
Morphological and Micromorphological Description of the Larvae of Two Endemic Species of Duvalius (Coleoptera, Carabidae, Trechini)



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
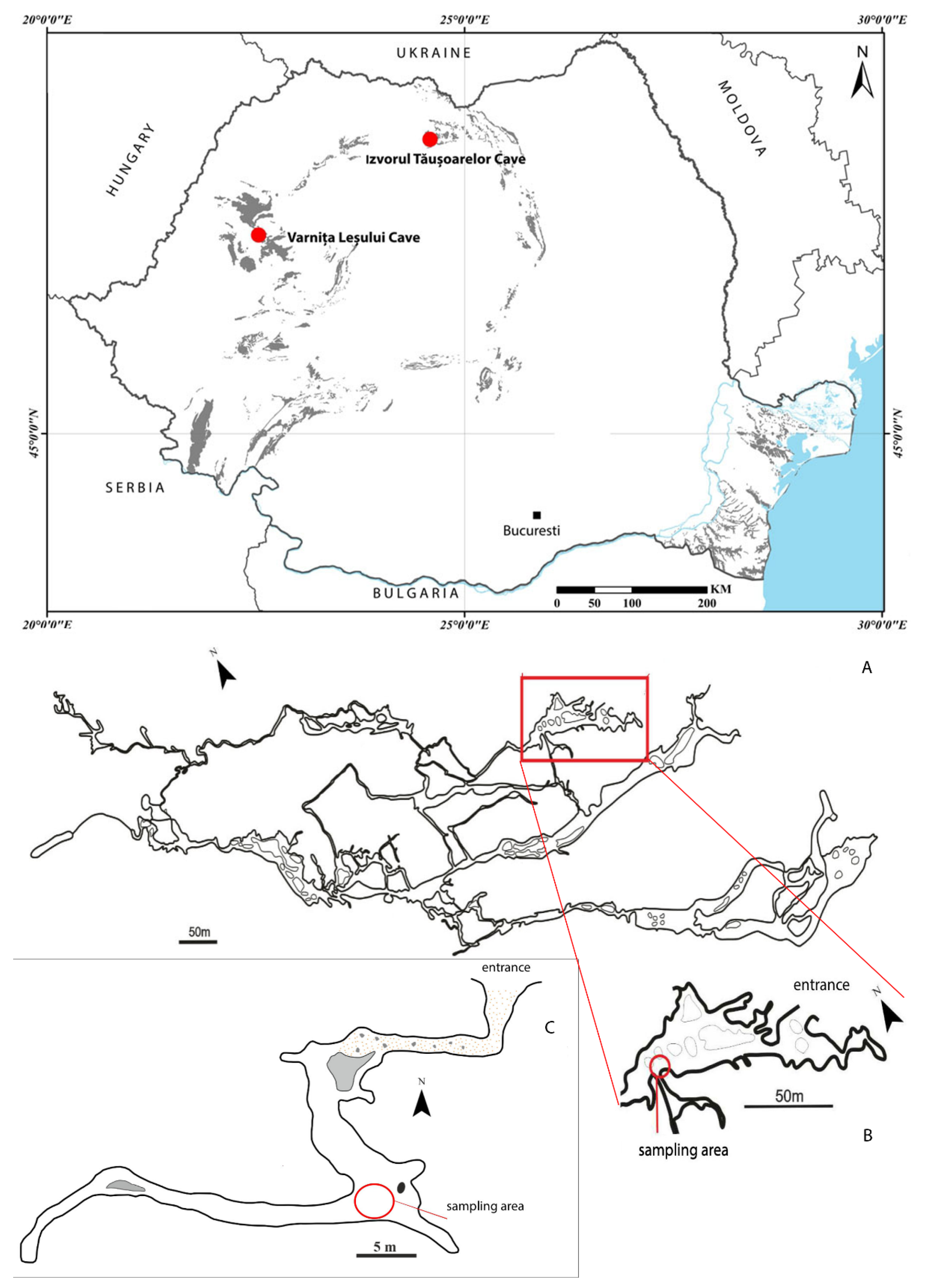
2.1. Sampling Sites
2.2. Microscopy Analysis
3. Results
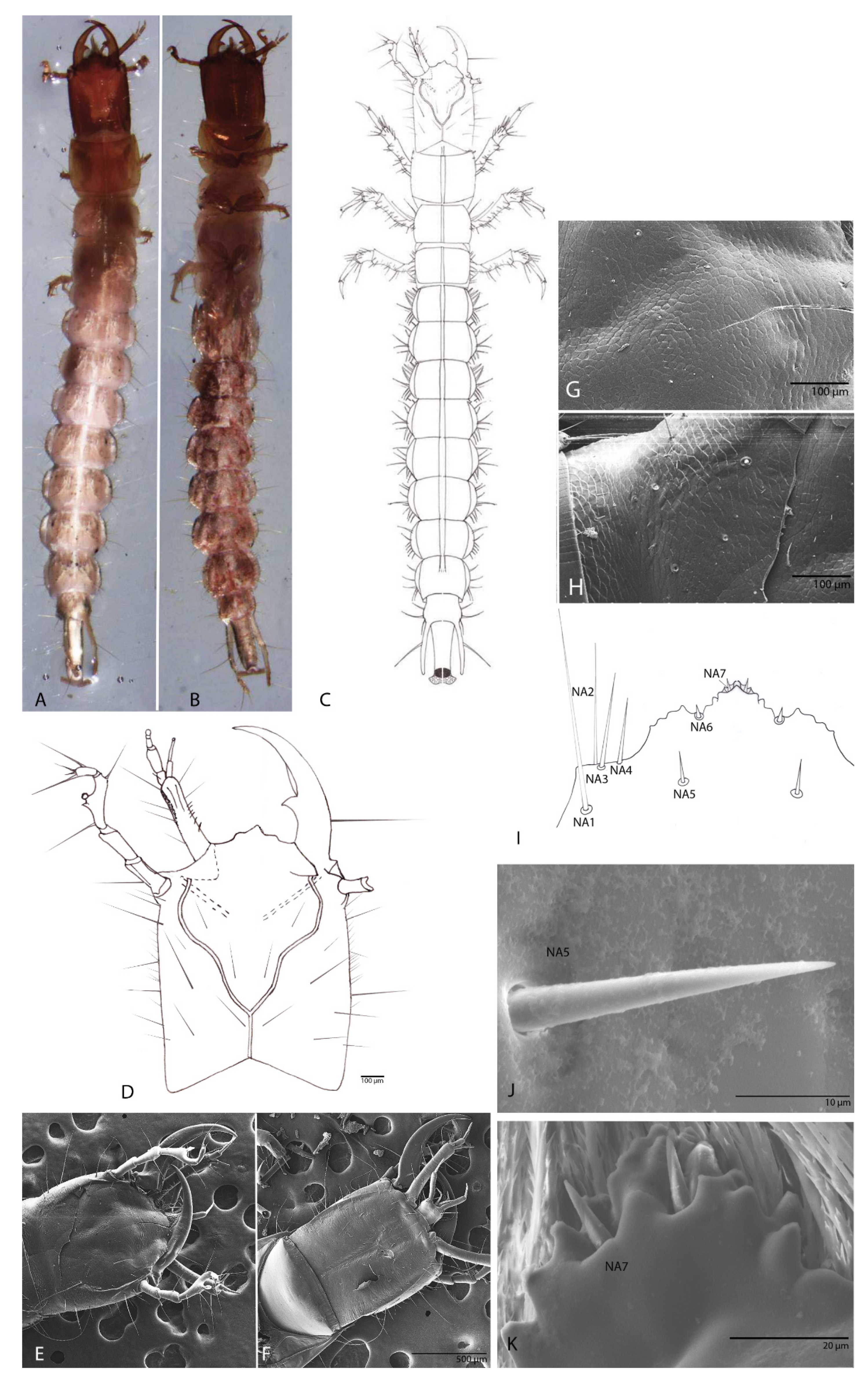
3.1. Description of Duvalius (Hungarotrechus) subterraneus (L. Miller, 1868) Larva
3.1.1. Habitus
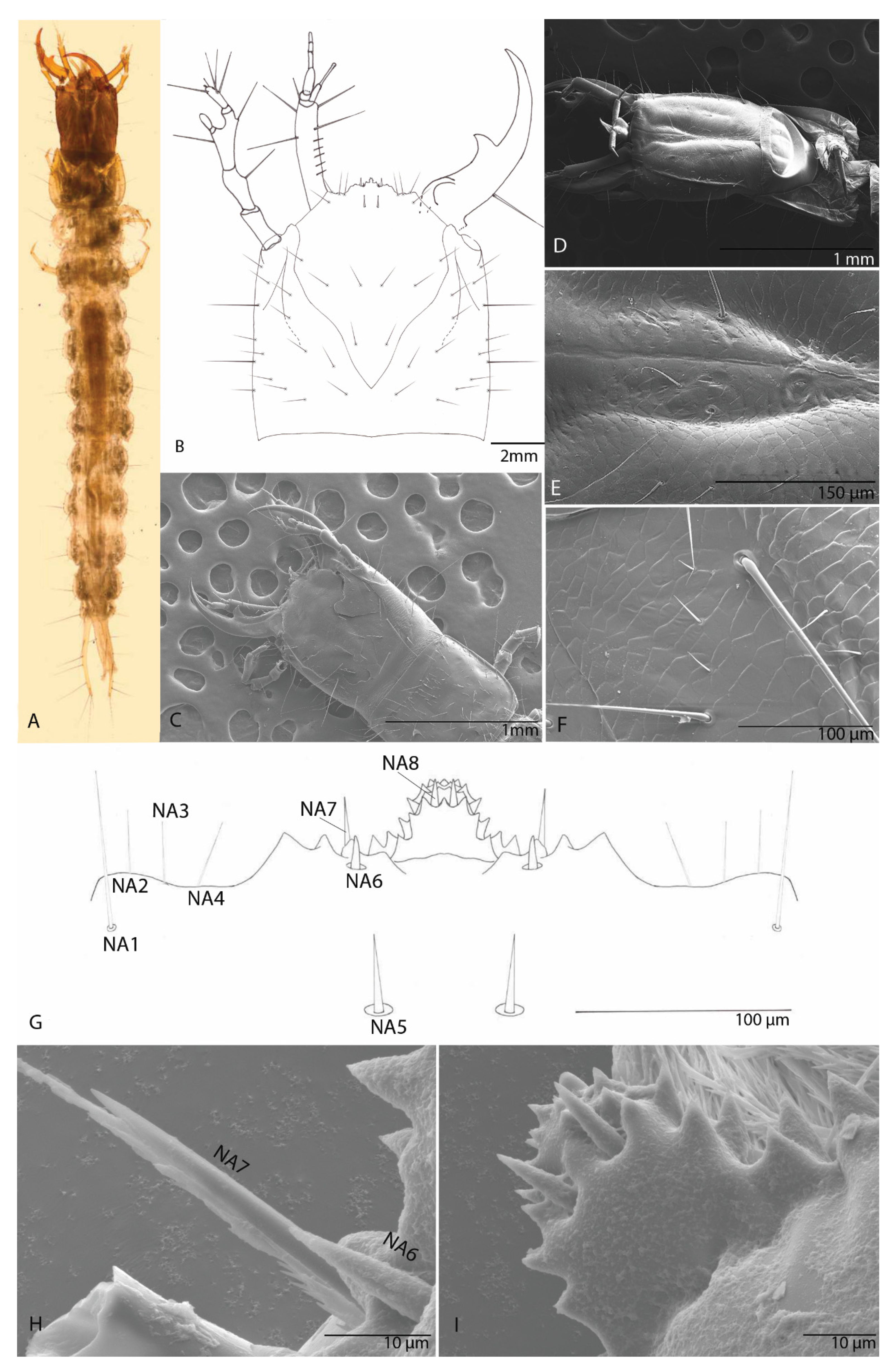
3.1.2. Head
3.1.3. The Nasale (Anterior Margin of Epistome)
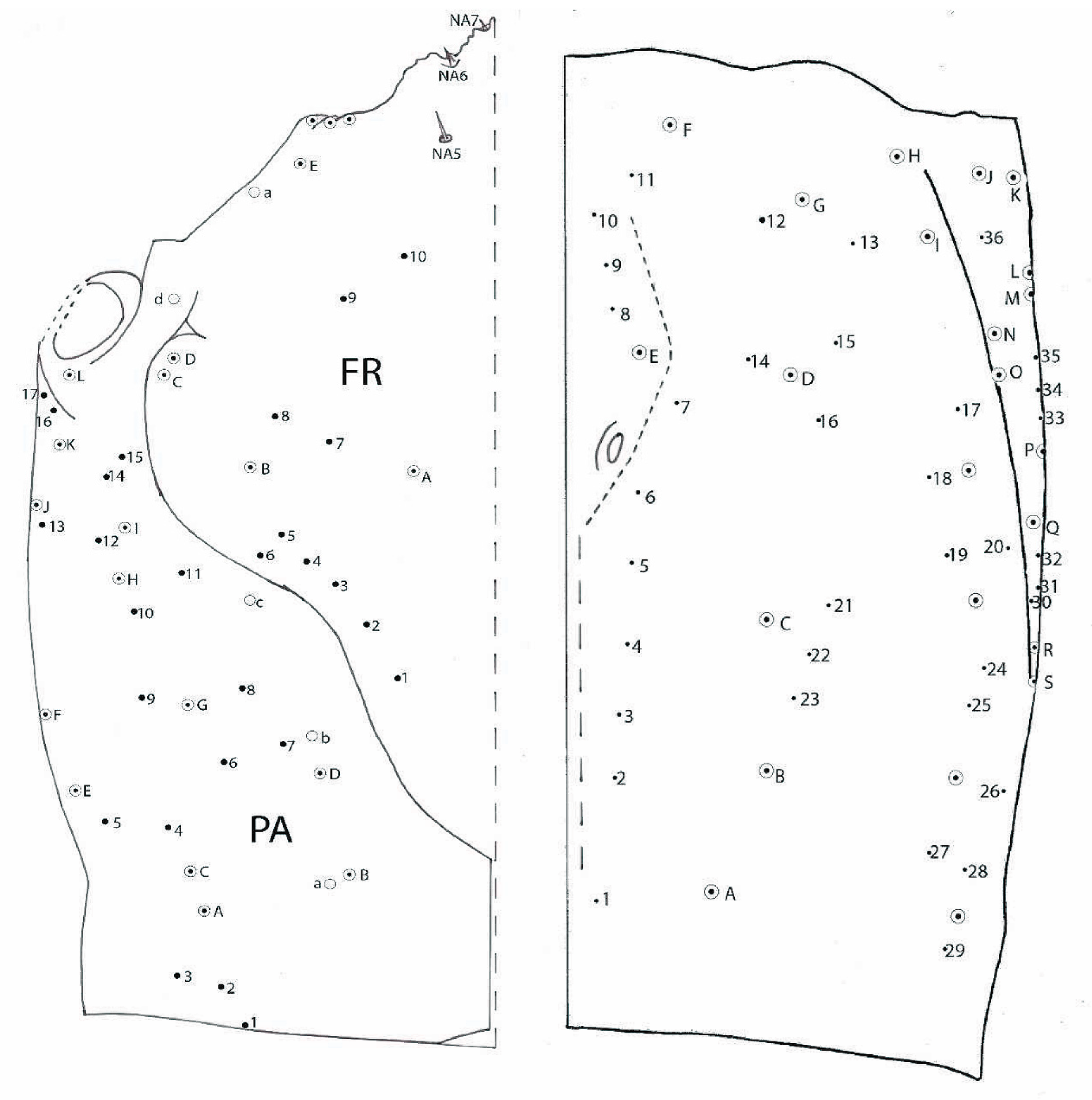
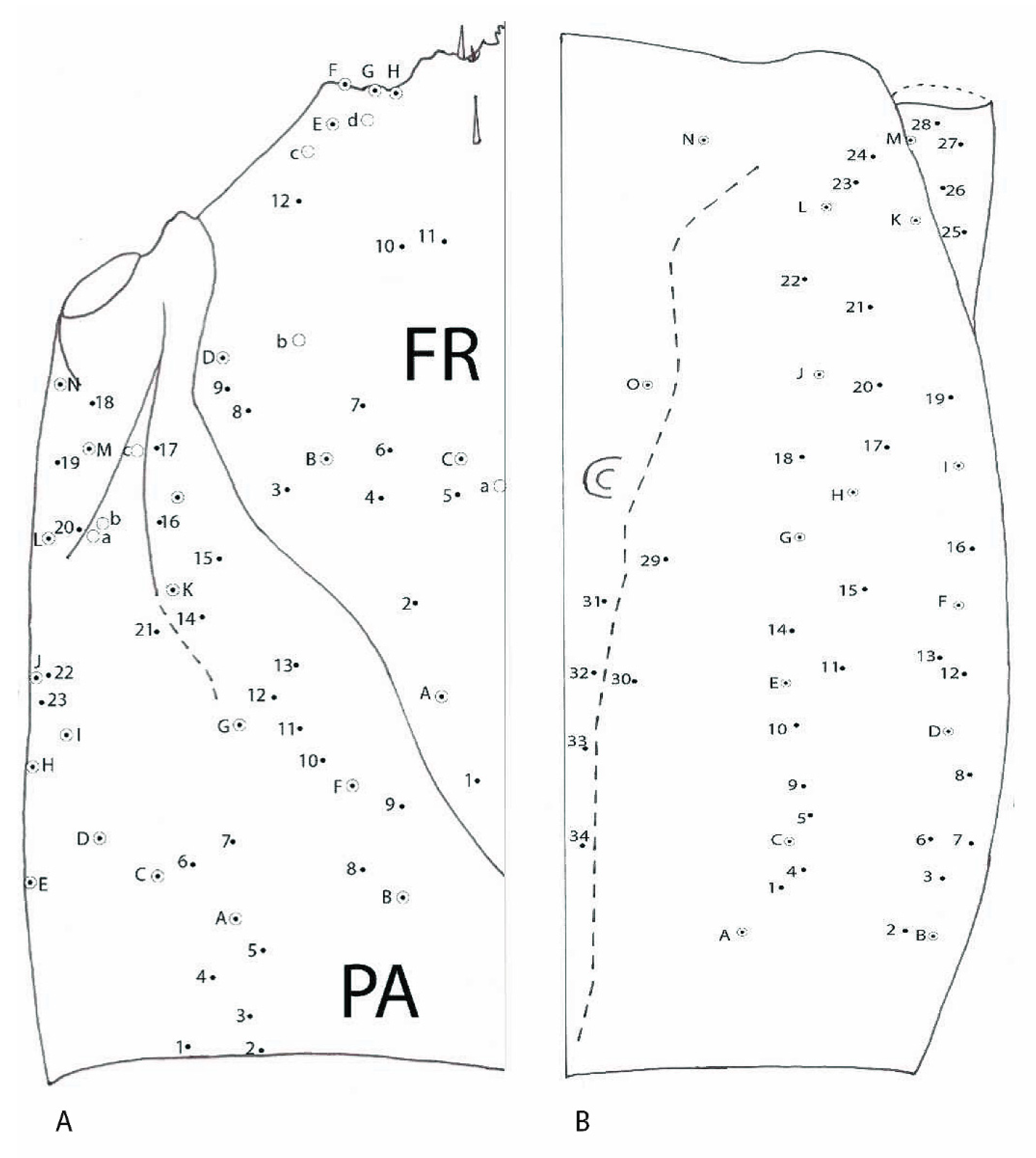
3.1.4. Chaetotaxy of the Cephalic Capsule
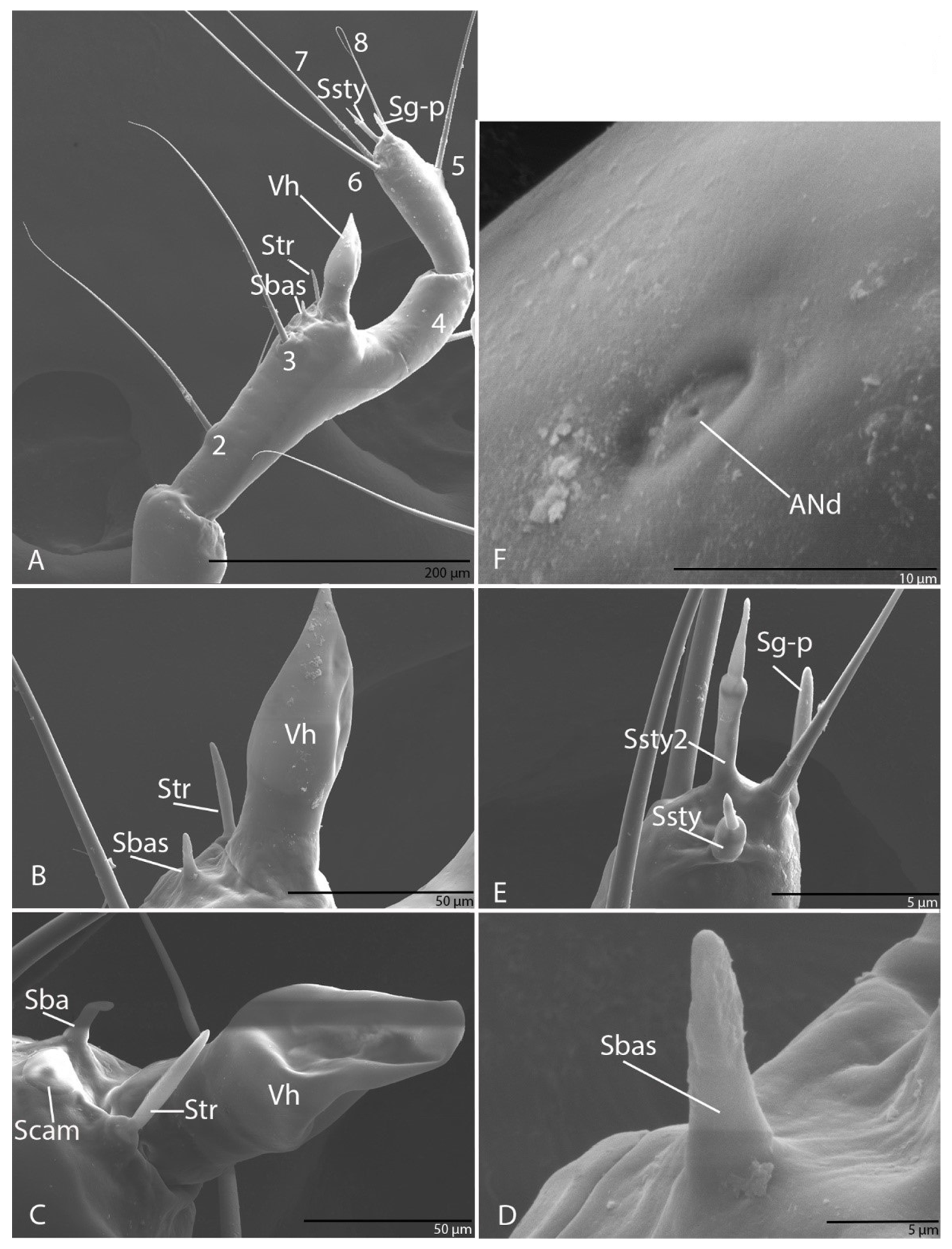
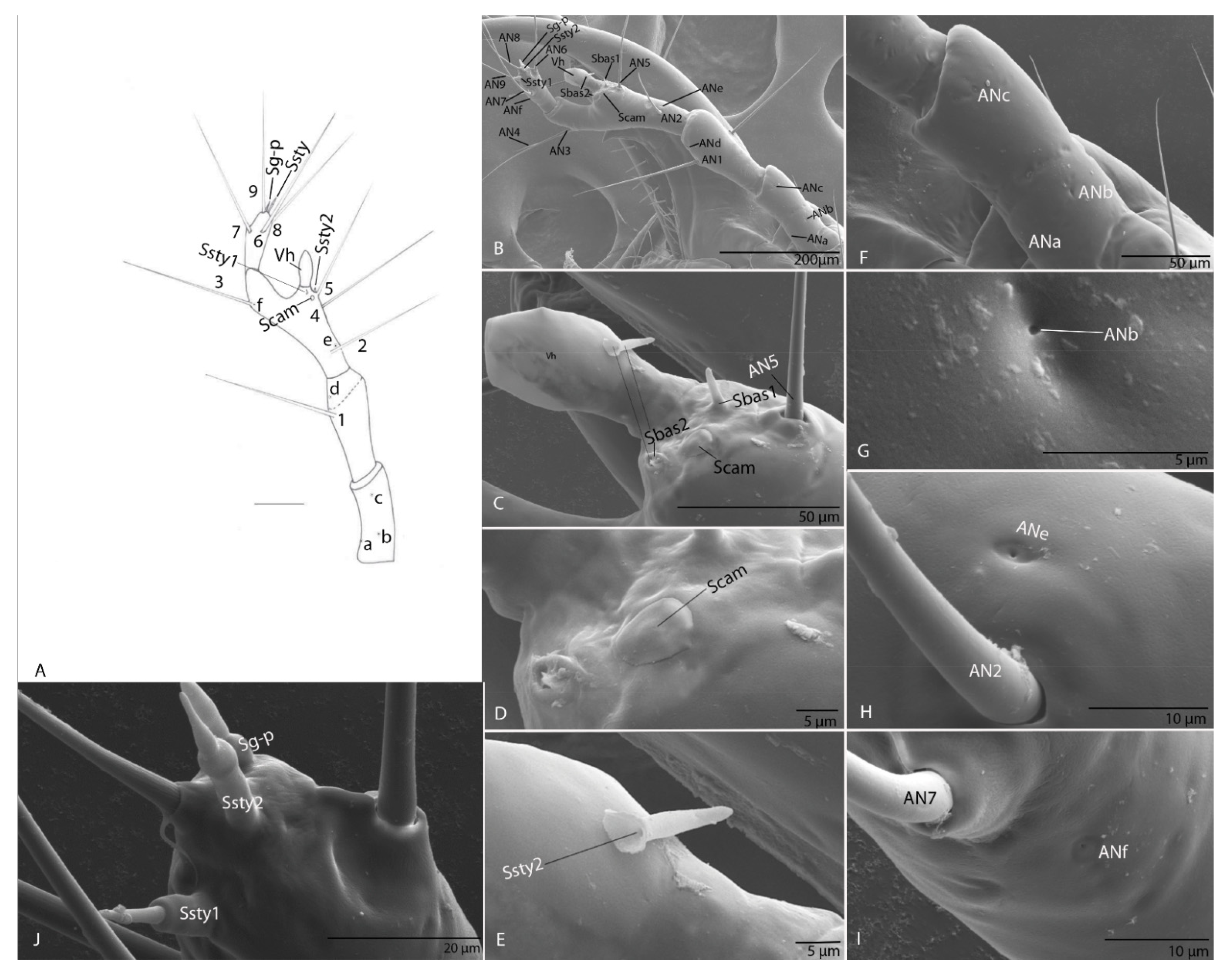
3.1.5. Antenna
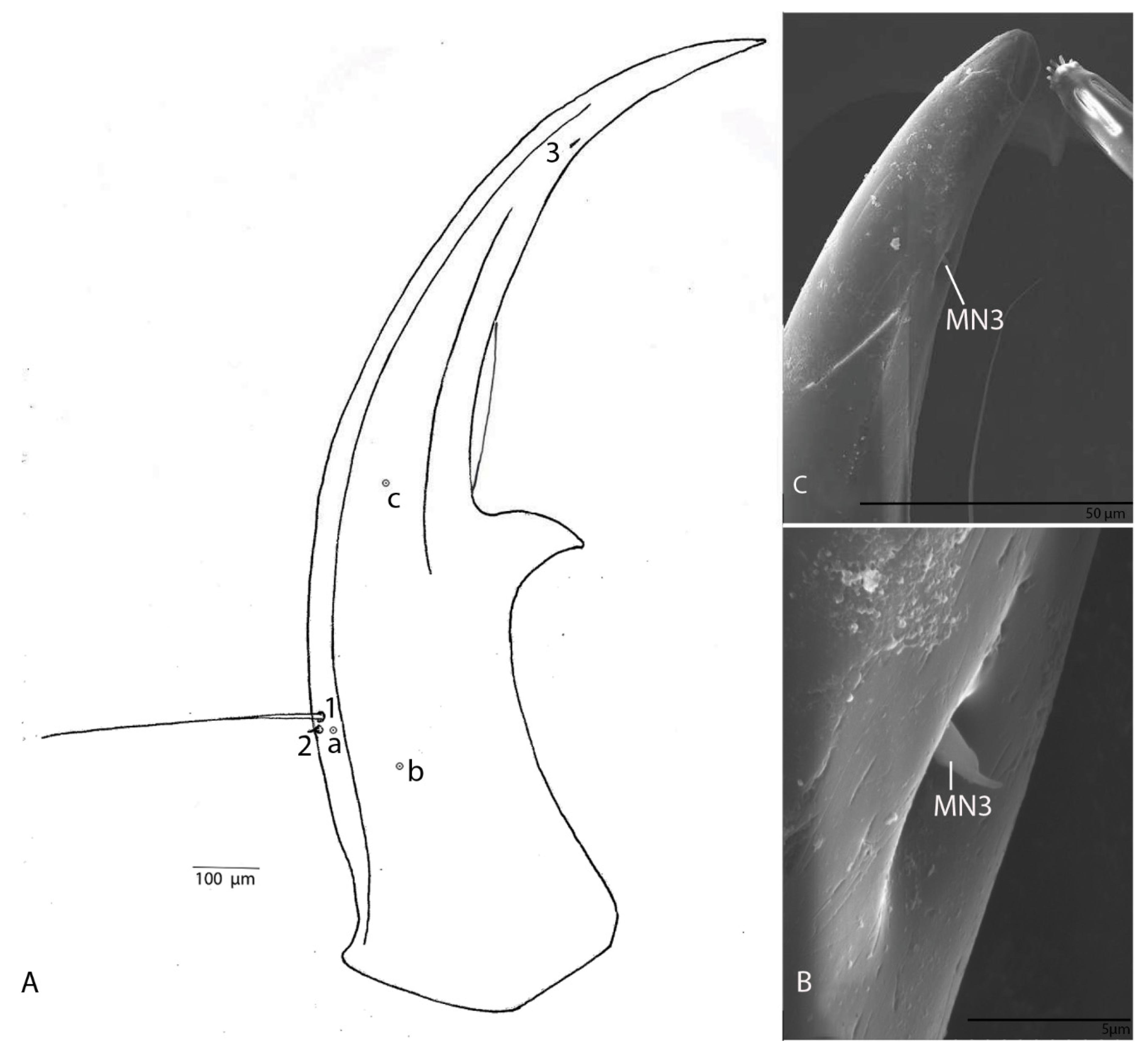
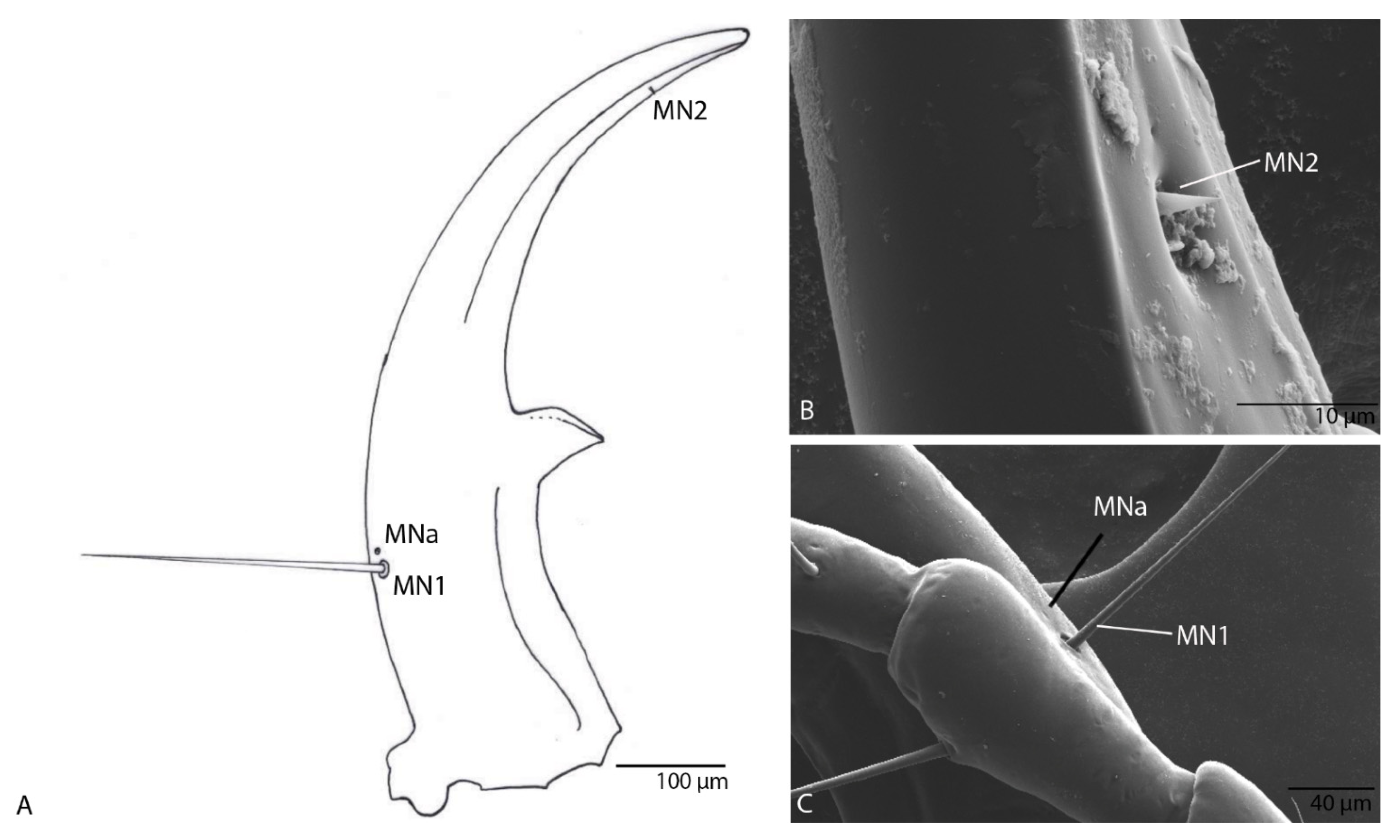
3.1.6. Mandible
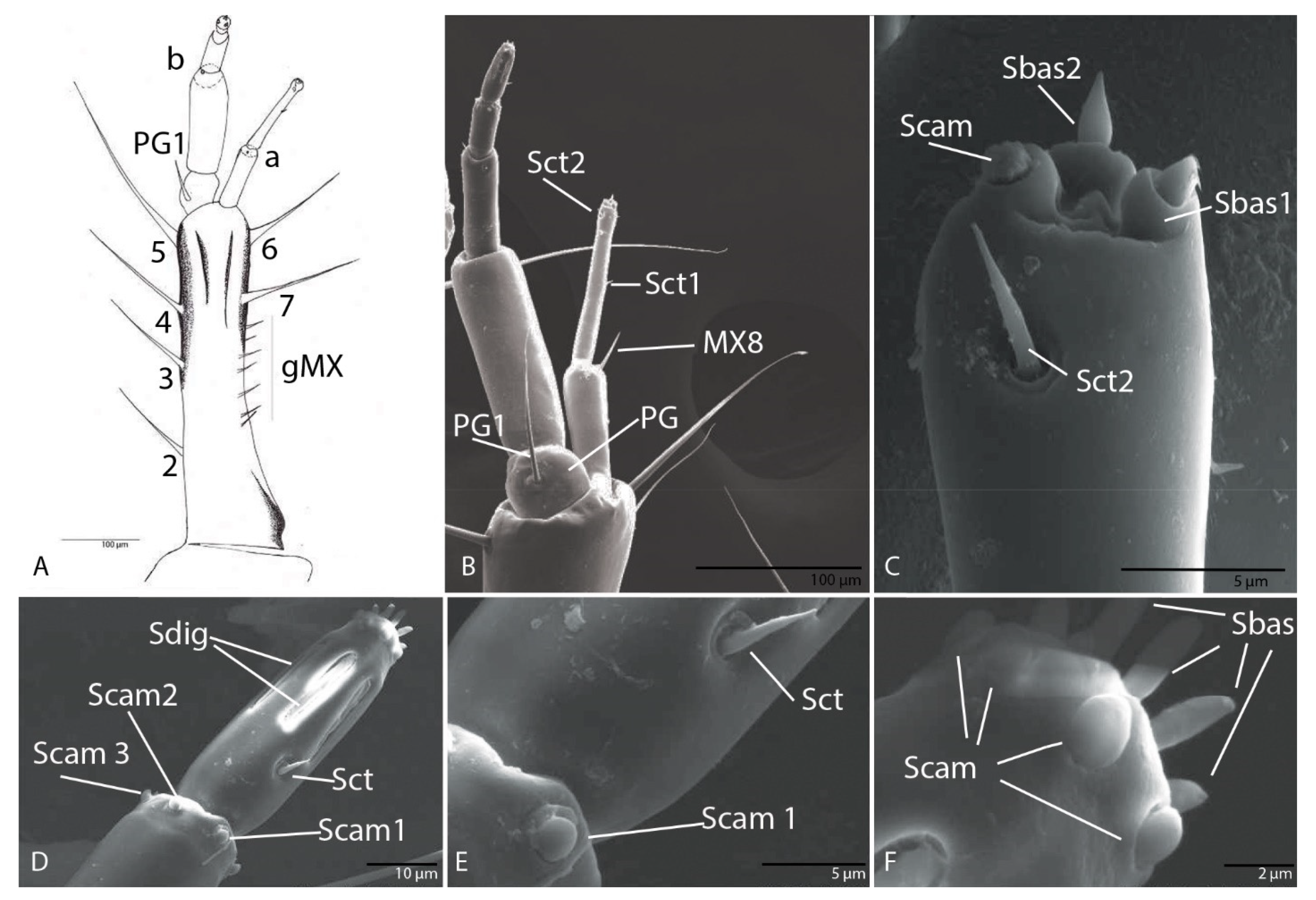
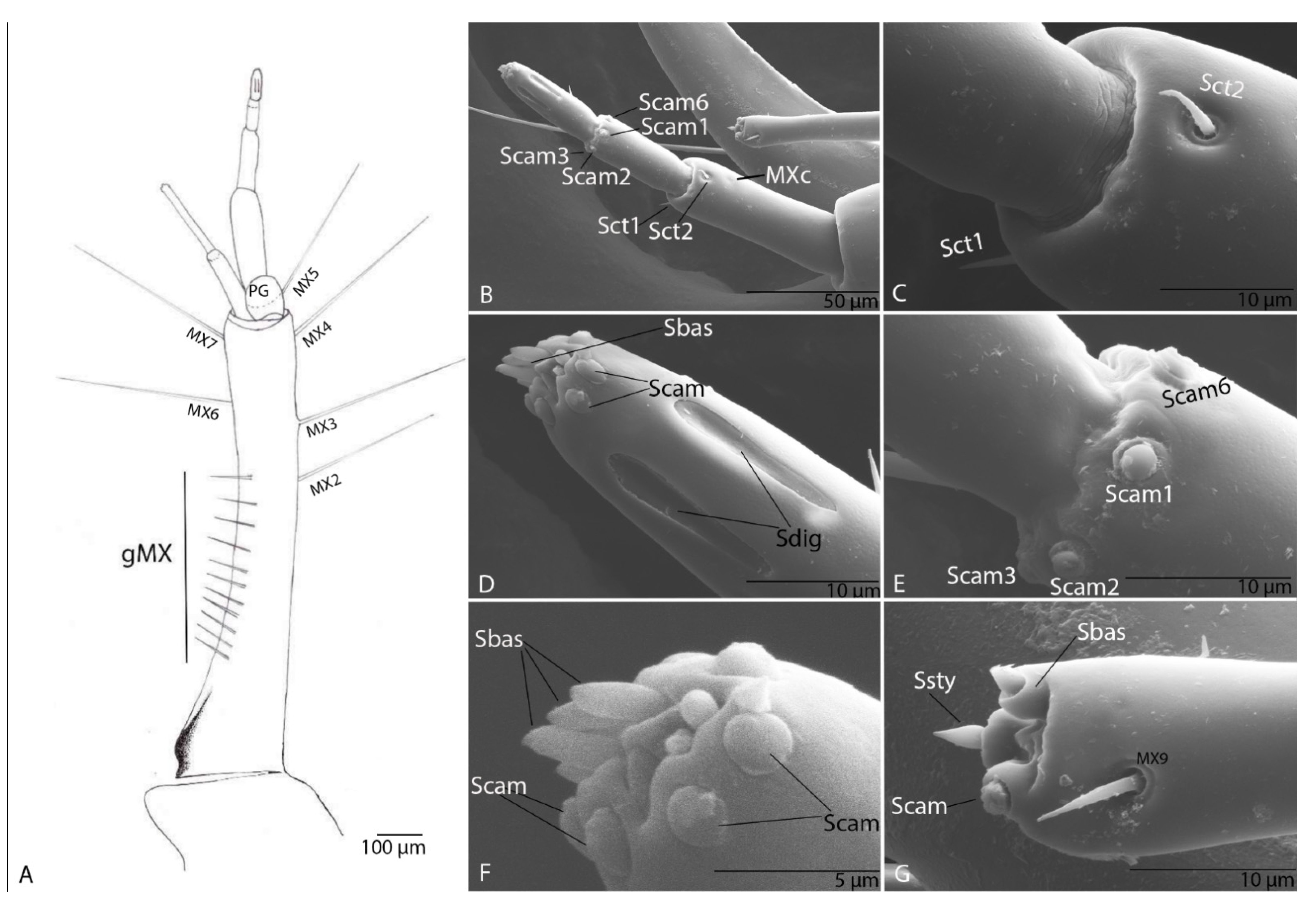
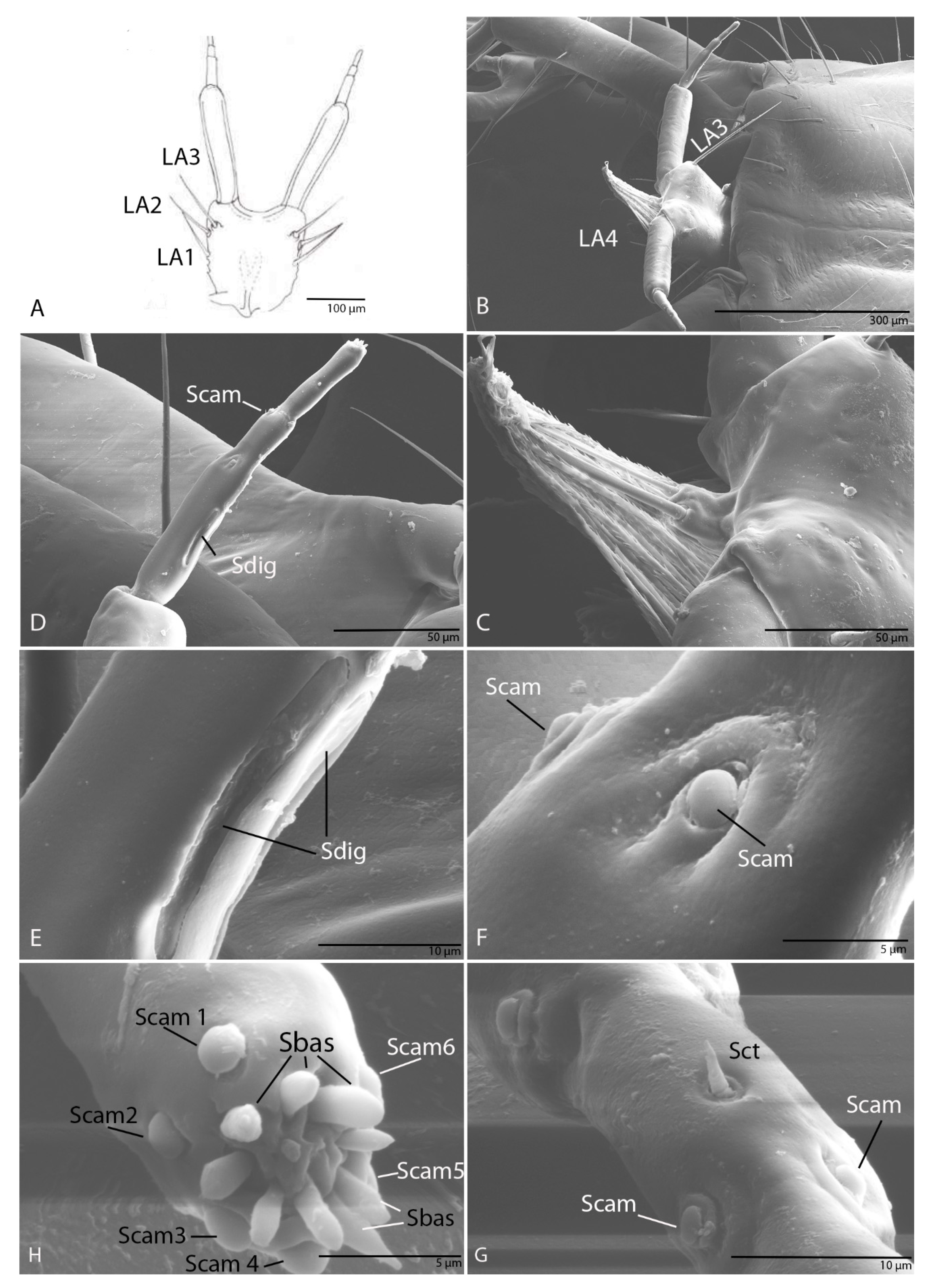
3.1.7. Maxilla
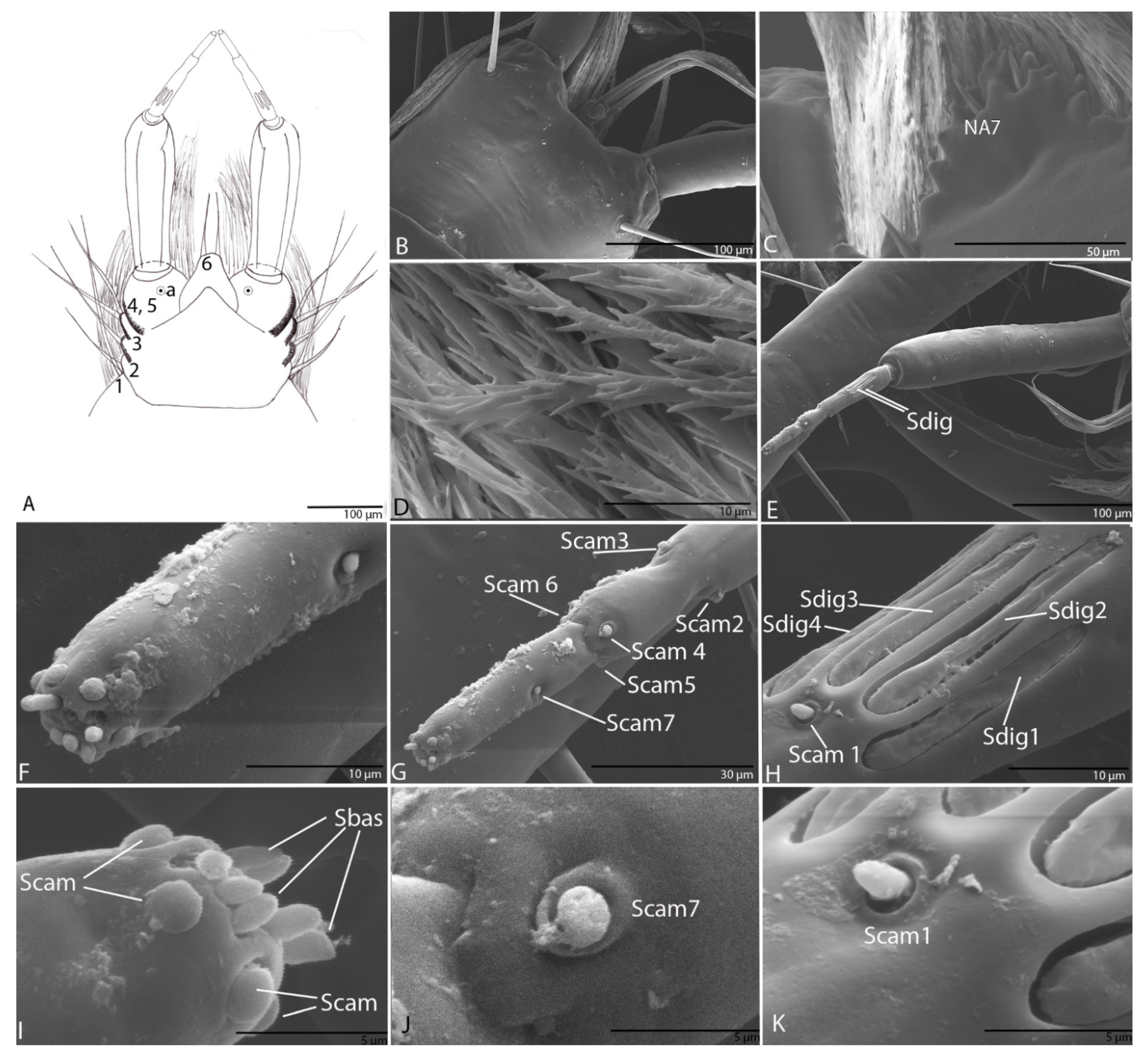
3.1.8. Labium
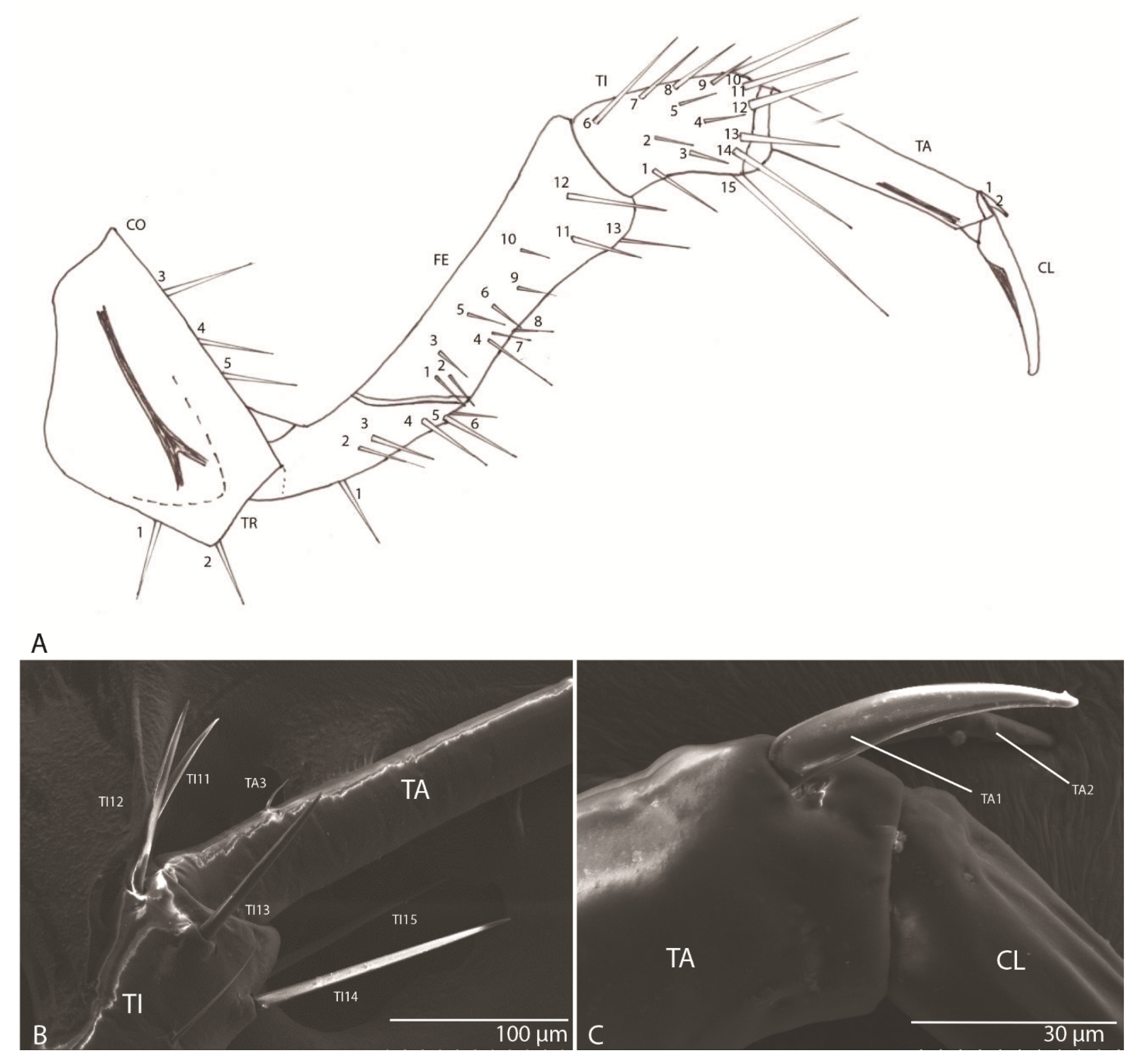
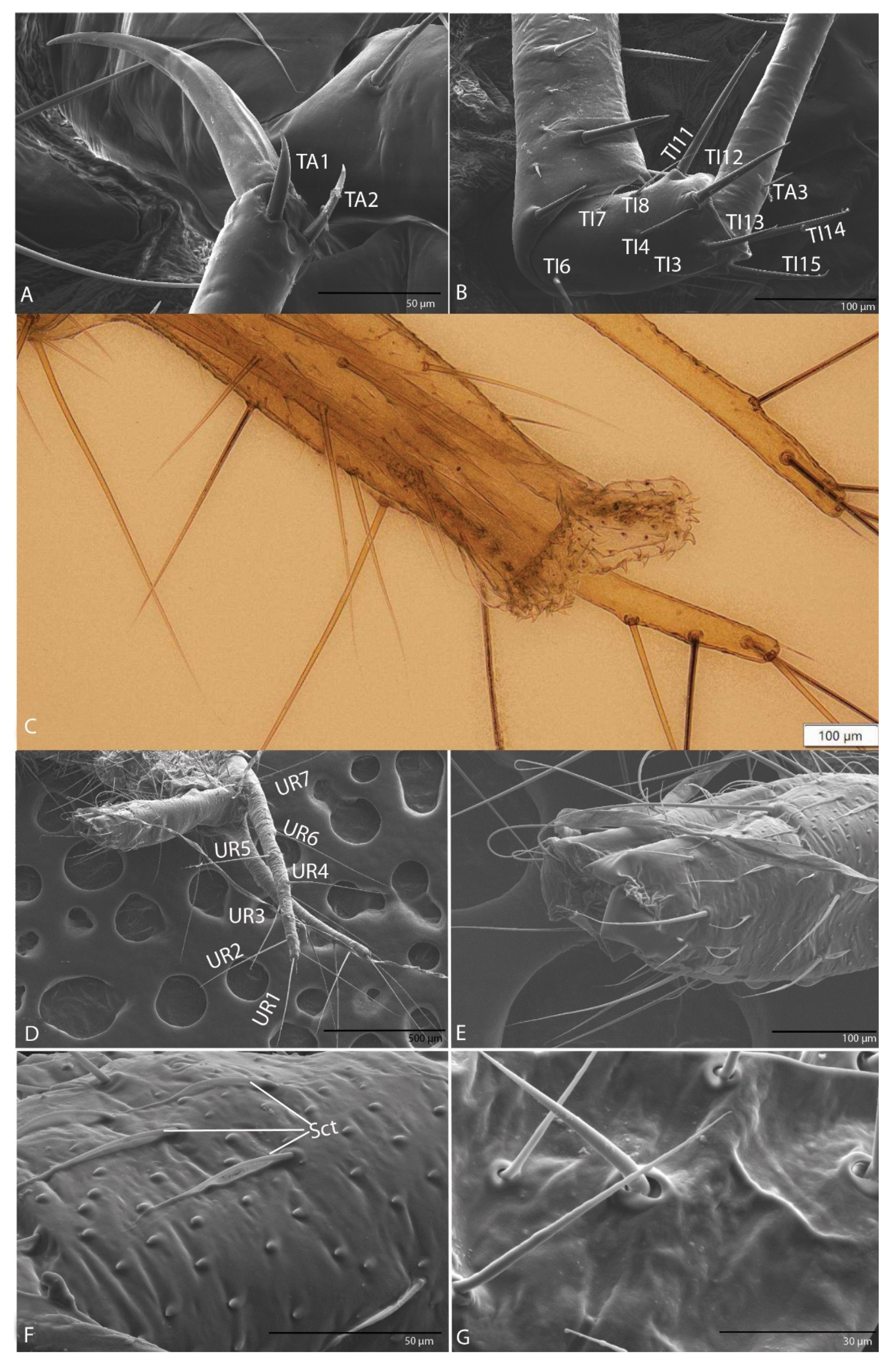
3.1.9. Leg
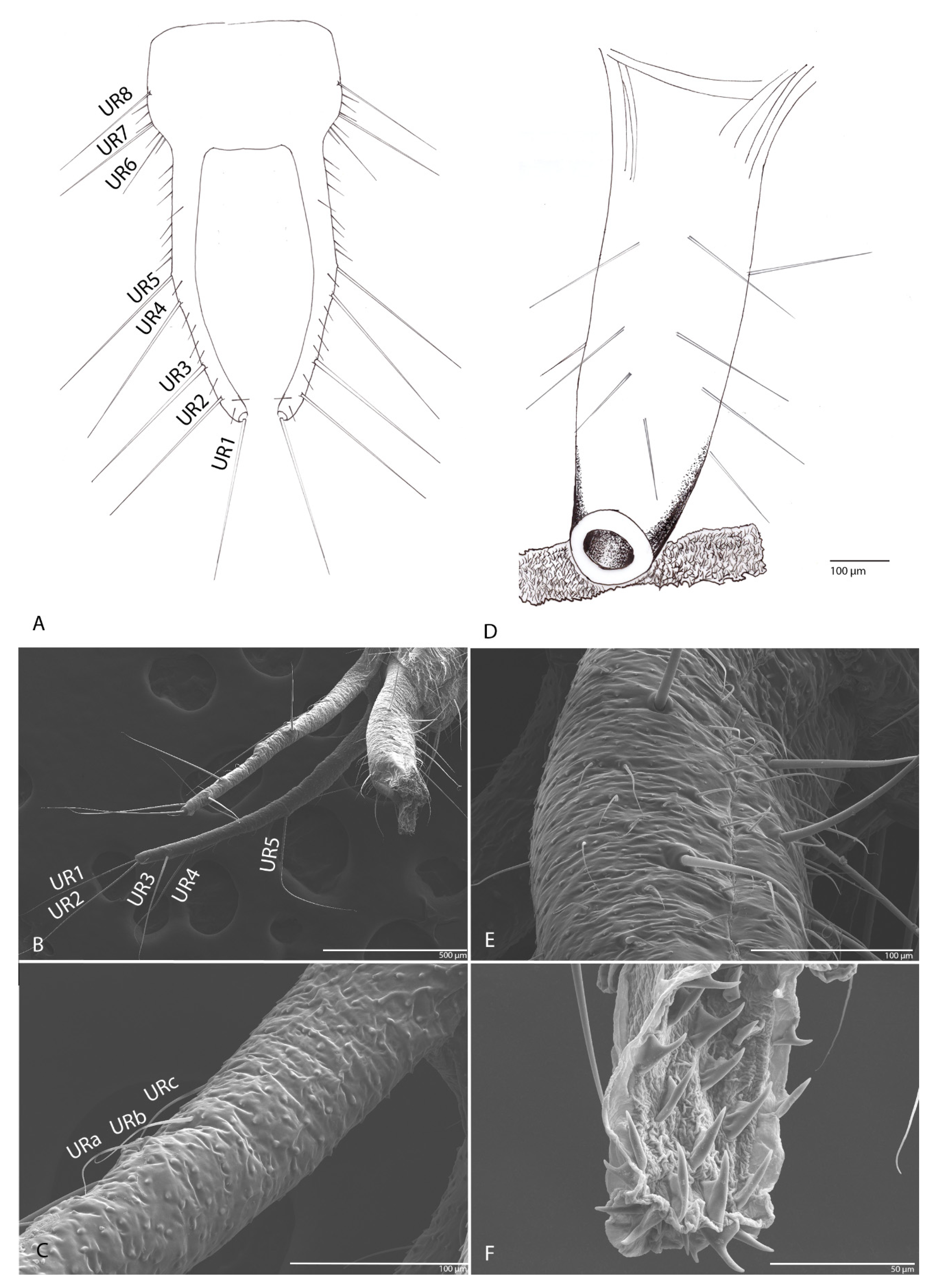
3.1.10. Urogomphi
3.1.11. Pygidium
3.2. Description of the Duvalius (Biharotrechus) paroecus (J. Frivaldszkyi, 1865) Larva
3.2.1. Habitus
3.2.2. Head
3.2.3. The Nasale (Anterior Margin of Epistome)
3.2.4. Chaetotaxy of Cephalic Capsule
3.2.5. Antenna
3.2.6. Mandible
3.2.7. Maxilla
3.2.8. Labium
3.2.9. Leg
3.2.10. Urogomphi
3.2.11. Pygidium
4. Discussion and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Casale, A.; Taglianti, A.V.; Juberthie, C. Coleoptera carabidae. In Encyclopaedia Biospéologica; CNRS: Paris, France, 1998; Volume 2, pp. 1047–1081. [Google Scholar]
- Moravec, P.; Uéno, S.I.; Belousov, I.A. Carabidae: Trechinae: Trechini. In Catalogue of Palaearctic Coleoptera; Löbl, I., Smetana, A., Eds.; Apollo Books: Stenstrup, Denmark, 2003; Volume 1, pp. 288–346. [Google Scholar]
- Faille, A.; Casale, A.; Balke, M.; Ribera, I. A molecular phylogeny of Alpine subterranean Trechini (Coleoptera: Carabidae). BMC Evol. Biol. 2013, 13, 248. [Google Scholar] [CrossRef] [Green Version]
- Jeannel, R. Coleopteres carabiques, I, II. In Faune de France; Lechevalier: Paris, France, 1941; pp. 1–571. [Google Scholar]
- Available online: https://www.gbif.org/species/4406363 (accessed on 13 March 2021).
- Ortuño, V.; Reboleira, A.S. Description of the larva and female genitalia of Trechus gamae with data on its ecology. Bull. Insectol. 2011, 64, 43–52. [Google Scholar]
- Makarov, K.V.; Koval, A.G. A contribution to the knowledge of the biology of a troglobiont Carabid species, Jeannelius birsteini Ljovuschkin, 1963 (Coleoptera, Carabidae, Trechini). Entomol. Rev. 2003, 83, 819–826. [Google Scholar]
- Boldori, L. Altri appunti sulle larve dei Trechini. Mem. Soc. Ent. Ital. 1932, 10, 149–167. [Google Scholar]
- Boldori, L. Considerazioni sui caratteri distintivi di larve di Carabidi. Atti. Acc. Naz. Ital. Ent. 1959, 7, 85–89. [Google Scholar]
- Jeannel, R. Les larves des Trechini. Biospéologica 42. Arch. Zool. Exp. Gen. 1920, 59, 509–542. [Google Scholar]
- Giachino, P.M. Morfologia larvale di alcuni Trechus alticoli delle Alpi Lepontine (Coleoptera Carabidae). Boll. Soc. Entomol. Ital. 1985, 117, 161–171. [Google Scholar]
- Giachino, P.M. Contributo alla conoscenza della morfologia larvale del genere Trechus. La larva di Trechus schatzmayri Focarile, 1949 (Coleoptera, Carabidae). Riv. Piemont. St. Nat. 1989, 10, 131–135. [Google Scholar]
- Luff, M.L. The Carabidae (Coleoptera) larvae of Fennoscandia and Denmark. In Fauna Entomologica Scandinavica; Scandinavian Science Press: Leiden, Copenhagen, 1993; Volume 27, pp. 1–186. [Google Scholar]
- Casale, A.; Marcia, P. Larval morphology of Sardaphaenops adelphus Casale, 2004, a highly specialized troglobitic beetle endemic to Sardinia (Coleoptera, Carabidae). Subterr. Biol. 2007, 5, 35–42. [Google Scholar]
- Ortuño, V.; Reboleira, A.S. Description of the third instar larva of a hypogean ground beetle, Trechus alicantinus (Coleoptera: Carabidae: Trechinae). Entomol. Fenn. 2010, 21, 33–42. [Google Scholar] [CrossRef] [Green Version]
- Giachino, P.M.; Magnaghi, S.; Lana, E.; Meregalli, M.; Taglianti, A.V. Morfologia Degli Stadi Larvali di Alcuni Trechini Paleartici e Australiani (Coleoptera, Carabidae); Stab. Tipo-Litografico del Littorio: Genova, Italy, 2020; Volume 113–118, pp. 199–321. [Google Scholar]
- Jeannel, R. Monographie des Trechinae. Morphologie Comparee et Distribution geographique d’un Groupe de Coleopteres; Premiere livraison; Abeille: Paris, France, 1926; Volume 32, pp. 221–550. [Google Scholar]
- Bousquet, Y.; Goulet, H. Notation of primary setae and pores on larvae of Carabidae (Coleoptera: Adephaga). Can. J. Zool. 1984, 62, 573–588. [Google Scholar] [CrossRef]
- Makarov, K.V. A key to the genera of the Groundbeetle larvae (Coleoptera, Carabidae) of the Palearctic region. Boll. Mus. Reg. Sci. Nat. Torino 1994, 12, 221–254. [Google Scholar]
- Grebennikov, V.V.; Maddison, D.R. Phylogenetic analysis of Trechitae (Coleoptera: Carabidae) based on larval morphology, with a description of first-instar Phrypeus and a key to genera. Syst. Entomol. 2005, 30, 38–59. [Google Scholar] [CrossRef]
- Solodovnikov, A.Y. Larval chaetotaxy of Coleoptera (Insecta) as a tool for evolutionary research and systematics: Less confusion, more clarity. J. Zoolog. Syst. Evol. Res. 2007, 45, 120–127. [Google Scholar] [CrossRef]
- Altner, H.; Bauer, T. Ultrastructure of a specialized, thrust-sensitive, insect mechanoreceptor: Stimulus-transmitting structures and sensory apparatus in the rostral horns of Notiophilus biguttatus. Cell Tissue Res. 1982, 226, 337–354. [Google Scholar] [CrossRef]
- Bauer, T.; Kredler, M. Adhesive mouthparts in a ground beetle larva (Coleoptera, Carabidae, Loricera pilicornis F.) and their function during predation. Zool. Anz. 1988, 221, 145–156. [Google Scholar]
- Moore, W.; Di Giulio, A. Out of the burrow and into the nest: Functional anatomy of three life history stages of Ozaena lemoulti (Coleoptera:Carabidae) reveals an obligate life with ants. PLoS ONE 2019, 14, e0209790. [Google Scholar] [CrossRef]
- Li, W.B.; Liu, N.Y.; Xu, Q.; Sun, M.; Fang, J.; Wang, S.Y. Ultrastructure structure of antennal sensilla of carabid beetle Carabus elysii Thomson, 1856 (Coleoptera: Carabidae). Entomol. Res. 2019, 49, 81–86. [Google Scholar] [CrossRef] [Green Version]
- Giglio, A.; Ferrero, E.A.; Perrotta, E.; Tripepi, S.; Zetto-Brandmayr, T. Ultrastructure and comparative morphology of mouth-part sensilla in ground beetle larvae (Insecta, Coleoptera, Carabidae). Zool. Anz. 2003, 242, 277–292. [Google Scholar] [CrossRef]
- Onac, B.P.; Drăgușin, V.; Papiu, F.; Theodorescu, C.T. Rodna mountains: Izvorul tăusoarelor cave (pestera de la izvorul tăusoarelor). In Cave and Karst Systems of Romania; Cave and Karst Systems of the World; Ponta, G., Onac, B., Eds.; Springer: Amsterdam, The Netherlands, 2019. [Google Scholar]
- Rusu, T. Carstul din Munţii Pădurea Craiului; Editura Dacia: Cluj, Romania, 1988; p. 254. [Google Scholar]
- Viehmann, I.; Racoviță, G.; Crăciun, C.S.A. Cluj-Napoca. In Carstul din Munţii Pădurea Craiului; Rusu, T., Ed.; Ed Dacia: Cluj-Napoca, Romania, 1988; 254p. [Google Scholar]
- Jeannel, R. Les Fossiles Vivants des Cavernes; Gallimard: Paris, France, 1943; p. 321. [Google Scholar]
- Emden, F.I. A key to the genera of larval Carabidae. Trans. R. Ent. Soc. 1942, 92, 1–99. [Google Scholar] [CrossRef]
- Sharova, I.K. Larvae of the carabids useful and harmful in agriculture. Uch. Zapiski Mosk. Gos. Pedag. Inst. Im. Lenina 1958, 12, 4–164. [Google Scholar]
- Sharova, I.K. Familia Carabidae—Carabid beetles. In Key to Soil Dwelling Insect Larvae; Ghilarov, M.S., Ed.; Nauka: Moscow, Russia, 1964; pp. 112–195. (In Russian) [Google Scholar]
- Luff, M.L. The larvae of the British Carabidae (Coleoptera) VII. Trechini and Pogonini. Entomol. Gaz. 1985, 36, 301–314. [Google Scholar]
- Arndt, E.; Grebennikov, V.V.; Zaballos, J. Description of the larvae of a representative of anillina with a key to the palaearctic genera of bembidiini. Koleopt. Runds 1999, 69, 11–17. [Google Scholar]
- Carabajal, E.; García, J.; Rodríguez, F. Descripción de un nuevo género y una nueva especie de Trechini (Coleoptera: Caraboidea: Trechidae) de la cordillera cantábrica. Elytron 1999, 13, 123–131. [Google Scholar]
- Laneyrie, R. Sur la systématique des Trechinae (Coleoptera: Trechidae). Nouv. Rev. Entomol. 1974, 4, 3–21. [Google Scholar]
- Moldovan, O.; Jalzic, B.; Erichsen, E. Diversity of mouthparts in some European representatives of subterranean Cholevidae (Coleoptera). Nat. Croat. 2004, 13, 1–18. [Google Scholar]
- Giglio, A.; Brandmayr, P.; Ferrero, E.A.; Giulianini, P.G.; Perrotta, E.; Talarico, F.F.; Zetto-Brandmayr, T. Ultrastructure of the antennal sensorial appendage of larvae of Ophonus ardosiacus (Lutshnik, 1922) (Coleoptera, Carabidae) and possible correlations between size and shape and the larval feeding habits. Zool. Anz. 2008, 247, 209–221. [Google Scholar] [CrossRef]
- Altner, H.; Prillinger, L. Ultrastructure of invertebrate chemo-, thermo-, and hygroreceptors and its functional significance. In International Review of Cytology; Academic Press: Cambridge, MA, USA, 1980; Volume 67, pp. 69–139. [Google Scholar]
- Keil, T.A. Morphology and development of the peripheral olfactory organs. In Insect Olfaction; Hansson, B.S., Ed.; Springer: Berlin/Heidelberg, Germany, 1999; pp. 22–47. [Google Scholar]
- Steinbrecht, R.A. Structure and function of insect olfactory sensilla. In Ciba Foundation Symposium 200-Olfaction in Mosquito-Host Interactions: Olfaction in Mosquito-Host Interactions: Ciba Foundation Symposium 200; John Wiley & Sons, Ltd.: Chichester, UK, 1996; pp. 158–174. [Google Scholar]
- Zacharuk, R.Y.; Mittler, T.E.; Radovsky, F.J.; Resh, V.H. Ultrastructure and function of insect chemosensilla. Annu. Rev. Entomol. 1980, 25, 27–47. [Google Scholar] [CrossRef]
- Altner, H.; Sass, H.; Altner, I. Relationship between structure and function of antennal chemo-, hygro-, and thermoreceptive sensilla in Periplaneta americana. Cell Tissue Res. 1977, 176, 389–405. [Google Scholar] [CrossRef] [PubMed]
- Zacharuk, R.Y. Antennae and sensilla. In Comprehensive Insect Physiology, Biochemistry and Pharmacology; Kerkut, G.A., Gilbert, L.I., Eds.; Pergamon Press: London, UK, 1985; pp. 1–70. [Google Scholar]
- Altner, H.; Loftus, R. Ultrastructure and function of insect thermo-and hygroreceptors. Annu. Rev. Entomol. 1985, 30, 273–295. [Google Scholar] [CrossRef]
- Steinbrecht, R.A.; Kittmann, R. Funktionelle Morphologie eines elektrophysiologisch identifizierten Hygrorezeptors beim Seidenspinner, Bombyx mori. Verh. Dtsch. Zool. Ges. 1986, 79, 111. [Google Scholar]
- Carle, T.; Toh, Y.; Yamawaki, Y.; Watanabe, H.; Yokohari, F. The antennal sensilla of the praying mantis Tenodera aridifolia: A new flagellar pseudo partition based on the antennal macro-, micro- and ultrastructures. Arthropod Struct. Dev. 2014, 43, 103–116. [Google Scholar] [CrossRef] [PubMed]
- Honomichl, K.; Guse, G.W. Digitiform sensilla on the maxillar palp of coleoptera: III fine structure in Tenebrio molitor L. and Dermestes maculatus De Geer. Acta Zool. 1981, 62, 17–25. [Google Scholar] [CrossRef]
- Honomichl, K. Die digitiformen sensillen auf dem maxillarpalpus von coleoptera. I. Vergleichend-topographisce untersuchung des kutikulären apparates. Zool. Anz. 1980, 204, 1–12. [Google Scholar]
- Bloom, J.W.; Zacharuk, R.Y.; Holodniuk, A.E. Ultrastructure of the larval antenna of Tenebrio molitor L. (Coleoptera: Tenebrionidae): Structure of the blunt-tipped peg and papillate sensilla. Can. J. Zool. 1982, 60, 1545–1556. [Google Scholar] [CrossRef]
- Bland, R.G.; Slaney, D.P.; Weinstein, P. Antennal sensilla on cave species of Australian Paratemnopteryx cockroaches (Blattaria: Blattellidae). Int. J. Insect Morphol. Embryol. 1998, 27, 83–93. [Google Scholar] [CrossRef]
- Lamoreux, J.; Morrison, J.; Ricketts, T.H.; Olson, D.M.; Dinerstein, E.; McKnight, M.W.; Shugart, H.H. Global tests of biodiversity concordance and the importance of endemism. Nature 2006, 440, 212–214. [Google Scholar] [CrossRef]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef]
- Milić, D.; Radenković, S.; Ačanski, J.; Vujić, A. The importance of hidden diversity for insect conservation: A case study in hoverflies (the Merodon atratus complex, Syrphidae, Diptera). J. Insect Conserv. 2019, 23, 29–44. [Google Scholar] [CrossRef]
- Carvalho, S.B.; Brito, J.C.; Crespo, E.J.; Possingham, H. From climate change predictions to actions—Conserving vulnerable animal groups in hotspots at a regional scale. Glob. Change Biol. 2010, 16, 3257–3270. [Google Scholar] [CrossRef]
- Thomas, C.D.; Cameron, A.; Green, R.E.; Bakkenes, M.; Beaumont, L.J.; Collingham, Y.C.; Erasmus, B.F.N.; de Siqueira, F.M.; Grainger, A.; Hannah, L.; et al. Extinction risk from climate change. Nature 2004, 427, 145–148. [Google Scholar] [CrossRef] [PubMed]
- Ceballos, G.; García, A.; Ehrlich, P.R. The sixth extinction crisis: Loss of animal populations and species. J. Cosmol. 2010, 8, 1821–1831. [Google Scholar]
- Ceballos, G.; Ehrlich, P.R.; Barnosky, A.D.; García, A.; Pringle, R.M.; Palmer, T.M. Accelerated modern human-induced species losses: Entering the sixth mass extinction. Sci. Adv. 2015, 1, e1400253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moldovan, O.T.; Iepure, S.; Brad, T.; Kenesz, M.; Mirea, I.C.; Năstase-Bucur, R. Database of Romanian cave invertebrates with a red list of cave species and a list of hotspot/coldspot caves. Biodiv. Data 2020, 8, e53571. [Google Scholar] [CrossRef] [PubMed]
- Brandmayr, P.; Giorgi, F.; Casale, A.; Colombetta, G.; Mariotti, L.; Vigna Taglianti, A.; Weber, F.; Pizzolotto, R. Hypogean carabid beetles as indicators of global warming? Environ. Res. Lett. 2013, 8, 044047. [Google Scholar] [CrossRef] [Green Version]
















Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sitar, C.; Barbu-Tudoran, L.; Moldovan, O.T. Morphological and Micromorphological Description of the Larvae of Two Endemic Species of Duvalius (Coleoptera, Carabidae, Trechini). Biology 2021, 10, 627. https://doi.org/10.3390/biology10070627
Sitar C, Barbu-Tudoran L, Moldovan OT. Morphological and Micromorphological Description of the Larvae of Two Endemic Species of Duvalius (Coleoptera, Carabidae, Trechini). Biology. 2021; 10(7):627. https://doi.org/10.3390/biology10070627
Chicago/Turabian StyleSitar, Cristian, Lucian Barbu-Tudoran, and Oana Teodora Moldovan. 2021. "Morphological and Micromorphological Description of the Larvae of Two Endemic Species of Duvalius (Coleoptera, Carabidae, Trechini)" Biology 10, no. 7: 627. https://doi.org/10.3390/biology10070627