
Phytotoxic Activity and Growth Inhibitory Substances from Albizia richardiana (Voigt.) King & Prain



1
Department of Applied Biological Science, Faculty of Agriculture, Kagawa University, Miki 761-0795, Japan
2
The United Graduate School of Agricultural Sciences, Ehime University, Matsuyama 790-8566, Japan
3
Department of Chemistry, Faculty of Science and Technology, Keio University, Kohoku, Yokohama 223-8522, Japan
*
Author to whom correspondence should be addressed.
Appl. Sci. 2021, 11(4), 1455; https://doi.org/10.3390/app11041455
Submission received: 24 December 2020
/
Revised: 18 January 2021
/
Accepted: 1 February 2021
/
Published: 5 February 2021
(This article belongs to the Special Issue Phytotoxic Substances: Characterization, Activity, and Application)
Abstract
:Albizia richardiana, a fast-growing, large deciduous tree belonging to the Fabaceae family, grows well in hot and humid areas but mainly grows in the tropics of the Old World. The medicinal and other uses of Albizia richardiana are well documented, but the phytotoxic effects of this tree have not yet been investigated. We conducted this study to investigate the phytotoxic activity of Albizia richardiana leaves and to identify growth inhibitory substances for controlling weeds in a sustainable way. Aqueous methanol extracts of Albizia richardiana leaves greatly suppressed the growth of cress and barnyard grass seedlings in a concentration- and species-dependent manner. Two phytotoxic substances were separated using several purification steps and characterized through spectral analysis as dehydrovomifoliol and loliolide. Dehydrovomifoliol and loliolide significantly arrested the seedling growth of cress in the concentrations of 0.1 and 0.01 mM, respectively. The extract concentrations needed for 50% growth inhibition (I50 values) of cress seedlings were 3.16–3.01 mM for dehydrovomifoliol and 0.03–0.02 mM for loliolide. The results suggest that these two allelopathic substances might play a vital role in the phytotoxicity of Albizia richardiana leaves.
1. Introduction
Sustainable agriculture is described as a method that improves the quality of the environment and the resources on which the agriculture sector relies, that supplies the basic human needs for food and fibers, and that improves the living standard of farmers and society in general [1,2]. It implies the reliable maintenance of agricultural practices for assessing and implementing sustainable crop cultivation. Protecting different field crops against weeds, other crop pests, and diseases is the key factor for the sustainable production of crops [3]. Weeds directly contend with crops for space, light, nutrition, and humidity. Therefore, the intrusion of weeds greatly affects the physiological behavior and development of crop plants [4,5]. Weeds cause significant crop yield losses—more than 50 percent for some crops if the fields are left unchecked [6]. The strategy for weed control performs a critical role and directly affects global food protection and food productivity.
Herbicides developed since the end of the Second World War have been used as the key tool for weed control. At present, the application of herbicides in agriculture creates problems for people and crops and results in environmental degradation and the growth of herbicide-resistant weeds [7]. Consequently, health hazards ensue, and sources of drinking water could become polluted, leading to adverse effects on plants, microorganisms, birds, and fish [8]. It is essential to find an alternative method to develop synthetic herbicides, which might be used for integrated weed control programs to grow crops sustainably. Allelopathy is the biological phenomenon in which plants or plant residues release allelochemicals that affect (usually adversely) other nearby plants [9] and is the best alternative for sustainably controlling weeds in crop fields. The first reason for research on allelopathy is to gain better insight into how plants interact with other life forms via allelochemicals/secondary metabolites. Allelopathy helps to reduce the use of synthetic herbicides, subsequently reducing environmental deterioration, and helps to develop successful methods for sustainable agricultural production [10].
In agro-ecosystems, allelopathy has a significant role not only in plant-to-plant, and plant-to-microorganisms interactions, but also in plant-to-insect or plant-to-herbivore contact [11]. Plants produce various types of allelochemicals, such as phenols and tannins [12]; saponins and alkaloids [13]; flavonoids and terpenoids [14]; amino acids, carbohydrates, and glycosides [15]; and coumaric acid [16]. These substances have herbicidal characteristics such as being able to restrict cell division [17] and reduce the rate of photosynthesis of nearby plants [18]. The effects of phytochemicals on seed germination, seedling growth, and plant development are controlled by the interaction, complexity, and concentration of plants’ secondary metabolites [19]. Studies have investigated the use of allelopathic plants and allelochemicals in agriculture for sustainable weed control [20,21,22]. Presently, phytochemicals are being investigated in the development of herbicides [23].
Albizia richardiana (Voigt.) King & Prain (local (Bangladesh) name: rajkoroi) is a large, fast-growing deciduous tree belonging to the family Fabaceae, sub-family Mimosaceae [24]. The leaves of Albizia richardiana are bipinnate, compound, sessile, and small. The flower is stalkless, small, and greenish white. The fruit is thin, long, and whitish brown. This tree species usually grows well in hot and humid areas in Asia, Australia, Madagascar, North America, and Africa, but mainly grows in the tropics of the Old World [25,26]. The tree can be found in different areas of Bangladesh like Sunamgonj, Chittagong, Barisal, Bagerhat, Jhalukati, Madaripur, and Pirujpur. It is also cultivated as an ornamental and roadside avenue tree [27] and is considered an auspicious tree in Bangladesh [28].
Albizia richardiana is an important part of villages and social forests in Bangladesh [29] where it is used for furniture, frame manufacturing, house posts, roofing, plywood, etc. [30,31]. Different parts of this tree, such as the roots, bark, fruit, and flowers, are used medicinally to treat appetite loss, tightness in the chest, depression, eye problems, back pain, and blurred vision, and to increase blood circulation [32]. This species contains many compounds in its bark including glycosides, carbohydrates, alkaloids, saponins, and glucosides. The antimicrobial activity and hypoglycemic and anti-inflammatory effects have also been evaluated [33,34]. However, despite reports in the literature of Albizia richardiana’s medicinal and other uses, there is no report on its phytotoxicity. This research was undertaken to explore the phytotoxic potential of Albizia richardiana and to identify its active phytotoxic substances.
2. Materials and Methods
2.1. Plant Materials
Albizia richardiana is a fast-growing tree mostly found in hot and humid climatic zones. The leaves of this plant were collected from the Bangladesh Agricultural University (BAU), Mymensingh, Bangladesh, in July and August 2019. The Albizia richardiana plant identification was confirmed by Sarwar Abul Khayer Mohammad Golam (Department of Crop Botany, BAU, Mymensingh, Bangladesh). For future reference, a voucher (voucher number HOTBAU 19OP-0001) for this tree species has been deposited with the Ornamental Plant Herbarium, Department of Horticulture, BAU. The collected leaves were washed under tap water to remove dust or other debris. The leaves were desiccated in a shady place to avoid direct sunlight, and desiccated leaves were ground to powder through a grinder. Finally, the leaf powders were stored in a plastic bag in a refrigerator at 2 °C until the extraction.
2.2. Test Plant Species
In this study, two test plant species, cress (Lepidium sativum L.), a dicot plant, and barnyard grass (Echinochloa crus-galli (L.) P. Beauv.), a monocot plant species, were used for a bioassay. Cress is a crop species and barnyard grass is a weed species. Cress has been tested in various laboratory conditions for its noted seedling growth characteristics and has shown susceptibility to allelopathic substances [35]. Barnyard grass is a common and noxious weed in crop fields.
2.3. Extract Preparation
Aqueous methanol (70% (v/v), 500 mL) was used for extracting 100 g of Albizia richardiana leaf powder for 48 h. The leaf extract was filtrated using a layer of filter paper (No. 2, 125 mm; Advantec, Toyo Roshi Kaisha Ltd., Tokyo, Japan). An equal quantity of cold methanol (500 mL) was utilized to extract the leaf residue for 24 h and resulting leaf residues were filtrated again. The extracted residues were mixed and evaporated (40 °C) to complete dryness through a rotary evaporator.
2.4. Growth Bioassay Experiments
The subsequent extract residues of Albizia richardiana (obtained from 100 g leaf powder) were dissolved in 250 mL of cold methanol to produce six bioassay concentrations, 0.003, 0.01, 0.03, 0.1, 0.3, and 1.0 g dry weight (DW) equivalent extract/mL. To check the growth suppression activity of the leaf extracts, an aliquot of extract residue was applied on one layer of filter paper (No. 2, 28 mm; Toyo Roshi Ltd., Tokyo, Japan) in the Petri dishes (28 mm) at different bioassay concentrations and stored in the fume chamber to remove methanol from the leaf extract. In each of the Petri dishes, the filter paper was soaked with 0.6 mL of an aqueous solution of Tween 20 (polyoxyethylene sorbitan monolaurate; Nacalai Tesque, Inc., Kyoto, Japan). The Tween 20 solution was applied as surfactant and had no harmful effect on the pre-germinated seeds or seeds of the test plants. After that, the cress (10 seeds) and the barnyard grass (10 pre-emergence seedlings) were placed in each Petri dish onto a sheet of filter paper (No. 2, 28 mm; Toyo Roshi Ltd., Tokyo, Japan). Barnyard grass seeds were pre-sprouted by moistening in water and the seeds were kept for 48 h at 25 °C in a growth chamber. The control treatment without extract residues was prepared as described above. Six replications were performed for each concentration using a completely randomized block design (CRBD). Seedling lengths (shoots and roots) were determined after two days of incubation in a growth incubator (25 °C under a dark condition), and the percentage of seedling growth was measured with reference to control seedling growth.
2.5. Isolation and Purification of the Growth Inhibitory Substances
The powdered Albizia richardiana leaves (3000 g) were (dissolved in 14 L of 70% aqueous methanol and 14 L of methanol) extracted as mentioned in the extract preparation and were concentrated (at 40 °C) using a rotary evaporator to make aqueous residues. The concentrated residues were then adjusted to a pH of 7.0 with 1 molar of phosphate buffer solution. Partitioning was done with the same amount of the ethyl acetate (seven times, 150 mL/time) and separated into aqueous fraction and ethyl acetate fraction. The growth suppression effects of the aqueous fraction and the ethyl acetate fraction were measured with a cress assay as mentioned previously. The ethyl acetate fraction displayed the highest activity, so this fraction was then evaporated until complete dryness after using anhydrous sodium sulphate (Na2SO4) to remove water for overnight. Firstly, the fraction of ethyl acetate was separated through a column of silica gel (60 g of silica gel 60, spherical, 70–230 mesh; Nacalai Tesque, Inc.) and eluted stepwise with n-hexane (150 mL/step), holding increasing quantities of ethyl acetate (10%/step, v/v) from 20% to 80%, ethyl acetate (150 mL), and two times cold methanol (300 mL) to produce 9 fractions. The biological effects of these nine fractions were measured using a cress assay. From the assay results, it was observed that fraction 8 (80% ethyl acetate in n-hexane) showed the highest biological activity. The residues were evaporated to dryness and separated through a column of Sephadex LH-20 (GE Healthcare, 50 g; Bio-Sciences AB, SE-751 84 Uppsala, Sweden). The Sephadex column was loaded with different percentages (fractions 1 to 5) of aqueous methanol (e.g., 20, 40, 60, and 80% (v/v)) (150 mL/step), and two times methanol (300 mL). After collection, these fractions were evaporated until dry and, to check the biological effects of these fractions, a cress bioassay was set. Assay chromatography showed that the 40% aqueous methanol fraction (fraction 2) was the most active. The residues were evaporated until dry, diluted with aqueous methanol 20% (v/v), and loaded onto a reverse-phase C18 cartridge (YMC Co. Ltd., Kyoto, Japan). The C18 cartridge was eluted with 20%, 30%, 40%, 50%, 60%, and 80% (v/v) aqueous methanol (15 mL per step) and two times methanol (30 mL per step). The aqueous methanol of 30% (fraction 2) showed the most activity against cress. The highest active fraction was purified using reverse-phase high-performance liquid chromatography (HPLC, 500 mm × 10 mm I.D. ODS AQ-325; YMC Co. Ltd.) at a flow rate of 1.5 mL/min with 40% aqueous methanol and was detected at a wavelength of 220 nm and an oven temperature of 40 °C. Two active peaks were found at retention times 65–70 min (compound 1) and 76–80 min (compound 2). These two active peak fractions were checked again using reverse-phase HPLC (5 μm, 4.6 mm × 250 mm I.D., Inertsil® ODS-3; GL Science Inc., Tokyo, Japan). The column was eluted at a flow rate of 0.8 mL/min with 20% (v/v) aqueous methanol for compound 1 and 25% (v/v) aqueous methanol for compound 2. The active peaks of these two compounds were detected at 220 nm, 40 °C, and at a retention time of 68–74 min as a colorless substance (compound 1) and at a retention time of 56–63 min as a whitish substance (compound 2). These two active compounds were then characterized through HRESIMS. ESIMS spectra were obtained on an LCT Premier XE time-of-flight (TOF) mass spectrometer using positive ion mode; 1H-NMR, NMR spectral data were recorded on a JEOL JNM-ECX400 spectrometer at 400 MHz. 1H NMR chemical shifts were referenced to residual CHD2OD observed at δH 3.31 and a specific rotation.
2.6. Growth Bioassay of the Isolated Substances
The isolated compounds were dissolved in cold methanol to create different bioassay concentrations of 0.003, 0.01, 0.03, 0.1, 0.3, 1.0, 3.0, and 4.0 mM for compound 1 and 0.001, 0.003, 0.01, 0.03, 0.1, 0.3, 1.0, and 1.5 mM for compound 2, added onto a one-layer sheet of filter paper (No. 2, 28 mm; Toyo Roshi Ltd., Tokyo, Japan) in the Petri dishes (28 mm) and stowed in a fume chamber to evaporate the solvent. The growth inhibitory effects of the isolated substances were measured using the cress assay as previously mentioned.
2.7. Statistical Analysis
All of the assay experiments were carried out following a CRBD with the three replications and were repeated two times. The mean values are stated as mean ± SE. The ANOVA (analysis of variance) was measured using SPSS software, version 16.0 for Windows (SPSS Inc., Chicago, IL, USA). The significant variations between the control and treatments were tested using Tukey’s HSD (Honestly Significant Difference) at a 0.05 probability level. The concentrations needed for 50% inhibition of the growth of the tested plants (I50 values) in the bioassay were measured using a logistic regression equation of the concentration–response curves.
3. Results
3.1. Allelopathic Effects of the Albizia richardiana Extracts
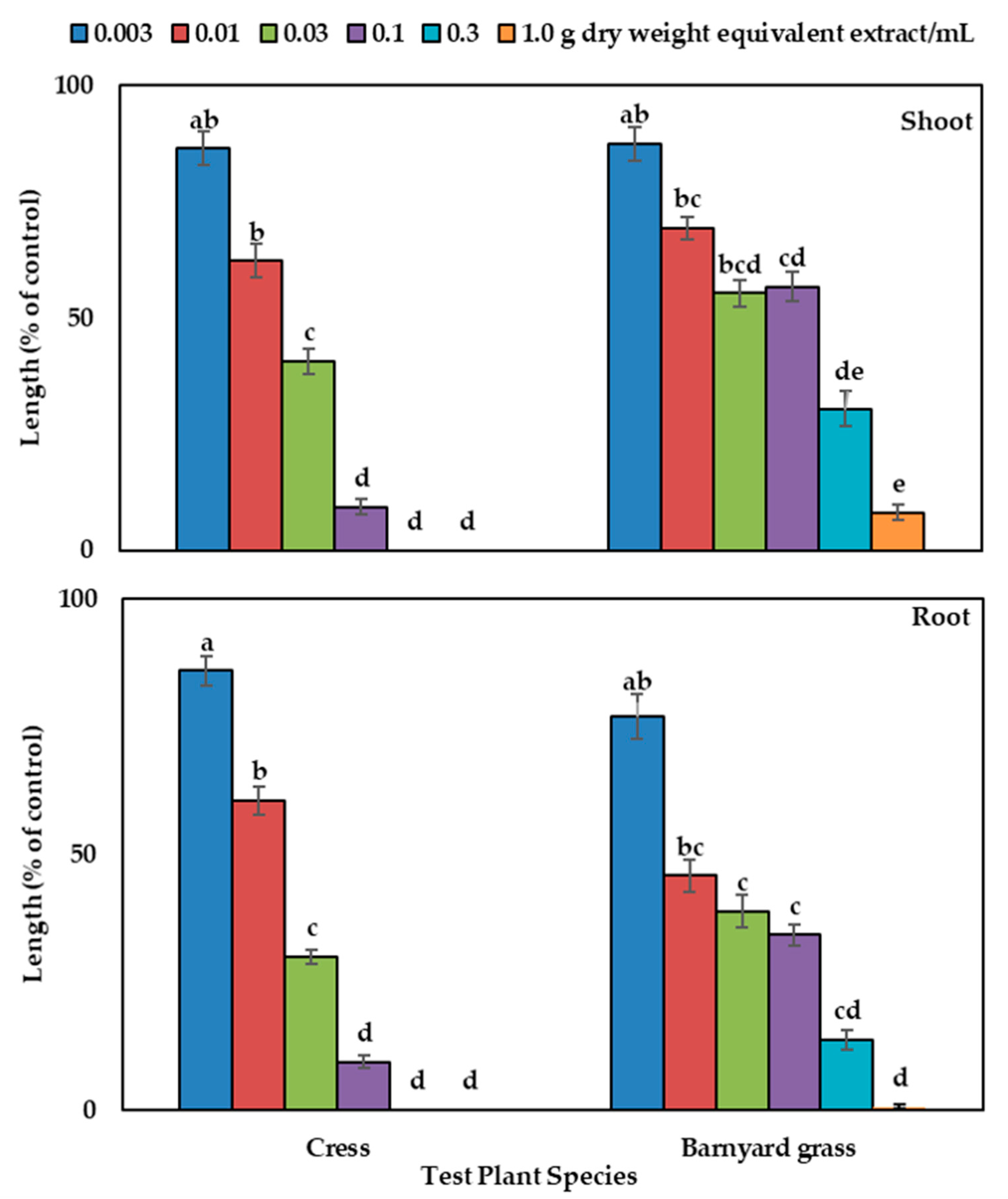
The aqueous methanol leaf extract of Albizia richardiana significantly restricted the seedling growth of the tested plants (Figure 1) the concentrations higher than the 0.003 g DW equivalent extract/mL. Cress seedlings were completely arrested at the concentration of 0.3 and 1.0 g DW equivalent extract/mL, whereas the barnyard grass seedlings shoot was restricted to 30.5% and 8.1% of the control seedling growth and the roots to 13.7% and 0.64%. Different levels of suppression of tested plants were also found with plant extracts when treated with different concentrations (Figure 1). The concentration needed for 50% suppression (I50 values) of growth of the tested species varied between 0.019 and 0.049 g DW equivalent extract/mL for the shoots and between 0.008 and 0.015 for the roots (Table 1), showing that root growth was more sensitive to the extracts than the shoots of all the tested plants.
3.2. Determination of the Structures of the Allelopathic Substances
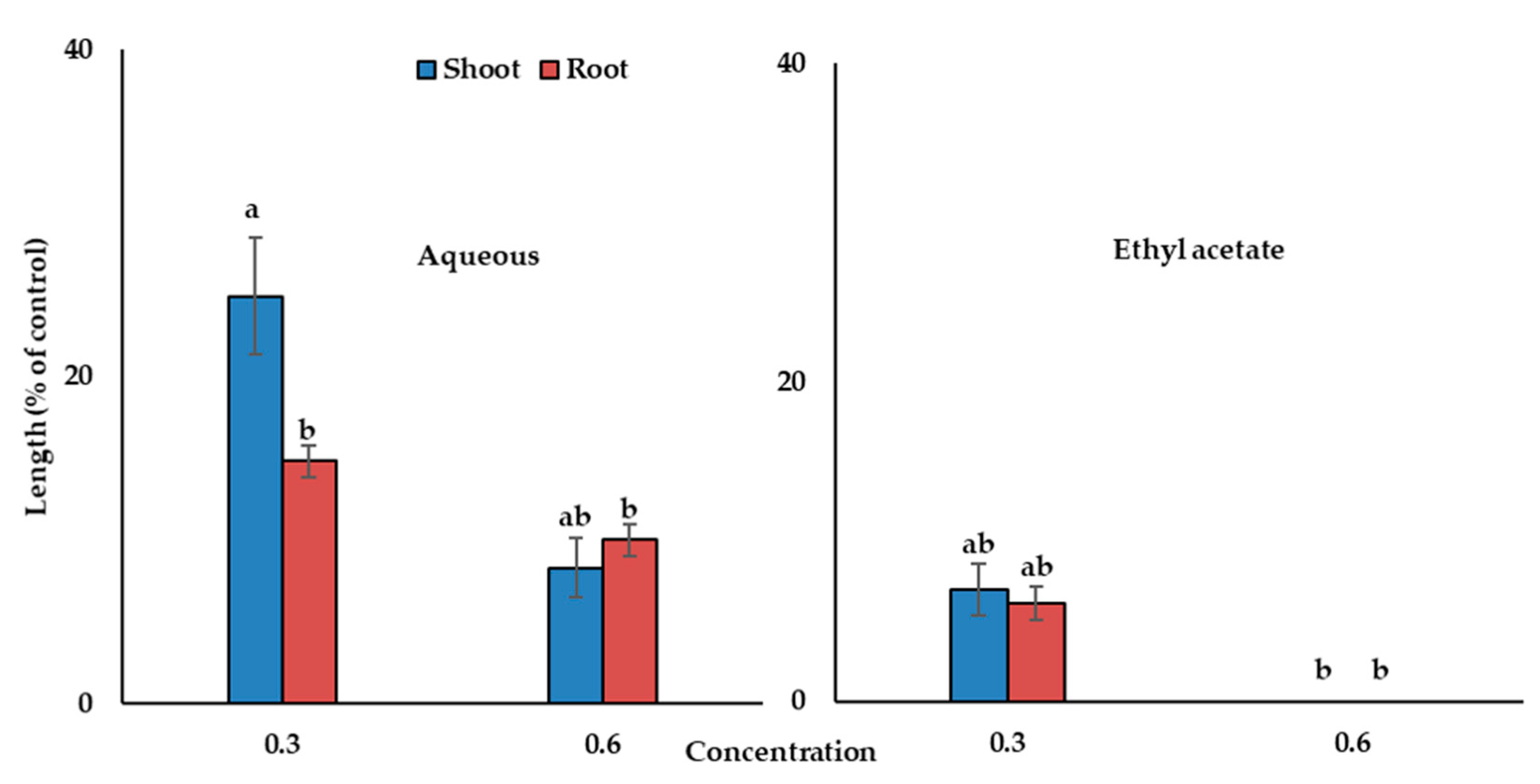
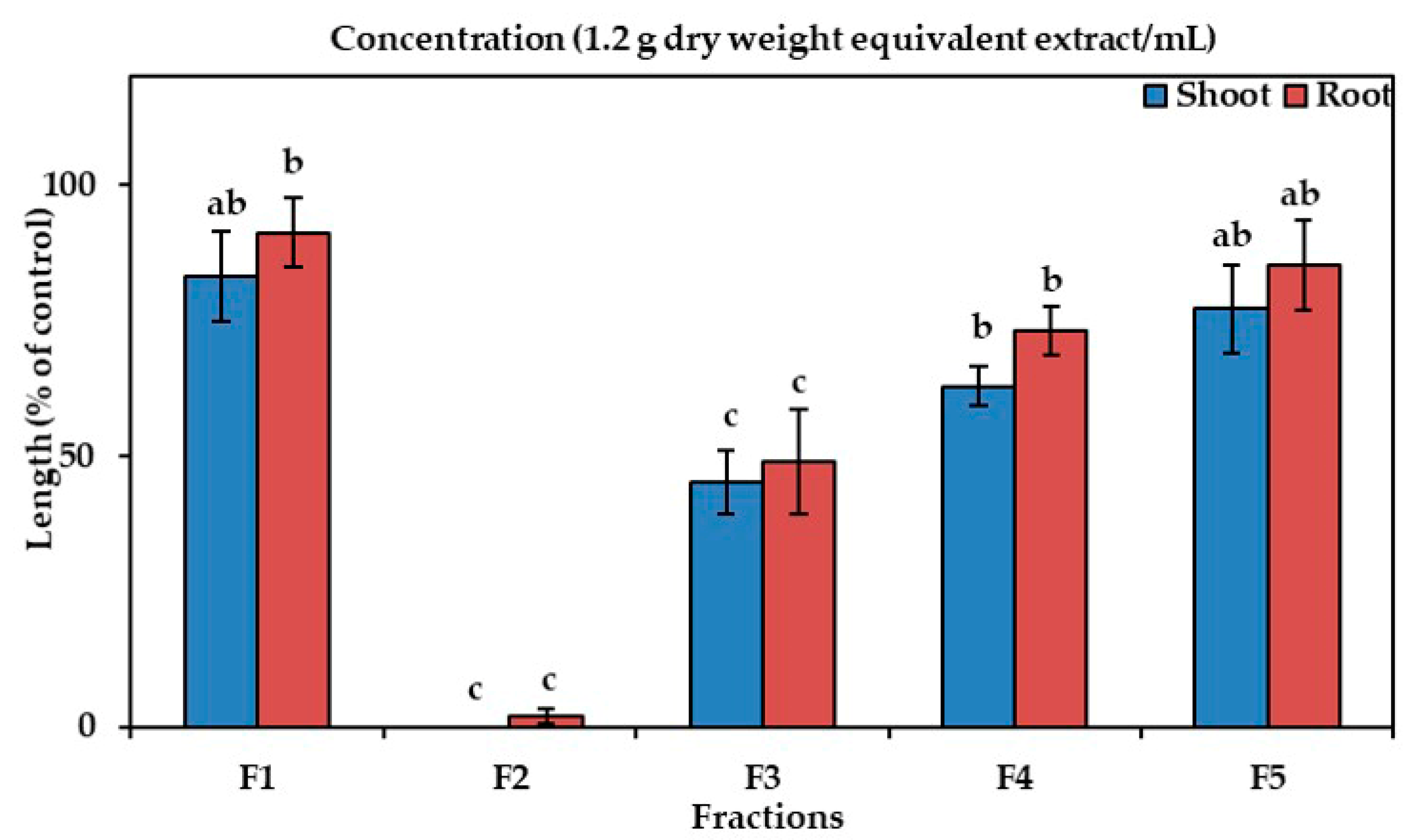
The aqueous fraction and the ethyl acetate fraction of Albizia richardiana extracts arrested the growth of the cress seedlings in a concentration-dependent manner (Figure 2). In the concentration 0.6 g DW equivalent extract/mL, the aqueous portion restricted shoot growth to 8.3% of the control and the growth of the roots to 10%, whereas the ethyl acetate portion completely suppressed the growth of the cress seedlings compared with the control. Hence, the ethyl acetate fraction was selected for the subsequent purification steps using a silica gel column, a Sephadex LH-20 column (in the Sephadex LH-20 purification step, fraction 2 (F2) showed the highest inhibitory activity, which is displayed in Figure 3), and reverse-phase C18 cartridges to check the allelopathic effects of all the fractions. Two active phytotoxic substances were isolated by using reverse-phase HPLC, and these substances were identified by comparing with earlier reported spectral data.
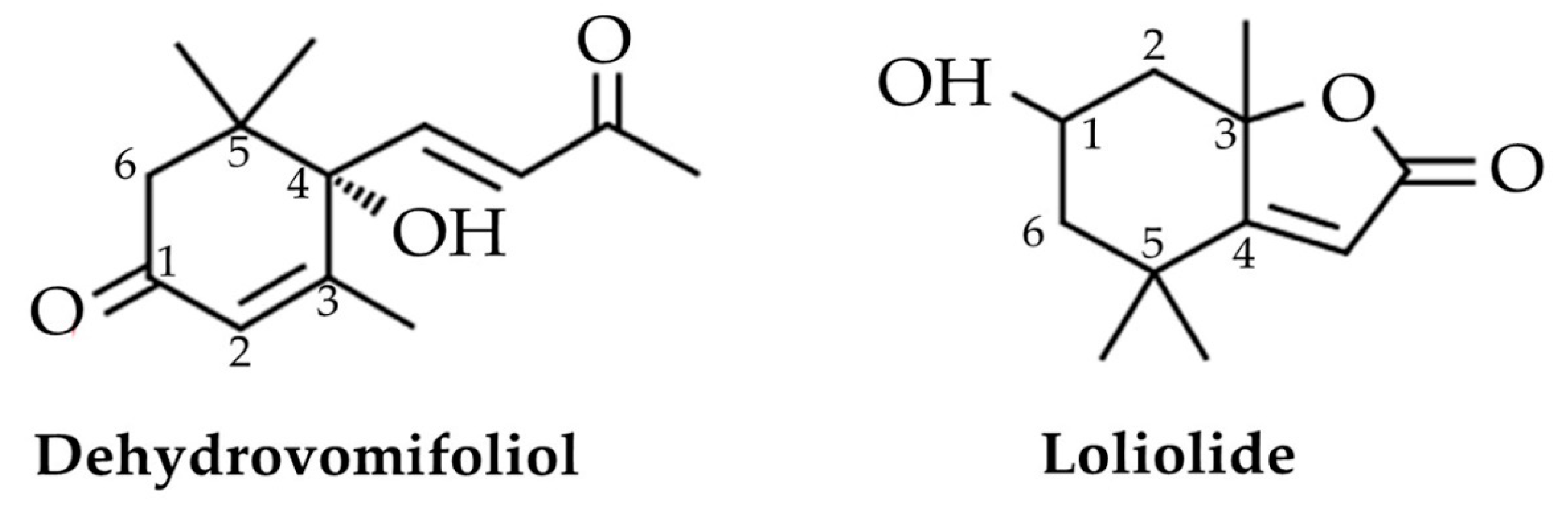
The molecular formula of compound 1 was measured as C13H19O3 through ESIMS at m/z 223.1407 [M+H]+ (calcd. for C13H19O3, 223.1334). The 1H NMR (400 MHz, CD3OD) spectrum of this compound was δH 6.99 (d, J = 16.2 Hz, 1 H, H7), 6.43 (d, J = 16.2 Hz, 1 H, H8), 5.93 (brs, 1 H, H4), 2.59 (d, J = 17.8 Hz, 1 H, H1), 2.30 (s, 3 H, H10), 2.28 (d, J = 17.8 Hz, 1 H, H2), 1.90 (d, J = 1.4 Hz, 3 H, H13), 1.06 (s, 3H, H11), 1.02 (s, 3 H, H12). The distinct rotation of this substance was [α]D32 = +50 (c = 0.14, CH3OH). Comparing this spectral data with earlier recorded data led to identifying this compound as dehydrovomifoliol (Figure 4) [36,37].
The molecular formula of compound 2 was measured as C11H17O3 through HRESIMS at m/z 197.1193 [M+H]+ (calcd. for C11H17O3, 197.1178). The 1H NMR (400 MHz, CD3OD) spectrum of this compound was δH 5.75 (s, 1 H, H7), 4.22 (m, 1 H, H3), 2.42 (ddd, J = 14.2, 2.8, 2.8 Hz, 1 H, H4b), 1.99 (ddd, J = 14.6, 3.2, 2.6 Hz, 1 H, H2b), 1.76 (s, 3 H, H11), 1.75 (m, 1 H, H4a), 1.53 (dd, J = 14.6, 3.8, Hz, 1 H, H2a), 1.47 (s, 3 H, H9), 1.28 (s, 3 H, H10). The distinct rotation of this compound was [α]D26 = −101 (c = 0.07, CHCl3). Comparing this spectral data with earlier recorded data led to identifying this compound as loliolide (Figure 4) [38,39,40].
3.3. The Biological Effects of the Isolated Compounds
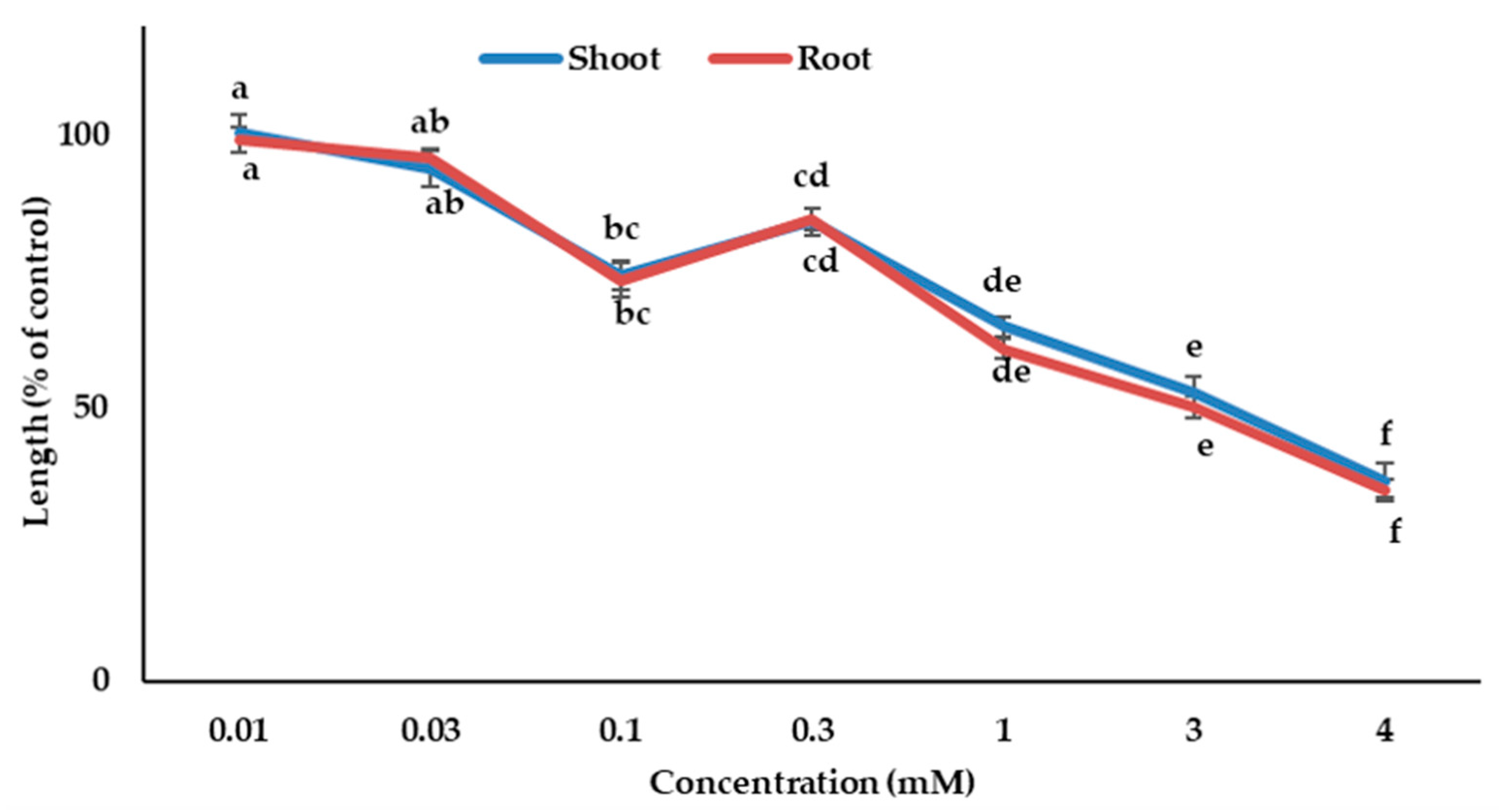
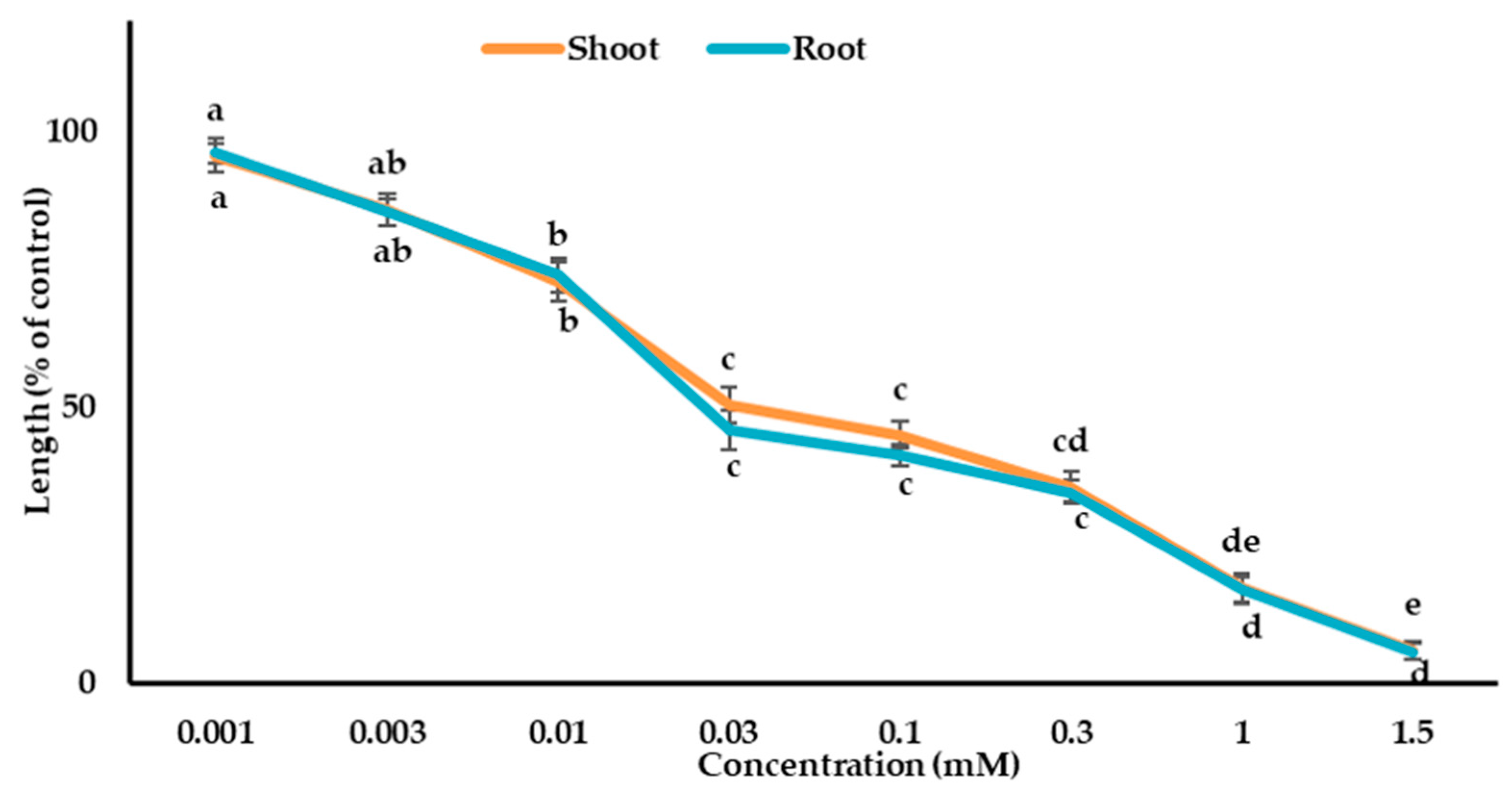
The biological effects of dehydrovomifoliol and loliolide on cress were evaluated. The results from the bioassay showed that cress seedling growth was significantly suppressed by both substances (Figure 5 and Figure 6). The inhibition level of the substances was raised by raising the concentration, indicating that suppression was dose-dependent. Dehydrovomifoliol and loliolide significantly inhibited the cress seedling growth in the concentrations of 0.1 and 0.01 mM, respectively (Figure 5 and Figure 6).
Dehydrovomifoliol showed the highest inhibition of 36.7% and 34.9% for the shoot and root growth of cress, respectively, at a concentration of 4.0 mM, and loliolide showed the highest inhibition of 5.9% and 5.8% for the shoot and root growth of cress, respectively, at a concentration of 1.5 mM, compared with the control seedling growth. The I50 values for dehydrovomifoliol against the cress were 3.16 mM for the shoots and 3.01 mM for the roots, and for loliolide the I50 values were 0.03 and 0.02 mM, respectively (Table 2). Accordingly, loliolide had higher growth suppressing potential against cress compared with dehydrovomifoliol. Moreover, the roots showed higher sensitivity to the compounds than the shoots.
4. Discussion
The aqueous methanol leaf extract of Albizia richardiana significantly suppressed the shoot and root growth of the tested cress and barnyard grass plants (Figure 2). The increasing suppression of the growth of seedlings of the tested species was related to the increased extract concentration. The I50 values of the tested species were different, indicating that the inhibitory effects were test-plant specific (Table 1). The concentration-dependent and test-species-dependent inhibitory effects of the leaf extract of Albizia richardiana indicate that this leaf extract might possess phytotoxic compounds, and this inhibitory activity has been reported in various studies [40,41,42,43].
Allelopathic substances can influence physiological activities and factors such as seed germination, photosynthesis, respiration, transpiration, ion uptake, water status, stomatal opening, enzyme activity, and the hormone levels of plants [44]. These phytotoxic substances also influence cell division and differentiation, gene expression, signal transduction, cell wall structure, cell membranes, and cell permeability [45,46]. The results of this study show that the shoots of the test species were less suspectable to the Albizia richardiana extracts than the roots. Plant roots play a vital role in the adaptation of the plant to edaphic restrictions, as well as biotic and abiotic factors [47]. The elongation of shoots and roots is usually used to measure allelopathic potential [48]. However, researchers also report that growth suppression activity of plant extracts are more active against roots than shoots [49,50]. Roots show greater sensitivity to plant extracts because of direct interaction with allelopathic substances [51] and because root tissues are more highly permeable to phytochemicals than shoot tissues [52,53]. In addition, root growth depends on the proliferation of cells, which is greatly influenced by allelochemicals, leading to the inhibition of root growth [54]. The plant extracts for this experiment were subjected to different purification steps, and the two allelopathic compounds were identified and characterized through spectral analysis as dehydrovomifoliol and loliolide. Both phytotoxic compounds have been reported as carotenoid metabolites [55].
Dehydrovomifoliol is reported in different plants such as Phaseolus vulgaris L. [56], rice husks [35], Beta vulgaris var. [57], Cucumis sativa [36], Arctium lappa [58], and Nitraria sibirica Pall. [59]; it has also been obtained by synthesizing C9-hydroxy ketone [60]. In addition, dehydrovomifoliol has been shown to have cytotoxic effects against human cancer cells [61].
In 1964, Hodges and Porte [37] first reported on loliolide in the plant Lolium perenne, and thereafter it was found in both land and sea ecosystems in different plant species and animal species [62], such as Helianthus tuberosus [55], Heliotropium angiospermum [63], Digitaria sanguinalis [64], Eichhornia crassipes [65], Sargassum horneri [66], and Oryza sativa L. ssp. indica (indigenous rice variety “Goria”) [67]; it has also been obtained by synthesizing C11-aldehyde [60]. Loliolide has various pharmaceutical functions [62]. It has been applied for its antioxidant [68], anticancer [58], antiviral, anti-melanogenic, anti-inflammatory, anti-aging [69], antituberculosis [70], antidiabetic [71], antibacterial, antifungal [64], and cell senescence inhibition activities [58]. Previously, the compounds dehydrovomifoliol and loliolide have also been identified from the plants Rollinia emarginata [72], Paspalum commersonii [73], and Vitex leptobotrys [74]. However, there are no reports found in the literature suggesting that dehydrovomifoliol and loliolide have been isolated from Albizia richardiana leaves.
The I50 values indicate that inhibitory effect of loliolide against cress shoot and root growth was more potent than that of dehydrovomifoliol. The difference in allelopathic activity may be due to the disparity in the chemical structures of the substances, because the allelopathic potential of phytotoxic substances is measured based on structural differences [75]. Dehydrovomifoliol contains 13 carbon atoms, in which cyclohexanone (2-cyclohexen-1-one) is substituted at positions 3, 5, and 5 by methyl groups, and by both a hydroxy group and a 3-oxobut-1-en-1-yl group at position 4. Alternatively, loliolide has 11 carbon atoms, in which 1,3-dihydroxy-3,5,5-trimethyl-substituted cyclohexylidene is connected to acetic acid lactone at position 4. Additionally, in dehydrovomifoliol, an OH group is situated at the C-4 position, whereas in loliolide, an OH group is situated at the C-1 and C-3 positions. Kobayashi [76] suggested that the OH group at the C-3 position of loliolide is responsible for its phytotoxic effects. For these reasons, the cress seedlings might exhibit greater sensitivity to loliolide than dehydrovomifoliol.
Therefore, the inhibitory activity of dehydrovomifoliol and loliolide indicate the allelopathic potential of Albizia richardiana. Accordingly, the allelopathic activity of Albizia richardiana could lead to this tree being used for the development of bioherbicides to increase sustainable agricultural production.
5. Conclusions
Extracts of the leaves of Albizia richardiana restricted the seedling growth of the test species, and the level of suppression depended on the concentration of the extracts and the test plant species. Two phytotoxic substances were obtained from the Albizia richardiana leaves through various purification steps and were characterized as dehydrovomifoliol and loliolide using spectral data. Both of the substances suppressed the seedling growth of the cress, which was subjected to different concentrations. The results from this study suggest that dehydrovomifoliol and loliolide have phytotoxic potential and that they contribute to the phytotoxic effects of the Albizia richardiana leaves. Therefore, Albizia richardiana might be a potent candidate for the biological control of weeds.
Author Contributions
Conceptualization, K.H. and H.K.-N.; methodology, K.H., K.S., A.I., and H.K.-N.; software, K.H.; validation, K.S., A.I., and H.K.-N.; formal analysis, K.H.; investigation, K.H.; resources, H.K.-N.; data curation, H.K.-N.; writing—original draft preparation, K.H.; writing—review and editing, H.K.-N.; visualization, K.H.; supervision, H.K.-N. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported through a MEXT scholarship (Grant Number MEXT-193490) from the Japanese government to conduct the study in Japan.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Acknowledgments
We are grateful to Dennis Murphy, the United Graduate School of Agricultural Sciences (UGAS), Ehime University, Japan, for editing the English of this manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Crews, T.E.; Mohler, C.L.; Power, A.G. Energetics and ecosystem integrity: The defining principles of sustainable agriculture. Am. J. Altern. Agric. 1991, 6, 146–149. [Google Scholar] [CrossRef]
- Kambewa, E.V. Contracting for sustainability: An Analysis of the Lake Victoria-EU. Nile Perch Chain; Wageningen Acad. Pub: Wageningen, The Netherlands, 2007; pp. 15–20. [Google Scholar]
- Begum, K.; Shammi, M.; Hasan, N.; Appiah, K.S.; Fujii, Y. Evaluation of Potential Volatile Allelopathic Plants from Bangladesh, with Sapindus mukorossi as a Candidate Species. Agronomy 2019, 10, 49. [Google Scholar] [CrossRef] [Green Version]
- Rajcan, I.; Swanton, C.J. Understanding maize–weed competition: Resource competition, light quality and the whole plant. Field Crop. Res. 2001, 71, 139–150. [Google Scholar] [CrossRef]
- Jabran, K.; Mahajan, G.; Sardana, V.; Chauhan, B.S. Allelopathy for weed control in agricultural systems. Crop. Prot. 2015, 72, 57–65. [Google Scholar] [CrossRef]
- Muñoz, M.; Torres-Pagán, N.; Peiró, R.; Guijarro, R.; Sánchez-Moreiras, A.M.; Verdeguer, M. Phytotoxic Effects of Three Natural Compounds: Pelargonic Acid, Carvacrol, and Cinnamic Aldehyde, against Problematic Weeds in Mediterranean Crops. Agronomy 2020, 10, 791. [Google Scholar] [CrossRef]
- Bhadoria, P.B.S. Allelopathy: A Natural Way towards Weed Management. Am. J. Exp. Agric. 2010, 1, 7–20. [Google Scholar] [CrossRef]
- Mada, D.; Duniya, N.; Adams, I.G. Effect of continuous application of herbicide on soil and environment with crop protection machinery in Southern Adamawa state. Int. Ref. J. Eng. Sci. 2013, 2, 4–9. [Google Scholar]
- Yang, X.; Kong, C.H. Interference of allelopathic rice with paddy weeds at the root level. Plant. Biol. 2017, 19, 584–591. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Cheng, Z.; Meng, H.; Yang, X.; Ahmad, I. Allelopathic effect of decomposed garlic (Allium sativum L.) stalk on lettuce (L. sativa var. crispa L.). Pak. J. Bot. 2013, 45, 225–233. [Google Scholar]
- Weir, T.L.; Park, S.-W.; Vivanco, J.M. Biochemical and physiological mechanisms mediated by allelochemicals. Curr. Opin. Plant. Biol. 2004, 7, 472–479. [Google Scholar] [CrossRef]
- Zaïdi, M.A.; Huda, A.; Crow, S.A. Biological activity and elemental composition of Arceuthobium oxycedri(Dwarf Mistletoe) of juniper forest of Pakistan. Acta Bot. Hung. 2008, 50, 223–230. [Google Scholar] [CrossRef]
- Favaretto, A.; Chini, S.O.; Basso, S.M.S.; Sobottka, A.M.; Bertol, C.D.; Perez, N.B. Pattern of allelochemical distribution in leaves and roots of tough lovegrass (Eragrostis plana Nees.). Aust. J. Crop Sci. 2015, 9, 1119–1125. [Google Scholar]
- Shaikh, A.C.; Gupta, A.; Chaphalkar, S.R. Identification of structurally unique molecules, phytochemical and immunological activity of medicinal plants. Int. J. Med. Pharm. Res. 2016, 4, 223–230. [Google Scholar]
- Tuyen, P.T.; Xuan, T.D.; Anh, T.T.T.; Van, T.M.; Ahmad, A.; Elzaawely, A.A.; Khanh, T.D. Weed Suppressing Potential and Isolation of Potent Plant Growth Inhibitors from Castanea crenata Sieb. et Zucc. Molecules 2018, 23, 345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selvi, E.K.; Turumatay, H.; Demir, A.; Turumatay, E.A. Phytochemical profiling and evaluation of the hepatoprotective effect of Cuscuta campestris by high-performance liquid chromatography with diode array detection. Anal. Lett. 2018, 51, 1–15. [Google Scholar] [CrossRef]
- Sołtys-Kalina, D.; Rudzińska-Langwald, A.; Kurek, W.; Gniazdowska, A.; Śliwińska, E.; Bogatek, R. Cyanamide mode of action during inhibition of onion (Allium cepa L.) root growth involves disturbances in cell division and cytoskeleton formation. Planta 2011, 234, 609–621. [Google Scholar] [CrossRef] [Green Version]
- Chen, F.; Liu, K.; Xie, Z.; Liu, M.; Chen, C. Effects of decomposing leaf litter of Leucaena leucocephala on photosynthetic traits of Cynodon dactylon and Medicago sativa. New For. 2018, 49, 667–679. [Google Scholar] [CrossRef]
- Saleh, A.M. In vitro assessment of allelopathic potential of olive processing waste on maize (Zea mays L.). Egypt. J. Exp. Biol. 2013, 9, 35–39. [Google Scholar]
- Vyvyan, J.R. Allelochemicals as leads for new herbicides and agrochemicals. Tetrahedron 2002, 58, 1631–1646. [Google Scholar] [CrossRef]
- Weston, L.A.; Duke, S.O. Weed and Crop Allelopathy. Crit. Rev. Plant. Sci. 2003, 22, 367–389. [Google Scholar] [CrossRef]
- Soltys, D.; Krasuska, U.; Bogatek, R.; Gniazdowska, R.B.A.A. Allelochemicals as Bioherbicides—Present and Perspectives. In Herbicides—Current Research and Case Studies in Use; IntechOpen: London, UK, 2013. [Google Scholar]
- Chung, I.M.; Kim, S.H.; Oh, Y.; Ali, M.; Ahmad, A. New constituents from Oryza sativa L. straw and their algicidal activities against blue-green algae. Allelopath. J. 2017, 40, 47–62. [Google Scholar] [CrossRef]
- Daniel, J.M.; Gillian, K.B.; Joseph, T.M.; Pauline, Y.L. Molecular phylogeny of Acacia Mill. (Mimosoideae: Leguminosae): Evidence for major clades and informal classification. Taxon 2010, 59, 7–19. [Google Scholar]
- Allan, G.J.; Porter, J.M. Tribal delimitation and phylogenetic relationships of Loteae and Coronilleae (Faboideae: Fabaceae) with special reference to Lotus: Evidence from nuclear ribosomal ITS sequences. Am. J. Bot. 2000, 87, 1871–1881. [Google Scholar] [CrossRef] [PubMed]
- Azad, S.; Paul, N.K.; Matin, A. Do pre-sowing treatments affect seed germination in Albizia richardiana and Lagerstroemia speciosa? Front. Agric. China 2010, 4, 181–184. [Google Scholar] [CrossRef]
- Salam, A.; Akhter, K.; Rahman, M.A.; Chowdhury, M.H.; Mridha, M.N.A.; Chowdhury, F.H. CCB preservative treatment of rajkoroi (Albizia richardiana King & Prain) wood by soaking method. Eco-Friendly Agril. J. 2019, 1, 74–77. [Google Scholar]
- Das, D.K.; Alam, M.K. Trees of Bangladesh; Bangladesh Forest Research Institute: Chittagong, Bangladesh, 2001. [Google Scholar]
- Al Faruq, M.A.; Zaman, S.; Katoh, M. Perceptions of Local People toward Community Development and Forest Conservation in Bangladesh: The Case of Sal Forests. J. For. Plan. 2017, 22, 1. [Google Scholar] [CrossRef]
- Rahman, M.M.; Das, A.K.; Asaduzzaman, M.; Biswas, S.K.; Hannan, M.O. Physical and mechanical properties of Raj Koroi (Albizia richardiana) plywood. Afr. J. Wood Sci. For. 2013, 2, 98–103. [Google Scholar]
- Joycharat, N.; Thammavong, S.; Limsuwan, S.; Homlaead, S.; Voravuthikunchai, S.P.; Yingyongnarongkul, B.-E.; Dej-Adisai, S.; Subhadhirasakul, S. Antibacterial substances from Albizia myriophylla wood against cariogenic Streptococcus mutans. Arch. Pharmacal. Res. 2013, 36, 723–730. [Google Scholar] [CrossRef] [PubMed]
- Xinrong, Y.; Anmin, C.; Fang, S.; Bingyi, F.; Jinlin, Q.; Yingfu, M.; Quan, L.; Yuan, G.; Shuqian, W.; Werner, H.; et al. Encyclopedic Reference of Traditional Chinese Medicine; Springer: Berlin/Heidelberg, Germany, 2003. [Google Scholar]
- Rahman, M.; Shetu, H.J.; Sukul, A.; Rahman, I. Phytochemical and biological evaluation of albizia richardiana benth, Fabaceae family. World J. Pharm. Res. 2015, 4, 168–176. [Google Scholar]
- Islam, M.N.; Tasnim, H.; Arshad, L.; Haque, A.; Tareq, S.M.; Kamal, A.T.M.M.; Rahman, M.; Reza, A.S.M.A.; Chowdhury, K.A.A.; Tareq, A.M. Stem extract of Albizia richardiana exhibits potent antioxidant, cytotoxic, antimicrobial, anti-inflammatory and thrombolytic effects through in vitro approach. Clin. Phytosci. 2020, 6, 1–9. [Google Scholar] [CrossRef]
- Xuan, T.D.; Shinkichi, T.; Khanh, T.D.; Chung, I.M. Biological control of weeds and plant pathogens in paddy rice by exploiting plant allelopathy: An overview. Crop. Prot. 2005, 24, 197–206. [Google Scholar] [CrossRef]
- Kato, T.; Tsunakawa, M.; Sasaki, N.; Aizawa, H.; Fujita, K.; Kitahara, Y.; Takahashi, N. Growth and germination inhibitors in rice husks. Phytochemistry 1977, 16, 45–48. [Google Scholar] [CrossRef]
- Kai, H.; Baba, M.; Okuyama, T. Two new megastigmanes from the leaves of Cucumis sativus. Chem. Pharm. Bull. 2007, 55, 133–136. [Google Scholar] [CrossRef] [Green Version]
- Hodges, R.; Porte, A.L. The structure of loliolide: A terpene from Lolium perenne. Tetrahedron 1964, 20, 1463–1467. [Google Scholar] [CrossRef]
- Valdes, L.J. Loliolide from Salvia divinorum. J. Nat. Prod. 1986, 49, 171. [Google Scholar] [CrossRef]
- Kimura, J.; Maki, N. New Loliolide Derivatives from the Brown AlgaUndaria pinnatifida. J. Nat. Prod. 2002, 65, 57–58. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.S.; Zaman, F.; Iwasaki, A.; Suenaga, K.; Kato-Noguchi, H. Phytotoxic potential of Chrysopogon aciculatus (Retz.) Trin. (Poaceae). Weed Biol. Manag. 2019, 19, 51–58. [Google Scholar] [CrossRef]
- Rob, M.; Hossen, K.; Iwasaki, A.; Suenaga, K.; Kato-Noguchi, H. Phytotoxic Activity and Identification of Phytotoxic Substances from Schumannianthus dichotomus. Plants 2020, 9, 102. [Google Scholar] [CrossRef] [Green Version]
- Hossen, K.; Das, K.R.; Okada, S.; Iwasaki, A.; Suenaga, K.; Kato-Noguchi, H. Allelopathic Potential and Active Substances from Wedelia Chinensis (Osbeck). Foods 2020, 9, 1591. [Google Scholar] [CrossRef]
- Hossen, K.; Kato-Noguchi, H. Determination of allelopathic properties of Acacia catechu (L.f.) Willd. Not. Bot. Horti Agrobot. Cluj-Napoca 2020, 48, 2050–2059. [Google Scholar] [CrossRef]
- Scognamiglio, M.; D’Abrosca, B.; Esposito, A.; Pacifico, S.; Monaco, P.; Fiorentino, A. Plant growth inhibitors: Allelopathic role or phytotoxic effects? Focus on Mediterranean biomes. Phytochem. Rev. 2013, 12, 803–830. [Google Scholar] [CrossRef]
- Reigosa, M.J.; Sánchez-Moreiras, A.; González, L. Ecophysiological Approach in Allelopathy. Crit. Rev. Plant. Sci. 1999, 18, 577–608. [Google Scholar] [CrossRef]
- Zhuo, Y.H.; Yu, J.Q. Allelochemicals and photosynthesis. In Allelopathy: A Physiological Process with Ecological Implications; Reigosa, M.J., Pedrol, N., González, L., Eds.; Kluwer Academic Publishers: Amsterdam, The Netherlands, 2006; pp. 127–139. [Google Scholar]
- Yan, X.; Lynch, J.P.; Beebe, S.E. Genetic Variation for Phosphorus Efficiency of Common Bean in Contrasting Soil Types: I. Vegetative Response. Crop. Sci. 1995, 35, 1086–1093. [Google Scholar] [CrossRef]
- Wu, H.; Pratley, J.; Lemerled, D.; Haig, T. Crop cultivars with allelopathic capability. Weed Res. 1999, 39, 171–180. [Google Scholar] [CrossRef]
- Netsere, A.; Mendesil, E. Allelopathic effects of Parthenium hysterophorus L. aqueous extracts on soybean (Glycine max L.) and haricot bean (Phaseolus vulgaris L.) seed germination, shoot and root growth and dry matter production. J. Appl. Bot. Food Qual. 2011, 84, 219–222. [Google Scholar]
- Liu, J.; Xie, M.; Li, X.; Jin, H.; Yang, X.; Yan, Z.; Su, A.; Qin, B. Main Allelochemicals from the Rhizosphere Soil of Saussurea lappa (Decne.) Sch. Bip. and Their Effects on Plants’ Antioxidase Systems. Molecules 2018, 23, 2506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Islam, M.S.; Kato-Noguchi, H. Phytotoxicity assessment of Cyperus difformis (L.) towards a sustainable weed management option. J. Anim. Plant. Sci. 2016, 2, 1765–1771. [Google Scholar]
- Nishida, N.; Tamotsu, S.; Nagata, N.; Saito, C.; Sakai, A. Allelopathic Effects of Volatile Monoterpenoids Produced by Salvia leucophylla: Inhibition of Cell Proliferation and DNA Synthesis in the Root Apical Meristem of Brassica campestris Seedlings. J. Chem. Ecol. 2005, 31, 1187–1203. [Google Scholar] [CrossRef]
- Tanveer, A.; Jabbar, M.K.; Kahliq, A.; Matloob, A.; Abbas, R.N.; Javaid, M.M. Allelopathic effects of aqueous and organic fractions of Euphorbia dracunculoides Lam. on germination and seedling growth of chickpea and wheat. Chil. J. Agric. Res. 2012, 72, 495–501. [Google Scholar] [CrossRef]
- Pan, L.; Sinden, M.R.; Kennedy, A.H.; Chai, H.; Watson, L.E.; Graham, T.L.; Kinghorn, A.D. Bioactive constituents of Helianthus tuberosus (Jerusalem artichoke). Phytochem. Lett. 2009, 2, 15–18. [Google Scholar] [CrossRef]
- Takasugi, M.; Anetai, M.; Katsui, N.; Masamune, T. The occurrence of vomifoliol, dehydrovomifoliol and dihydrophaseic acid in the roots of “kidney bean” (Phaseolus vulgaris L.). Chem. Lett. 1973, 2, 245–248. [Google Scholar] [CrossRef]
- Kim, I.; Chin, Y.-W.; Lim, S.W.; Kim, Y.C.; Kim, J. Norisoprenoids and hepatoprotective flavone glycosides from the aerial parts of Beta vulgaris var. cicla. Arch. Pharmacal Res. 2004, 27, 600–603. [Google Scholar] [CrossRef]
- Machado, F.B.; Yamamoto, R.E.; Zanoli, K.; Nocchi, S.R.; Novello, C.R.; Schuquel, I.T.A.; Sakuragui, C.M.; Luftmann, H.; Ueda-Nakamura, T.; Nakamura, C.V.; et al. Evaluation of the Antiproliferative Activity of the Leaves from Arctium lappa by a Bioassay-Guided Fractionation. Molecules 2012, 17, 1852–1859. [Google Scholar] [CrossRef]
- Yang, Y.; Bakri, M.; Gu, D.; Aisa, H.A. Separation of (s)-dehydrovomifoliol from leaves of nitraria sibirica pall. by high-speed counter-current chromatography. J. Liq. Chromatogr. Relat. Technol. 2012, 36, 573–582. [Google Scholar] [CrossRef]
- Mayer, H. Synthesis of optically active carotenoids and related compounds. Pure Appl. Chem. 1979, 51, 535–564. [Google Scholar] [CrossRef] [Green Version]
- Ren, Y.; Shen, L.; Zhang, D.-W.; Dai, S.-J. Two New Sesquiterpenoids from Solanum lyratum with Cytotoxic Activities. Chem. Pharm. Bull. 2009, 57, 408–410. [Google Scholar] [CrossRef] [Green Version]
- Grabarczyk, M.; Winska, K.; Maczka, W.; Potaniec, B.; Anioł, M. Loliolide‒The most ubiquitous lactone. Folia Biol. Oecol. 2015, 11, 1–8. [Google Scholar] [CrossRef]
- Erosa-Rejón, G.; Peña-Rodríguez, L.M.; Sterner, O. Secondary Metabolites from Heliotropium angiospermum. J. Mex. Chem. Soc. 2019, 53, 44–47. [Google Scholar] [CrossRef]
- Zhou, B.; Kong, C.H.; Li, Y.-H.; Wang, P.; Xu, X.-H. Crabgrass (Digitaria sanguinalis) Allelochemicals That Interfere with Crop Growth and the Soil Microbial Community. J. Agric. Food Chem. 2013, 61, 5310–5317. [Google Scholar] [CrossRef]
- Kato-Noguchi, H.; Moriyasu, M.; Ohno, O.; Suenaga, K. Growth limiting effects on various terrestrial plant species by an allelopathic substance, loliolide, from water hyacinth. Aquat. Bot. 2014, 117, 56–61. [Google Scholar] [CrossRef]
- Masum, S.M.; Akamine, H.; Hossain, M.A.; Sakagami, J.I.; Ishii, T.; Gima, S. Assessment of the allelopathic potential and iden-tification of the phytotoxic substances from the straw of Bangladeshi indigenous rice variety ‘Goria’. Appl. Ecol. Environ. Res. 2020, 18, 5547–5560. [Google Scholar] [CrossRef]
- Yang, X.; Kang, M.-C.; Lee, K.-W.; Kang, S.-M.; Lee, W.-W.; Jeon, Y.-J. Antioxidant activity and cell protective effect of loliolide isolated from Sargassum ringgoldianum subsp. coreanum. ALGAE 2011, 26, 201–208. [Google Scholar] [CrossRef]
- Kim, H.-S.; Wang, L.; Fernando, I.P.S.; Je, J.-G.; Ko, S.-C.; Kang, M.C.; Lee, J.M.; Yim, M.-J.; Jeon, Y.-J.; Lee, D.-S. Antioxidant efficacy of (−)-loliolide isolated from Sargassum horneri against AAPH-induced oxidative damage in Vero cells and zebrafish models in vivo. Environ. Biol. Fishes 2020, 32, 3341–3348. [Google Scholar] [CrossRef]
- Park, S.H.; Kim, D.S.; Kim, S.; Lorz, L.R.; Choi, E.; Lim, H.Y.; Hossain, M.A.; Chang, S.; Choi, Y.I.; Park, K.J.; et al. Loliolide Presents Antiapoptosis and Antiscratching Effects in Human Keratinocytes. Int. J. Mol. Sci. 2019, 20, 651. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Evangelopoulos, D.; Bhakta, S.; Gray, A.I.; Seidel, V. Antitubercular activity of Arctium lappa and Tussilago farfara extracts and constituents. J. Ethnopharmacol. 2014, 155, 796–800. [Google Scholar] [CrossRef]
- Hunyadi, A.; Veres, K.; Danko, B.; Kele, Z.; Wéber, E.; Hetenyi, A.; Zupkó, I.; Hsieh, T.-J. In vitroAnti-diabetic Activity and Chemical Characterization of an Apolar Fraction ofMorus albaLeaf Water Extract. Phytother. Res. 2012, 27, 847–851. [Google Scholar] [CrossRef]
- Colom, O.A.; Popich, S.; Bardon, A. Bioactive constituents from Rollinia emarginata (Annonaceae). Nat. Prod. Res. 2007, 21, 254–259. [Google Scholar] [CrossRef]
- Zaman, F.; Iwasaki, A.; Suenaga, K.; Kato-Noguchi, H. Two allelopathic substances fromPaspalum commersoniiLam. Acta Agric. Scand. Sect. B Plant. Soil Sci. 2017, 68, 342–348. [Google Scholar] [CrossRef]
- Pan, W.; Liu, K.; Guan, Y.; Tan, G.T.; Van Hung, N.; Cuong, N.M.; Soejarto, D.D.; Pezzuto, J.M.; Fong, H.H.S.; Hongjie, Z. Bioactive Compounds from Vitex leptobotrys. J. Nat. Prod. 2014, 77, 663–667. [Google Scholar] [CrossRef] [Green Version]
- DellaGreca, M.; Fiorentino, A.; Monaco, P.; Previtera, L.; Temussi, F.; Zarrelli, A. New dimeric phenanthrenoids from the rhizomes of Juncus acutus. Structure determination and antialgal activity. Tetrahedron 2003, 59, 2317–2324. [Google Scholar] [CrossRef]
- Kobayashi, M.; Ueda, M.; Furumoto, T.; Kawanami, Y. Retarding Activity of 6-O-Acyl-D-alloses against Plant Growth. Biosci. Biotechnol. Biochem. 2010, 74, 216–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Phytotoxic effects of Albizia richardiana aqueous methanol extracts on shoot and root growth of the cress and barnyard grass. The test plant species were treated at the following concentrations: 0.003, 0.01, 0.03, 0.1, 0.3, and 1.0 g dry weight (DW) equivalent extract of Albizia richardiana/mL. The mean ± standard error from the two independent experiments with three replications for every treatment are displayed (seedling number per treatment = 10, n = 60). The vertical bar denotes standard error of the mean. Various letters denote the significant differences according to Tukey’s HSD test at a 0.05 probability level.
Figure 1.
Phytotoxic effects of Albizia richardiana aqueous methanol extracts on shoot and root growth of the cress and barnyard grass. The test plant species were treated at the following concentrations: 0.003, 0.01, 0.03, 0.1, 0.3, and 1.0 g dry weight (DW) equivalent extract of Albizia richardiana/mL. The mean ± standard error from the two independent experiments with three replications for every treatment are displayed (seedling number per treatment = 10, n = 60). The vertical bar denotes standard error of the mean. Various letters denote the significant differences according to Tukey’s HSD test at a 0.05 probability level.

Figure 2.
The phytotoxic effects on cress seedling growth of aqueous and ethyl acetate fractions obtained by the partitioning of Albizia richardiana extracts at the concentrations of 0.3 and 0.6 g DW equivalent extract/mL. The means ± standard errors for every treatment from the two separate experiments with 10 seedlings are displayed. Different letters denote the significant differences according to Tukey’s HSD test at a 0.05 probability level.
Figure 2.
The phytotoxic effects on cress seedling growth of aqueous and ethyl acetate fractions obtained by the partitioning of Albizia richardiana extracts at the concentrations of 0.3 and 0.6 g DW equivalent extract/mL. The means ± standard errors for every treatment from the two separate experiments with 10 seedlings are displayed. Different letters denote the significant differences according to Tukey’s HSD test at a 0.05 probability level.

Figure 3.
The allelopathic effect of Albizia richardiana extracts on shoot and root growth of cress. The cress seeds were treated in the concentration of 1.2 g DW equivalent extract/mL using the following fractions: F1 (20% aqueous methanol), F2 (40% aqueous methanol), F3 (60% aqueous methanol), F4 (80% aqueous methanol), and F5 (methanol). The means ± standard errors for every treatment from the two separate experiments with 10 seedlings are displayed. Various letters denote the significant differences according to Tukey’s HSD test at a 0.05 probability level.
Figure 3.
The allelopathic effect of Albizia richardiana extracts on shoot and root growth of cress. The cress seeds were treated in the concentration of 1.2 g DW equivalent extract/mL using the following fractions: F1 (20% aqueous methanol), F2 (40% aqueous methanol), F3 (60% aqueous methanol), F4 (80% aqueous methanol), and F5 (methanol). The means ± standard errors for every treatment from the two separate experiments with 10 seedlings are displayed. Various letters denote the significant differences according to Tukey’s HSD test at a 0.05 probability level.

Figure 4.
The chemical structures of the isolated allelopathic compounds dehydrovomifoliol and loliolide from the extracts of Albizia richardiana.
Figure 4.
The chemical structures of the isolated allelopathic compounds dehydrovomifoliol and loliolide from the extracts of Albizia richardiana.

Figure 5.
The phytotoxic effects of dehydrovomifoliol on the growth of the cress seedling. The mean ± standard error from the two independent experiments with three replications (10 seedlings per replication) for each experiment is presented. Different letters denote the significant differences according to Tukey’s HSD test at a 0.05 probability level.
Figure 5.
The phytotoxic effects of dehydrovomifoliol on the growth of the cress seedling. The mean ± standard error from the two independent experiments with three replications (10 seedlings per replication) for each experiment is presented. Different letters denote the significant differences according to Tukey’s HSD test at a 0.05 probability level.

Figure 6.
The phytotoxic effects of loliolide on the growth of the cress seedling. The mean ± standard error from the two independent experiments with three replications (10 seedlings per replication) for each experiment is presented. Different letters denote the significant differences according to Tukey’s HSD test at a 0.05 probability level.
Figure 6.
The phytotoxic effects of loliolide on the growth of the cress seedling. The mean ± standard error from the two independent experiments with three replications (10 seedlings per replication) for each experiment is presented. Different letters denote the significant differences according to Tukey’s HSD test at a 0.05 probability level.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Concentrations needed for the 50% inhibition of growth (I50 values) of the tested plants by the Albizia richardiana aqueous methanol extracts.
Table 1.
Concentrations needed for the 50% inhibition of growth (I50 values) of the tested plants by the Albizia richardiana aqueous methanol extracts.
| Tested Species | I50 Values (g Dry Weight Equivalent Extract/mL) | ||
|---|---|---|---|
| Shoot | Root | ||
| Dicot | Cress | 0.019 | 0.015 |
| Monocot | Barnyard grass | 0.049 | 0.008 |
Table 2.
The concentration needed for the 50% inhibition of growth (I50 values) of the tested plant by dehydrovomifoliol and loliolide.
Table 2.
The concentration needed for the 50% inhibition of growth (I50 values) of the tested plant by dehydrovomifoliol and loliolide.
| Test Plant | Dehydrovomifoliol | Loliolide | |
|---|---|---|---|
| (mM) | |||
| Cress | Shoot | 3.1633 | 0.0341 |
| Root | 3.0155 | 0.0256 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Hossen, K.; Iwasaki, A.; Suenaga, K.; Kato-Noguchi, H. Phytotoxic Activity and Growth Inhibitory Substances from Albizia richardiana (Voigt.) King & Prain. Appl. Sci. 2021, 11, 1455. https://doi.org/10.3390/app11041455
AMA Style
Hossen K, Iwasaki A, Suenaga K, Kato-Noguchi H. Phytotoxic Activity and Growth Inhibitory Substances from Albizia richardiana (Voigt.) King & Prain. Applied Sciences. 2021; 11(4):1455. https://doi.org/10.3390/app11041455
Chicago/Turabian StyleHossen, Kawsar, Arihiro Iwasaki, Kiyotake Suenaga, and Hisashi Kato-Noguchi. 2021. "Phytotoxic Activity and Growth Inhibitory Substances from Albizia richardiana (Voigt.) King & Prain" Applied Sciences 11, no. 4: 1455. https://doi.org/10.3390/app11041455
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.