Oxyresveratrol Induces Autophagy via the ER Stress Signaling Pathway, and Oxyresveratrol-Induced Autophagy Stimulates MUC2 Synthesis in Human Goblet Cells



Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cell Culture
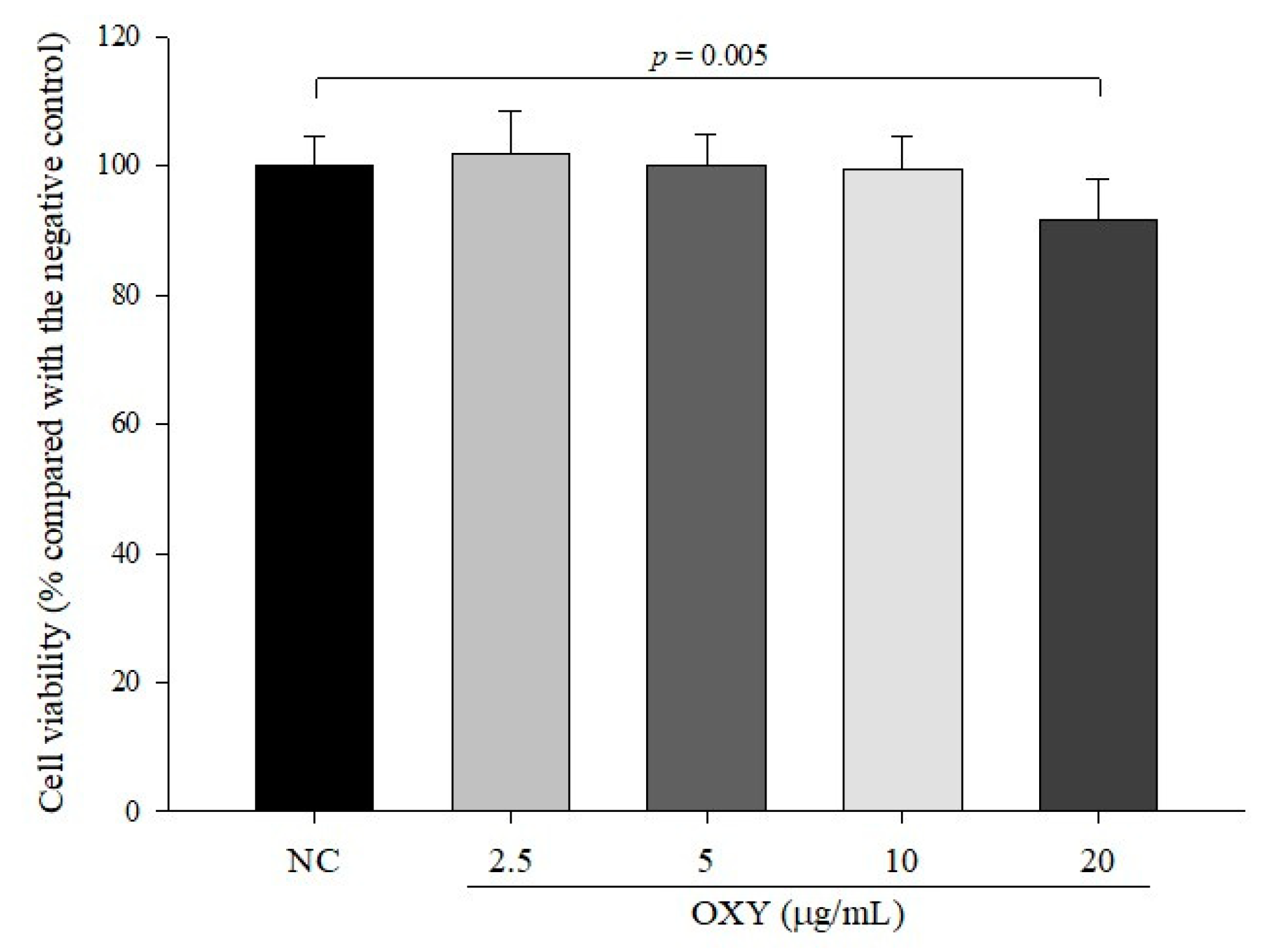
2.3. MTT Measurement of OXY Cytotoxicity
2.4. Quantitative Real-Time Polymerase Chain Reaction (qPCR)
2.5. Western Blot Analysis
2.6. Two-Dimensional Electrophoresis (2DE) and Peptide Mass Fingerprinting (PMF)
2.7. Autophagy Detection
2.8. Immunocytochemistry (ICC)
2.9. Statistical Analysis
3. Results
3.1. Cytotoxicity of OXY in LS 174T Goblet Cells
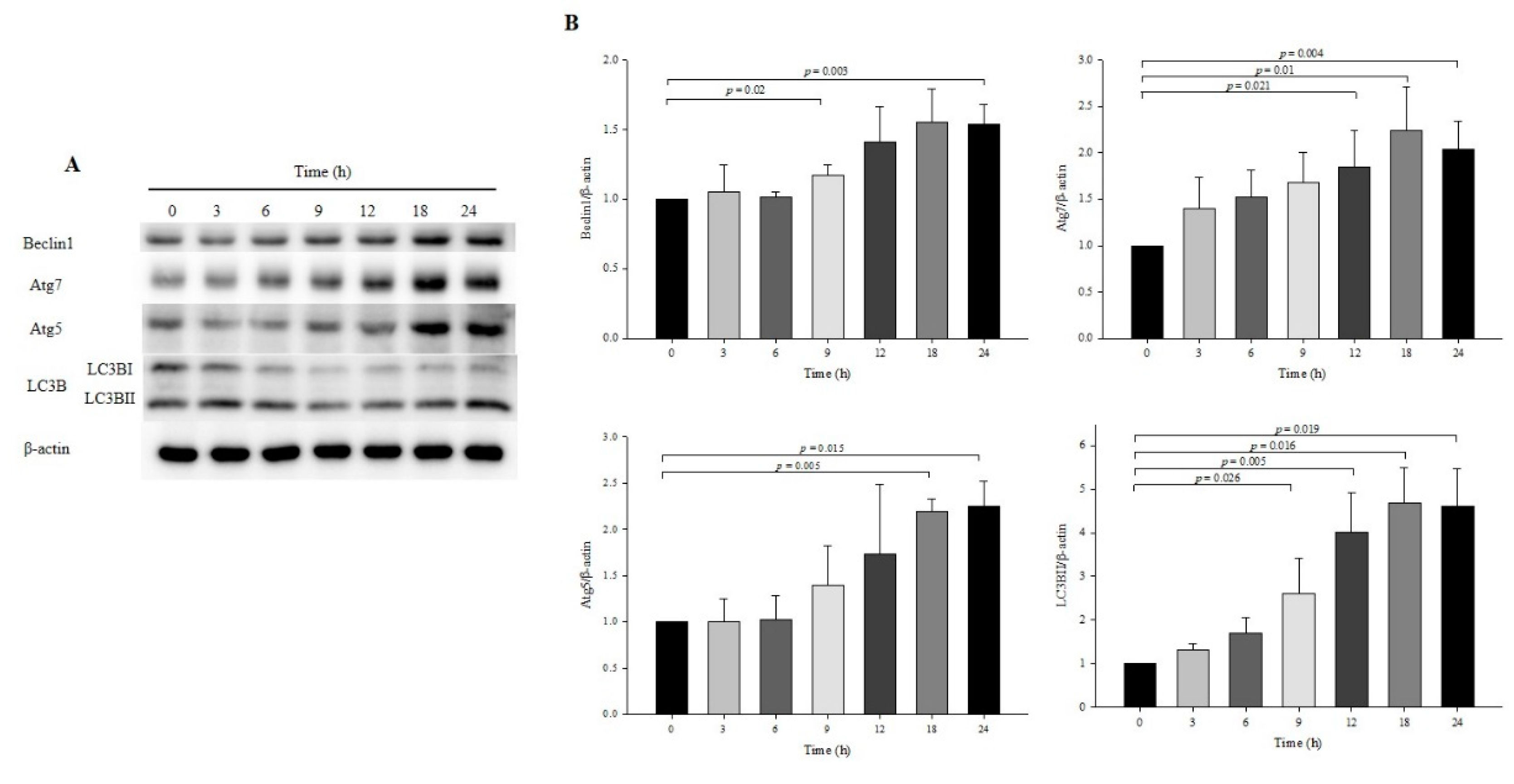
3.2. Effect of OXY on Autophagy Development at Specific Times
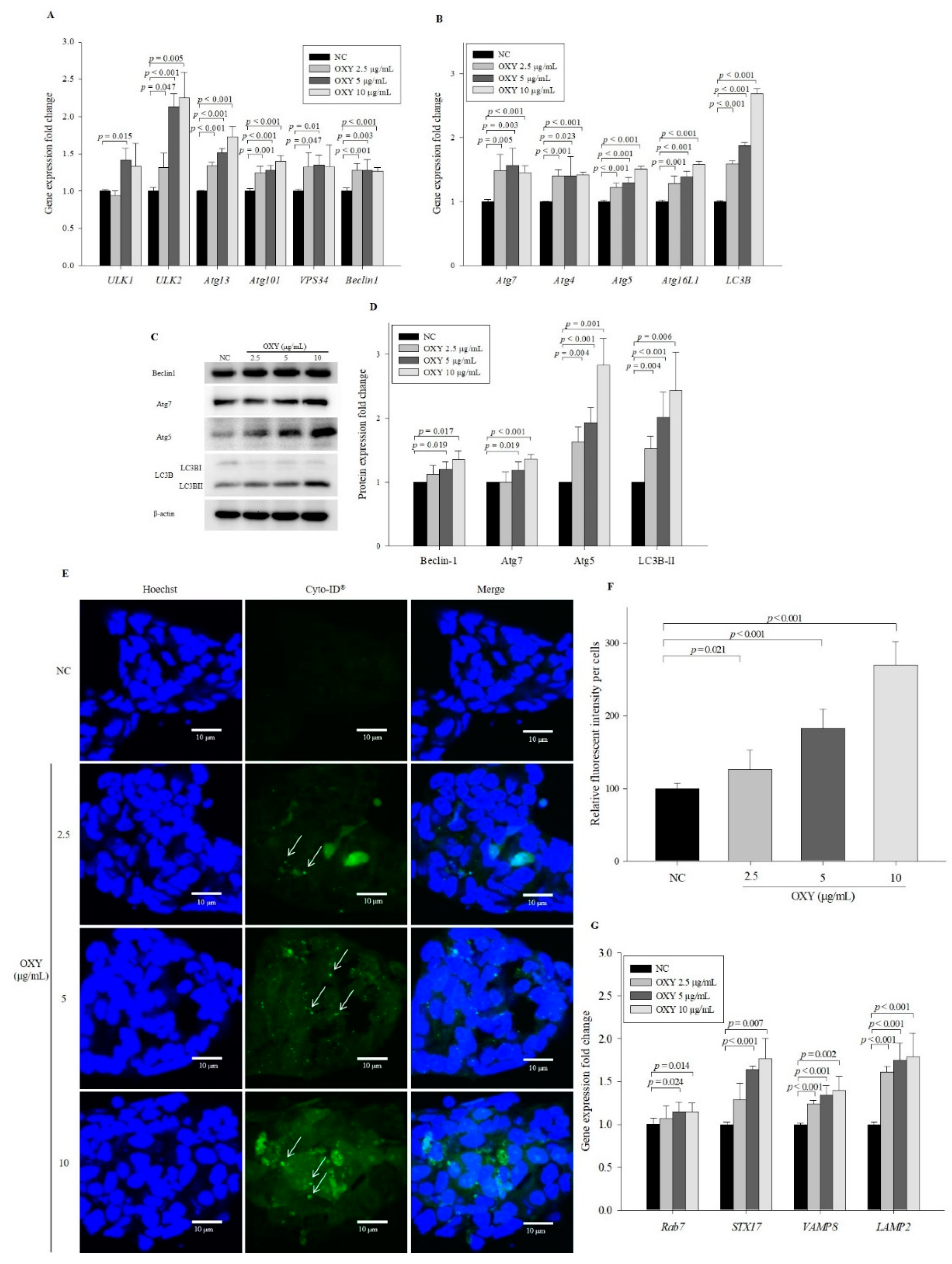
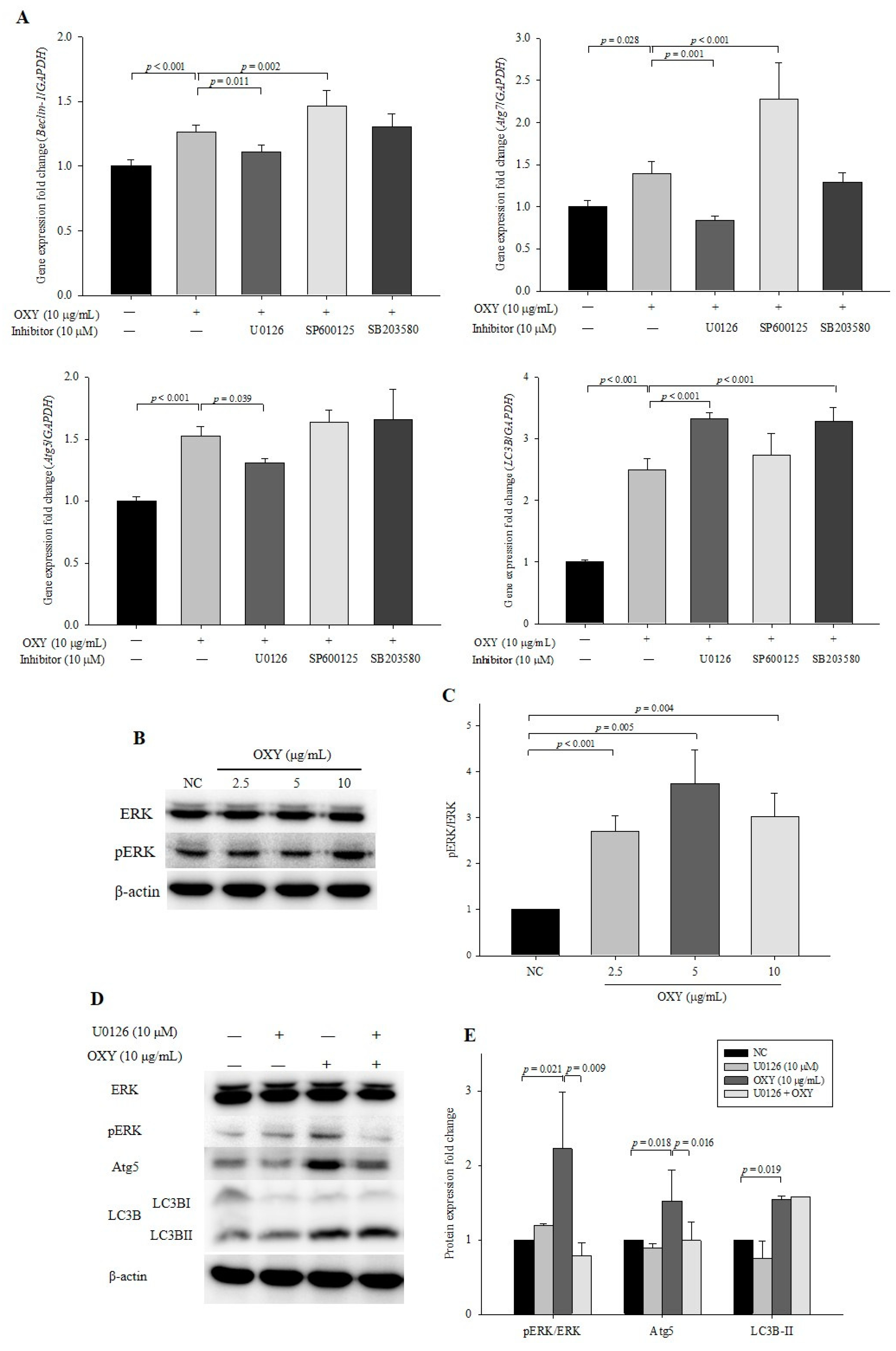
3.3. OXY Increases Expression of Autophagy-Related Factors
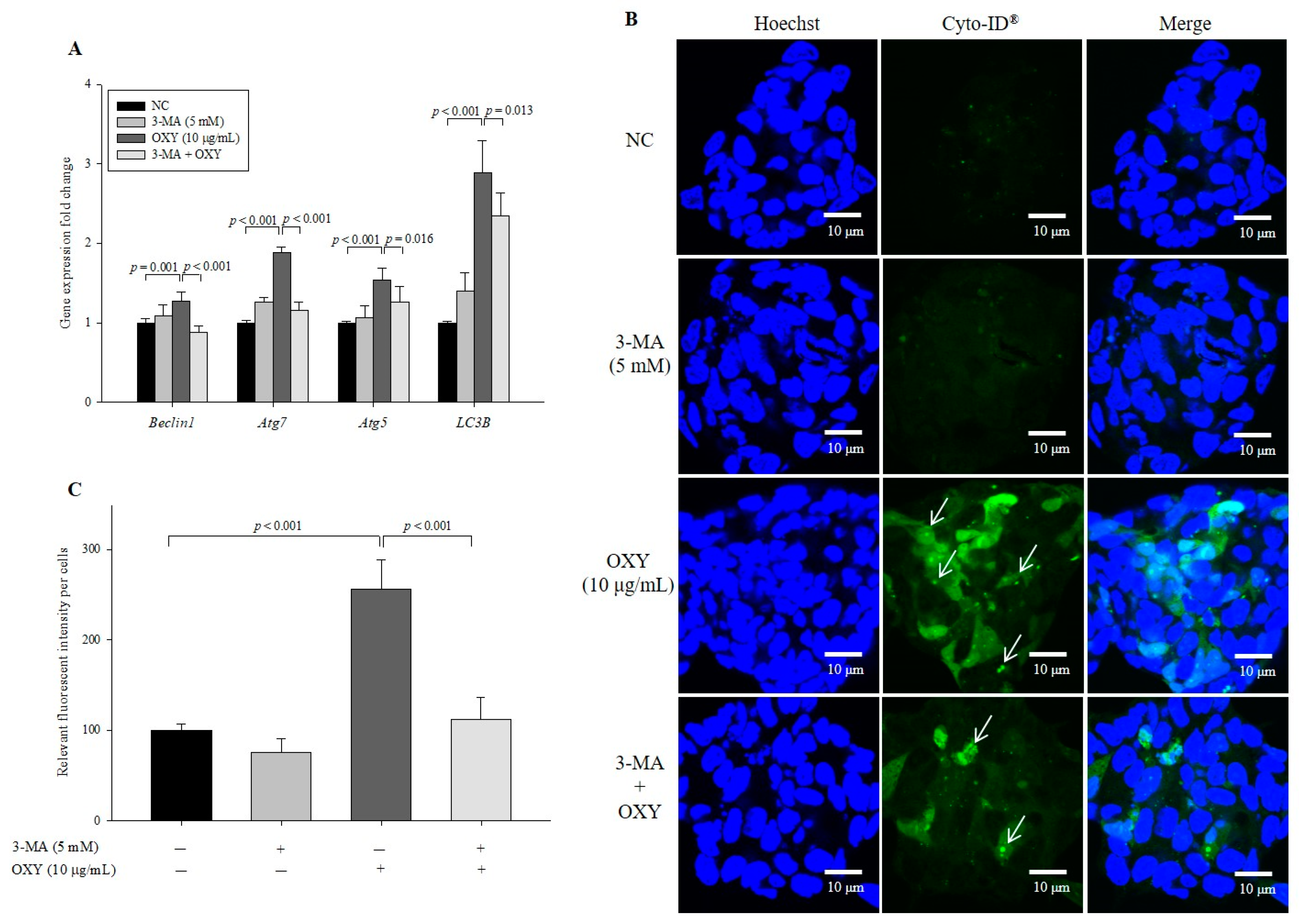
3.4. 3-Methyladenine (3-MA) Decreases the Effect of OXY on Autophagy
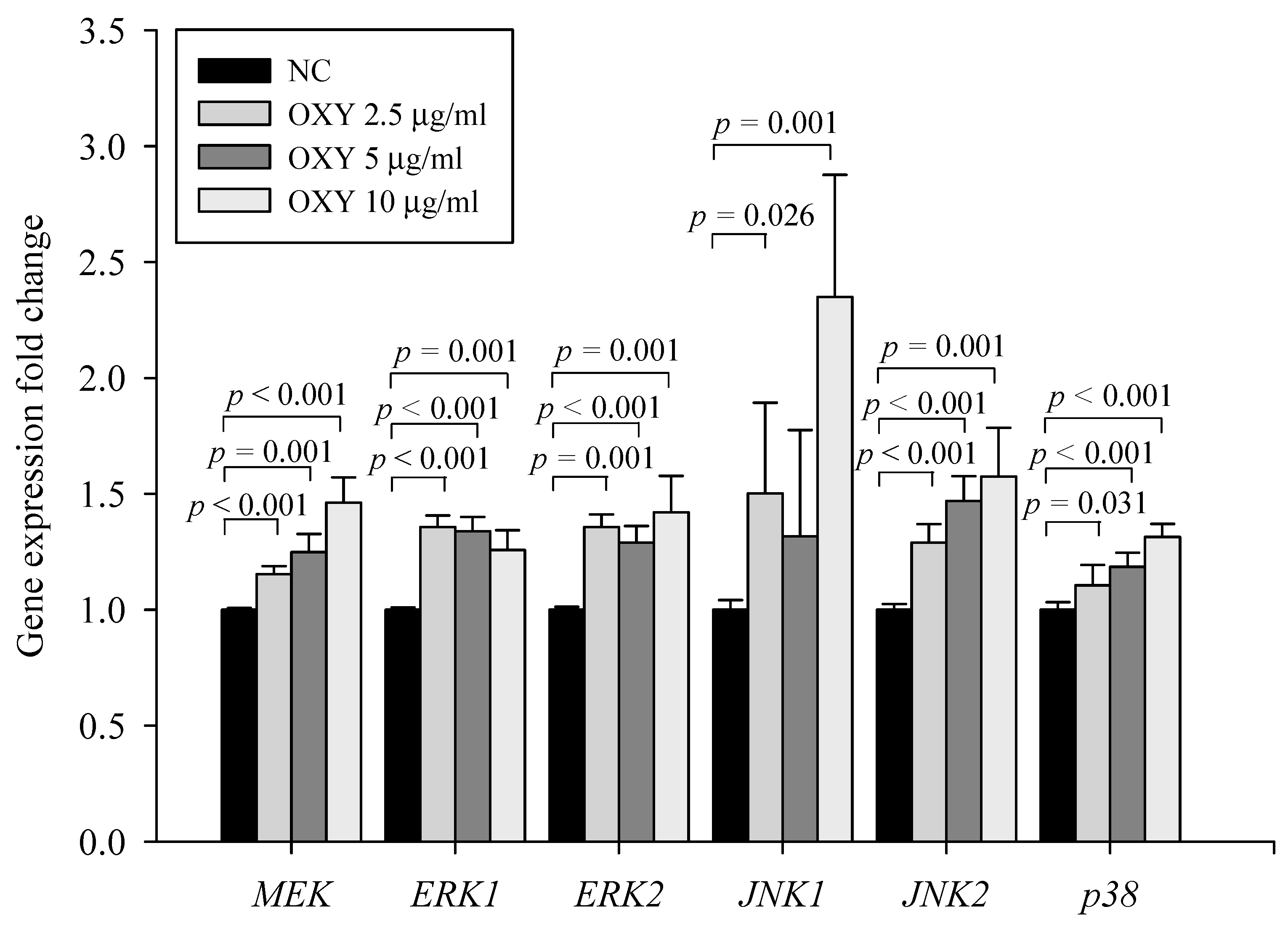
3.5. OXY Stimulates the Mitogen-Activated Protein Kinase (MAPK) Signaling Pathway
3.6. Effect of MAPK kinase Inhibitors on Expression Levels of Autophagy-Related Genes and Proteins
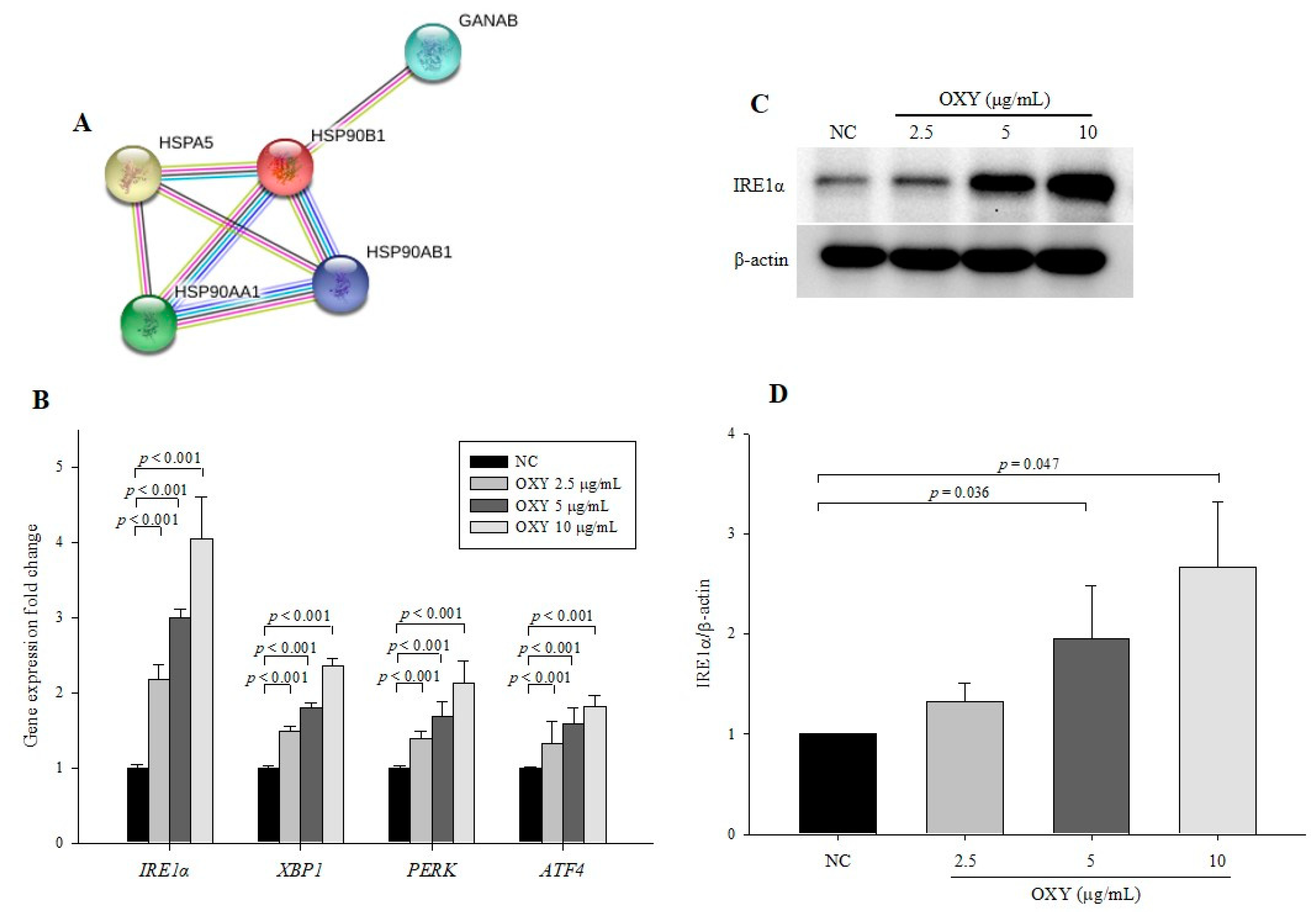
3.7. OXY Stimulates the ER Stress Signaling Pathway
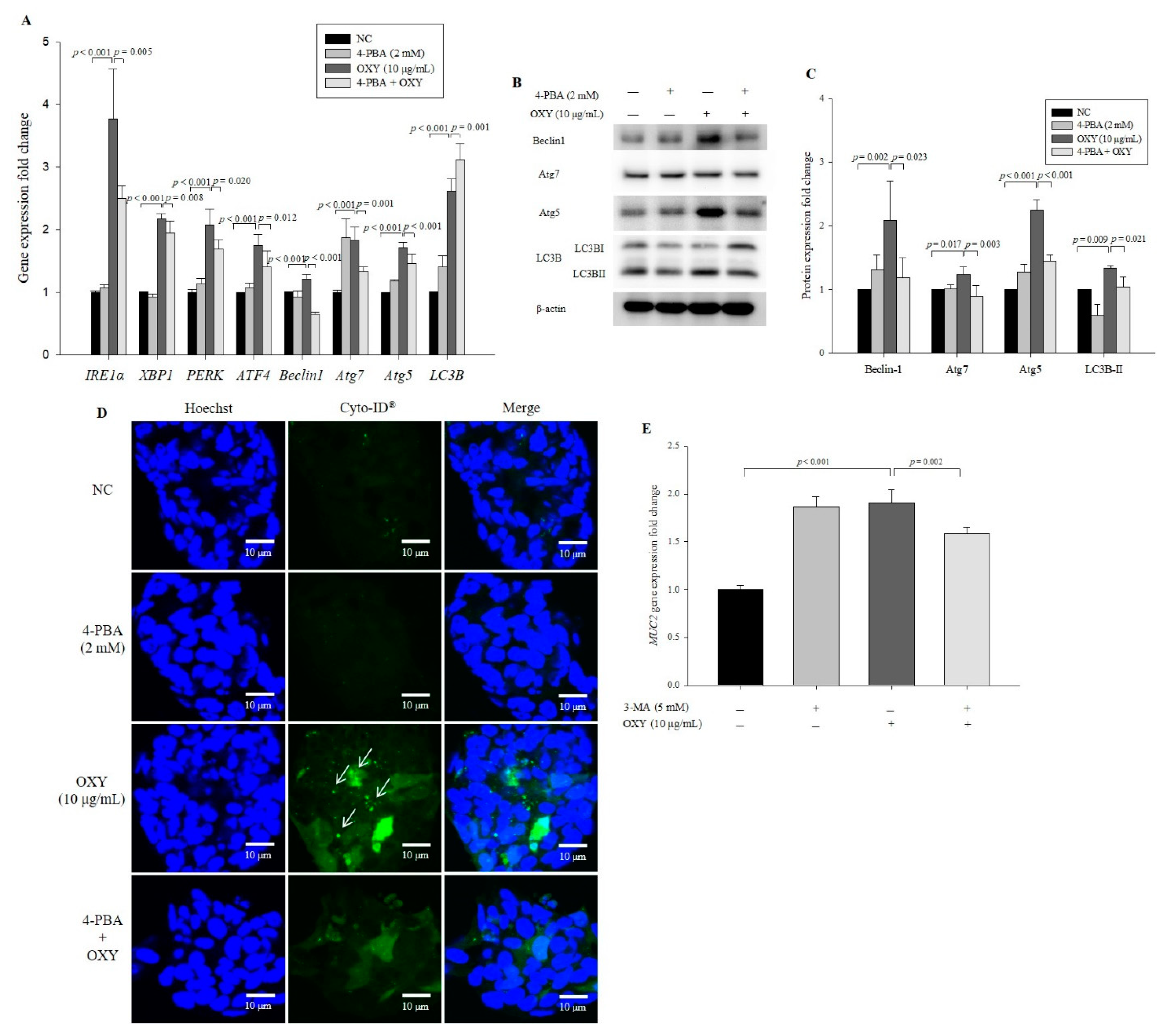
3.8. An ER Stress Inhibitor Decreases Expression Levels of Autophagy-Related Genes and Proteins
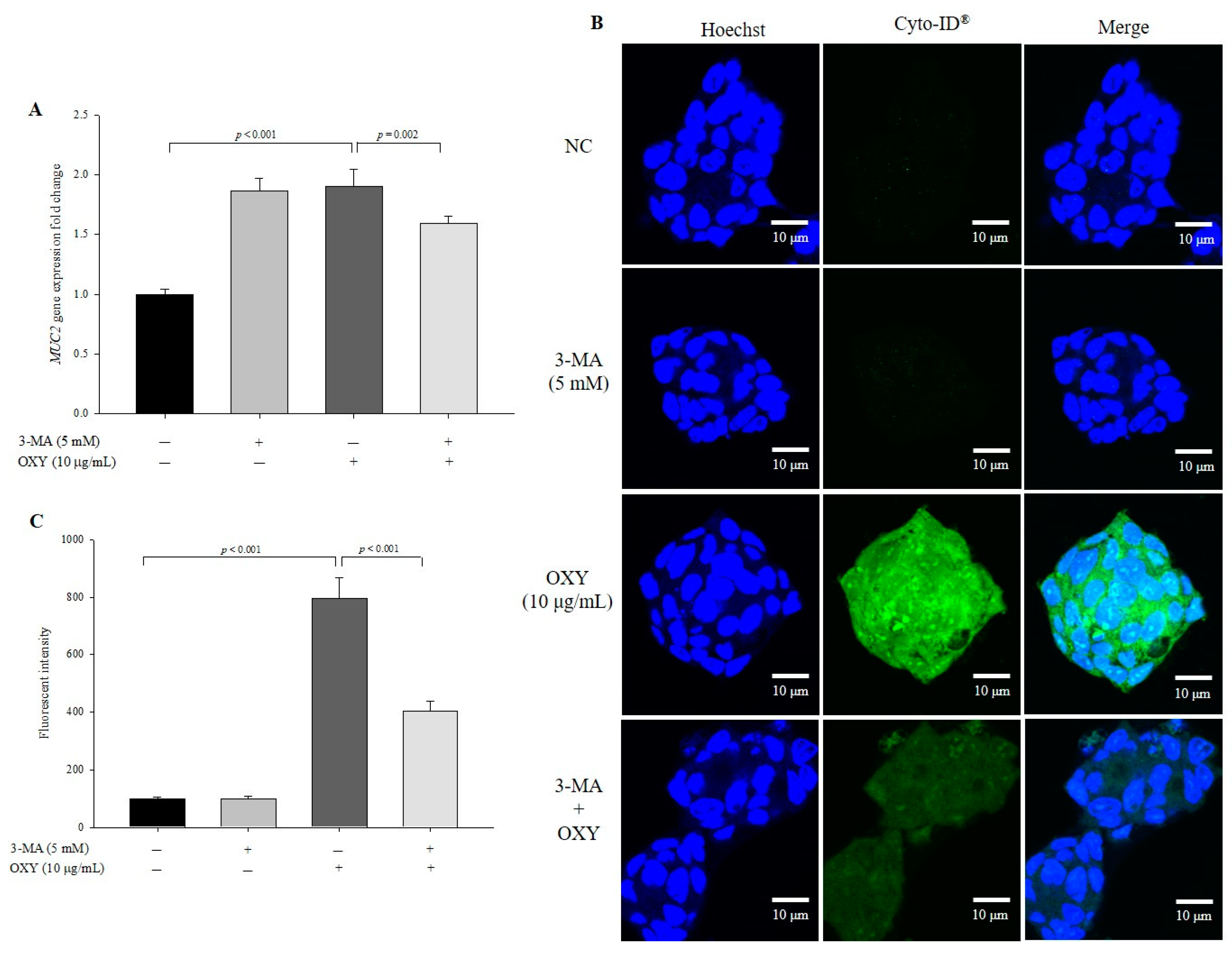
3.9. OXY-Induced Autophagy Affects Expression of MUC2
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| OXY | oxyresveratrol |
| LC3 | microtubule-associated protein 1A/1B-light chain 3 |
| PE | phosphatidylethanolamine |
| ATGs | autophagy-related genes |
| ULK | unc-51-like kinase |
| PI3K | phosphatidylinositol 3-kinase |
| RPMI | Roswell Park Memorial Institute |
| DMSO | dimethyl sulfoxide |
| 3-MA | 3-methyladenine |
| 2 DE | two-dimensional electrophoresis |
| PMF | peptide mass fingerprinting |
| IEF | isoelectric focusing |
| DTT | dithiothreitol |
| ICC | immunocytochemistry |
| ER | endoplasmic reticulum |
| 4-PBA | 4-phenylbutyric acid |
References
- Glick, D.; Barth, S.; Macleod, K.F. Autophagy: Cellular and molecular mechanisms. J. Pathol. 2010, 221, 3–12. [Google Scholar] [CrossRef] [Green Version]
- Badadani, M. Autophagy mechanism, regulation, functions, and disorders. ISRN Cell Biol. 2012, 2012, 927064. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Klionsky, D.J. The regulation of autophagy–unanswered questions. J. Cell Sci. 2011, 124, 161–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saha, S.; Panigrahi, D.P.; Patil, S.; Bhutia, S.K. Autophagy in health and disease: A comprehensive review. Biomed. Pharmacother. 2018, 104, 485–495. [Google Scholar] [CrossRef] [PubMed]
- Sun, K.; Xu, L.; Jing, Y.; Han, Z.; Chen, X.; Cai, C.; Zhao, P.; Zhao, X.; Yang, L.; Wei, L. Autophagy-deficient Kupffer cells promote tumorigenesis by enhancing mtROS-NF-κB-IL1α/β-dependent inflammation and fibrosis during the preneoplastic stage of hepatocarcinogenesis. Cancer Lett. 2017, 388, 198–207. [Google Scholar] [CrossRef]
- Takamura, A.; Komatsu, M.; Hara, T.; Sakamoto, A.; Kishi, C.; Waguri, S.; Eishi, Y.; Hino, O.; Tanaka, K.; Mizushima, N. Autophagy-deficient mice develop multiple liver tumors. Genes Dev. 2011, 25, 795–800. [Google Scholar] [CrossRef] [Green Version]
- Fujikake, N.; Shin, M.; Shimizu, S. Association between autophagy and neurodegenerative diseases. Front. Neurosci. 2018, 12, 255. [Google Scholar] [CrossRef] [Green Version]
- Peng, X.; Wang, Y.; Li, H.; Fan, J.; Shen, J.; Yu, X.; Zhou, Y.; Mao, H. ATG5-mediated autophagy suppresses NF-κB signaling to limit epithelial inflammatory response to kidney injury. Cell Death Dis. 2019, 10, 253. [Google Scholar] [CrossRef] [Green Version]
- Feng, Y.; He, D.; Yao, Z.; Klionsky, D.J. The machinery of macroautophagy. Cell Res. 2014, 24, 24–41. [Google Scholar] [CrossRef] [Green Version]
- Van der Sluis, M.; De Koning, B.A.; De Bruijn, A.C.; Velcich, A.; Meijerink, J.P.; Van Goudoever, J.B.; Büller, H.A.; Dekker, J.; Van Seuningen, I.; Renes, I.B. Muc2-deficient mice spontaneously develop colitis, indicating that MUC2 is critical for colonic protection. Gastroenterology 2006, 131, 117–129. [Google Scholar] [CrossRef]
- Johansson, M.E. Mucus layers in inflammatory bowel disease. Inflamm. Bowel Dis. 2014, 20, 2124–2131. [Google Scholar] [CrossRef]
- Wei, J.; Chen, J.-R.; Pais, E.M.A.; Wang, T.-Y.; Miao, L.; Li, L.; Li, L.-Y.; Qiu, F.; Hu, L.-M.; Gao, X.-M. Oxyresveratrol is a phytoestrogen exerting anti-inflammatory effects through NF-κB and estrogen receptor signaling. Inflammation 2017, 40, 1285–1296. [Google Scholar] [CrossRef] [PubMed]
- Joung, D.K.; Mun, S.H.; Choi, S.H.; Kang, O.H.; Kim, S.B.; Lee, Y.S.; Zhou, T.; Kong, R.; Choi, J.G.; Shin, D.W. Antibacterial activity of oxyresveratrol against methicillin-resistant Staphylococcus aureus and its mechanism. Exp. Ther. Med. 2016, 12, 1579–1584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jo, H.; Hwang, D.; Kim, J.-K.; Lim, Y.-H. Oxyresveratrol improves tight junction integrity through the PKC and MAPK signaling pathways in Caco-2 cells. Food Chem. Toxicol. 2017, 108, 203–213. [Google Scholar] [CrossRef] [PubMed]
- Lorenz, P.; Roychowdhury, S.; Engelmann, M.; Wolf, G.; Horn, T.F. Oxyresveratrol and resveratrol are potent antioxidants and free radical scavengers: Effect on nitrosative and oxidative stress derived from microglial cells. Nitric Oxide 2003, 9, 64–76. [Google Scholar] [CrossRef] [PubMed]
- Hwang, D.; Jo, H.; Kim, J.-K.; Lim, Y.-H. Oxyresveratrol-containing Ramulus mori ethanol extract attenuates acute colitis by suppressing inflammation and increasing mucin secretion. J. Funct. Foods 2017, 35, 146–158. [Google Scholar] [CrossRef]
- Hwang, D.; Jo, H.; Ma, S.-H.; Lim, Y.-H. Oxyresveratrol stimulates mucin production in an NAD+-dependent manner in human intestinal goblet cells. Food Chem. Toxicol. 2018, 118, 880–888. [Google Scholar] [CrossRef]
- Zhu, Y.; Zhao, K.-k.; Tong, Y.; Zhou, Y.-l.; Wang, Y.-X.; Zhao, P.-Q.; Wang, Z.-Y. Exogenous NAD+ decreases oxidative stress and protects H2O2-treated RPE cells against necrotic death through the up-regulation of autophagy. Sci. Rep. 2016, 6, 26322. [Google Scholar] [CrossRef] [Green Version]
- Ke, P.; Shao, B.-Z.; Xu, Z.-Q.; Chen, X.-W.; Liu, C. Intestinal autophagy and its pharmacological control in inflammatory bowel disease. Front. Immunol. 2017, 7, 695. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Anderson, N.; Esquer-Blasco, R.; HoffmanJ, P.; Anderson, N.G. A two-dimensional gel database of rat liver proteins useful in gene regulation and drug effects studies. Electrophoresis 1991, 12, 907–930. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, J.; Gharahdaghi, F.; Mische, S.M. Routine identification of proteins from sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) gels or polyvinyl difluoride membranes using matrix assisted laser desorption/ionization-time of flight-mass spectrometry (MALDI-TOF-MS). Electrophoresis 1998, 19, 1036–1045. [Google Scholar] [CrossRef] [PubMed]
- Yoshii, S.R.; Mizushima, N. Monitoring and measuring autophagy. Int. J. Mol. Sci. 2017, 18, 1865. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Salvati, A.; Boya, P. Lysosome-dependent cell death and deregulated autophagy induced by amine-modified polystyrene nanoparticles. Open Biol. 2018, 8, 170271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lane, J.D.; Korolchuk, V.I.; Murray, J.T.; Zachari, M.; Ganley, I.G. The mammalian ULK1 complex and autophagy initiation. Essays Biochem. 2017, 61, 585–596. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, S.; Yoshimori, T. New insights into autophagosome–lysosome fusion. J. Cell Sci. 2017, 130, 1209–1216. [Google Scholar] [CrossRef] [Green Version]
- Ravikumar, B.; Moreau, K.; Jahreiss, L.; Puri, C.; Rubinsztein, D.C. Plasma membrane contributes to the formation of pre-autophagosomal structures. Nat. Cell Biol. 2010, 12, 747–757. [Google Scholar] [CrossRef]
- Bendich, A. Physiological role of antioxidants in the immune system. J. Dairy Sci. 1993, 76, 2789–2794. [Google Scholar] [CrossRef]
- Brambilla, D.; Mancuso, C.; Scuderi, M.R.; Bosco, P.; Cantarella, G.; Lempereur, L.; Di Benedetto, G.; Pezzino, S.; Bernardini, R. The role of antioxidant supplement in immune system, neoplastic, and neurodegenerative disorders: A point of view for an assessment of the risk/benefit profile. Nutr. J. 2008, 7, 29. [Google Scholar] [CrossRef]
- De la Fuente, M. Effects of antioxidants on immune system ageing. Eur. J. Clin. Nutr. 2002, 56, S5–S8. [Google Scholar] [CrossRef]
- Crimeen-Irwin, B.; Scalzo, K.; Gloster, S.; Mottram, P.; Plebanski, M. Failure of immune homeostasis-the consequences of under and over reactivity. Curr. Drug Targets Immune Endocr. Metabol. Disord. 2005, 5, 413–422. [Google Scholar] [CrossRef]
- Kuballa, P.; Nolte, W.M.; Castoreno, A.B.; Xavier, R.J. Autophagy and the immune system. Annu. Rev. Immunol. 2012, 30, 611–646. [Google Scholar] [CrossRef]
- Park, D.; Jeong, H.; Lee, M.N.; Koh, A.; Kwon, O.; Yang, Y.R.; Noh, J.; Suh, P.-G.; Park, H.; Ryu, S.H. Resveratrol induces autophagy by directly inhibiting mTOR through ATP competition. Sci. Rep. 2016, 6, 21772. [Google Scholar] [CrossRef] [Green Version]
- Rahman, M.A.; Bishayee, K.; Sadra, A.; Huh, S.-O. Oxyresveratrol activates parallel apoptotic and autophagic cell death pathways in neuroblastoma cells. Biochim. Biophys. Acta Gen. Subj. 2017, 1861, 23–36. [Google Scholar] [CrossRef]
- Burada, F.; Nicoli, E.R.; Ciurea, M.E.; Uscatu, D.C.; Ioana, M.; Gheonea, D.I. Autophagy in colorectal cancer: An important switch from physiology to pathology. World J. Gastrointest. Oncol. 2015, 7, 271–284. [Google Scholar] [CrossRef]
- Lamb, C.A.; Yoshimori, T.; Tooze, S.A. The autophagosome: Origins unknown, biogenesis complex. Nat. Rev. Mol. Cell Biol. 2013, 14, 759–774. [Google Scholar] [CrossRef]
- Tanida, I. Autophagosome formation and molecular mechanism of autophagy. Antioxid. Redox Signal. 2011, 14, 2201–2214. [Google Scholar] [CrossRef]
- Wirawan, E.; Lippens, S.; Vanden Berghe, T.; Romagnoli, A.; Fimia, G.M.; Piacentini, M.; Vandenabeele, P. Beclin1: A role in membrane dynamics and beyond. Autophagy 2012, 8, 6–17. [Google Scholar] [CrossRef] [Green Version]
- Shinojima, N.; Yokoyama, T.; Kondo, Y.; Kondo, S. Roles of the Akt/mTOR/p70S6K and ERK1/2 signaling pathways in curcumin-induced autophagy. Autophagy 2007, 3, 635–637. [Google Scholar] [CrossRef] [Green Version]
- Roux, P.P.; Blenis, J. ERK and p38 MAPK-activated protein kinases: A family of protein kinases with diverse biological functions. Microbiol. Mol. Biol. Rev. 2004, 68, 320–344. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Li, J.; Cao, N.; Li, Z.; Han, J.; Li, L. Resveratrol, an activator of SIRT1, induces protective autophagy in non-small-cell lung cancer via inhibiting Akt/mTOR and activating p38-MAPK. Onco Targets Ther. 2018, 11, 7777–7786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, K.-L.; Lin, K.-J.; Wang, P.-W.; Chuang, J.-H.; Lin, H.-Y.; Chen, S.-D.; Chuang, Y.-C.; Huang, S.-T.; Tiao, M.-M.; Chen, J.-B. Resveratrol provides neuroprotective effects through modulation of mitochondrial dynamics and ERK1/2 regulated autophagy. Free Radic. Res. 2018, 52, 1371–1386. [Google Scholar] [CrossRef] [PubMed]
- Codogno, P.; Meijer, A.J. Atg5: More than an autophagy factor. Nat. Cell Biol. 2006, 8, 1045–1047. [Google Scholar] [CrossRef]
- Subramani, S.; Malhotra, V. Non-autophagic roles of autophagy-related proteins. EMBO Rep. 2013, 14, 143–151. [Google Scholar] [CrossRef]
- Schönthal, A.H. Endoplasmic reticulum stress and autophagy as targets for cancer therapy. Cancer Lett. 2009, 275, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Hollien, J. Evolution of the unfolded protein response. Biochim. Biophys. Acta Mol. Cell Res. 2013, 1833, 2458–2463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Wey, S.; Zhang, Y.; Ye, R.; Lee, A.S. Role of the unfolded protein response regulator GRP78/BiP in development, cancer, and neurological disorders. Antioxid. Redox Signal. 2009, 11, 2307–2316. [Google Scholar] [CrossRef]
- So, J.-S. Roles of endoplasmic reticulum stress in immune responses. Mol. Cells 2018, 41, 705–716. [Google Scholar]
- Margariti, A.; Li, H.; Chen, T.; Martin, D.; Vizcay-Barrena, G.; Alam, S.; Karamariti, E.; Xiao, Q.; Zampetaki, A.; Zhang, Z. XBP1 mRNA splicing triggers an autophagic response in endothelial cells through BECLIN-1 transcriptional activation. J. Biol. Chem. 2013, 288, 859–872. [Google Scholar] [CrossRef] [Green Version]
- Engevik, M.A.; Luk, B.; Chang-Graham, A.L.; Hall, A.; Herrmann, B.; Ruan, W.; Endres, B.T.; Shi, Z.; Garey, K.W.; Hyser, J.M. Bifidobacterium dentium Fortifies the Intestinal Mucus Layer via Autophagy and Calcium Signaling Pathways. mBio 2019, 10, e01087-19. [Google Scholar] [CrossRef] [Green Version]
- Iida, T.; Yokoyama, Y.; Wagatsuma, K.; Hirayama, D.; Nakase, H. Impact of autophagy of innate immune cells on inflammatory bowel disease. Cells 2019, 8, 7. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward (5′ to 3′) | Reverse (5′ to 3′) |
|---|---|---|
| GAPDH | GAGTCAACGGATTTGGTCGT | GACAAGCTTCCCGTTCTCAG |
| ULK1 | CAGAACTACCAGCGCATTGA | TCCACCCAGAGACATCTTCC |
| ULK2 | GTGGGAACGTGAGGAAGAGG | CTGGGAGTTTCAGGGTGGTC |
| Atg13 | AAAGTCACCTCCCAGTGTGG | GAAAAGCTCCACAGGACAGC |
| Atg101 | GAAGTGTGGACGGTCAAGGT | CACGTTATCCACCTCCGACT |
| VPS34 | AAGCAGTGCCTGTAGGAGGA | TGTCGATGAGCTTTGGTGAG |
| Beclin-1 | AGGTTGAGAAAGGCGAGACA | AATTGTGAGGACACCCAAGC |
| Atg7 | GGCCTAGGTACCTGTGACCA | GAAGCTCCCATGAGCTGAAC |
| Atg4 | GGCTTACCAAGGGCTACCTC | ACAGCACTGGAAAGGGACAC |
| Atg5 | ACCAGAAACACTTCGCTGCT | GACCTTCAGTGGTCCGGTAA |
| Atg16L1 | CACAAGAAACGTGGGGAGTT | ACAAAGCTTAGTGCGCAGGT |
| LC3B | CCACACCCAAAGTCCTCACT | CACTGCTGCTTTCCGTAACA |
| Rab7 | TGGATGACAGGCTAGTCACG | CTGGCCTGGATGAGAAACTC |
| STX17 | GGGTGAAGCCAGGAATGTTA | ATGCCACACCCAGCTAATTC |
| VAMP8 | CATCTCCGCAACAAGACAGA | GACCCTCTTGGCACACATTT |
| LAMP2 | ACAACTCACTCCACAGGCAG | TGCAATGCTGAAAACGGAGC |
| MEK | GCTTGGGGCTATTTGTGTGT | TCTCACAAGGCTCCCTCCTA |
| ERK1 | TCA GAC TCC AAA GCC CTT GAC | CGT GCT GTC TCC TGG AAG ATG |
| ERK2 | TCC AAC AGG CCC ATC TTT CC | CCA GAG CTT TGG AGT CAG CA |
| IRE1α | CGGCCTTTGCAGATAGTCTC | ACGTCCCCAGATTCACTGTC |
| XBP1 | GGAGTTAAGACAGCGCTTGG | ACTGGGTCCAAGTTGTCCAG |
| PERK | CTCACAGGCAAAGGAAGGAG | AACAACTCCAAAGCCACCAC |
| ATF4 | TCAAACCTCATGGGTTCTCC | GAAGGTCATCTGGCATGGTT |
| MUC2 | ACCCGCACTATGTCACCTTC | GGACAGGACACCTTGTCGTT |
| Protein | Protein Identification | Fold Change |
|---|---|---|
| EIF5A | eukaryotic translation initiation factor 5A-1 isoform B | 9.94 |
| PHB | prohibitin | 0.003 |
| AP1B1 | AP-1 complex subunit beta-1 isoform b | 19.206 |
| GANAB | glucosidase, alpha; neutral AB, isoform CRA_a | 0.059 |
| HSPA5 | GRP78, endoplasmic reticulum chaperone BiP precursor | 3.42 |
| IQGAP1 | IQGAP1 protein | 3.421 |
| ACTN4 | alpha-actinin-4 isoform 1 | 3.994 |
| PDCD6IP | programmed cell death 6-interacting protein isoform 2 | 0.096 |
| HSP90AB1 | heat shock protein HSP 90-beta isoform c | 3.464 |
| HSP90B1 | tumor rejection antigen (gp96) 1 | 3.081 |
| HSP90AA1 | HSP90AA1 protein | 2.469 |
| AHSA1 | activator of 90kDa heat shock protein ATPase homolog 1 isoform1 | 0.149 |
| AARS | alanyl-tRNA synthetase variant | 2.356 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yeom, J.; Ma, S.; Lim, Y.-H. Oxyresveratrol Induces Autophagy via the ER Stress Signaling Pathway, and Oxyresveratrol-Induced Autophagy Stimulates MUC2 Synthesis in Human Goblet Cells. Antioxidants 2020, 9, 214. https://doi.org/10.3390/antiox9030214
Yeom J, Ma S, Lim Y-H. Oxyresveratrol Induces Autophagy via the ER Stress Signaling Pathway, and Oxyresveratrol-Induced Autophagy Stimulates MUC2 Synthesis in Human Goblet Cells. Antioxidants. 2020; 9(3):214. https://doi.org/10.3390/antiox9030214
Chicago/Turabian StyleYeom, Jiah, Seongho Ma, and Young-Hee Lim. 2020. "Oxyresveratrol Induces Autophagy via the ER Stress Signaling Pathway, and Oxyresveratrol-Induced Autophagy Stimulates MUC2 Synthesis in Human Goblet Cells" Antioxidants 9, no. 3: 214. https://doi.org/10.3390/antiox9030214