1. Introduction
Obesity and metabolic syndrome (MetS) are becoming a global epidemic worldwide, and their prevalence has dramatically increased during the last 15 years in both children and adults [
1]. MetS is composed of a variable cluster of cardiometabolic risk factors and comorbidities, such as abdominal obesity, reduced HDL and elevated LDL cholesterol, elevated triglycerides, glucose intolerance, and hypertension, that portend high risk of both cardiovascular disease and type 2 diabetes. It is responsible for huge socio-economic costs, resulting morbidity and mortality in most countries [
1,
2]. Indeed, the different components of MetS have been linked, individually or combined, with a higher prevalence of sudden cardiac death (SCD) [
3], but the exact underlying mechanisms remain poorly understood.
Heart rate responds dynamically to physiologic and pathologic perturbations. This cyclic fluctuation of RR intervals can be analyzed using heart rate variability (HRV), which can be used as a quantitative marker of autonomic activity [
4]. One of the proposed mechanisms of cardiac arrhythmia leading to SCD is an abnormal modulation of autonomic activity. Indeed, experimental and clinical evidence has found an association between propensity for lethal arrhythmias and signs of either increased sympathetic/parasympathetic activity or reduced vagal activity [
5,
6,
7].
Recently, we developed a diet-induced experimental model of MetS in New Zealand White (NZW) rabbits, which reproduced the main components of human MetS, namely, central obesity, a state of prediabetes characterized by impaired fasting glucose and glucose intolerance, mild hypertension, and alterations in the lipid profile (increased triglycerides and LDL, decreased HDL and no changes in total cholesterol) [
8]. Our aim was to investigate the time-course modifications of short-term HRV in an experimental rabbit model of diet-induced MetS during its development in order to identify the early changes in HRV and its potential relationship with cardiac arrhythmias and SCD.
4. Discussion
This study includes a comprehensive analysis of HRV alterations, which is a non-invasive measure of cardiac autonomic function, found in a diet-induced model of MetS using short-term ECG recordings. The analysis of HRV is a very useful and inexpensive tool that enables the analysis of heart rate dynamics and the detection of any early stage indication of cardiac dysfunction. In our study, we used short-term (15-min) recordings to analyze HRV during the course of MetS development using high-fat, high-sucrose diet in NZW rabbits. The advantages of short term HRV analysis include easier usability and analysis, the ability to collect data in a very controlled environment, and better suitability for spectral analysis that require stationary conditions [
5,
14]. In our study, using conventional HRV analyses, we found significant alterations in heart rate dynamics including time-domain, frequency-domain and non-linear domain modifications.
We analyzed the time-domain parameters of HRV before diet administration, at week 14 and at week 28. We found a decrease in the SDHR at week 28 in the MetS group, which is in agreement with other clinical studies that used short-time recordings in patients diagnosed with MetS [
15,
16,
17], without apparent modifications in other standard time-domain parameters. Previous studies found that SDNN was consistently reduced when one or more risk factors were present as compared to none or smaller number of MetS components, however, the best discrimination of MetS was provided by non-linear complexity and irregularity analyses of the RR series in agreement with our results [
4,
18,
19].
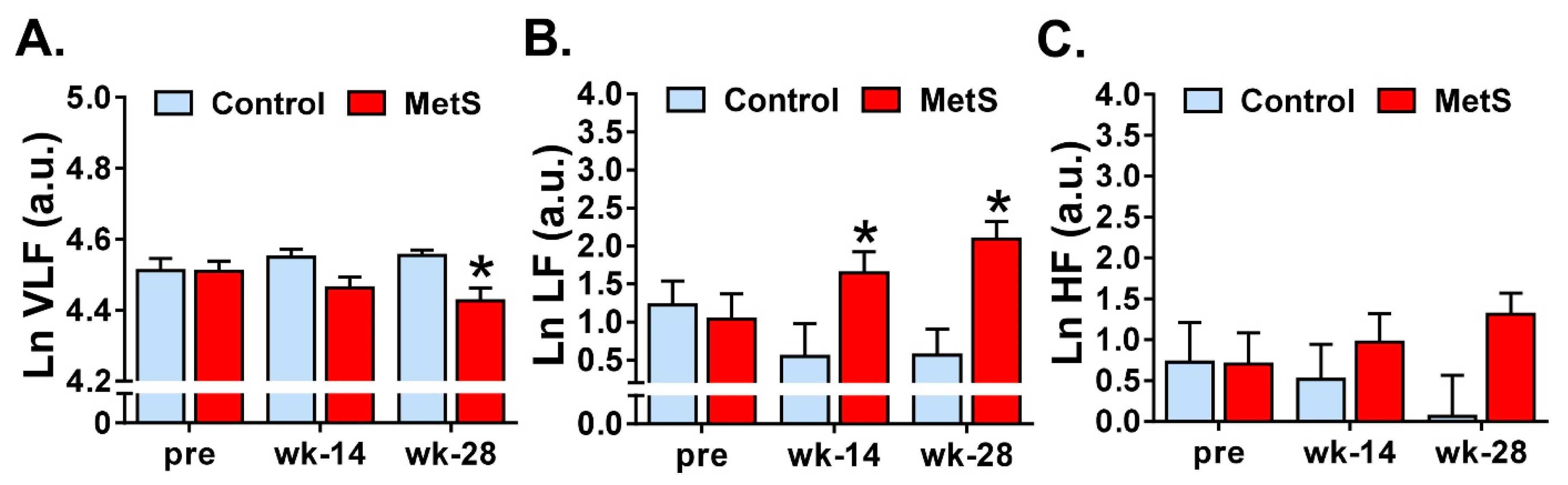
Regarding frequency-domain analysis, several changes were found in the lower frequency bands in the MetS group: a decrease in LnVLF at week 28 and an increase in LnLF at both week 14 and week 28. Regarding the former, the results in our experimental model are comparable to those found in the clinical setting where a decrease of VLF was found in humans [
16,
20,
21]. VLF band is linked to sympatho-vagal activity, sympathetic activity reduces the VLF band component while vagal activity increases it [
22]. Even though there is uncertainty about the mechanisms responsible for activity within this band, this component is directly correlated with SDHR and it has been related to the heart’s intrinsic nervous system activity [
13]. VLF has been used as a predictor of prognosis, linked to cardiovascular disease and MetS; however, the mechanisms of those associations are not well understood [
23]. Furthermore, low VLF power has been associated with arrhythmic death more than all-cause death, a correlation that was stronger than LH and HF components [
24], which we hypothesise could point towards a possible mechanism for the increased incidence of SCD in MetS, athough we did not explore this evidence in detail in our experiments.
We observed a significant increase in LnLF components already from week 14. With respect to LF, most of the studies have reported a decrease [
14,
16,
20,
25] or no changes in this frequency band [
17]. Since, in resting conditions, LF power reflects baroreflex activity and not sympathetic activity [
26], the increased LF power that we found could be related to alterations in baroreflex control. Results about HF and LF/HF are more heterogeneous. We found no changes in HF, in line with several studies performed in patients [
20,
25], whereas others reported a decrease of this parameter [
14,
16,
17]. On the other hand, LF/HF balance was also unaltered in our experiments in MetS animals, similar to Koskinen et al. [
14] and Stuckey et al. [
25], but other studies have observed either increased [
17] or decreased [
20] LF/HF. Strong discrepancies between studies regarding frequency-domain parameters might be due to heterogeneity in ECG duration, which has a great impact in non-parametric spectral methods, and further, that not all the studies distinguished between VLF and LF bands.
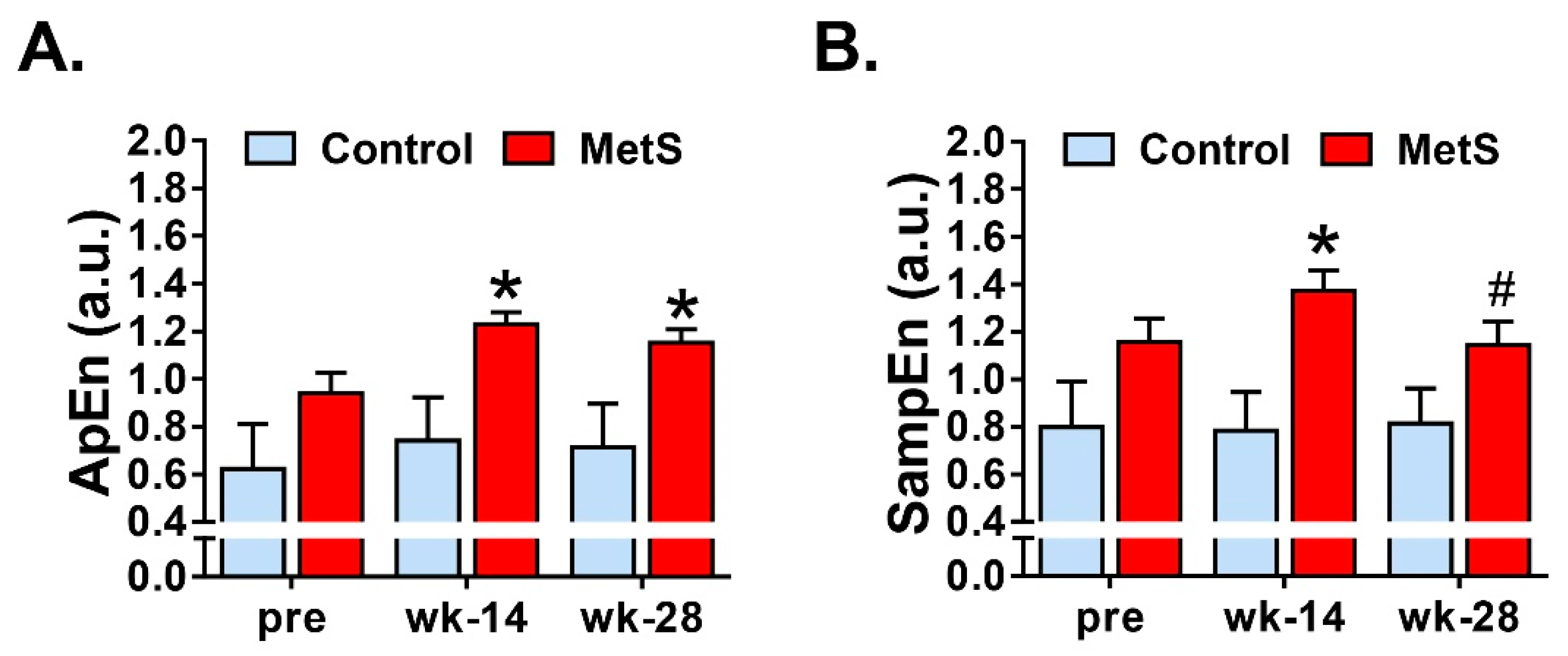
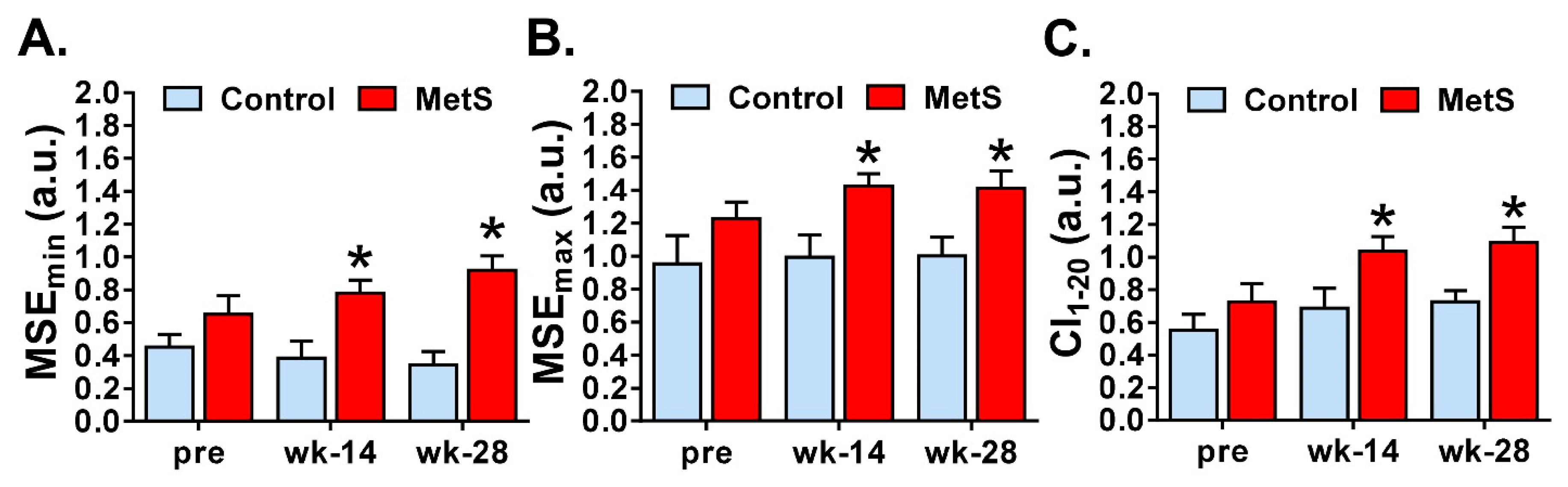
Finally, we studied the non-linear dynamics of the RR time series. Although there is still some uncertainty about the physiological significance of nonlinear parameters for HRV analysis, these measures have shown to be useful for the assessment of cardiovascular risk and sudden cardiac death [
5] and are a well-recognized way to distinguish between physiological and pathological conditions [
18]. We analyzed entropy-derived physiological markers and our results showed that MetS animals presented an increase of ApEn, SampEn, MSE
min, MSE
max and CI
1–20. In agreement with previous studies [
4], we observed that entropy measures, as well as the complexity index derived from MSE analyses had a great impact of discrimination among the groups at weeks 14 and 28. Both ApEn and SampEn quantified the irregularity and randomness of a time series, the latter being less biased and more reliable when obtained with short length time series [
5]. MSE, on the other hand, is a measure of the complexity of physiological time series, taking into account the complex temporal fluctuations conveyed in different scales. Derived from MSE, the CI
1–20, which includes the global entropy of all 20 scales, can be computed [
18]. Unfortunately, studies that have analyzed the nonlinear dynamics and the complexity of HRV using short-term RR time series in MetS are scarce. Nevertheless, collectively, the increase in the different measures of entropy found in our study indicates low predictability of cardiac fluctuations and higher randomness and complexity in cardiac activity, which other researchers have related to pathological states such as myocardial infarction, arrhythmias, sinus sick syndrome, or mutations in intracellular Ca
2+ or surface membrane ion channel proteins [
27,
28].
The main limitation of our study is the use of short-term RR interval time series, since cardiac autonomic nervous system function was only partially captured. Yet, altered sympathovagal balance can be inferred by both short-term and long-term HRV given its predictive power [
5,
18]. In order to explore all the changes that MetS produces in cardiac activity, future studies should aim to study long-term time series (i.e., 24 h), which would be necessary to identify all aspects of abnormal autonomic regulation that could be produced by MetS. Furthermore, information on the short-term complexity of HRV analysis in MetS is almost non-existent. Most of the studies have been performed using holter recordings with long-term analyses. This is of crucial importance for interpretation since there are differences in the same parameters between short-term and 24-h HRV in the same population of patients [
19]. On the other hand, the main strengths of the study are (1) the use of a diet-induced model in rabbits, which exhibited many of the characteristics of the pathology in humans [
8], (2) the controlled experimental conditions and working with experimental models is useful to avoid the interference of confounding factors in a more controlled environment, which are more present in clinical studies and could account for the heterogeneous results found in patients, and (3) we only used male NZW rabbits, but future studies should take gender into account as a factor, given that some changes have been described between men and women [
19].
 ,
, 

{kind=link}
{kind=link}
{kind=link}


