
The Balancing of Peroxynitrite Detoxification between Ferric Heme-Proteins and CO2: The Case of Zebrafish Nitrobindin
 ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Methods
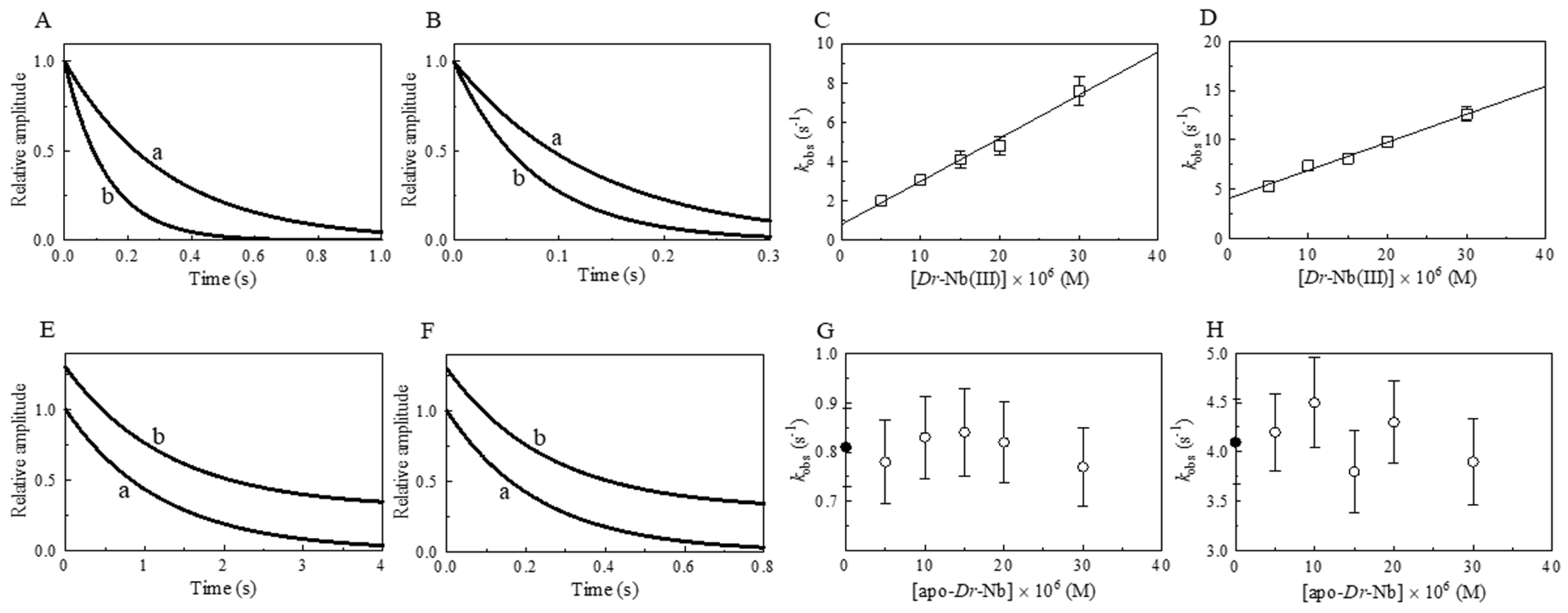
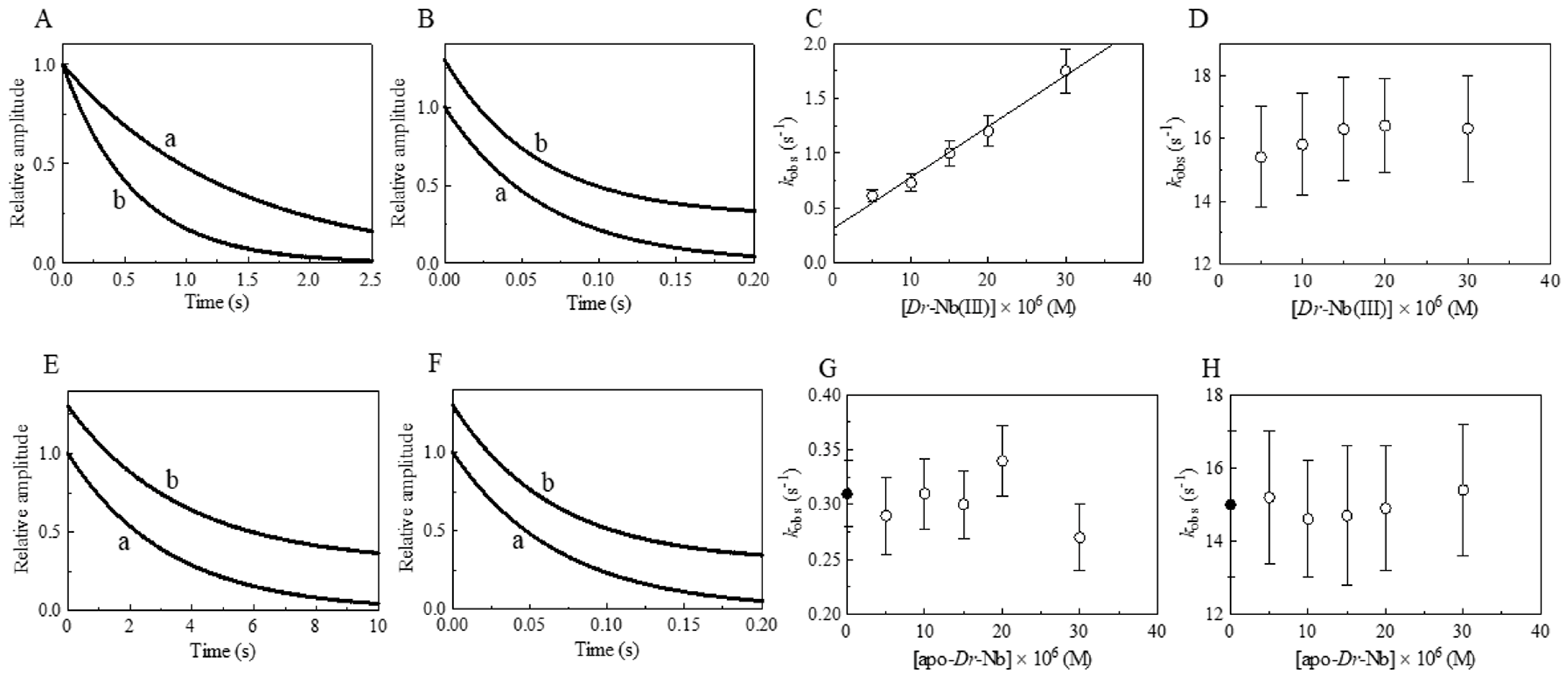
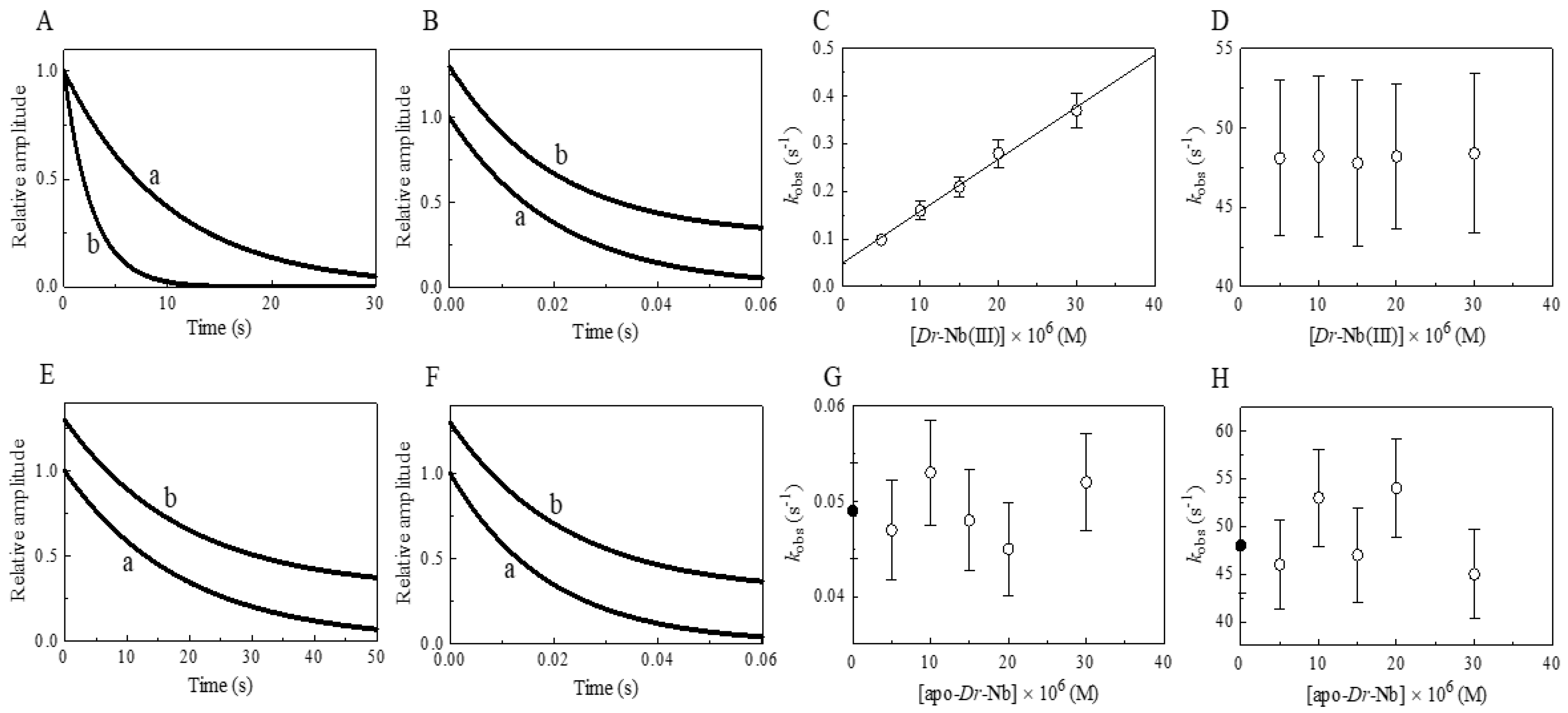
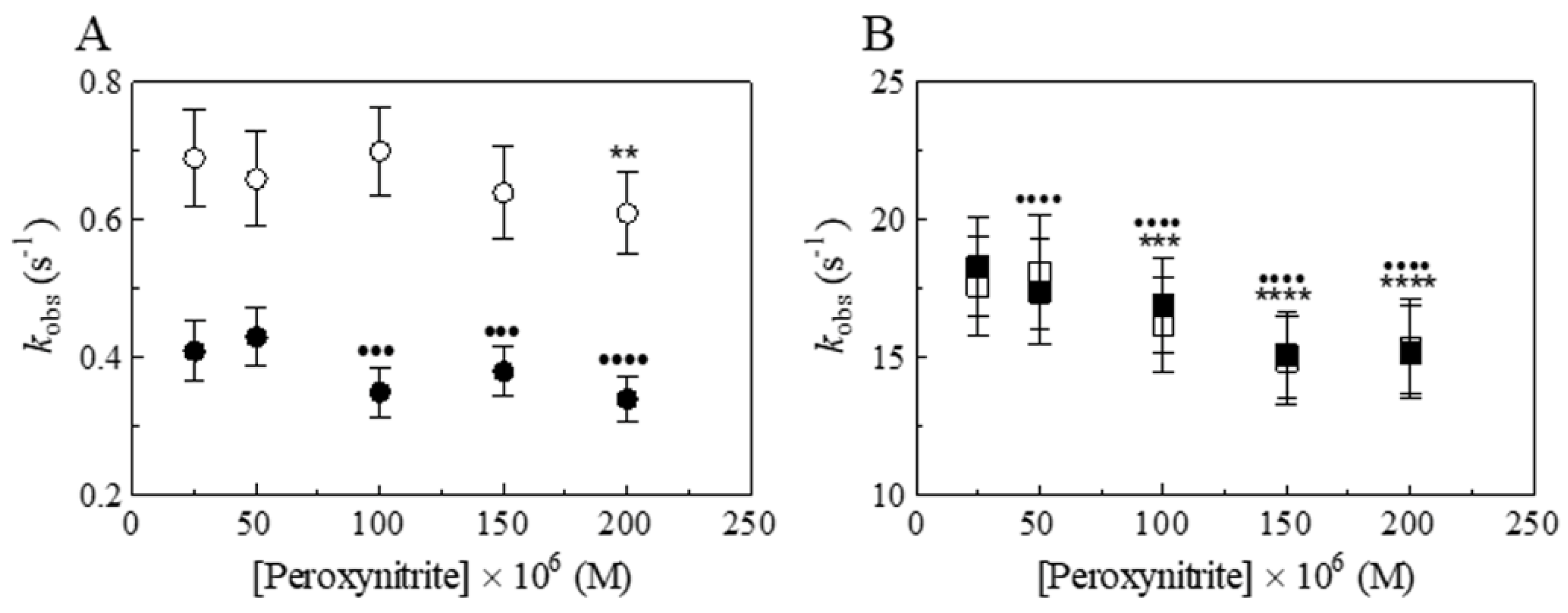
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cantó, A.; Olivar, T.; Romero, F.J.; Miranda, M. Nitrosative Stress in Retinal Pathologies: Review. Antioxidants 2019, 8, 543. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, A.; Ahsan, H. Biomarkers of inflammation and oxidative stress in ophthalmic disorders. J. Immunoass. Immunochem. 2020, 41, 257–271. [Google Scholar] [CrossRef] [PubMed]
- Lei, Y.; Gao, Y.; Song, M.; Cao, W.; Sun, X. Peroxynitrite is a novel risk factor and treatment target of glaucoma. Nitric Oxide 2020, 99, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Beckman, J.S.; Koppenol, W.H. Nitric oxide, superoxide, and peroxynitrite: The good, the bad, and ugly. Am. J. Physiol. 1996, 271, C1424–C1437. [Google Scholar] [CrossRef] [PubMed]
- Ducrocq, C.; Blanchard, B.; Pignatelli, B.; Ohshima, H. Peroxynitrite: An endogenous oxidizing and nitrating agent. Cell. Mol. Life Sci. 1999, 55, 1068–1077. [Google Scholar] [CrossRef] [PubMed]
- Clementi, E.; Nisoli, E. Nitric oxide and mitochondrial biogenesis: A key to long-term regulation of cellular metabolism. Comp. Biochem. Physiol. 2005, 142, 102–110. [Google Scholar] [CrossRef]
- DeNicola, A.; Radi, R. Peroxynitrite and drug-dependent toxicity. Toxicology 2005, 208, 273–288. [Google Scholar] [CrossRef]
- Pacher, P.; Beckman, J.S.; Liaudet, L. Nitric Oxide and Peroxynitrite in Health and Disease. Physiol. Rev. 2007, 87, 315–424. [Google Scholar] [CrossRef]
- Goldstein, S.; Merényi, G. The Chemistry of Peroxynitrite: Implications for Biological Activity. Methods Enzymol. 2008, 436, 49–61. [Google Scholar] [CrossRef]
- Jones, L.H. Chemistry and Biology of Biomolecule Nitration. Chem. Biol. 2012, 19, 1086–1092. [Google Scholar] [CrossRef] [Green Version]
- Radi, R. Oxygen radicals, nitric oxide, and peroxynitrite: Redox pathways in molecular medicine. Proc. Natl. Acad. Sci. USA 2018, 115, 5839–5848. [Google Scholar] [CrossRef]
- Goldstein, S.; Lind, J.; Merényi, G. Chemistry of peroxynitrites and peroxynitrates. Chem. Rev. 2005, 105, 2457–2470. [Google Scholar] [CrossRef]
- Ascenzi, P.; Leboffe, L.; Santucci, R.; Coletta, M. Ferric microperoxidase-11 catalyzes peroxynitrite isomerization. J. Inorg. Biochem. 2015, 144, 56–61. [Google Scholar] [CrossRef]
- Lymar, S.V.; Hurst, J.K. Rapid reaction between peroxynitrite ion and carbon dioxide: Implications for biological activity. J. Am. Chem. Soc. 1995, 117, 8867–8868. [Google Scholar] [CrossRef]
- DeNicola, A.; Freeman, B.A.; Trujillo, M.; Radi, R. Peroxynitrite Reaction with Carbon Dioxide/Bicarbonate: Kinetics and Influence on Peroxynitrite-Mediated Oxidations. Arch. Biochem. Biophys. 1996, 333, 49–58. [Google Scholar] [CrossRef]
- Bonini, M.G.; Radi, R.; Ferrer-Sueta, G.; Ferreira, A.M.D.C.; Augusto, O. Direct EPR Detection of the Carbonate Radical Anion Produced from Peroxynitrite and Carbon Dioxide. J. Biol. Chem. 1999, 274, 10802–10806. [Google Scholar] [CrossRef]
- Radi, R.; Denicola, A.; Freeman, B.A. Peroxynitrite reactions with carbon dioxide-bicarbonate. Methods Enzymol. 1999, 301, 353–367. [Google Scholar] [CrossRef]
- Meli, R.; Nauser, T.; Latal, P.; Koppenol, W.H. Reaction of peroxynitrite with carbon dioxide: Intermediates and determination of the yield of CO3− and NO2●. J. Biol. Inorg. Chem. 2002, 7, 31–36. [Google Scholar] [CrossRef]
- Squadrito, G.L.; Pryor, W.A. Mapping the reaction of peroxynitrite with CO2: Energetics, reactive species, and biological im-plications. Chem. Res. Toxicol. 2002, 15, 885–895. [Google Scholar] [CrossRef]
- Augusto, O.; Goldstein, S.; Hurst, J.K.; Lind, J.; Lymar, S.V.; Merenyi, G.; Radi, R. Carbon dioxide-catalyzed peroxynitrite reactivity—The resilience of the radical mechanism after two decades of research. Free Radic. Biol. Med. 2019, 135, 210–215. [Google Scholar] [CrossRef]
- Augusto, O.; Truzzi, D.R. Carbon dioxide redox metabolites in oxidative eustress and oxidative distress. Biophys. Rev. 2021, 13, 889–891. [Google Scholar] [CrossRef]
- Mehl, M.; Daiber, A.; Herold, S.; Shoun, H.; Ullrich, V. Peroxynitrite reaction with heme proteins. Nitric Oxide 1999, 3, 142–152. [Google Scholar] [CrossRef]
- Shimanovich, R.; Groves, J.T. Mechanisms of Peroxynitrite Decomposition Catalyzed by FeTMPS, a Bioactive Sulfonated Iron Porphyrin. Arch. Biochem. Biophys. 2001, 387, 307–317. [Google Scholar] [CrossRef] [PubMed]
- Jensen, M.P.; Riley, D.P. Peroxynitrite Decomposition Activity of Iron Porphyrin Complexes. Inorg. Chem. 2002, 41, 4788–4797. [Google Scholar] [CrossRef] [PubMed]
- Herold, S.; Shivashankar, K. Metmyoglobin and Methemoglobin Catalyze the Isomerization of Peroxynitrite to Nitrate. Biochemistry 2003, 42, 14036–14046. [Google Scholar] [CrossRef] [PubMed]
- Herold, S.; Fago, A.; Weber, R.E.; Dewilde, S.; Moens, L. Reactivity studies of the Fe(III) and Fe(II)NO forms of human neu-roglobin reveal a potential role against oxidative stress. J. Biol. Chem. 2004, 279, 22841–22847. [Google Scholar] [CrossRef] [PubMed]
- Herold, S.; Kalinga, S.; Matsui, A.T.; Watanabe, Y. Mechanistic Studies of the Isomerization of Peroxynitrite to Nitrate Catalyzed by Distal Histidine Metmyoglobin Mutants. J. Am. Chem. Soc. 2004, 126, 6945–6955. [Google Scholar] [CrossRef]
- Herold, S.; Puppo, A. Kinetics and mechanistic studies of the reactions of metleghemoglobin, ferrylleghemoglobin, and nitrosylleghemoglobin with reactive nitrogen species. J. Biol. Inorg. Chem. 2005, 10, 946–957. [Google Scholar] [CrossRef]
- Ascenzi, P.; Visca, P. Scavenging of Reactive Nitrogen Species by Mycobacterial Truncated Hemoglobins. Methods Enzymol. 2008, 436, 317–337. [Google Scholar] [CrossRef]
- Ascenzi, P.; di Masi, A.; Coletta, M.; Ciaccio, C.; Fanali, G.; Nicoletti, F.P.; Smulevich, G.; Fasano, M. Ibuprofen impairs allo-sterically peroxynitrite isomerization by ferric human serum heme-albumin. J. Biol. Chem. 2009, 284, 31006–31017. [Google Scholar] [CrossRef] [Green Version]
- Ascenzi, P.; Bolli, A.; Di Masi, A.; Tundo, G.R.; Fanali, G.; Coletta, M.; Fasano, M. Isoniazid and rifampicin inhibit allosterically heme binding to albumin and peroxynitrite isomerization by heme–albumin. J. Biol. Inorg. Chem. 2010, 16, 97–108. [Google Scholar] [CrossRef]
- Ascenzi, P.; Ciaccio, C.; Sinibaldi, F.; Santucci, R.; Coletta, M. Cardiolipin modulates allosterically peroxynitrite detoxification by horse heart cytochrome c. Biochem. Biophys. Res. Commun. 2011, 404, 190–194. [Google Scholar] [CrossRef]
- Ascenzi, P.; Ciaccio, C.; Sinibaldi, F.; Santucci, R.; Coletta, M. Peroxynitrite detoxification by horse heart carboxymethylated cytochrome c is allosterically modulated by cardiolipin. Biochem. Biophys. Res. Commun. 2011, 415, 463–467. [Google Scholar] [CrossRef]
- Ascenzi, P.; Coletta, A.; Cao, Y.; Trezza, V.; Leboffe, L.; Fanali, G.; Fasano, M.; Pesce, A.; Ciaccio, C.; Marini, S.; et al. Isoniazid Inhibits the Heme-Based Reactivity of Mycobacterium tuberculosis Truncated Hemoglobin N. PLoS ONE 2013, 8, e69762. [Google Scholar] [CrossRef]
- Coppola, D.; Giordano, D.; Tinajero-Trejo, M.; di Prisco, G.; Ascenzi, P.; Poole, R.K.; Verde, C. Antarctic bacterial haemoglobin and its role in the protection against nitrogen reactive species. Biochim. Biophys. Acta 2013, 1834, 1923–1931. [Google Scholar] [CrossRef]
- Ascenzi, P.; Pesce, A. Peroxynitrite scavenging by Campylobacter jejuni truncated hemoglobin P. J. Biol. Inorg. Chem. 2017, 22, 1141–1150. [Google Scholar] [CrossRef]
- Ascenzi, P.; Coletta, M. Peroxynitrite Detoxification by Human Haptoglobin:Hemoglobin Complexes: A Comparative Study. J. Phys. Chem. B 2018, 122, 11100–11107. [Google Scholar] [CrossRef]
- Coppola, D.; Giordano, D.; Milazzo, L.; Howes, B.D.; Ascenzi, P.; di Prisco, G.; Smulevich, G.; Poole, R.K.; Verde, C. Coex-istence of multiple globin genes conferring protection against nitrosative stress to the Antarctic bacterium Pseudoalteromonas haloplanktis TAC125. Nitric Oxide 2018, 73, 39–51. [Google Scholar] [CrossRef]
- De Simone, G.; di Masi, A.; Polticelli, F.; Ascenzi, P. Human nitrobindin: The first example of an all-β-barrel ferric heme-protein that catalyzes peroxynitrite detoxification. FEBS Open Bio 2018, 8, 2002–2010. [Google Scholar] [CrossRef]
- De Simone, G.; Di Masi, A.; Vita, G.M.; Polticelli, F.; Pesce, A.; Nardini, M.; Bolognesi, M.; Ciaccio, C.; Coletta, M.; Turilli, E.S.; et al. Mycobacterial and Human Nitrobindins: Structure and Function. Antioxidants Redox Signal. 2020, 33, 229–246. [Google Scholar] [CrossRef]
- Giordano, D.; Pesce, A.; Vermeylen, S.; Abbruzzetti, S.; Nardini, M.; Marchesani, F.; Berghmans, H.; Seira, C.; Bruno, S.; Luque, F.J.; et al. Structural and functional properties of Antarctic fish cytoglobins-1: Cold-reactivity in multi-ligand reactions. Comput. Struct. Biotechnol. J. 2020, 18, 2132–2144. [Google Scholar] [CrossRef] [PubMed]
- Radi, R.; Beckman, J.S.; Bush, K.M.; Freeman, B.A. Peroxynitrite oxidation of sulfhydryls. The cytotoxic potential of superoxide and nitric oxide. J. Biol. Chem. 1991, 266, 4244–4250. [Google Scholar] [CrossRef]
- Gow, A.; Duran, D.; Thom, S.R.; Ischiropoulos, H. Carbon dioxide enhancement of peroxynitrite mediated protein tyrosine nitration. Arch. Biochem. Biophys. 1996, 333, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Scorza, G.; Minetti, M. One electron oxidation pathway of thyols by peroxynitrite in biological fluids: Bicarbonate and ascorbate promote the formation of albumin disulphide dimers in human blood plasma. Biochem. J. 1998, 329, 405–413. [Google Scholar] [CrossRef]
- Bonini, M.G.; Augusto, O. Carbon Dioxide Stimulates the Production of Thiyl, Sulfinyl, and Disulfide Radical Anion from Thiol Oxidation by Peroxynitrite. J. Biol. Chem. 2001, 276, 9749–9754. [Google Scholar] [CrossRef]
- Augusto, O.; Bonini, M.G.; Amanso, A.M.; Linares, E.; Santos, C.C.; De Menezes, S.L. Nitrogen dioxide and carbonate radical anion: Two emerging radicals in biology. Free Radic. Biol. Med. 2002, 32, 841–859. [Google Scholar] [CrossRef]
- Elks, P.M.; van der Vaart, M.; van Hensbergen, V.; Schutz, E.; Redd, M.J.; Murayama, E.; Spaink, H.P.; Meijer, A.H. Myco-bacteria counteract a TLR-mediated nitrosative defense mechanism in a zebrafish infection model. PLoS ONE 2014, 9, e100928. [Google Scholar] [CrossRef]
- Shahab, M.; Rosati, R.; Meyer, D.N.; Shields, J.N.; Crofts, E.; Baker, T.R.; Jamesdaniel, S. Cisplatin-induced hair cell loss in zebrafish neuromasts is accompanied by protein nitration and Lmo4 degradation. Toxicol. Appl. Pharmacol. 2020, 410, 115342. [Google Scholar] [CrossRef]
- Vossen, L.E.; Jutfelt, F.; Cocco, A.; Thörnqvist, P.-O.; Winberg, S. Zebrafish (Danio rerio) behavior is largely unaffected by el-evated pCO2. Conserv. Physiol. 2016, 4, cow065. [Google Scholar] [CrossRef]
- Damsgaard, C. Physiology and evolution of oxygen secreting mechanism in the fisheye. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2020, 252, 110840. [Google Scholar] [CrossRef]
- Tummanapalli, S.S.; Kuppusamy, R.; Yeo, J.H.; Kumar, N.; New, E.J.; Willcox, M.D. The role of nitric oxide in ocular surface physiology and pathophysiology. Ocul. Surf. 2021, 21, 37–51. [Google Scholar] [CrossRef]
- De Simone, G.; Tundo, G.R.; Coletta, A.; Coletta, M.; Ascenzi, P. Hydroxylamine-induced oxidation of ferrous nitrobindins. J. Biol. Inorg. Chem. 2022, 27, 443–453. [Google Scholar] [CrossRef]
- Antonini, E.; Brunori, M. Hemoglobin and Myoglobin in Their Reactions with Ligands; North Holland Publishing Co.: Amsterdam, The Netherlands; London, UK, 1971. [Google Scholar]
- Herold, S.; Exner, M.; Boccini, F. The Mechanism of the Peroxynitrite-Mediated Oxidation of Myoglobin in the Absence and Presence of Carbon Dioxide. Chem. Res. Toxicol. 2003, 16, 390–402. [Google Scholar] [CrossRef]
- Ascenzi, P.; Leboffe, L.; Pesce, A.; Ciaccio, C.; Sbardella, D.; Bolognesi, M.; Coletta, M. Nitrite-Reductase and Peroxynitrite Isomerization Activities of Methanosarcina acetivorans Protoglobin. PLoS ONE 2014, 9, e95391. [Google Scholar] [CrossRef]
- Miranda, K.M.; Espey, M.G.; Wink, D.A. A Rapid, Simple Spectrophotometric Method for Simultaneous Detection of Nitrate and Nitrite. Nitric Oxide 2001, 5, 62–71. [Google Scholar] [CrossRef]
- Perutz, M.F. Regulation of Oxygen Affinity of Hemoglobin: Influence of Structure of the Globin on the Heme Iron. Annu. Rev. Biochem. 1979, 48, 327–386. [Google Scholar] [CrossRef]
- Bolognesi, M.; Bordo, D.; Rizzi, M.; Tarricone, C.; Ascenzi, P. Nonvertebrate hemoglobins: Structural bases for reactivity. Prog. Biophys. Mol. Biol. 1997, 68, 29–68. [Google Scholar] [CrossRef]
- Bianchetti, C.M.; Blouin, G.C.; Bitto, E.; Olson, J.S.; Phillips, G.N., Jr. The structure and NO binding properties of the nitro-phorin-like heme-binding protein from Arabidopsis thaliana gene locus At1 g79260.1. Proteins 2010, 78, 917–931. [Google Scholar] [CrossRef]
- Bianchetti, C.M.; Bingman, C.A.; Phillips, G.N., Jr. Structure of the C-terminal heme-binding domain of THAP domain con-taining protein 4 from Homo sapiens. Proteins 2011, 79, 1337–1341. [Google Scholar] [CrossRef]
- Ascenzi, P.; Brunori, M. A molecule for all seasons: The heme. J. Porphyrins Phthalocyanines 2016, 20, 134–149. [Google Scholar] [CrossRef]
- Kissner, R.; Koppenol, W.H. Product Distribution of Peroxynitrite Decay as a Function of pH, Temperature, and Concentration. J. Am. Chem. Soc. 2001, 124, 234–239. [Google Scholar] [CrossRef] [PubMed]
- Ascenzi, P.; Bocedi, A.; Visca, P.; Minetti, M.; Clementi, E. Does CO2 modulate peroxynitrite specificity? IUBMB Life 2006, 58, 611–613. [Google Scholar] [CrossRef] [PubMed]
- Brunori, M. Molecular Adaptation to Physiological Requirements: The Hemoglobin System of Trout. Curr. Top. Cell. Regul. 1975, 9, 1–39. [Google Scholar] [CrossRef] [PubMed]
- Brunori, M.; Coletta, M.; Giardina, B.; Wyman, J. A macromolecular transducer as illustrated by trout hemoglobin IV. Proc. Natl. Acad. Sci. USA 1978, 75, 4310–4312. [Google Scholar] [CrossRef]
- Ciaccio, C.; Coletta, A.; Coletta, M. Role of hemoglobin structural-functional relationships in oxygen transport. Mol. Asp. Med. 2021, 84, 101022. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
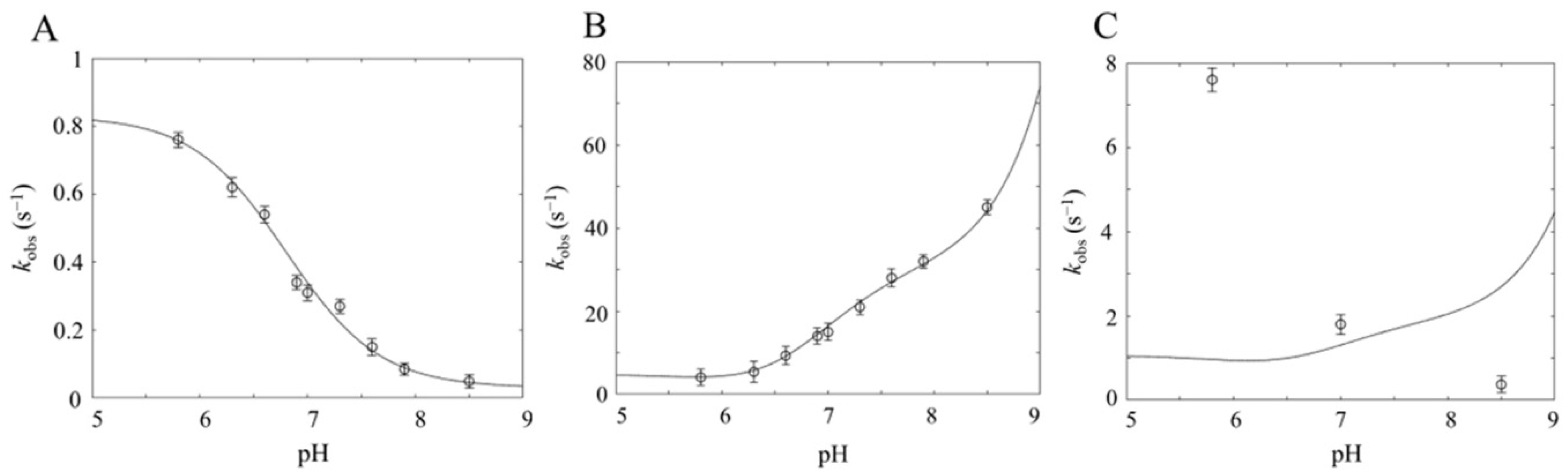{kind=link}
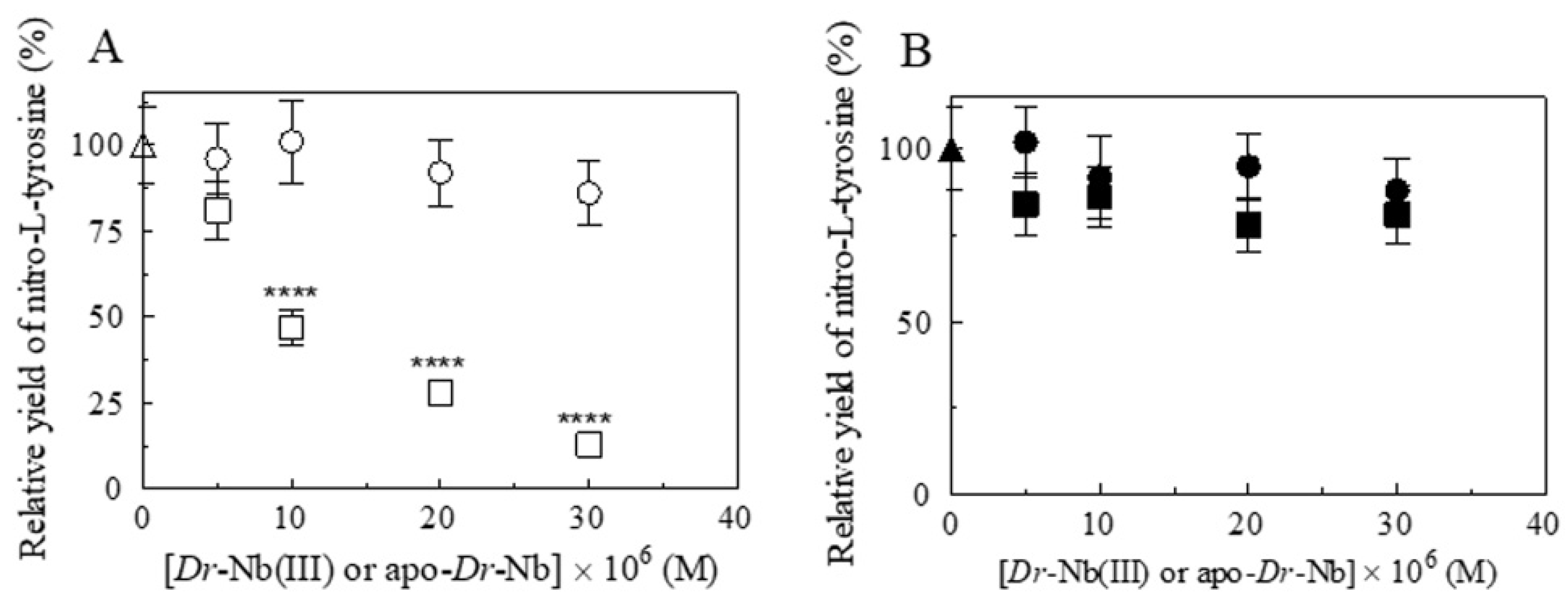{kind=link}
{kind=link}
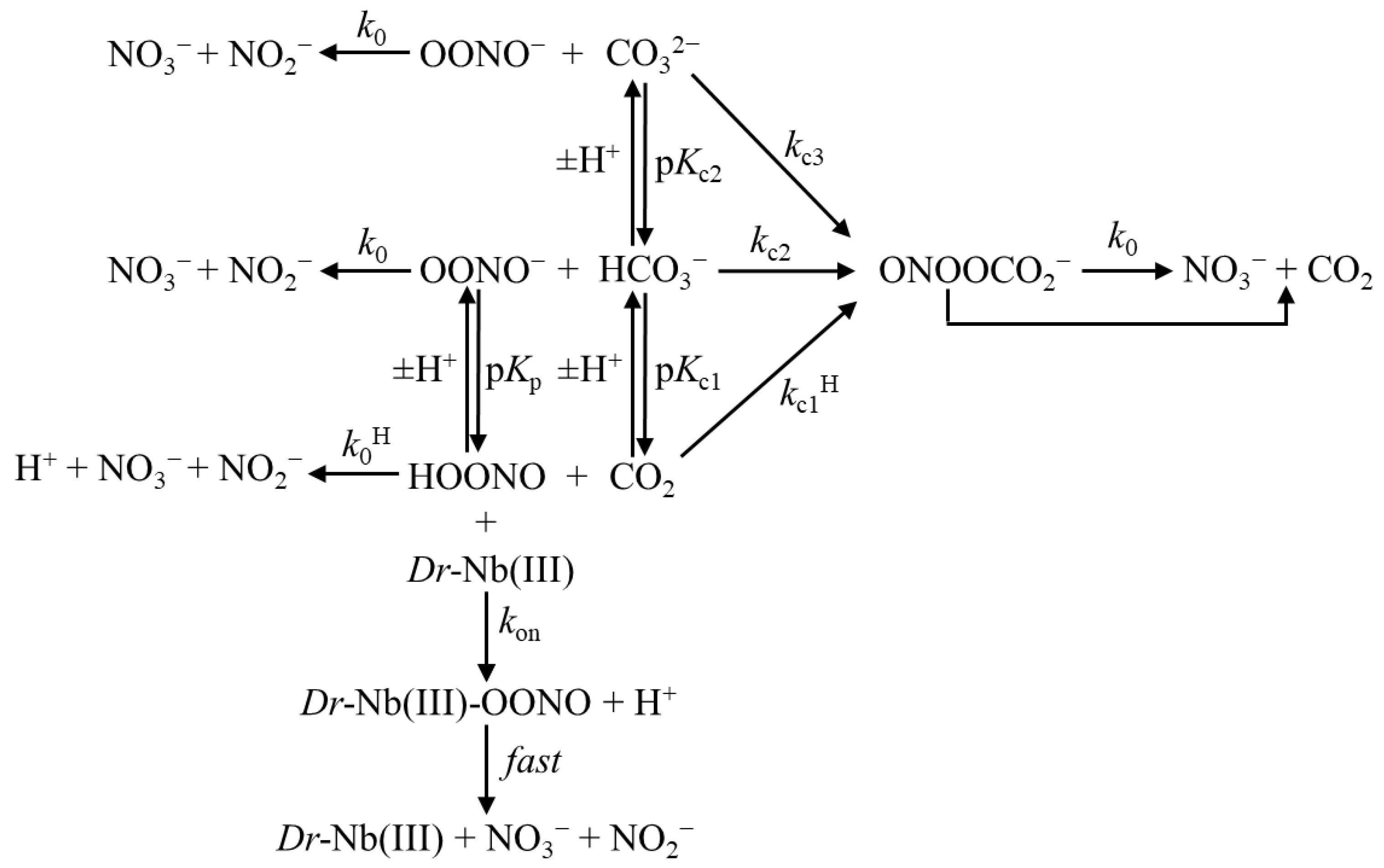{kind=link}
| pH | − CO2 | + CO2 | ||
|---|---|---|---|---|
 |  | |||
| kon (M−1 s−1) | k0 (s−1) | kon (M−1 s−1) | k0 (s−1) | |
| 5.8 | (2.2 ± 0.3) × 105 | (8.1 ± 0.8) × 10−1 | (2.8 ± 0.3) × 105 | 4.1 ± 0.4 |
| 7.0 | (4.7 ± 0.5) × 104 | (3.1 ± 0.3) × 10−1 | n.d. | (1.5 ± 0.2) × 101 |
| 8.5 | (1.1 ± 0.1) × 104 | (4.9 ± 0.5) × 10−2 | n.d. | (4.8 ± 0.5) × 101 |
| Heme-Protein | − CO2 | + CO2 | ||
|---|---|---|---|---|
 |  | |||
| kon (M−1 s−1) | k0 (s−1) | kon (M−1 s−1) | k0 (s−1) | |
| Ma-Pgb(III) a | 3.8 × 104 | 2.8 × 10−1 | n.d. | n.d. |
| Mt-trHbN(III) b | 6.2 × 104 | 2.7 × 10−1 | n.d. | n.d. |
| Ph-trHbO(III) c | 2.9 × 104 | 2.8 × 10−1 | n.d. | n.d. |
| Cj-trHbP(III) d | 9.6 × 105 | 3.0 × 10−1 | 8.8 × 105 | 2.1 × 101 |
| Efc-Mb(III) e | 2.9 × 104 | 3.5 × 10−1 | 7.7 × 104 | 1.7 × 101 |
| Pc-Mb(III) f | 1.6 × 104 | n.d. | n.d. | n.d. |
| Hs-Hb(III) e | 1.2 × 104 | 3.0 × 10−1 | 3.9 × 104 | 1.7 × 101 |
| Mt-Nb(III) g | 6.9 × 104 | 2.6 × 10−1 | n.d. | n.d. |
| At-Nb(III) g | 3.7 × 104 | 3.0 × 10−1 | n.d. | n.d. |
| Dr-Nb(III) h | 4.7 × 104 | 3.1 × 10−1 | n.d. | 1.5 × 101 |
| Hs-Nb(III) i | 3.4 × 104 | 2.6 × 10−1 | n.d. | n.d. |
| Parameter | Value |
|---|---|
| k0 (s−1) | (3.0 ± 0.5) × 10−2 |
| k0H (s−1) | (8.3 ± 1.0) × 10−1 |
| KP (M) | (1.6 ± 0.3) × 10−7 |
| Kc1 (M) | (4.6 ± 0.9) × 10−7 |
| Kc2 (M) | (5.6 ± 1.2) × 10−11 |
| kc2 (M−1s−1) | (6.2 ± 1.3) × 101 |
| kc3 (M−1s−1) | (1.7 ± 0.4) × 103 |
| kc1H (M−1s−1) | 8.0 ± 2.1 |
| Dr-Nb(III) | Apo-Dr-Nb | CO2 | NO3− | NO2− |
|---|---|---|---|---|
| (M) | (M) | (M) | (%) | (%) |
| 0.0 | 0.0 | 0.0 | 73 ± 8 | 28 ± 3 |
| 0.0 | 0.0 | 1.2 × 10−3 | 69 ± 7 | 30 ± 3 |
| 0.0 | 3.5 × 10−5 | 0.0 | 71 ± 8 | 29 ± 3 |
| 0.0 | 3.5 × 10−5 | 1.2 × 10−3 | 74 ± 7 | 25 ± 2 |
| 3.5 × 10−5 | 0.0 | 0.0 | 89 ± 9 | 12 ± 2 |
| 3.5 × 10−5 | 0.0 | 1.2 × 10−3 | 93 ± 8 | 7 ± 1 |
| 3.5 × 10−5 | 3.5 × 10−5 | 0.0 | 91 ± 9 | 10 ± 1 |
| 3.5 × 10−5 | 3.5 × 10−5 | 1.2 × 10−3 | 92 ± 9 | 8 ± 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Simone, G.; Coletta, A.; di Masi, A.; Coletta, M.; Ascenzi, P. The Balancing of Peroxynitrite Detoxification between Ferric Heme-Proteins and CO2: The Case of Zebrafish Nitrobindin. Antioxidants 2022, 11, 1932. https://doi.org/10.3390/antiox11101932
De Simone G, Coletta A, di Masi A, Coletta M, Ascenzi P. The Balancing of Peroxynitrite Detoxification between Ferric Heme-Proteins and CO2: The Case of Zebrafish Nitrobindin. Antioxidants. 2022; 11(10):1932. https://doi.org/10.3390/antiox11101932
Chicago/Turabian StyleDe Simone, Giovanna, Andrea Coletta, Alessandra di Masi, Massimo Coletta, and Paolo Ascenzi. 2022. "The Balancing of Peroxynitrite Detoxification between Ferric Heme-Proteins and CO2: The Case of Zebrafish Nitrobindin" Antioxidants 11, no. 10: 1932. https://doi.org/10.3390/antiox11101932