
Ice Nucleation Activity of Alpine Bioaerosol Emitted in Vicinity of a Birch Forest

 , , , and
, , , and
Abstract
:1. Introduction
2. Experiments
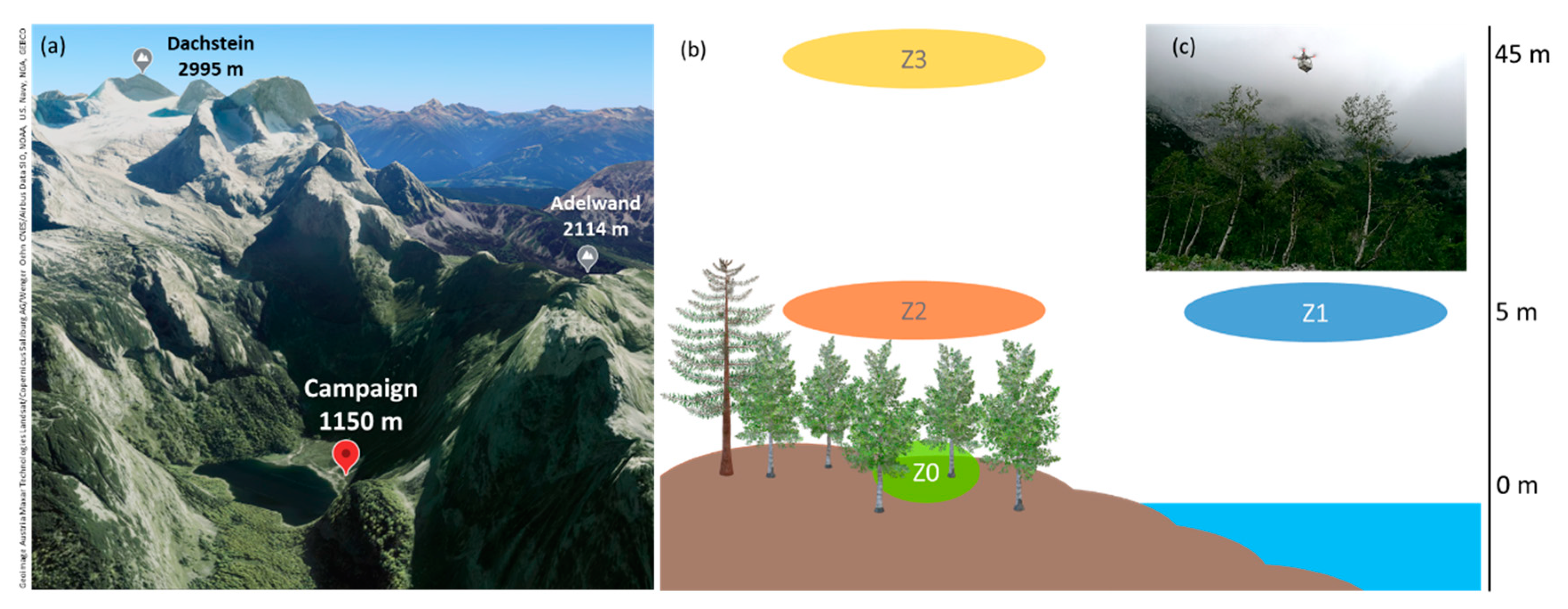
2.1. Field Campaign
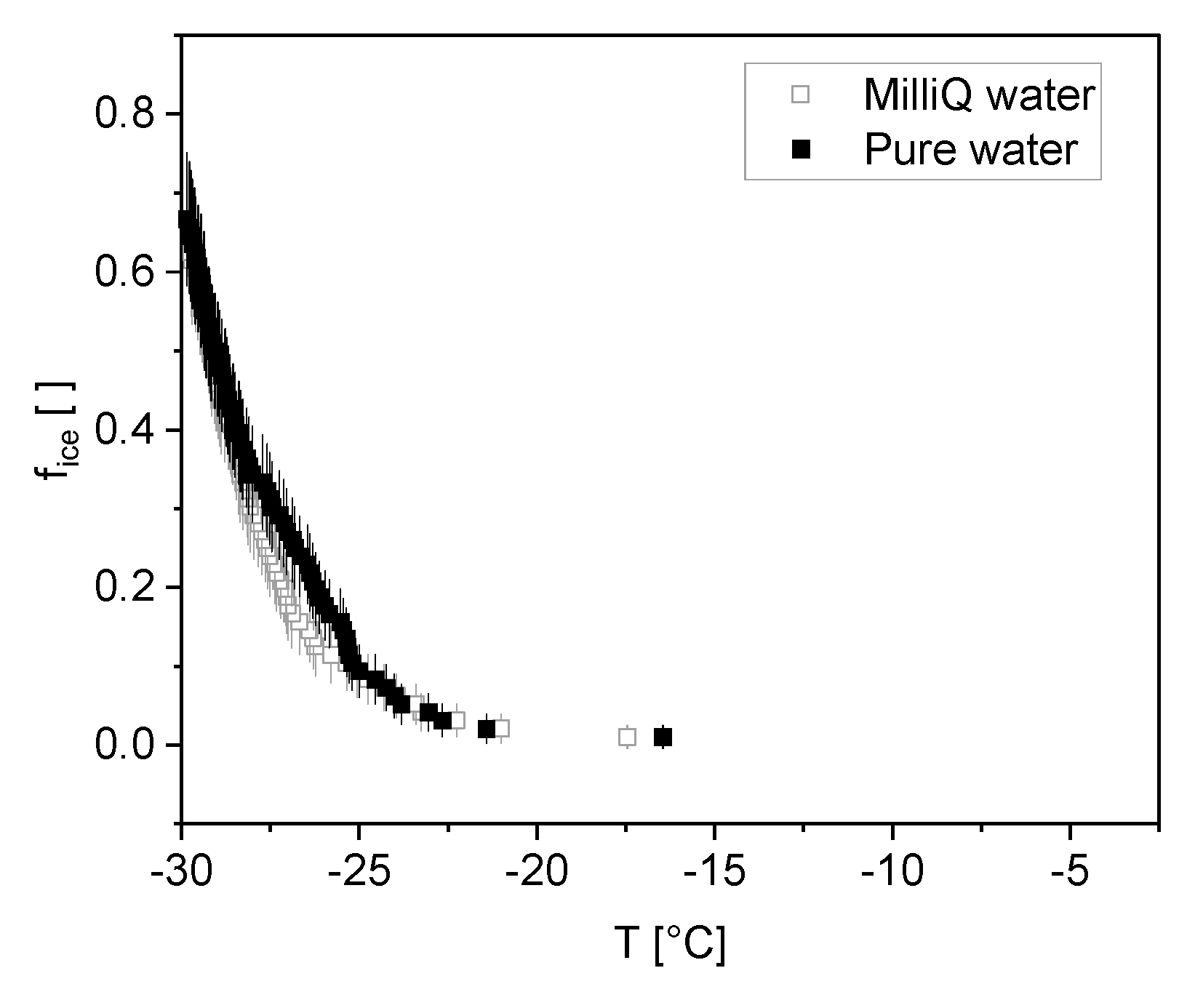
2.2. Ice Nucleation
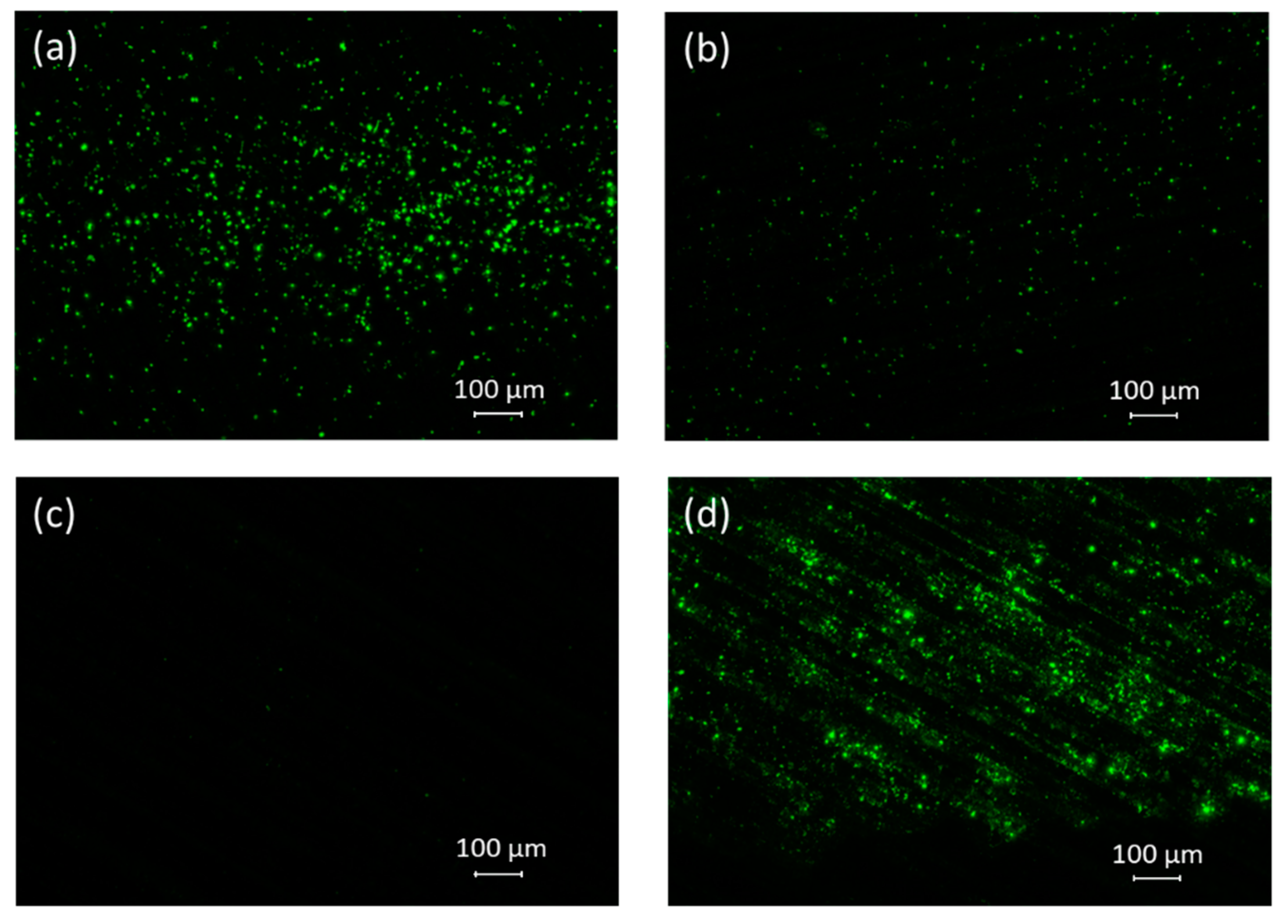
2.3. Microscopy (Fluorescence, SEM)
2.4. Aerosol Particle Characterization
3. Results
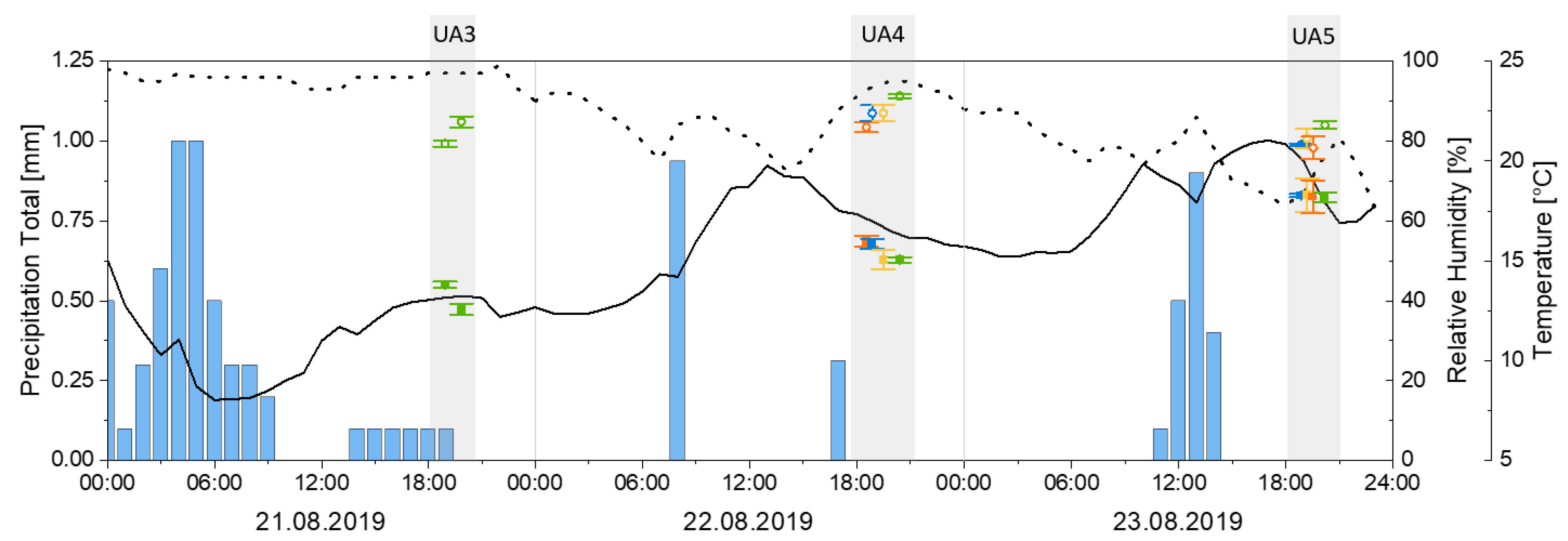
3.1. Meteorological and Environmental Data
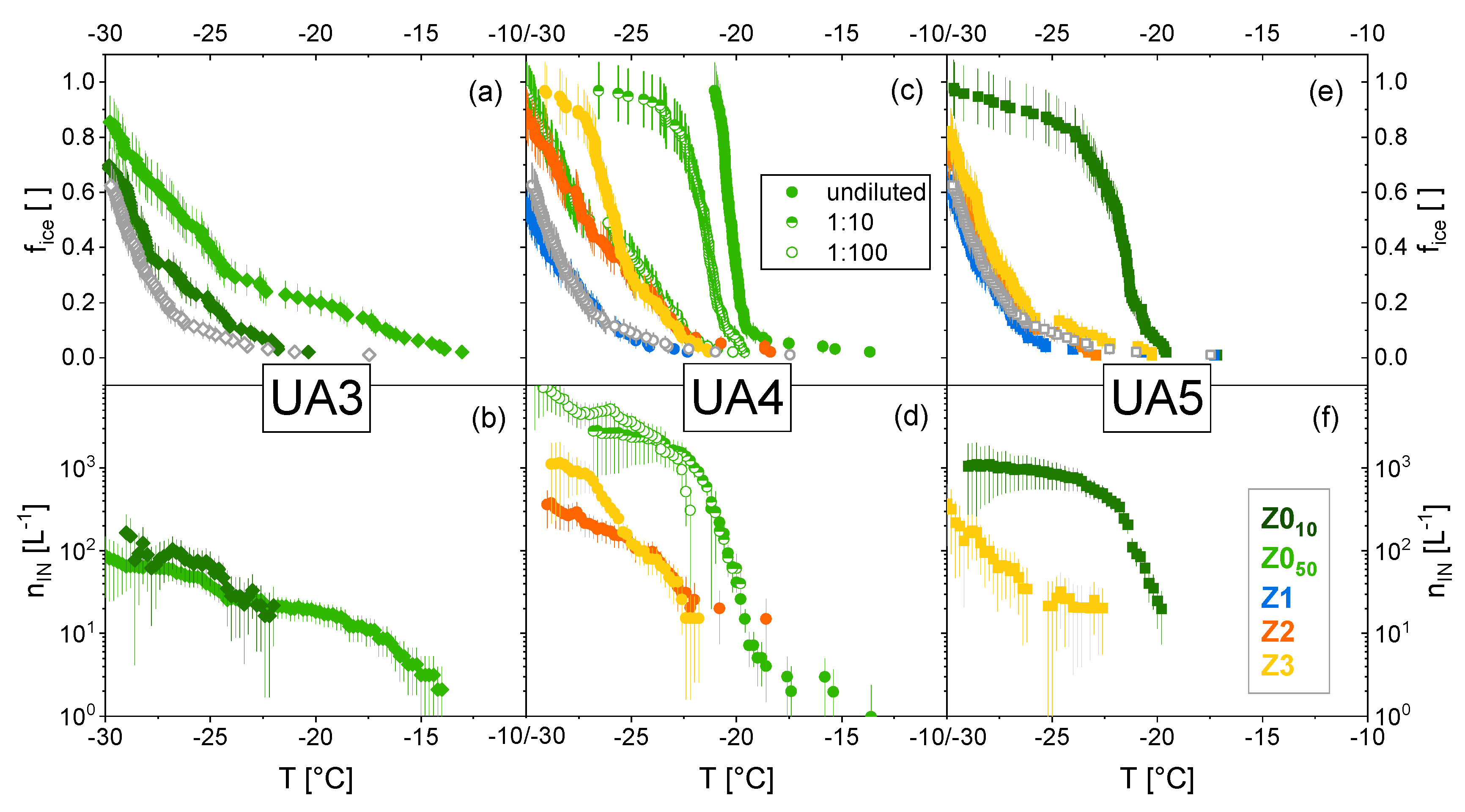
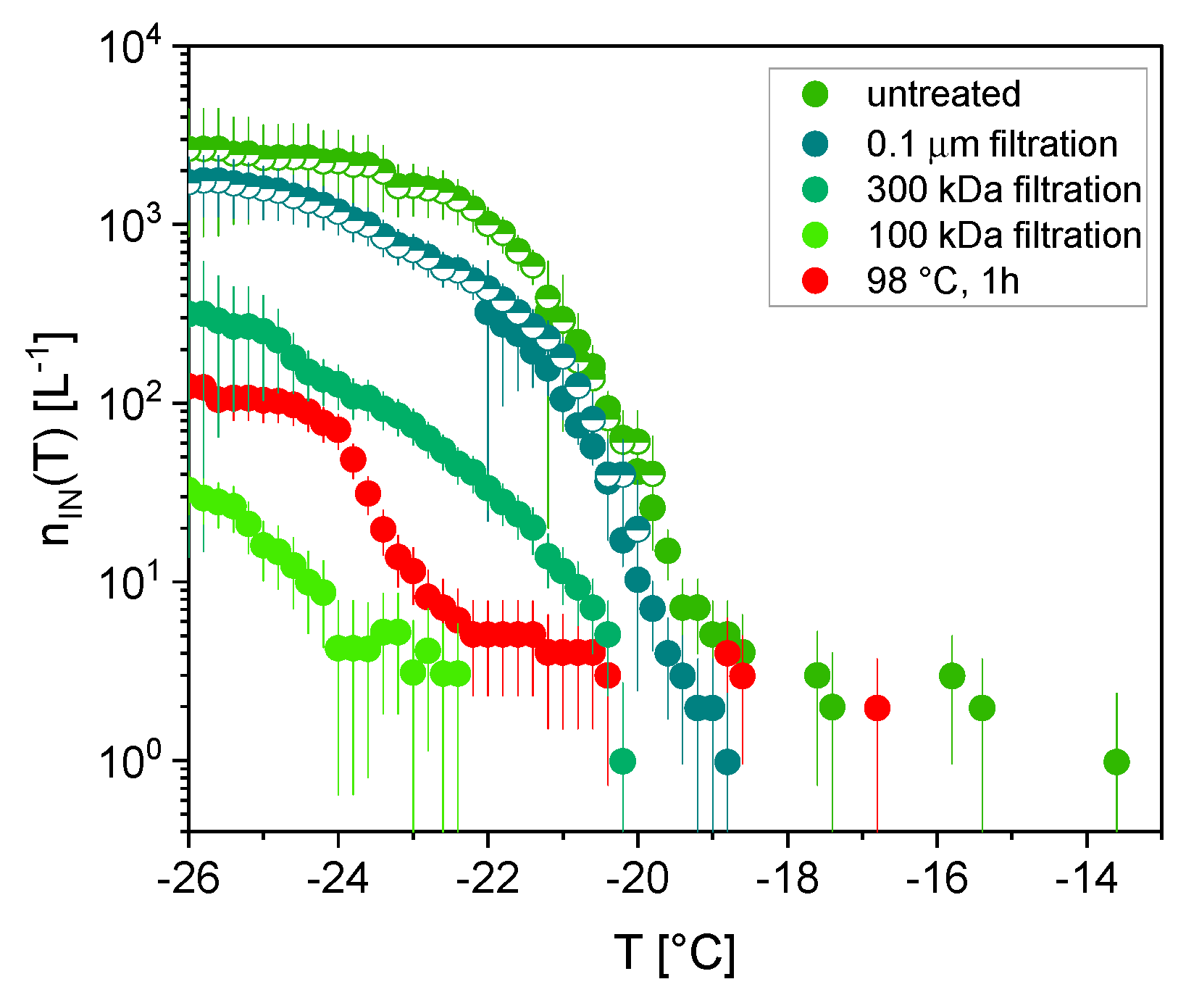
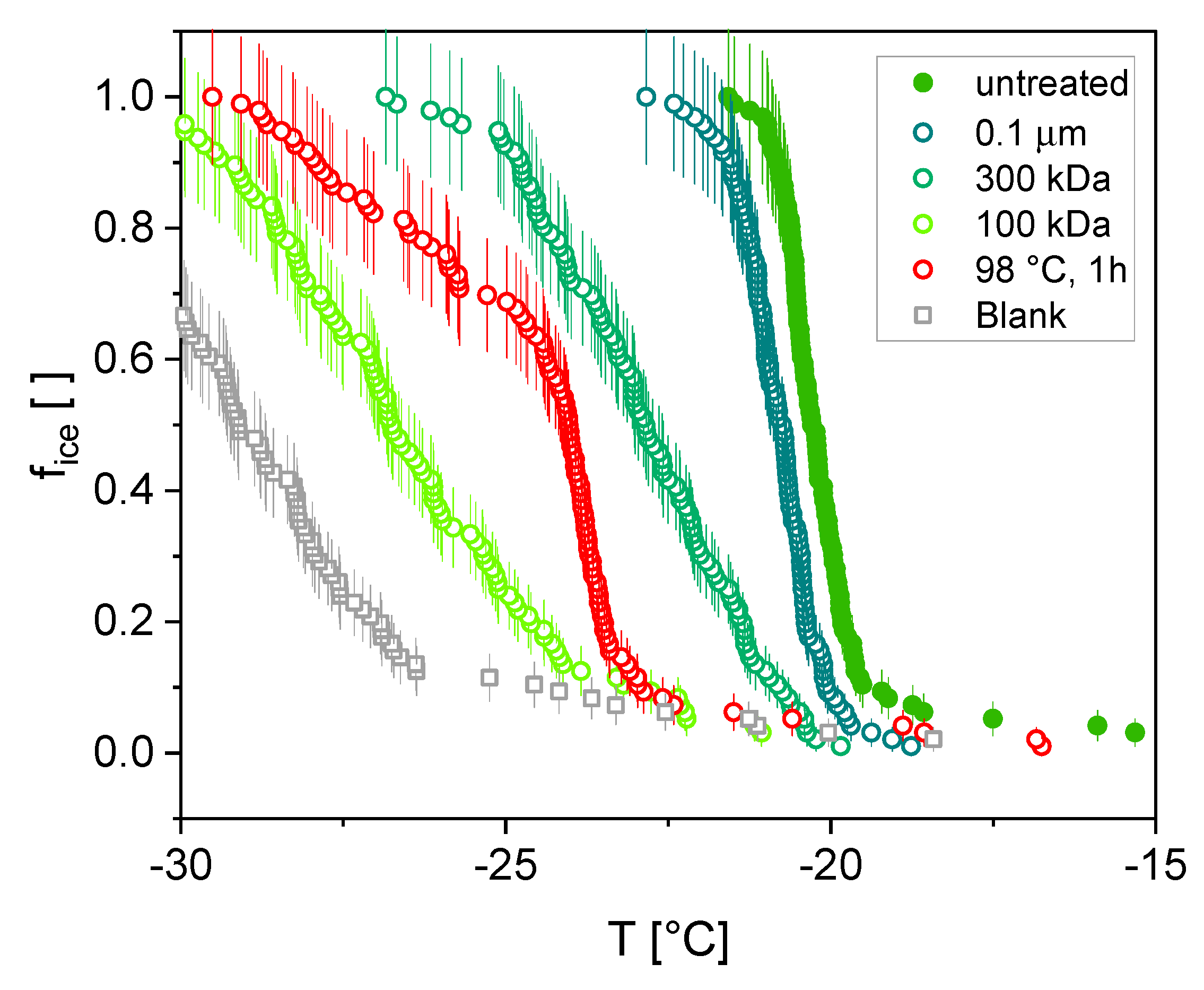
3.2. Ice Nucleation
3.3. Microscopy (Fluorescence, SEM)
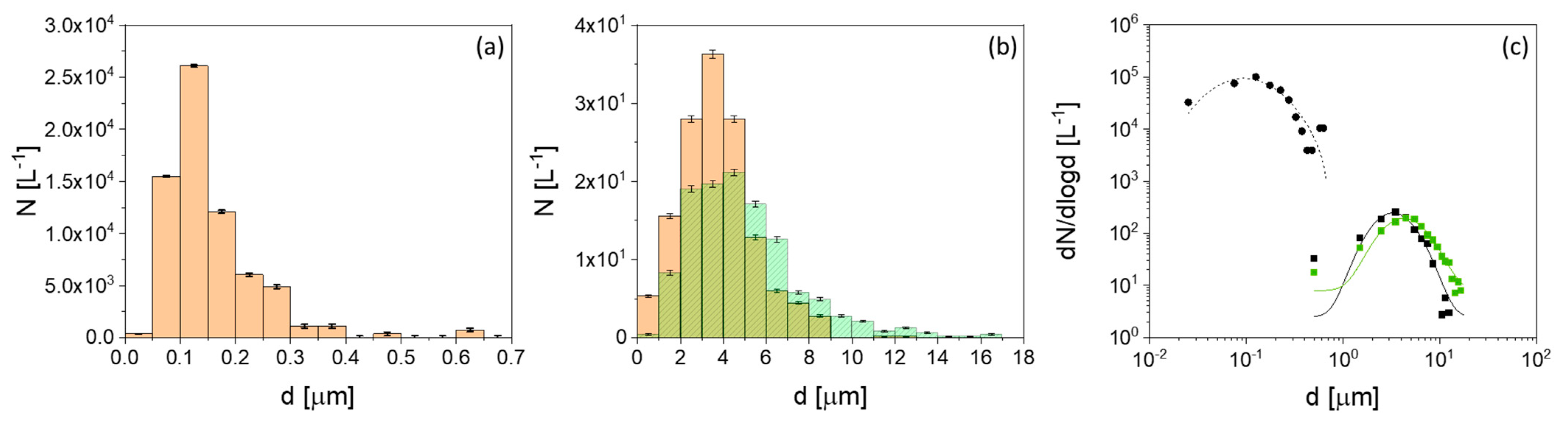
3.4. Aerosol Particle Characterization
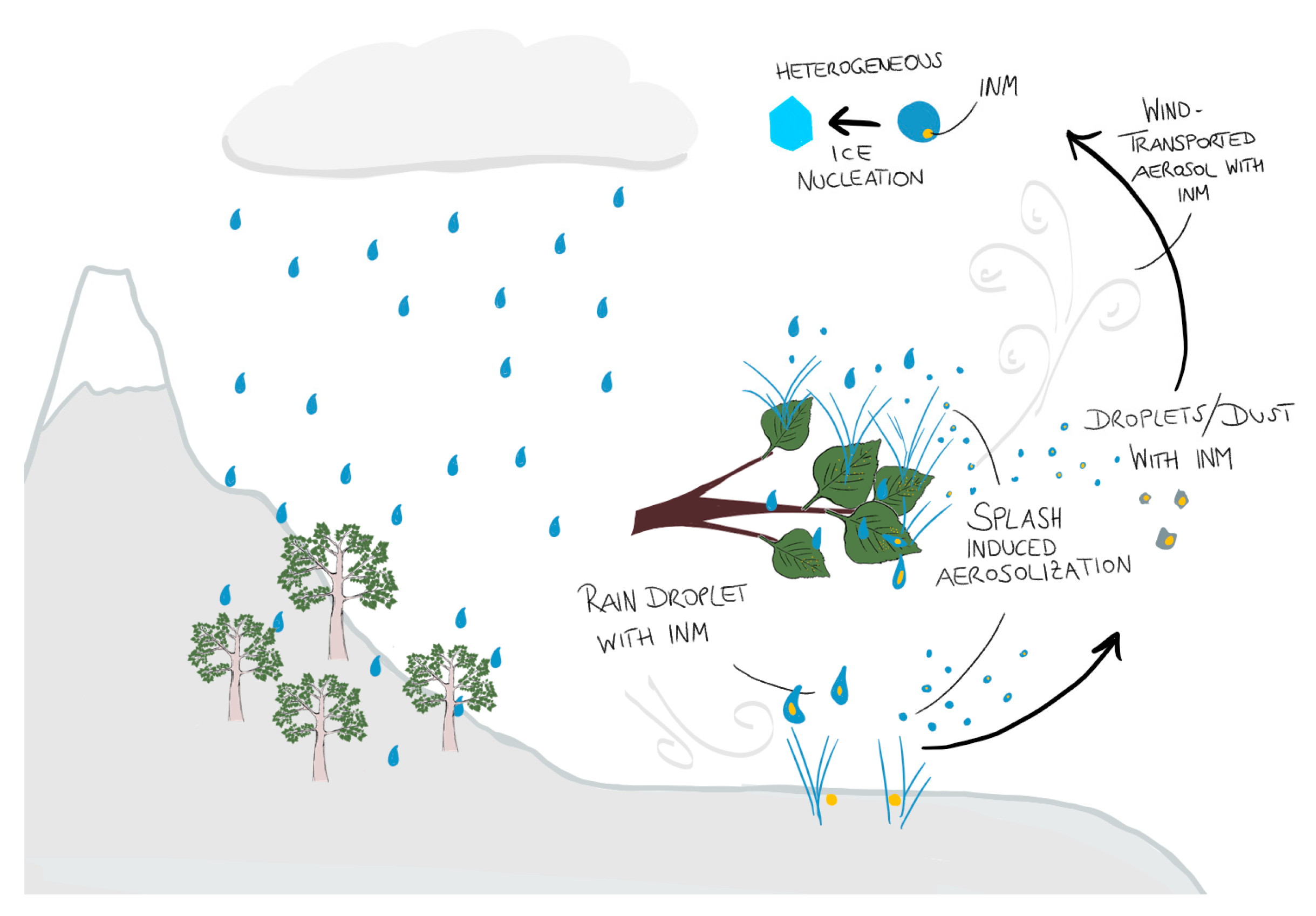
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A







References
- Morris, E.C.; Sands, D.C.; Vinatzer, A.B.; Glaux, C.; Guilbaud, C.; Buffière, A.; Yan, S. The life history of the plant pathogen Pseudomonas syringae is linked to the water cycle. ISME J. 2008, 2, 321–334. [Google Scholar] [CrossRef] [Green Version]
- Benson, J.; Hanlon, R.; Seifried, T.M.; Baloh, P.; Powers, C.W.; Grothe, H.; Schmale, D.G. Microorganisms Collected from the Surface of Freshwater Lakes Using a Drone Water Sampling System (DOWSE). Water 2019, 11, 157. [Google Scholar] [CrossRef] [Green Version]
- Felgitsch, L.; Baloh, P.; Burkart, J.; Mayr, M.; Momken, M.E.; Seifried, T.M.; Winkler, P.; Iii, D.G.S.; Grothe, H. Birch leaves and branches as a source of ice-nucleating macromolecules. Atmos. Chem. Phys. Discuss. 2018, 18, 16063–16079. [Google Scholar] [CrossRef] [Green Version]
- Pruppacher, H.R. A New Look at Homogeneous Ice Nucleation in Supercooled Water Drops. J. Atmos. Sci. 1995, 52, 1924–1933. [Google Scholar] [CrossRef] [Green Version]
- Matus, A.V.; L’Ecuyer, T.S. The role of cloud phase in Earth’s radiation budget. J. Geophys. Res. Atmos. 2017, 122, 2559–2578. [Google Scholar] [CrossRef]
- Boucher, O. Clouds and Aerosols. In Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Stocker, T.F., Qin, D., Plattner, G.-K., Tignor, M., Allen, S.K., Boschung, J., Nauels, A., Xia, Y., Bex, V., Midgley, P.M., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013. [Google Scholar]
- Creamean, J.M.; Suski, K.J.; Rosenfeld, D.; Cazorla, A.; DeMott, P.J.; Sullivan, R.; White, A.B.; Ralph, F.M.; Minnis, P.; Comstock, J.; et al. Dust and Biological Aerosols from the Sahara and Asia Influence Precipitation in the Western U.S. Science 2013, 339, 1572–1578. [Google Scholar] [CrossRef] [Green Version]
- DeMott, P.J.; Prenni, A.J. New Directions: Need for defining the numbers and sources of biological aerosols acting as ice nuclei. Atmos. Environ. 2010, 44, 1944–1945. [Google Scholar] [CrossRef]
- Petters, M.D.; Wright, T.P. Revisiting ice nucleation from precipitation samples. Geophys. Res. Lett. 2015, 42, 8758–8766. [Google Scholar] [CrossRef]
- Bigg, E.K.; Miles, G.T. The Results of Large–Scale Measurements of Natural Ice Nuclei. J. Atmos. Sci. 1964, 21, 396–403. [Google Scholar] [CrossRef] [Green Version]
- Isono, K.; Tanaka, T. Sudden Increase of Ice Nucleus Concentration Associated with Thunderstorm. J. Meteorol. Soc. Jpn. 1966, 44, 255–259. [Google Scholar] [CrossRef] [Green Version]
- Huffman, J.A.; Prenni, A.J.; DeMott, P.J.; Pöhlker, C.; Mason, R.H.; Robinson, N.H.; Fröhlich-Nowoisky, J.; Tobo, Y.; Després, V.R.; Garcia, E.; et al. High concentrations of biological aerosol particles and ice nuclei during and after rain. Atmos. Chem. Phys. Discuss. 2013, 13, 6151–6164. [Google Scholar] [CrossRef] [Green Version]
- Prenni, A.J.; Tobo, Y.; Garcia, E.; DeMott, P.J.; Huffman, J.A.; McCluskey, C.S.; Kreidenweis, S.M.; Prenni, J.; Pöhlker, C.; Pöschl, U. The impact of rain on ice nuclei populations at a forested site in Colorado. Geophys. Res. Lett. 2013, 40, 227–231. [Google Scholar] [CrossRef]
- Tobo, Y.; Prenni, A.J.; DeMott, P.J.; Huffman, J.A.; McCluskey, C.; Tian, G.; Pöhlker, C.; Pöschl, U.; Kreidenweis, S.M. Biological aerosol particles as a key determinant of ice nuclei populations in a forest ecosystem. J. Geophys. Res. Atmos. 2013, 118, 10–100. [Google Scholar] [CrossRef]
- Hara, K.; Maki, T.; Kobayashi, F.; Kakikawa, M.; Wada, M.; Matsuki, A. Variations of ice nuclei concentration induced by rain and snowfall within a local forested site in Japan. Atmos. Environ. 2016, 127, 1–5. [Google Scholar] [CrossRef]
- Iwata, A.; Imura, M.; Hama, M.; Maki, T.; Tsuchiya, N.; Kunihisa, R.; Matsuki, A. Release of Highly Active Ice Nucleating Biological Particles Associated with Rain. Atmosphere 2019, 10, 605. [Google Scholar] [CrossRef] [Green Version]
- Conen, F.; Eckhardt, S.; Gundersen, H.; Stohl, A.; Yttri, K.E. Rainfall drives atmospheric ice-nucleating particles in the coastal climate of southern Norway. Atmos. Chem. Phys. Discuss. 2017, 17, 11065–11073. [Google Scholar] [CrossRef] [Green Version]
- Knackstedt, K.A.; Moffett, B.F.; Hartmann, S.; Wex, H.; Hill, T.C.J.; Glasgo, E.D.; Reitz, L.A.; Augustin-Bauditz, S.; Beall, B.F.N.; Bullerjahn, G.S.; et al. Terrestrial Origin for Abundant Riverine Nanoscale Ice-Nucleating Particles. Environ. Sci. Technol. 2018, 52, 12358–12367. [Google Scholar] [CrossRef] [PubMed]
- O′sullivan, D.; Murray, B.J.; Ross, J.; Whale, T.F.; Price, H.C.; Atkinson, J.D.; Umo, N.S.; Webb, M.E. The relevance of nanoscale biological fragments for ice nucleation in clouds. Sci. Rep. 2015, 5, 8082. [Google Scholar] [CrossRef] [Green Version]
- Joung, Y.S.; Ge, Z.; Buie, C.R. Bioaerosol generation by raindrops on soil. Nat. Commun. 2017, 8, 14668. [Google Scholar] [CrossRef]
- Kim, S.; Park, H.; Gruszewski, H.A.; Schmale, D.G.; Jung, S. Vortex-induced dispersal of a plant pathogen by raindrop impact. Proc. Natl. Acad. Sci. USA 2019, 116, 4917–4922. [Google Scholar] [CrossRef] [Green Version]
- Blanchard, D.C.; Syzdek, L.D.; Weber, M.E. Bubble scavenging of bacteria in freshwater quickly produces bacterial enrichment in airborne jet drops1. Limnol. Oceanogr. 1981, 26, 961–964. [Google Scholar] [CrossRef]
- Spiel, D.E. The number and size of jet drops produced by air bubbles bursting on a fresh water surface. J. Geophys. Res. Space Phys. 1994, 99, 10289–10296. [Google Scholar] [CrossRef]
- Villermaux, E. Fragmentation. Annu. Rev. Fluid Mech. 2007, 39, 419–446. [Google Scholar] [CrossRef]
- Pietsch, R.B.; Grothe, H.; Hanlon, R.; Powers, C.W.; Jung, S.; Ross, S.D.; Schmale, D.G., III. Wind-driven spume droplet production and the transport of Pseudomonas syringae from aquatic environments. PeerJ 2018, 6, e5663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Powers, C.W.; Hanlon, R.; Grothe, H.; Prussin, A.J.I.; Marr, L.C.; Schmale, D.G.I. Coordinated Sampling of Microorganisms Over Freshwater and Saltwater Environments Using an Unmanned Surface Vehicle (USV) and a Small Unmanned Aircraft System (sUAS). Front. Microbiol. 2018, 9, 1668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elbert, W.; Taylor, P.E.; Andreae, M.O.; Pöschl, U. Contribution of fungi to primary biogenic aerosols in the atmosphere: Wet and dry discharged spores, carbohydrates, and inorganic ions. Atmos. Chem. Phys. Discuss. 2007, 7, 4569–4588. [Google Scholar] [CrossRef] [Green Version]
- Jones, A.M.; Harrison, R.M. The effects of meteorological factors on atmospheric bioaerosol concentrations—A review. Sci. Total. Environ. 2004, 326, 151–180. [Google Scholar] [CrossRef] [PubMed]
- Rockett, T.R.; Kramer, C.L. Periodicity and Total Spore Production by Lignicolous Basidiomycetes. Mycologia 1974, 66, 817–829. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, C.J.; Pöhlker, C.; Aalto, P.; Hiltunen, V.; Petäjä, T.; Kulmala, M.; Pöschl, U.; Huffman, J.A. Seasonal cycles of fluorescent biological aerosol particles in boreal and semi-arid forests of Finland and Colorado. Atmos. Chem. Phys. Discuss. 2013, 13, 11987–12001. [Google Scholar] [CrossRef] [Green Version]
- Kunert, A.T.; Pöhlker, M.L.; Tang, K.; Krevert, C.S.; Wieder, C.; Speth, K.R.; Hanson, L.E.; Morris, C.E.; Iii, D.G.S.; Pöschl, U.; et al. Macromolecular fungal ice nuclei in Fusarium: Effects of physical and chemical processing. Biogeosciences 2019, 16, 4647–4659. [Google Scholar] [CrossRef] [Green Version]
- Pouleur, S.; Richard, C.; Martin, J.-G.; Antoun, H. Ice Nucleation Activity in Fusarium acuminatum and Fusarium avenaceum. Appl. Environ. Microbiol. 1992, 58, 2960–2964. [Google Scholar] [CrossRef] [Green Version]
- Diehl, K.; Quick, C.; Matthias-Maser, S.; Mitra, S.K.; Jaenicke, R. The ice nucleating ability of pollen Part I: Laboratory studies in deposition and condensation freezing modes. Atmos. Res. 2001, 58, 75–87. [Google Scholar] [CrossRef]
- Pummer, B.G.; Bauer, H.; Bernardi, J.; Bleicher, S.; Grothe, H. Suspendable macromolecules are responsible for ice nucleation activity of birch and conifer pollen. Atmos. Chem. Phys. Discuss. 2012, 12, 2541–2550. [Google Scholar] [CrossRef] [Green Version]
- Taylor, P.E.; Flagan, R.C.; Miguel, A.G.; Valenta, R.; Glovsky, M.M. Birch pollen rupture and the release of aerosols of respirable allergens. Clin. Exp. Allergy 2004, 34, 1591–1596. [Google Scholar] [CrossRef] [PubMed]
- Grote, M.; Valenta, R.; Reichelt, R. Abortive pollen germination: A mechanism of allergen release in birch, alder, and hazel revealed by immunogold electron microscopy. J. Allergy Clin. Immunol. 2003, 111, 1017–1023. [Google Scholar] [CrossRef]
- Burkart, J.; Gratzl, J.; Seifried, T.M.; Bieber, P.; Grothe, H. Subpollen particles (SPP) of birch as carriers of ice nucleating macromolecules. Biogeosciences Discuss. 2021, 1–15. [Google Scholar] [CrossRef]
- Seifried, T.M.; Bieber, P.; Felgitsch, L.; Vlasich, J.; Reyzek, F.; Iii, D.G.S.; Grothe, H. Surfaces of silver birch (Betula pendula) are sources of biological ice nuclei: In Vivo and in situ investigations. Biogeosciences 2020, 17, 5655–5667. [Google Scholar] [CrossRef]
- Bieber, P.; Seifried, T.M.; Burkart, J.; Gratzl, J.; Kasper-Giebl, A.; Schmale, D.G.; Grothe, H. A Drone-Based Bioaerosol Sampling System to Monitor Ice Nucleation Particles in the Lower Atmosphere. Remote Sens. 2020, 12, 552. [Google Scholar] [CrossRef] [Green Version]
- Kunert, A.T.; Lamneck, M.; Helleis, F.; Pöschl, U.; Pöhlker, M.L.; Fröhlich-Nowoisky, J. Twin-plate Ice Nucleation Assay (TINA) with infrared detection for high-throughput droplet freezing experiments with biological ice nuclei in laboratory and field samples. Atmos. Meas. Tech. 2018, 11, 6327–6337. [Google Scholar] [CrossRef] [Green Version]
- Vali, G. Quantitative Evaluation of Experimental Results and the Heterogeneous Freezing Nucleation of Supercooled Liquids. J. Atmos. Sci. 1971, 28, 402–409. [Google Scholar] [CrossRef]
- Vali, G. Revisiting the differential freezing nucleus spectra derived from drop-freezing experiments: Methods of calculation, applications, and confidence limits. Atmos. Meas. Tech. 2019, 12, 1219–1231. [Google Scholar] [CrossRef] [Green Version]
- David, R.O.; Cascajo-Castresana, M.; Brennan, K.P.; Rösch, M.; Els, N.; Werz, J.; Weichlinger, V.; Boynton, L.S.; Bogler, S.; Borduas-Dedekind, N.; et al. Development of the DRoplet Ice Nuclei Counter Zurich (DRINCZ): Validation and application to field-collected snow samples. Atmos. Meas. Tech. 2019, 12, 6865–6888. [Google Scholar] [CrossRef] [Green Version]
- Fröhlich-Nowoisky, J.; Hill, T.C.J.; Pummer, B.G.; Yordanova, P.; Franc, G.D.; Poeschl, U. Ice nucleation activity in the widespread soil fungus Mortierella alpina. Biogeosciences 2015, 12, 1057–1071. [Google Scholar] [CrossRef] [Green Version]
- Zachariassen, K.E.; Kristiansen, E. Ice Nucleation and Antinucleation in Nature. Cryobiology 2000, 41, 257–279. [Google Scholar] [CrossRef] [PubMed]
- Sakai, A.; Larcher, W. Frost Survival of Plants: Responses and Adaptation to Freezing Stress; Springer Science: Berlin, Germany, 1987; Volume 62, p. 321. [Google Scholar]
- Burke, M.J.; Gusta, L.V.; Quamme, H.A.; Weiser, C.J.; Li, P.H. Freezing and Injury in Plants. Annu. Rev. Plant Physiol. 1976, 27, 507–528. [Google Scholar] [CrossRef]
- Pearce, R.S. Plant Freezing and Damage. Ann. Bot. 2001, 87, 417–424. [Google Scholar] [CrossRef]
- Storey, J.M.; Storey, K.B. Cold Hardiness and Freeze Tolerance. Funct. Metab. 2005, 473–503. [Google Scholar] [CrossRef]
- Rathnayake, C.M.; Metwali, N.; Jayarathne, T.; Kettler, J.; Huang, Y.; Thorne, P.S.; O’Shaughnessy, P.T.; Stone, E.A. Influence of rain on the abundance of bioaerosols in fine and coarse particles. Atmos. Chem. Phys. Discuss. 2017, 17, 2459–2475. [Google Scholar] [CrossRef] [Green Version]
- Heo, K.J.; Kim, H.B.; Lee, B.U. Concentration of environmental fungal and bacterial bioaerosols during the monsoon season. J. Aerosol Sci. 2014, 77, 31–37. [Google Scholar] [CrossRef]
- Lugauer, M.; Baltensperger, U.; Furger, M.; Gäggeler, H.W.; Jost, D.T.; Schwikowski, M.; Wanner, H. Aerosol transport to the high Alpine sites Jungfraujoch (3454 m asl) and Colle Gnifetti (4452 m asl). Tellus B 1998, 50, 76–92. [Google Scholar] [CrossRef]
- Felgitsch, L.; Bichler, M.; Burkart, J.; Fiala, B.; Häusler, T.; Hitzenberger, R.; Grothe, H. Heterogeneous Freezing of Liquid Suspensions Including Juices and Extracts from Berries and Leaves from Perennial Plants. Atmosphere 2019, 10, 37. [Google Scholar] [CrossRef] [Green Version]
- DeMott, P.J.; Möhler, O.; Cziczo, D.J.; Hiranuma, N.; Petters, M.D.; Petters, S.S.; Belosi, F.; Bingemer, H.G.; Brooks, S.D.; Budke, C.; et al. The Fifth International Workshop on Ice Nucleation phase 2 (FIN-02): Laboratory intercomparison of ice nucleation measurements. Atmos. Meas. Tech. 2018, 11, 6231–6257. [Google Scholar] [CrossRef] [Green Version]
- Erickson, H.P. Size and shape of protein molecules at the nanometer level determined by sedimentation, gel filtration, and electron microscopy. Biol. Proced. Online 2009, 11, 32–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolber, P.K.; Deininger, C.A.; Southworth, M.W.; Vandekerckhove, J.; van Montagu, M.; Warren, G.J. Identification and purification of a bacterial ice-nucleation protein. Proc. Natl. Acad. Sci. USA 1986, 83, 7256–7260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, Y.; Hudait, A.; Molinero, V. How Size and Aggregation of Ice-Binding Proteins Control Their Ice Nucleation Efficiency. J. Am. Chem. Soc. 2019, 141, 7439–7452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zolles, T.; Burkart, J.; Häusler, T.; Pummer, B.; Hitzenberger, R.; Grothe, H. Identification of Ice Nucleation Active Sites on Feldspar Dust Particles. J. Phys. Chem. A 2014, 119, 2692–2700. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, D.; Adams, M.P.; Tarn, M.D.; Harrison, A.D.; Vergara-Temprado, J.; Porter, G.C.E.; Holden, M.A.; Sanchez-Marroquin, A.; Carotenuto, F.; Whale, T.F.; et al. Contributions of biogenic material to the atmospheric ice-nucleating particle population in North Western Europe. Sci. Rep. 2018, 8, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Conen, F.; Morris, C.E.; Leifeld, J.; Yakutin, M.V.; Alewell, C. Biological residues define the ice nucleation properties of soil dust. Atmos. Chem. Phys. Discuss. 2011, 11, 9643–9648. [Google Scholar] [CrossRef] [Green Version]
- Haga, D.I.; Iannone, R.; Mason, R.; Polishchuk, E.A.; Fetch, T.; Van Der Kamp, B.J.; McKendry, I.G.; Bertram, A.K.; Wheeler, M.J. Ice nucleation properties of rust and bunt fungal spores and their transport to high altitudes, where they can cause heterogeneous freezing. J. Geophys. Res. Atmos. 2013, 118, 7260–7272. [Google Scholar] [CrossRef]
- Wang, B.; Lambe, A.T.; Massoli, P.; Onasch, T.B.; Davidovits, P.; Worsnop, D.R.; Knopf, D.A. The deposition ice nucleation and immersion freezing potential of amorphous secondary organic aerosol: Pathways for ice and mixed-phase cloud formation. J. Geophys. Res. Space Phys. 2012, 117, 1–12. [Google Scholar] [CrossRef]
- Kanji, Z.A.; Abbatt, J.P.D. Ice Nucleation onto Arizona Test Dust at Cirrus Temperatures: Effect of Temperature and Aerosol Size on Onset Relative Humidity. J. Phys. Chem. A 2010, 114, 935–941. [Google Scholar] [CrossRef] [PubMed]
- Broadley, S.L.; Murray, B.; Herbert, R.J.; Atkinson, J.D.; Dobbie, S.; Malkin, T.L.; Condliffe, E.; Neve, L. Immersion mode heterogeneous ice nucleation by an illite rich powder representative of atmospheric mineral dust. Atmos. Chem. Phys. Discuss. 2012, 12, 287–307. [Google Scholar] [CrossRef] [Green Version]
- Pöhlker, C.; Huffman, J.A.; Pöschl, U. Autofluorescence of atmospheric bioaerosols—fluorescent biomolecules and potential interferences. Atmos. Meas. Tech. 2012, 5, 37–71. [Google Scholar] [CrossRef] [Green Version]
- Healy, D.A.; Huffman, J.A.; Oconnor, D.J.; Pöhlker, C.; Pöschl, U.; Sodeau, J.R. Ambient measurements of biological aerosol particles near Killarney, Ireland: A comparison between real-time fluorescence and microscopy techniques. Atmos. Chem. Phys. Discuss. 2014, 14, 8055–8069. [Google Scholar] [CrossRef] [Green Version]
- Haga, D.I.; Burrows, S.M.; Iannone, R.; Wheeler, M.J.; Mason, R.H.; Chen, J.; Bertram, A.K. Ice nucleation by fungal spores from the classes Agaricomycetes, Ustilaginomycetes, and Eurotiomycetes and the effect on the atmospheric transport of these spores. Atmos. Chem. Phys. 2014, 14, 8611–8630. [Google Scholar] [CrossRef] [Green Version]
- Chang, K.-F.; Blenis, P.V. Survival of Endocronartium harknessii teliospores in a simulated airborne state. Can. J. Bot. 1989, 67, 928–932. [Google Scholar] [CrossRef]
- Lindow, S.E.; Arny, D.C.; Upper, C.D. Bacterial Ice Nucleation: A Factor in Frost Injury to Plants. Plant Physiol. 1982, 70, 1084–1089. [Google Scholar] [CrossRef] [PubMed]
- Morris, C.E.; Georgakopoulos, D.; Sands, D.C. Ice nucleation active bacteria and their potential role in precipitation. Le J. de Phys. Colloq. 2004, 121, 87–103. [Google Scholar] [CrossRef]
- Huffman, J.A.; Sinha, B.; Garland, R.M.; Snee-Pollmann, A.; Gunthe, S.S.; Artaxo, P.; Martin, S.T.; Andreae, M.O.; Pöschl, U. Size distributions and temporal variations of biological aerosol particles in the Amazon rainforest characterized by microscopy and real-time UV-APS fluorescence techniques during AMAZE-08. Atmos. Chem. Phys. Discuss. 2012, 12, 11997–12019. [Google Scholar] [CrossRef] [Green Version]
- Bogler, S.; Borduas-Dedekind, N. Lignin’s ability to nucleate ice via immersion freezing and its stability towards physicochemical treatments and atmospheric processing. Atmos. Chem. Phys. 2020, 20, 14509–14522. [Google Scholar] [CrossRef]
- Dreischmeier, K.; Budke, C.; Wiehemeier, L.; Kottke, T.; Koop, T. Boreal pollen contain ice-nucleating as well as ice-binding ‘antifreeze’ polysaccharides. Sci. Rep. 2017, 7, srep41890. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample ID (Day-Device-Zone) | Date | Zone | Altitude (m) | Start Time (CEST) | Duration (min) | Flow Rate (L min−1) | Air Volume (L) |
|---|---|---|---|---|---|---|---|
| UA3-Imp-Z0 | 21. 08. 2019 | 0 | 1 | 18:50 | 10 | 1.00 ± 0.03 | 10.0 |
| UA3-Pac-Z0 | 21. 08. 2019 | 0 | 1 | 18:50 | 10 | 8.92 ± 0.04 | 89.2 |
| UA3-Imp-Z0 | 21. 08. 2019 | 0 | 1 | 19:23 | 50 | 1.00 ± 0.03 | 50.0 |
| UA3-Pac-Z0 | 21. 08. 2019 | 0 | 1 | 19:23 | 50 | 8.92 ± 0.04 | 446 |
| UA4-Imp-Z0 | 22. 08. 2019 | 0 | 1 | 20:01 | 50 | 1.07 ± 0.04 | 53.5 |
| UA4-Pac-Z0 | 22. 08. 2019 | 0 | 1 | 20:01 | 50 | 8.94 ± 0.06 | 447 |
| UA4-Imp-Z1 | 22. 08. 2019 | 1 | 5 | 18:48 | 10 | 1.07 ± 0.04 | 10.7 |
| UA4-Imp-Z2 | 22. 08. 2019 | 2 | 5 | 18:28 | 10 | 1.07 ± 0.04 | 10.7 |
| UA4-Imp-Z3 | 22. 08. 2019 | 3 | 45 | 19:25 | 10 | 1.07 ± 0.04 | 10.7 |
| UA5-Imp-Z0 | 23. 08. 2019 | 0 | 1 | 19:54 | 10 | 1.08 ± 0.02 | 10.8 |
| UA5-Pac-Z0 | 23. 08. 2019 | 0 | 1 | 19:15 | 50 | 9.08 ± 0.06 | 454 |
| UA5-Imp-Z1 | 23. 08. 2019 | 1 | 5 | 18:55 | 10 | 1.08 ± 0.02 | 10.8 |
| UA5-Imp-Z2 | 23. 08. 2019 | 2 | 5 | 19:10 | 10 | 1.08 ± 0.02 | 10.8 |
| UA5-Imp-Z3 | 23. 08. 2019 | 3 | 45 | 19:25 | 10 | 1.08 ± 0.02 | 10.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seifried, T.M.; Bieber, P.; Kunert, A.T.; Schmale, D.G., III; Whitmore, K.; Fröhlich-Nowoisky, J.; Grothe, H. Ice Nucleation Activity of Alpine Bioaerosol Emitted in Vicinity of a Birch Forest. Atmosphere 2021, 12, 779. https://doi.org/10.3390/atmos12060779
Seifried TM, Bieber P, Kunert AT, Schmale DG III, Whitmore K, Fröhlich-Nowoisky J, Grothe H. Ice Nucleation Activity of Alpine Bioaerosol Emitted in Vicinity of a Birch Forest. Atmosphere. 2021; 12(6):779. https://doi.org/10.3390/atmos12060779
Chicago/Turabian StyleSeifried, Teresa M., Paul Bieber, Anna T. Kunert, David G. Schmale, III, Karin Whitmore, Janine Fröhlich-Nowoisky, and Hinrich Grothe. 2021. "Ice Nucleation Activity of Alpine Bioaerosol Emitted in Vicinity of a Birch Forest" Atmosphere 12, no. 6: 779. https://doi.org/10.3390/atmos12060779