. (Frullaniaceae, Porellales), a new leafy liverwort from the mid-Cretaceous of Myanmar. Cretac. Res. 2020, 108, 104341. [Google Scholar] [CrossRef]
- Li, Y.; Li, L.-Q.; Feldberg, K.; Wu, P.-C.; Schmidt, A.R.; Schneider, H.; Wang, Y.-D. Re-appraisal of two fossil Frullaniaceae species (Marchantiophyta, Porellales) from the mid-Cretaceous Burmese amber. Cretac. Res. 2021, 124, 104803. [Google Scholar] [CrossRef]
- Li, Y.; Wang, Y.-D.; Feldberg, K.; Wang, Q.; Yang, X.-J. A new leafy liverwort of Frullania (Frullaniaceae, Porellales) from the mid-Cretaceous Kachin amber, Myanmar. Geol. J. 2021, 56, 5046–5057. [Google Scholar] [CrossRef]
- Wang, Q.; Li, Y.; Feldberg, K.; Wang, Y.-D.; Yang, X.-J. Radula heinrichsii (Radulaceae, Porellales), a leafy liverwort from the mid-Cretaceous of Myanmar. Palaeoworld 2022, 31, 679–687. [Google Scholar] [CrossRef]
- Feldberg, K.; Schäfer-Verwimp, A.; Li, Y.; Renner, M.A.M. Extending the diversity of the bryoflora in Kachin amber (Myanmar), with the description of Radula patrickmuelleri, sp. nov. and R. tanaiensis, sp. nov. (Jungermanniopsida, Porellales, Radulaceae). Foss. Rec. 2022, 25, 213–230. [Google Scholar]
- Bell, N.E.; York, P.V. Vetiplanaxis pyrrhobryoides, a new fossil moss genus and species from Middle Cretaceous Burmese amber. Bryologist 2007, 110, 514–520. [Google Scholar] [CrossRef]
- Hedenäs, L.; Heinrichs, J.; Schmidt, A.R. Bryophytes of the Burmese amber forest: Amending and expanding the circumscription of the Cretaceous moss genus Vetiplanaxis. Rev. Palaeobot. Palynol. 2014, 209, 1–10. [Google Scholar] [CrossRef]
- Li, Y.; Wang, Y.-D.; Feldberg, K.; Wang, S.; Shi, C.; Cui, Y.-M.; Zhang, X.-Q. New insights into the moss genus Vetiplanaxis with a description of V. obtusus sp. nov. from the mid-Cretaceous Kachin amber, Myanmar. Rev. Palaeobot. Palynol. 2022, 301, 104659. [Google Scholar] [CrossRef]
- Schmidt, A.R.; Regalado, L.; Weststrand, S.; Korall, P.; Sadowski, E.-M.; Schneider, H.; Jansen, E.; Bechteler, J.; Krings, M.; Müller, P.; et al. Selaginella was hyperdiverse already in the Cretaceous. New Phytol. 2020, 228, 1176–1182. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Li, Y.-D.; Wang, Y.-D.; Schneider, H.; Shi, G.-L. Re-appraisal of lacewing mimicry of liverworts from the mid-Cretaceous Kachin amber, Myanmar with a description of Selaginella cretacea sp. nov. (Selaginellales, Selaginellaceae). Cretac. Res. 2022, 133, 105143. [Google Scholar] [CrossRef]
- Schmidt, A.R.; Korall, P.; Krings, M.; Weststrand, S.; Bergschneider, L.; Sadowski, E.-M.; Bechteler, J.; Rikkinen, J.; Regalado, L. Selaginella in Cretaceous amber from Myanmar. Willdenowia 2022, 52, 179–245. [Google Scholar] [CrossRef]
- Poinar, G.O., Jr.; Buckley, A.R. Cretacifilix fungiformis gen. and sp. nov., an eupolypod fern (Polypodiales) in Early Cretaceous Burmese amber. J. Bot. Res. Inst. Texas 2008, 2, 1175–1182. [Google Scholar]
- Schneider, H.; Schmidt, A.R.; Heinrichs, J. Burmese amber fossils bridge the gap in the Cretaceous record of polypod ferns. Perspect. Plant Ecol. Evol. Syst. 2016, 18, 70–78. [Google Scholar] [CrossRef] [Green Version]
- Regalado, L.; Schmidt, A.R.; Appelhans, M.S.; Ilsemann, B.; Schneider, H.; Krings, M.; Heinrichs, J. A fossil species of the enigmatic early polypod fern genus Cystodium (Cystodiaceae) in Cretaceous amber from Myanmar. Sci. Rep. 2017, 7, 14615. [Google Scholar] [CrossRef] [Green Version]
- Regalado, L.; Schmidt, A.R.; Müller, P.; Kobbert, M.J.; Schneider, H.; Heinrichs, J. The first fossil of Lindsaeaceae (Polypodiales) from the Cretaceous amber forest of Myanmar. Cretac. Res. 2017, 72, 8–12. [Google Scholar] [CrossRef]
- Regalado, L.; Schmidt, A.R.; Krings, M.; Bechteler, J.; Schneider, H.; Heinrichs, J. Fossil evidence of eupolypod ferns in the mid-Cretaceous of Myanmar. Plant Syst. Evol. 2018, 304, 1–13. [Google Scholar] [CrossRef]
- Regalado, L.; Schmidt, A.R.; Müller, P.; Niedermeier, L.; Schneider, H. Heinrichsia cheilanthoides gen. et sp. nov., a fossil fern in the family Pteridaceae (Polypodiales) from the Cretaceous amber forests of Myanmar. J. Syst. Evol. 2019, 57, 329–338. [Google Scholar] [CrossRef] [Green Version]
- Li, C.-X.; Moran, R.C.; Ma, J.-Y.; Wang, B.; Hao, J.-S. A new fossil record of Lindsaeaceae (Polypodiales) from the mid-Cretaceous amber of Myanmar. Cretac. Res. 2020, 105, 104040. [Google Scholar] [CrossRef]
- Li, C.-X.; Moran, R.C.; Ma, J.-Y.; Wang, B.; Hao, J.-S.; Yang, Q. A mid-Cretaceous tree fern of Thyrsopteridaceae (Cyatheales) preserved in Myanmar amber. Cretac. Res. 2020, 105, 104050. [Google Scholar] [CrossRef]
- Zhang, H.-R.; Shi, C.; Long, X.-X.; Feng, Q.; Cai, H.-H.; Lü, Y.-T.; Wang, S. A new fossil record of Thyrsopteridaceae (Cyatheales) from the mid-Cretaceous amber of Myanmar. Palaeoworld 2021, 31, 478–484. [Google Scholar] [CrossRef]
- Wang, S.; Long, X.-X.; Zhang, H.-R.; Cai, H.-H.; Engel, M.S.; Shi, C. A semi-aquatic fern (Marsileaceae) from the mid-Cretaceous amber of northern Myanmar. Cretac. Res. 2022, 133, 105119. [Google Scholar] [CrossRef]
- Poinar, G., Jr. A new fern, Cladarastega burmanica gen. et sp. nov. (Dennstaedtiaceae: Polypodiales) in mid-Cretaceous Burmese amber. Palaeodiversity 2021, 14, 153–160. [Google Scholar] [CrossRef]
- Rothfels, C.J.; Li, F.-W.; Sigel, E.M.; Huiet, L.; Larsson, A.; Burge, D.O.; Ruhsam, M.; Deyholos, M.; Soltis, D.E.; Stewart, C.N., Jr.; et al. The evolutionary history of ferns inferred from 25 low-copy nuclear genes. Am. J. Bot. 2015, 102, 1089–1107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Testo, W.; Sundue, M. A 4000-species dataset provides new insight into the evolution of ferns. Mol. Phylogenet. Evol. 2016, 105, 200–211. [Google Scholar] [CrossRef] [Green Version]
- The Pteridophyte Phylogeny Group. A community-derived classification for extant lycophytes and ferns. J. Syst. Evol. 2016, 54, 563–603. [Google Scholar] [CrossRef]
- Shen, H.; Jin, D.-M.; Shu, J.-P.; Zhou, X.-L.; Lei, M.; Wei, R.; Shang, H.; Wei, H.-J.; Zhang, R.; Liu, L.; et al. Large-scale phylogenomic analysis resolves a backbone phylogeny in ferns. GigaScience 2018, 7, gix116. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.-X.; Zhang, Q.-Y.; Ebihara, A.; Iwatsuki, K. Hymenophyllaceae. In Flora of China, Vols. 2–3 Pteridophytes; Wu, Z.-Y., Raven, P.H., Hong, D.-Y., Eds.; Science Press: Beijing, China; Missouri Botanical Garden Press: St. Louis, MO, USA, 2013; pp. 93–109. [Google Scholar]
- Ebihara, A.; Dubuisson, J.-Y.; Iwatsuki, K.; Hennequin, S.; Ito, M. A taxonomic revision of Hymenophyllaceae. Blumea 2006, 51, 221–280. [Google Scholar] [CrossRef] [Green Version]
- Ebihara, A.; Iwatsuki, K.; Ito, M.; Hennequin, S.; Dubuisson, J.-Y. A global molecular phylogeny of the fern genus Trichomanes (Hymenophyllaceae) with special reference to stem anatomy. Bot. J. Linn. Soc. 2007, 155, 1–27. [Google Scholar] [CrossRef] [Green Version]
- Farrar, D.R. Hymenophyllaceae. In Flora of North America North of Mexico, Vol. 2 Pteridophytes and Gymnosperms; Flora of North America Editorial Committee, Ed.; Oxford University Press: New York, NY, USA; Oxford, UK, 1993; pp. 190–197. [Google Scholar]
- Iwatsuki, K. Hymenophyllaceae. In The Families and Genera of Vascular Plants, Vol. 1 Pteridophytes and Gymnosperms; Kramer, K.U., Green, P.S., Eds.; Springer-Verlag: Berlin/Heidelberg, Germany, 1990; pp. 157–163. [Google Scholar]
- Vasques, D.T.; Ebihara, A.; Hirai, R.Y.; Prado, J.; Motomi, I. Phylogeny of Hymenophyllum subg. Mecodium (Hymenophyllaceae), with special focus on the diversity of the Hymenophyllum polyanthos species complex. Plant Syst. Evol. 2019, 305, 811–825. [Google Scholar] [CrossRef]
- Christensen, C. Filicinae. In Manual of Pteridology; Verdoorn, F., Ed.; Nijhoff: Hague, The Netherlands, 1938; pp. 522–550. [Google Scholar]
- Bower, F.O. The Ferns (Filicales); Cambridge University Press: London, UK, 1926; Volume II. [Google Scholar]
- Jongmans, W.; Dijkstra, S.J. Pars 43. Filicales, Pteridospermae, Cycadales. In Fossilium Catalogus. II. Plantae; Jongmans, W., Ed.; Uitgeverj Dr W. Junk: Hague, The Netherlands, 1960; pp. 1357–1452. [Google Scholar]
- Jongmans, W.; Dijkstra, S.J. Pars 63. Filicales, Pteridospermae, Cycadales. In Fossilium Catalogus. II. Plantae; Dijkstra, S.J., Ed.; Uitgeverj Dr W. Junk: Hague, The Netherlands, 1965; pp. 3421–3516. [Google Scholar]
- Axsmith, B.J.; Krings, M.; Taylor, T.N. A filmy fern from the Upper Triassic of North Carolina (USA). Am. J. Bot. 2001, 88, 1558–1567. [Google Scholar] [CrossRef] [PubMed]
- Seward, A.C. Fossil Plants, a Text-Book for Students of Botany and Geology; Cambridge University Press: Cambridge, UK, 1910; Volume II, p. 624. [Google Scholar]
- Taylor, T.N.; Taylor, E.L.; Krings, M. Paleobotany: The Biology and Evolution of Fossil Plants, 2nd ed.; Academic Press: Cambridge, MA, USA, 2009. [Google Scholar]
- Zeiller, R. Flore Fossile du Bassin Houiller de Valenciennes; Maison Quantin: Paris, France, 1886. [Google Scholar]
- Tenison-Woods, J.E. On the fossil flora of the coal deposits of Australia. Proc. Linn. Soc. New South Wales 1883, 8, 37–167. [Google Scholar] [CrossRef]
- Kidston, R. Fossil plants from the Carboniferous rocks of Great Britain, part 4, Memoirs of the Geological Survey of Great Britain. Palaeontology 1923, 2, 275–376. [Google Scholar]
- Vakhrameev, V.A. Stratigraphy and fossil flora of the Cretaceous deposits in Western Kazakhstan. In Regional Stratigraphy of the USSR, Part 1; Nalivkin, D.V., Ed.; Akad. Nauk SSSR: Moscow, Russia, 1952; pp. 1–340. (In Russian) [Google Scholar]
- Deng, S.-H. Eogonocormus—A new Early Cretaceous fern of Hymenophyllaceae from China. Aust. Syst. Bot. 1997, 10, 59–67. [Google Scholar] [CrossRef]
- Deng, S.-H. Ecology of the Early Cretaceous ferns of Northeast China. Rev. Palaeobot. Palynol. 2002, 119, 93–112. [Google Scholar] [CrossRef]
- Herrera, F.; Moran, R.C.; Shi, G.-L.; Ichinnorov, N.; Takahashi, M.; Crane, P.R.; Herendeen, P.S. An exquisitely preserved filmy fern (Hymenophyllaceae) from the Early Cretaceous of Mongolia. Am. J. Bot. 2017, 104, 1370–1381. [Google Scholar] [CrossRef] [Green Version]
- Reed, C.F. The phylogeny and ontogeny of the Pteropsida. I. Schizaeales. Bol. Soc. Broteriana 1947, 21, 71–197. [Google Scholar]
- Skog, J.E.; Sender, L.M. New information and family relationship (Hymenophyllaceae) for the fossil fern genus Acrostichopteris Fontaine and a new species from the Lower Cretaceous (Albian) of Spain. Am. J. Bot. 2022, 109, 1443–1455. [Google Scholar] [CrossRef]
- Deng, S.-H.; Ren, S.-Q.; Chen, F. Early Cretaceous Flora of Hailar, Inner Mongolia, China; Geological Publishing House: Beijing, China, 1997. [Google Scholar]
- Yang, X.-J. New material of fossil plants from the Early Cretaceous Muling Formation of the Jixi Basin, Eastern Heilongjiang Province, China. Acta Palaeontol. Sin. 2003, 42, 561–584. [Google Scholar]
- Zheng, S.-L.; Zhang, W. Fossil plants from Longzhaogou and Jixi Groups in Eastern Heilongjiang Province. Bull. Shenyang Inst. Geol. Miner. Resour. Chin. Acad. Geol. Sci. 1982, 5, 277–349, (In Chinese, with an English Abstract). [Google Scholar]
- Menéndez, C.A. Paleophytología Kurtziana. III. 8. La flora del Cretácico Superior de Cerro Guido, Chile (1–2). Ameghiniana 1972, 9, 209–212. [Google Scholar]
- Pigg, K.B.; DeVore, M.L.; Greenwood, D.R.; Sundue, M.A.; Schwartsburd, P.; Basinger, J.F. Fossil Dennstaedtiaceae and Hymenophyllaceae from the Early Eocene of the Pacific Northwest. Int. J. Plant Sci. 2021, 182, 793–807. [Google Scholar] [CrossRef]
- Knowlton, F.H. The Flora of the Denver and Associated Formations of Colorado; U.S. Geological Survey Professional Paper 155; U.S. Government Printing Office: Washington, DC, USA, 1930; pp. 1–142. [Google Scholar]
- Collinson, M.E. Cainozoic ferns and their distribution. Brittonia 2001, 53, 173–235. [Google Scholar] [CrossRef]
- Grimaldi, D.A.; Engel, M.S.; Nascimbene, P.C. Fossiliferous Cretaceous amber from Myanmar (Burma): Its rediscovery, biotic diversity, and paleontological significance. Am. Mus. Novit. 2002, 3361, 1–72. [Google Scholar] [CrossRef]
- Kania, I.; Wang, B.; Szwedo, J. Dicranoptycha Osten Sacken, 1860 (Diptera, Limoniidae) from the earliest Cenomanian Burmese amber. Cretac. Res. 2015, 52, 522–530. [Google Scholar] [CrossRef]
- Zheng, D.-R.; Chang, S.-C.; Perrichot, V.; Dutta, S.; Rudra, A.; Mu, L.; Kelly, R.S.; Li, S.; Zhang, Q.; Zhang, Q.-Q.; et al. A Late Cretaceous amber biota from central Myanmar. Nat. Commun. 2018, 9, 3170. [Google Scholar] [CrossRef]
- Xing, L.-D.; Qiu, L. Zircon U–Pb age constraints on the mid-Cretaceous Hkamti amber biota in northern Myanmar. Paleogeogr. Paleoclimatol. Paleoecol. 2020, 558, 109960. [Google Scholar] [CrossRef]
- Nyunt, T.-T.; Cho, C.; Kyaw, N.B.B.; Krishnaswamy, M.; Ying, L.-H.; Sun, T.-T.; Chutimum Chamaung, N. Geology, occurrence and gemmology of Khamti amber from Sagaing region, Myanmar. Thai Geosci. J. 2021, 2, 61–71. [Google Scholar]
- Ross, A.J. Burmese (Myanmar) amber checklist and bibliography 2018. Palaeoentomology 2019, 2, 22–84. [Google Scholar] [CrossRef]
- Yu, T.-T.; Kelly, R.; Mu, L.; Ross, A.; Kennedy, J.; Broly, P.; Xia, F.-Y.; Zhang, H.-C.; Wang, B.; Dilcher, D. An ammonite trapped in Burmese amber. Proc. Natl. Acad. Sci. USA 2019, 116, 11345–11350. [Google Scholar] [CrossRef] [Green Version]
- Davies, E.H. Palynological Analysis and Age Assignments of Two Burmese Amber Sample Sets; Unpublished Report by Branta Biostratigraphy Ltd. for Leeward Capital Corp.; 2001. [Google Scholar]
- Shi, G.-H.; Grimaldi, D.A.; Harlow, G.E.; Wang, J.; Wang, J.; Yang, M.-C.; Lei, W.-Y.; Li, Q.-L.; Li, X.-H. Age constraint on Burmese amber based on U–Pb dating of zircons. Cretac. Res. 2012, 37, 155–163. [Google Scholar] [CrossRef]
- Haug, J.T.; Azar, D.; Ross, A.; Szwedo, J.; Wang, B.; Arillo, A.; Baranov, V.; Bechteler, J.; Beutel, R.; Blagoderov, V.; et al. Comment on the letter of the Society of Vertebrate Paleontology (SVP) dated April 21, 2020 regarding “Fossils from conflict zones and reproducibility of fossil-based scientific data”: Myanmar amber. Palz 2020, 94, 431–437. [Google Scholar] [CrossRef]
- Wang, H.; Zhao, G.-H.; Ravi, B.X.; Lu, S.-G. Crepidomanes shenzhenense (subg. Crepidomanes; Hymenophyllaceae), a new filmy fern species from Guangdong, southern China. Phytotaxa 2020, 440, 101–107. [Google Scholar]
- Valdespino, I.A. Selaginellaceae. In Flora of North America North of Mexico, vol. 2, Pteridophytes and Gymnosperms; Flora of North America Editorial Committee, Ed.; Oxford University Press: New York, NY, USA, 1993; pp. 38–63. [Google Scholar]
- Weststrand, S.; Korall, P. A subgeneric classification of Selaginella (Selaginellaceae). Am. J. Bot. 2016, 103, 2160–2169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weststrand, S.; Korall, P. Phylogeny of Selaginellaceae: There is value in morphology after all! Am. J. Bot. 2016, 103, 2136–2159. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.-C.; Nooteboom, H.P.; Kato, M. Selaginellaceae. In Flora of China, vol. 2–3, Pteridophytes; Wu, Z.-Y., Raven, P.H., Hong, D.-Y., Eds.; Science Press: Beijing, China; Missouri Botanical Garden Press: St. Louis, MO, USA, 2013; pp. 37–66. [Google Scholar]
- Thomas, B.A. Upper Carboniferous herbaceous lycopsids. Rev. Palaeobot. Palynol. 1997, 95, 129–153. [Google Scholar] [CrossRef]
- Chen, F.; Meng, X.-Y.; Ren, S.-Q.; Wu, C.-L. The Early Cretaceous Flora of Fuxin Basin and Tiefa Basin, Liaoning Province; Geological Publishing House: Beijing, China, 1988. [Google Scholar]
- Knowlton, F.H. Description of a collection of Kootanie plants from the Great Falls coal field of Montana. Smithson. Misc. Collect. 1908, 50, 105–128. [Google Scholar]
- Fontaine, W.M. The Potomac or Younger Mesozoic flora; Monographs of the United States Geological Survey 15; U.S. Government Printing Office: Washington, DC, USA, 1889; pp. 1–377. [Google Scholar]
- Hennequin, S.; Schuettpelz, E.; Pryer, K.M.; Ebihara, A.; Dubuisson, J.-Y. Divergence times and the evolution of epiphytism in filmy ferns (Hymenophyllaceae) revisited. Int. J. Plant Sci. 2008, 169, 1278–1287. [Google Scholar] [CrossRef] [Green Version]
- Dubuisson, J.-Y.; Hennequin, S.; Douzery, E.J.P.; Cranfill, R.B.; Smith, A.R.; Pryer, K.M. rbcL Phylogeny of the Fern Genus Trichomanes (Hymenophyllaceae), with Special Reference to Neotropical Taxa. Int. J. Plant Sci. 2003, 164, 753–761. [Google Scholar] [CrossRef] [Green Version]
- Cruickshank, R.D.; Ko, K. Geology of an amber locality in the Hukawng Valley, Northern Myanmar. J. Asian Earth Sci. 2003, 21, 441–455. [Google Scholar] [CrossRef]
- Pryer, K.M.; Schuettpelz, E.; Wolf, P.G.; Schneider, H.; Smith, A.R.; Cranfill, R. Phylogeny and evolution of ferns (monilophytes) with a focus on the early leptosporangiate divergences. Am. J. Bot. 2004, 91, 1582–1598. [Google Scholar] [CrossRef] [PubMed]
- Dubuisson, J.-Y.; Péchon, T.L.; Hennequin, S.; Rouhan, G.; Salino, A.; Deblauwe, V.; Droissart, V.; Tuomisto, H.; Lehtonen, S.; Ebihara, A. New insights into the diversity, taxonomy and history of the fern genus Trichomanes (Hymenophyllaceae, Polypodiidae), with a focus on Africa and the western Indian Ocean. Bot. J. Linn. Soc. 2022, 198, 215–239. [Google Scholar] [CrossRef]
- Schuettpelz, E.; Pryer, K.M. Evidence for a Cenozoic radiation of ferns in an angiosperm-dominated canopy. Proc. Natl. Acad. Sci. USA 2009, 106, 11200–11205. [Google Scholar] [CrossRef]
Figure 1.
Hymenophyllites angustus sp. nov. from the mid-Cretaceous Kachin amber. (A) Top view of the amber showing seven lamina fragments. PB200744b–h. Scale bar = 5 mm. (B) Bottom view of the amber showing four lamina fragments. PB200744e–h. Scale bar = 5 mm. (C) Lateral side view of the amber showing four lamina fragments. PB200744a–d. Scale bar = 5 mm. (D) A tripinnatifid lamina fragment from middle portion of a lamina showing closely spaced pinnae and anadromous pinnules. Holotype PB200744a. Scale bar = 2 mm. (E) Portion of a pinna showing closely spaced and overlapped pinnules. Holotype PB200744a. Scale bar = 1 mm. (F,G) Pinnule fragments divided into simple or forked ultimate segments. PB200744d, c. Scale bars = 1 mm. (H) A young lamina fragment possibly from apical portion of a lamina. PB200744g. Scale bar = 2 mm. (I) Enlargement of lamina showing cell arrangement. PB200744c. Scale bar = 200 μm. (J) Laminar cells with straight cell walls. PB200744a. Scale bar = 100 μm.
Figure 1.
Hymenophyllites angustus sp. nov. from the mid-Cretaceous Kachin amber. (A) Top view of the amber showing seven lamina fragments. PB200744b–h. Scale bar = 5 mm. (B) Bottom view of the amber showing four lamina fragments. PB200744e–h. Scale bar = 5 mm. (C) Lateral side view of the amber showing four lamina fragments. PB200744a–d. Scale bar = 5 mm. (D) A tripinnatifid lamina fragment from middle portion of a lamina showing closely spaced pinnae and anadromous pinnules. Holotype PB200744a. Scale bar = 2 mm. (E) Portion of a pinna showing closely spaced and overlapped pinnules. Holotype PB200744a. Scale bar = 1 mm. (F,G) Pinnule fragments divided into simple or forked ultimate segments. PB200744d, c. Scale bars = 1 mm. (H) A young lamina fragment possibly from apical portion of a lamina. PB200744g. Scale bar = 2 mm. (I) Enlargement of lamina showing cell arrangement. PB200744c. Scale bar = 200 μm. (J) Laminar cells with straight cell walls. PB200744a. Scale bar = 100 μm.

Figure 2.
Hymenophyllites kachinensis sp. nov. (A–H) and its syninclusions (I) from the mid-Cretaceous Kachin amber. (A) The piece of amber containing three lamina fragments of Hymenophyllites kachinensis sp. nov. (PB200745b–d), as well as a sterile shoot of Selaginella (PB200745a). Scale bar = 2 mm. (B) The piece of amber containing two lamina fragments of Hymenophyllites kachinensis sp. nov. (PB200746c, d) and two fertile shoots of Selaginella (PB200746a, b). Scale bar = 2 mm. (C) The pinna or pinnule. Holotype PB18081914d. Scale bar = 2 mm. (D) A pinna or pinnule fragment and a sporangium. PB200745b,e. Scale bar = 2 mm. (E) Back view of the pinna or pinnule fragment. PB200746d. Scale bar = 2 mm. (F) Bases of two overlapped pinna or pinnule fragments. PB200745c,d. Scale bar = 1 mm. (G) Laminar cells with straight to slightly wavy cell walls. PB200745d. Scale bar = 300 μm. (H) A juvenile sorus born at apex of ultimate segment. PB200745b. Scale bar = 500 μm. (I) An accompanying sporangium. PB200745e. Scale bar = 50 μm.
Figure 2.
Hymenophyllites kachinensis sp. nov. (A–H) and its syninclusions (I) from the mid-Cretaceous Kachin amber. (A) The piece of amber containing three lamina fragments of Hymenophyllites kachinensis sp. nov. (PB200745b–d), as well as a sterile shoot of Selaginella (PB200745a). Scale bar = 2 mm. (B) The piece of amber containing two lamina fragments of Hymenophyllites kachinensis sp. nov. (PB200746c, d) and two fertile shoots of Selaginella (PB200746a, b). Scale bar = 2 mm. (C) The pinna or pinnule. Holotype PB18081914d. Scale bar = 2 mm. (D) A pinna or pinnule fragment and a sporangium. PB200745b,e. Scale bar = 2 mm. (E) Back view of the pinna or pinnule fragment. PB200746d. Scale bar = 2 mm. (F) Bases of two overlapped pinna or pinnule fragments. PB200745c,d. Scale bar = 1 mm. (G) Laminar cells with straight to slightly wavy cell walls. PB200745d. Scale bar = 300 μm. (H) A juvenile sorus born at apex of ultimate segment. PB200745b. Scale bar = 500 μm. (I) An accompanying sporangium. PB200745e. Scale bar = 50 μm.

Figure 3.
Hymenophyllites setosus sp. nov. from the mid-Cretaceous Kachin amber. PB200747. (A,B) Overview of the pinna or pinnule fragment in front and back views. Scale bars = 1 mm. (C) An ultimate segment showing the retuse apex and hairy surface with some hairs located close to the leaf margin (cyan arrows). Scale bar = 500 μm. (D) Enlargement of the pinna or pinnule showing obtuse to retuse apex of the segment. Scale bar = 500 μm. (E) Enlargement of leaf margin showing a darkened submarginal line (white arrow) and two hairs located very close to or nearly on the leaf margin (cyan arrows). Scale bar = 200 μm. (F) Poorly preserved laminar cells. Scale bar = 200 μm.
Figure 3.
Hymenophyllites setosus sp. nov. from the mid-Cretaceous Kachin amber. PB200747. (A,B) Overview of the pinna or pinnule fragment in front and back views. Scale bars = 1 mm. (C) An ultimate segment showing the retuse apex and hairy surface with some hairs located close to the leaf margin (cyan arrows). Scale bar = 500 μm. (D) Enlargement of the pinna or pinnule showing obtuse to retuse apex of the segment. Scale bar = 500 μm. (E) Enlargement of leaf margin showing a darkened submarginal line (white arrow) and two hairs located very close to or nearly on the leaf margin (cyan arrows). Scale bar = 200 μm. (F) Poorly preserved laminar cells. Scale bar = 200 μm.

Figure 4.
Schematic drawings of the pinnule of Hymenophyllites angustus sp. nov. (A), and the pinnae/pinnules of Hymenophyllites kachinensis sp. nov. (B) and Hymenophyllites setosus sp. nov. (C). Scale bar = 1 mm.
Figure 4.
Schematic drawings of the pinnule of Hymenophyllites angustus sp. nov. (A), and the pinnae/pinnules of Hymenophyllites kachinensis sp. nov. (B) and Hymenophyllites setosus sp. nov. (C). Scale bar = 1 mm.

Figure 5.
Accompanying Selaginella alata sp. nov. from the mid-Cretaceous Kachin amber. (A,B) A fertile shoot in dorsal and ventral views. PB200746a. Scale bars = 1 mm. (C) Another fertile shoot. PB200746b. Scale bar = 500 μm. (D) A sterile shoot showing the axillary leaf (AL). PB200745a. Scale bar = 1 mm. (E,F) Magnifications of the shoot showing the axillary, median and lateral vegetative leaves (AL, ML and LL). PB200746a. Scale bar = 500 μm. (G) Magnifications of the shoot showing the median and lateral vegetative leaves. PB200746b. Scale bar = 300 μm. (H,I) Strobili PB200746b. Scale bar = 500 μm. (J) Magnification of the strobilus (I) showing the alary serrulate keel (black arrows) on the abaxial side of the midrib of sporophylls. PB200746b. Scale bars = 200 μm.
Figure 5.
Accompanying Selaginella alata sp. nov. from the mid-Cretaceous Kachin amber. (A,B) A fertile shoot in dorsal and ventral views. PB200746a. Scale bars = 1 mm. (C) Another fertile shoot. PB200746b. Scale bar = 500 μm. (D) A sterile shoot showing the axillary leaf (AL). PB200745a. Scale bar = 1 mm. (E,F) Magnifications of the shoot showing the axillary, median and lateral vegetative leaves (AL, ML and LL). PB200746a. Scale bar = 500 μm. (G) Magnifications of the shoot showing the median and lateral vegetative leaves. PB200746b. Scale bar = 300 μm. (H,I) Strobili PB200746b. Scale bar = 500 μm. (J) Magnification of the strobilus (I) showing the alary serrulate keel (black arrows) on the abaxial side of the midrib of sporophylls. PB200746b. Scale bars = 200 μm.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Comparison of the present fossils with related Hymenophyllaceae fossils.
| Taxon | Lamina Shape | Lamina Surface | Pinnule Shape | Lobe Width (mm) | False Veinlets | Age | Locality | Reference |
|---|---|---|---|---|---|---|---|---|
| Acrostichopteris alcainensis Sender | Pinnate to pinnate-pinnatifid | ? | Flabellate | ? | ? | Early Cretaceous | Alcaine, Teruel Province, Spain | [62] |
| Acrostichopteris fimbriata Knowlton | Pinnate | ? | Flabellate | ? | ? | Early Cretaceous | Montana, USA | [87] |
| Acrostichopteris interpinnula Meng et Chen | Tripinnate | ? | Flabellate | 0.5 | ? | Early Cretaceous | Fuxin Basin, Liaoning, China | [86] |
| Acrostichopteris longipennis Fontaine | Pinnate-pinnatifid | ? | Flabellate | ? | ? | Early Cretaceous | Potomac Group, Baltimore, Maryland and Virginia, USA | [62,88] |
| Eogonocormus cretaceum Deng | Fan-like | ? | – | 1.0 | ? | Early Cretaceous | Huolinhe Basin, Inner Mongolia, China | [58] |
| Eogonocormus linearifolius Deng | Fan-like | ? | – | 1.0 | ? | Early Cretaceous | Huolinhe Basin, Inner Mongolia, China | [58] |
| Hopetedia praetermissa Axsmith et al. | Tripinnate | ? | Pinnate to deeply pinnatifid | 1.0 | ? | Late Triassic | North Carolina, USA | [51] |
| Hymenophyllites angustus sp. nov. | Tripinnatifid | Glabrous | Deeply pinnatifid | 0.1–0.4 | Absent | Mid-Cretaceous | Kachin amber, Myanmar | This paper |
| Hymenophyllites kachinensis sp. nov. | Pinnatifid | Glabrous | Deeply pinnatifid | 0.7–1.0 | Absent | Mid-Cretaceous | Kachin amber, Myanmar | This paper |
| Hymenophyllites macrosporangiatus Vachrameev | Fan-like | ? | – | ? | ? | Early Cretaceous | Kazakhstan | [57] |
| Hymenophyllites setosus sp. nov. | Pinnatifid | With bristle-like hairs | Pinnatifid | 0.7–1.3 | Absent | Mid-Cretaceous | Kachin amber, Myanmar | This paper |
| Hymenophyllum axsmithii Pigg et al. | Once pinnate-pinnatifid | ? | – | 0.5 | ? | Early Eocene | Boot Hill, Washington, USA | [67] |
| Hymenophyllum iwatsukii Herrera et al. | Bipinnate | Probably with trichomes | Shallowly pinnatifid | ? | ? | Early Cretaceous | Tevshiin Govi and Tugrug, Mongolia | [60] |
? means unknown character, and – stands for inapplicable character.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Li, Y.; Wang, Y.-D.; Nosova, N.; Lu, N.; Xu, Y.-Y. Filmy Ferns (Hymenophyllaceae) and Associated Spike-Mosses (Selaginellaceae) from the Mid-Cretaceous Kachin Amber, Myanmar. Biology 2022, 11, 1629. https://doi.org/10.3390/biology11111629
AMA Style
Li Y, Wang Y-D, Nosova N, Lu N, Xu Y-Y. Filmy Ferns (Hymenophyllaceae) and Associated Spike-Mosses (Selaginellaceae) from the Mid-Cretaceous Kachin Amber, Myanmar. Biology. 2022; 11(11):1629. https://doi.org/10.3390/biology11111629
Chicago/Turabian StyleLi, Ya, Yong-Dong Wang, Natalya Nosova, Ning Lu, and Yuan-Yuan Xu. 2022. "Filmy Ferns (Hymenophyllaceae) and Associated Spike-Mosses (Selaginellaceae) from the Mid-Cretaceous Kachin Amber, Myanmar" Biology 11, no. 11: 1629. https://doi.org/10.3390/biology11111629
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.