Supplementing Tropical Cattle for Improved Nutrient Utilization and Reduced Enteric Methane Emissions

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design and Animals
2.2. Feeding
2.3. Quantification of Feed Intake, Feces, and Urine Excretion
2.4. Determination of Digesta Passage
2.5. Quantification of Enteric Methane Emissions
2.6. Chemical Analysis of Samples
2.7. Data Analysis
3. Results
3.1. Feed Intake, Ingesta Composition, Digestibility, and Digesta Passage
3.2. Nitrogen Balance and Efficiency of Microbial Protein Synthesis
3.3. Methane Emissions
4. Discussion
4.1. Supplementation Effects on Intake, Digestibility, and Digesta Passage
4.2. Supplementation Effects on Nitrogen Balance and Microbial Protein Synthesis
4.3. Supplementation Effects on Enteric Methane Emissions
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Duncan, A.J.; Tarawali, S.A.; Thorne, P.J.; Valbuena, D.; Descheemaeker, K.; Homann-Kee Tui, S. Integrated crop-livestock systems—A key to sustainable intensification in Africa. Trop. Grassl. 2013, 1, 202–206. [Google Scholar] [CrossRef]
- Valbuena, D.; Erenstein, O.; Homann-Kee Tui, S.; Abdoulaye, T.; Claessens, L.; Duncan, A.J.; Gérard, B.; Rufino, M.C.; Teufel, N.; van Rooyen, A.; et al. Conservation agriculture in mixed crop–livestock systems: Scoping crop residue trade-offs in sub-Saharan Africa and South Asia. Field Crops Res. 2012, 132, 175–184. [Google Scholar] [CrossRef]
- Nyaata, O.Z.; Dorward, P.T.; Keatinge, J.D.H.; O’Neill, M.K. Availability and use of dry season feed resources on smallholder dairy farms in central Kenya. Agroforest. Syst. 2000, 50, 315–331. [Google Scholar] [CrossRef]
- Goopy, J.P.; Onyango, A.A.; Dickhoefer, U.; Butterbach-Bahl, K. A new approach for improving emission factors for enteric methane emissions of cattle in smallholder systems of East Africa—Results for Nyando, Western Kenya. Agric. Syst. 2018, 161, 72–80. [Google Scholar] [CrossRef]
- Otte, M.J.; Chilonda, P. Cattle and Small Ruminant Production Systems in Sub-Saharan Africa; FAO: Rome, Italy, 2002. [Google Scholar]
- Koralagama, K.D.N.; Mould, F.L.; Fernandez-Rivera, S.; Hanson, J. The effect of supplementing maize stover with cowpea (Vigna unguiculata) haulms on the intake and growth performance of Ethiopian sheep. Animal 2008, 2, 954–961. [Google Scholar] [CrossRef]
- Abdou, N.; Nsahlai, I.V.; Chimonyo, M. Effects of groundnut haulms supplementation on millet stover intake; digestibility and growth performance of lambs. Anim. Feed Sci. Technol. 2011, 169, 176–184. [Google Scholar] [CrossRef]
- Garg, M.R.; Gupta, B.N. Effect of supplementing urea molasses mineral block lick to straw based diet on DM intake and nutrient utilization. Asian Australas. J. Anim. Sci. 1992, 5, 39–44. [Google Scholar] [CrossRef]
- Van Soest, P.J. Rice straw, the role of silica and treatments to improve quality. Anim. Feed Sci. Technol. 2006, 130, 137–171. [Google Scholar] [CrossRef]
- Archimède, H.; Eugène, M.; Marie Magdeleine, C.; Boval, M.; Martin, C.; Morgavi, D.P.; Lecomte, P.; Doreau, M. Comparison of methane production between C3 and C4 grasses and legumes. Anim. Feed Sci. Technol. 2011, 166–167, 59–64. [Google Scholar] [CrossRef]
- Kurihara, M.; Magner, T.; Hunter, R.A.; McCrabb, G.J. Methane production and energy partition of cattle in the tropics. Br. J. Nutr. 1999, 81, 227–234. [Google Scholar] [CrossRef]
- Piñeiro-Vázquez, A.T.; Jiménez-Ferrer, G.O.; Chay-Canul, A.J.; Casanova-Lugo, F.; Díaz-Echeverría, V.F.; Ayala-Burgos, A.J.; Solorio-Sánchez, F.J.; Aguilar-Pérez, C.F.; Ku-Vera, J.C. Intake; digestibility; nitrogen balance and energy utilization in heifers fed low-quality forage and Leucaena leucocephala. Anim. Feed Sci. Technol. 2017, 228, 194–201. [Google Scholar] [CrossRef]
- Umunna, N.N.; Osuji, P.O.; Nsahlai, I.V.; Khalili, H.; Mohamed-Saleem, M.A. Effect of supplementing oat hay with lablab, sesbania, tagasaste or wheat middlings on voluntary intake, N utilization and weight gain of Ethiopian Menz sheep. Small Rumin. Res. 1995, 18, 113–120. [Google Scholar] [CrossRef]
- An, L.V.; Frankow-Lindberg, B.E.; Lindberg, J.E. Effect of harvesting interval and defoliation on yield and chemical composition of leaves; stems and tubers of sweet potato (Ipomoea batatas L. (Lam.)) plant parts. Field Crops Res. 2003, 82, 49–58. [Google Scholar] [CrossRef]
- Ondabu, N.; Kitilit, J.K.; Mwangi, J. Evaluation of sweet potato vine cultivars as animal feed in Nakuru district, Kenya. In Proceedings of the African Crop Science Conference, Entebbe, Uganda, 17–21 December 2005; pp. 575–577. [Google Scholar]
- Hadgu, G.Z.; Negesse, T.; Nurfeta, A. Chemical composition and in vitro dry matter digestibility of vines and roots of four sweet potato (Ipomoea batatas) varieties grown in southern Ethiopia. Trop. Subtrop. Agroecosyst. 2014, 17, 547–555. [Google Scholar]
- CIP (Centro Internacional de la Papa). Sweetpotato in Africa; International Potato Centre: Lima, Peru, 2011; Available online: https://cipotato.org/research/sweetpotato-in-africa/ (accessed on 13 April 2018).
- Etela, I.; Larbi, A.; Ikhatua, U.J.; Bamikole, M.A. Supplementing Guinea grass with fresh sweet potato foliage for milk production by Bunaji and N’Dama cows in early lactation. Livest. Sci. 2009, 120, 87–95. [Google Scholar] [CrossRef]
- Phesatcha, K.; Wanapat, M. Performance of lactating dairy cows fed a diet based on treated rice straw and supplemented with pelleted sweet potato vines. Trop. Anim. Health Prod. 2013, 45, 533–538. [Google Scholar] [CrossRef]
- Makkar, H.P.S. Feed supplementation block technology—Past, present and future. In Urea-Molasses Multinutrient Blocks: Simple and Effective Feed Supplement Technology for Ruminant Agriculture; Makkar, H.P.S., Sánchez, M., Speedy, A.W., Eds.; FAO: Rome, Italy, 2007; pp. 1–12. [Google Scholar]
- Waruiru, R.M.; Onyando, C.O.; Machuka, R.O. Effect of feeding urea-molasses blocks with incorporated fenbendazole on grazing dairy heifers naturally infected with gastrointestinal nematodes. J. S. Afr. Vet. Assoc. 2003, 74, 49–52. [Google Scholar] [CrossRef] [PubMed]
- Srinivas, B.; Gupta, B.N. Rumen fermentation, bacterial and total volatile fatty acid (TVFA) production rates in cattle fed on urea-molasses-mineral block licks supplement. Anim. Feed Sci. Technol. 1997, 65, 275–286. [Google Scholar] [CrossRef]
- Hristov, A.N.; Ropp, J.K.; Grandeen, K.L.; Abedi, S.; Etter, R.P.; Melgar, A.; Foley, A.E. Effect of carbohydrate source on ammonia utilization in lactating dairy cows. J. Anim. Sci. 2005, 83, 408–421. [Google Scholar] [CrossRef] [PubMed]
- Schwab, C.G.; Huhtanen, P.; Hunt, C.W.; Hvelplund, T. Nitrogen requirements of cattle. In Nitrogen and Phosphorous Nutrition of Cattle; Pfeffer, E., Hristov, A.N., Eds.; CAB International: Wallingford, UK, 2005; pp. 13–70. [Google Scholar]
- Lukuyu, B.; Gachuiri, C.K.; Agili, S.; Leon-Velarde, C.; Kirui, J. Making High Quality Sweetpotato Silage: An Improved Tube Silage Making Method; International Potato Center: Nairobi, Kenya, 2012; Available online: https://cgspace.cgiar.org/handle/10568/89025 (accessed on 10 April 2018).
- Ahnert, S.; Dickhoefer, U.; Schulz, F.; Susenbeth, A. Influence of ruminal Quebracho tannin extract infusion on apparent nutrient digestibility, nitrogen balance; and urinary purine derivatives excretion in heifers. Livest. Sci. 2015, 177, 63–70. [Google Scholar] [CrossRef]
- Teeter, R.G.; Owens, F.N.; Mader, T.L. Ytterbium chloride as a marker for particulate matter in the rumen. J. Anim. Sci. 1984, 58, 465–473. [Google Scholar] [CrossRef]
- Uden, P.; Colucci, P.E.; Van Soest, P.J. Investigation of chromium, cerium and cobalt as markers in digesta rate of passage studies. J. Sci. Food Agric. 1980, 31, 625–632. [Google Scholar] [CrossRef]
- Richter, H.; Schlecht, E. Accounting for marker disassociation when modelling time-dependent ruminal escape of particles based on the faecal excretion of Ytterbium. Anim. Feed Sci. Technol. 2006, 128, 135–154. [Google Scholar] [CrossRef]
- AOAC (Association of Official Analytical Chemists). Official Methods of Analysis, 15th ed.; AOAC: Washington, DC, USA, 1990. [Google Scholar]
- VDLUFA (Verband Deutscher Landwirtschaftlicher Untersuchungs- und Forschungsanstalten). Handbuch der landwirtschaftlichen Versuchs- und Untersuchungsmethodik Band III: Die Chemische Untersuchung von Futtermitteln; VDLUFA: Darmstadt, Germany, 2012. [Google Scholar]
- Anderson, D.L.; Henderson, L.J. Sealed chamber digestion for plant nutrient analyses. Agron. J. 1986, 78, 937–939. [Google Scholar] [CrossRef]
- Chen, X.B.; Gomez, M.J. Estimation of Microbial Protein Supply to Sheep and Cattle based on Urinary Excretion of Purine Derivatives—An Overview of the Technical Details; Rowett Research Institute: Bucksburn, UK, 1992. [Google Scholar]
- Wassie, S.; Ali, A.I.M.; Korir, D.; Butterbach-Bahl, K.; Goopy, J.P.; Merbold, L.; Schlecht, E.; Dickhoefer, U. Effects of feed intake level on efficiency of microbial protein synthesis and nitrogen balance in Boran steers consuming tropical poor-quality forage. Arch. Anim. Nutr. 2019, 73, 140–157. [Google Scholar] [CrossRef] [PubMed]
- Aregheore, E.M. Nutritive value of sweet potato (Ipomea batatas (L) Lam) forage as goat feed: Voluntary intake; growth and digestibility of mixed rations of sweet potato and Batiki grass (Ischaemum aristatum var. indicum). Small Rumin. Res. 2004, 51, 235–241. [Google Scholar] [CrossRef]
- Korir, D.; Goopy, J.P.; Gachuiri, C.; Butterbach-Bahl, K. Supplementation with Calliandra calothyrsus improves nitrogen retention in cattle fed low-protein diets. Anim. Prod. Sci. 2016, 56, 619–626. [Google Scholar] [CrossRef]
- Mlay, P.S.; Pereka, A.E.; Weisbjerg, M.R.; Hvelplund, T.; Madsen, J. Digestion and passage kinetics of fibre in mature dairy heifers maintained on poor quality hay as affected by the source and level of nitrogen supplementation. Anim. Feed Sci. Technol. 2003, 109, 19–33. [Google Scholar] [CrossRef]
- Das, A.; Singh, G.P. Effect of different levels of berseem (Trifolium alexdrinum) supplementation of wheat straw on some physical factors regulating intake and digestion. Anim. Feed Sci. Technol. 1999, 81, 133–149. [Google Scholar] [CrossRef]
- Giger-Reverdin, S. Characterisation of feedstuffs for ruminants using some physical parameters. Anim. Feed Sci. Technol. 2000, 86, 53–69. [Google Scholar] [CrossRef]
- Ishida, H.; Suzuno, H.; Sugiyama, N.; Innami, S.; Tadokoro, T.; Maekawa, A. Nutritive evaluation on chemical components of leaves, stalks and stems of sweet potatoes (Ipomoea batatas Poir). Food Chem. 2000, 68, 359–367. [Google Scholar] [CrossRef]
- Mu, T.H.; Tan, S.S.; Xue, Y.L. The amino acid composition; solubility and emulsifying properties of sweet potato protein. Food Chem. 2009, 112, 1002–1005. [Google Scholar] [CrossRef]
- Abdulrazak, S.A.; Muinga, R.W.; Thorpe, W.; Ørskov, E.R. The effects of supplementation with Gliricidia sepium or Leucaena leucocephala forage on intake, digestion and live-weight gains of Bos taurus × Bos indicus steers offered Napier grass. Anim. Sci. 1996, 63, 381–388. [Google Scholar] [CrossRef]
- Mathis, C.P.; Cochran, R.C.; Heldt, J.S.; Woods, B.C.; Abdelgadir, I.E.; Olson, K.C.; Titgemeyer, E.C.; Vanzant, E.S. Effects of supplemental degradable intake protein on utilization of medium- to low-quality forages. J. Anim. Sci. 2000, 78, 224–232. [Google Scholar] [CrossRef]
- Aguerre, M.J.; Wattiaux, M.A.; Powell, J.M.; Broderick, G.A.; Arndt, C. Effect of forage-to-concentrate ratio in dairy cow diets on emission of methane, carbon dioxide, and ammonia, lactation performance, and manure excretion. J. Dairy Sci. 2011, 94, 3081–3093. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, P.M.; Charmley, E. Methane yields from Brahman cattle fed tropical grasses and legumes. Anim. Prod. Sci. 2012, 52, 225–239. [Google Scholar] [CrossRef]
- McGinn, S.M.; Beauchemin, K.A.; Coates, T.; Colombatto, D. Methane emissions from beef cattle: Effects of monensin, sunflower oil, enzymes, yeast, and fumaric acid. J. Anim. Sci. 2004, 82, 3346–3356. [Google Scholar] [CrossRef] [PubMed]
- Chaokaur, A.; Nishida, T.; Phaowphaisal, I.; Sommart, K. Effects of feeding level on methane emissions and energy utilization of Brahman cattle in the tropics. Agric. Ecosyst. Environ. 2015, 199, 225–230. [Google Scholar] [CrossRef]
- Hammond, K.J.; Pacheco, D.; Burke, J.L.; Koolaard, J.P.; Muetzel, S.; Waghorn, G.C. The effects of fresh forages and feed intake level on digesta kinetics and enteric methane emissions from sheep. Anim. Feed Sci. Technol. 2014, 193, 32–43. [Google Scholar] [CrossRef]
- Janssen, P.H. Influence of hydrogen on rumen methane formation and fermentation balances through microbial growth kinetics and fermentation thermodynamics. Anim. Feed Sci. Technol. 2010, 160, 1–22. [Google Scholar] [CrossRef]
- Buxton, D.R.; Redfearn, D.D. Plant limitations to fiber digestion and utilization. J. Nutr. 1997, 127, 814S–818S. [Google Scholar] [CrossRef] [PubMed]
- Coleman, S.W.; Hart, S.P.; Sahlu, T. Relationships among forage chemistry, rumination and retention time with intake and digestibility of hay by goats. Small Rumin. Res. 2003, 50, 129–140. [Google Scholar] [CrossRef]
- Goopy, J.P.; Donaldson, A.; Hegarty, R.; Vercoe, P.E.; Haynes, F.; Barnett, M.; Oddy, V.H. Low-methane yield sheep have smaller rumens and shorter rumen retention time. Br. J. Nutr. 2014, 111, 578–585. [Google Scholar] [CrossRef] [PubMed]
- Petersen, S.O.; Sommer, S.G.; Béline, F.; Burton, C.; Dach, J.; Dourmad, J.Y.; Leip, A.; Misselbrook, T.; Nicholson, F.; Poulsen, H.D.; et al. Recycling of livestock manure in a whole-farm perspective. Livest. Sci. 2007, 112, 180–191. [Google Scholar] [CrossRef]
- Hristov, A.N.; Oh, J.; Lee, C.; Meinen, R.; Montes, F.; Ott, T.; Firkins, J.; Rotz, A.; Dell, C.; Adesogan, A.; et al. Mitigation of Greenhouse Gas Emissions in Livestock Production—A Review of Technical Options for non-CO2 Emissions; FAO Animal Production and Health Paper 177; FAO: Rome, Italy, 2013. [Google Scholar]


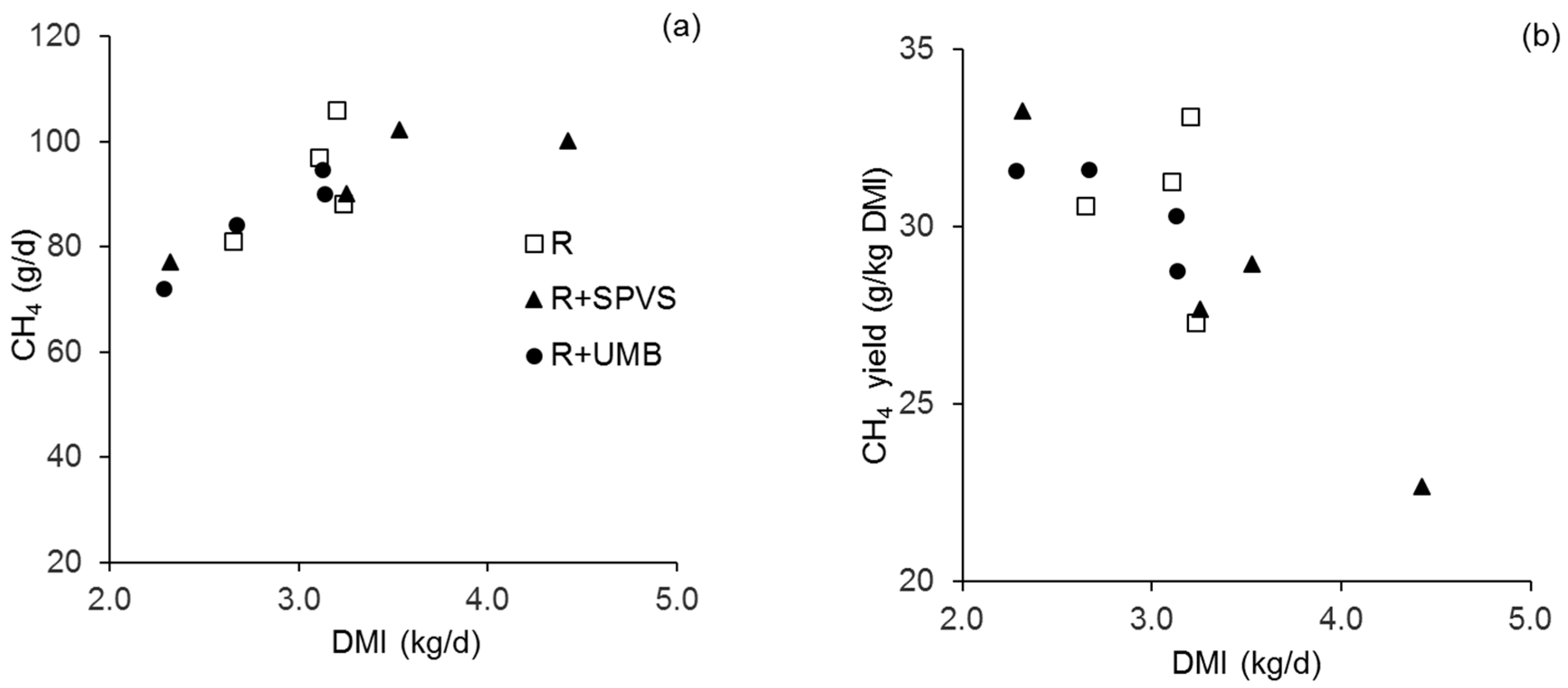{kind=link}
| Feed | Period 2 | DM | OM | CP | NDF | ADF |
|---|---|---|---|---|---|---|
| (g/kg FM) | (g/kg DM) | |||||
| Roughage 1 | 1 | 813 | 889 | 70.3 | 731 | 479 |
| 2 | 742 | 895 | 72.2 | 738 | 460 | |
| SPVS | 1 | 193 | 875 | 136.1 | 537 | 393 |
| 2 | 199 | 877 | 143.7 | 537 | 393 | |
| UMB | 1, 2 | 899 | 502 | 373.6 | 27 | 17 |
| Variable | R | R + SPVS | R + UMB | SEM | p-Value |
|---|---|---|---|---|---|
| Daily intake (g/kg0.75 LW) and (share of supplement, g/100 g) | |||||
| DM | 70.9 | 76.0 (19.3) | 66.8 (1.7) | 3.08 | 0.170 |
| OM | 63.1 | 67.5 (19.0) | 59.0 (1.0) | 2.79 | 0.158 |
| NDF | 50.4 | 52.6 (15.0) | 47.7 (0.1) | 2.22 | 0.230 |
| ADF | 32.3 | 33.9 (17.0) | 30.2 (0.1) | 1.30 | 0.311 |
| CP | 5.6 | 6.7 (30.5) | 5.5 (7.7) | 0.31 | 0.113 |
| Ingesta composition (g/kg DM) | |||||
| OM | 890 | 887 | 884 | 2.21 | 0.129 |
| NDF | 709 | 691 | 715 | 9.31 | 0.122 |
| ADF | 456 | 447 | 453 | 3.62 | 0.163 |
| CP | 79 a | 88 b | 82 a | 0.21 | 0.040 |
| Digestibility (g/kg) | |||||
| DM | 474 a | 510 b | 480 ab | 10.07 | 0.041 |
| OM | 509 A | 539 B | 512 AB | 7.78 | 0.071 |
| NDF | 496 A | 530 B | 506 AB | 8.28 | 0.058 |
| ADF | 428 | 458 | 429 | 6.79 | 0.134 |
| Variable | R | R + SPVS | R + UMB | SEM | p-Value |
|---|---|---|---|---|---|
| Liquid digesta passage | |||||
| λl (%/h) | 9.2 | 9.2 | 9.0 | 0.43 | 0.972 |
| TTl (h) | 4.3 | 3.1 | 4.4 | 0.39 | 0.230 |
| CMRTl (h) | 22.0 | 22.0 | 23.2 | 1.08 | 0.854 |
| TMRTl (h) | 26.3 | 25.0 | 27.6 | 0.97 | 0.567 |
| Solid digesta passage | |||||
| λs (%/h) | 3.5 | 3.8 | 3.4 | 0.14 | 0.262 |
| TTs (h) | 18.9 | 16.8 | 16.2 | 0.77 | 0.368 |
| CMRTs (h) | 56.8 | 54.4 | 60.2 | 2.08 | 0.298 |
| TMRTs (h) | 75.8 | 71.2 | 76.4 | 2.08 | 0.460 |
| Variable | Liquid Digesta Passage | Solid Digesta Passage | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| λl (%/h) | TTl (h) | CMRTl (h) | TMRTl (h) | λs (%/h) | TTs (h) | CMRTs (h) | TMRTs (h) | |||||||||
| Feed intake (g/kg0.75 LW) | ||||||||||||||||
| OM | 0.29 | n.s. | −0.36 | n.s. | −0.29 | n.s. | −0.57 | (*) | 0.92 | *** | 0.45 | n.s. | −0.92 | *** | −0.75 | ** |
| CP | 0.42 | n.s. | −0.26 | n.s. | −0.42 | n.s. | −0.69 | * | 0.82 | ** | 0.31 | n.s. | −0.82 | ** | −0.66 | * |
| NDF | 0.29 | n.s. | −0.29 | n.s. | −0.29 | n.s. | −0.52 | (*) | 0.95 | *** | 0.48 | n.s. | −0.95 | *** | −0.78 | ** |
| ADF | 0.31 | n.s. | −0.36 | n.s. | −0.31 | n.s. | −0.60 | * | 0.91 | *** | 0.45 | n.s. | −0.91 | *** | −0.74 | ** |
| Ingesta composition (g/kg DM) | ||||||||||||||||
| OM | −0.26 | n.s. | −0.31 | n.s. | 0.26 | n.s. | 0.05 | n.s. | 0.45 | n.s. | 0.57 | (*) | −0.45 | n.s. | −0.33 | n.s. |
| CP | 0.25 | n.s. | −0.20 | n.s. | −0.25 | n.s. | −0.48 | n.s. | 0.31 | n.s. | −0.15 | n.s. | −0.31 | n.s. | −0.30 | n.s. |
| NDF | −0.14 | n.s. | −0.06 | n.s. | 0.14 | n.s. | 0.21 | n.s. | 0.21 | n.s. | 0.08 | n.s. | −0.21 | n.s. | −0.21 | n.s. |
| ADF | −0.55 | (*) | 0.07 | n.s. | 0.55 | (*). | 0.65 | * | −0.70 | * | −0.19 | n.s. | 0.70 | * | 0.57 | (*). |
| Digestibility (g/kg) | ||||||||||||||||
| DM | 0.13 | n.s. | −0.62 | * | −0.13 | n.s. | −0.41 | n.s. | 0.65 | * | −0.10 | n.s. | −0.65 | * | −0.73 | ** |
| OM | 0.00 | n.s. | −0.70 | * | 0.00 | n.s. | −0.31 | n.s. | 0.45 | n.s. | −0.17 | n.s. | −0.45 | n.s. | −0.55 | (*) |
| NDF | 0.20 | n.s. | −0.55 | (*) | −0.20 | n.s. | −0.43 | n.s. | 0.64 | * | −0.11 | n.s. | −0.64 | * | −0.76 | ** |
| ADF | 0.10 | n.s. | −0.69 | * | −0.10 | n.s. | −0.37 | n.s. | 0.68 | * | −0.02 | n.s. | −0.68 | * | −0.76 | ** |
| Duodenal microbial N flow (g N/day) | 0.66 | * | −0.04 | n.s. | −0.66 | * | −0.76 | ** | 0.48 | n.s. | 0.00 | n.s. | −0.48 | n.s. | −0.53 | (*) |
| EMPS | ||||||||||||||||
| (g N/kg OM intake) | 0.42 | n.s. | 0.09 | n.s. | −0.42 | n.s. | −0.39 | n.s. | 0.01 | n.s. | −0.02 | n.s. | −0.01 | n.s. | −0.07 | n.s. |
| (g N/kg dOM) | 0.42 | n.s. | 0.09 | n.s. | −0.42 | n.s. | −0.39 | n.s. | 0.01 | n.s. | −0.02 | n.s. | −0.01 | n.s. | −0.07 | n.s. |
| (g N/g N intake) | 0.28 | n.s. | −0.01 | n.s. | −0.28 | n.s. | −0.21 | n.s. | −0.03 | n.s. | 0.06 | n.s. | 0.03 | n.s. | −0.06 | n.s. |
| Variable | R | R + SPVS | R + UMB | SEM | p Value |
|---|---|---|---|---|---|
| Nitrogen balance (g/kg0.75 LW) | |||||
| N intake | 0.90 | 1.08 | 0.88 | 0.050 | 0.113 |
| Fecal N excretion | 0.60 | 0.65 | 0.60 | 0.028 | 0.367 |
| Urine N excretion | 0.45 | 0.48 | 0.45 | 0.026 | 0.655 |
| N retention | −0.15 A | −0.05 B | −0.17 A | 0.021 | 0.051 |
| Fecal-to-urine N ratio | 1.33 | 1.44 | 1.32 | 0.066 | 0.576 |
| Urine PD excretion (mmol/day) | |||||
| Allantoin | 35.62 | 40.55 | 29.62 | 2.524 | 0.281 |
| Uric acid | 7.21 | 9.87 | 6.07 | 0.849 | 0.324 |
| Xanthine + hypoxanthine | 0.03 | 0.06 | 0.02 | 0.006 | 0.150 |
| Total PD excretion | 42.86 | 50.48 | 35.72 | 3.274 | 0.283 |
| Creatinine (mmol/day) | 29.66 | 33.63 | 25.70 | 1.357 | 0.452 |
| PD-to-creatinine ratio | 1.45 | 1.50 | 1.38 | 0.078 | 0.572 |
| Duodenal microbial N flow (g/day) | 27.73 | 33.95 | 21.85 | 2.696 | 0.512 |
| EMPS | |||||
| (g N/kg OM intake) | 10.23 | 11.63 | 8.76 | 0.849 | 0.139 |
| (g N/kg dOM) | 20.20 | 21.61 | 16.98 | 1.601 | 0.617 |
| (g N/g N intake) | 0.71 | 0.73 | 0.59 | 0.052 | 0.126 |
| Methane (g) Emitted | R | R + SPVS | R + UMB | SEM | p-value |
|---|---|---|---|---|---|
| /animal·day | 93.1 | 92.4 | 85.3 | 3.00 | 0.611 |
| /kg DMI | 30.6 | 28.1 | 30.6 | 0.85 | 0.240 |
| /kg OMI | 34.4 | 31.7 | 34.6 | 0.99 | 0.238 |
| /kg NDFI | 43.2 | 40.8 | 42.7 | 1.33 | 0.538 |
| /kg ADFI | 67.0 | 62.9 | 67.4 | 1.76 | 0.395 |
| /kg dDM | 64.7 b | 55.2 a | 63.8 ab | 2.33 | 0.041 |
| /kg dOM | 67.8 B | 58.9 A | 67.5 B | 2.28 | 0.094 |
| /kg dNDF | 88.1 | 77.0 | 84.6 | 3.52 | 0.167 |
| /kg dADF | 157.5 | 137.7 | 157.1 | 5.40 | 0.133 |
| Parameter | Methane (g) emitted | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| /Animal·Day | /kg DMI | /kg OMI | /kg NDFI | /kg dDM | /kg dOM | /kg dNDF | ||||||||
| Liquid digesta passage | ||||||||||||||
| λl (%/h) | 0.78 | ** | −0.13 | n.s. | −0.15 | n.s. | −0.15 | n.s. | −0.14 | n.s. | −0.06 | n.s. | −0.16 | n.s. |
| TTl (h) | 0.20 | n.s. | 0.39 | n.s. | 0.34 | n.s. | 0.39 | n.s. | 0.54 | (*) | 0.57 | (*) | 0.52 | (*) |
| TMRTl (h) | −0.84 | *** | 0.40 | n.s. | 0.41 | n.s. | 0.41 | n.s. | 0.43 | n.s. | 0.36 | n.s. | 0.43 | n.s. |
| Solid digesta passage | ||||||||||||||
| λs (%/h) | 0.55 | (*) | −0.90 | *** | −0.91 | *** | −0.90 | *** | −0.84 | *** | −0.81 | ** | −0.83 | *** |
| TTs (h) | 0.03 | n.s. | −0.41 | n.s. | −0.40 | n.s. | −0.28 | n.s. | −0.16 | n.s. | −0.22 | n.s. | −0.04 | n.s. |
| TMRTs (h) | −0.62 | * | 0.68 | * | 0.70 | * | 0.75 | ** | 0.76 | ** | 0.67 | * | 0.84 | *** |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ali, A.I.M.; Wassie, S.E.; Korir, D.; Merbold, L.; Goopy, J.P.; Butterbach-Bahl, K.; Dickhoefer, U.; Schlecht, E. Supplementing Tropical Cattle for Improved Nutrient Utilization and Reduced Enteric Methane Emissions. Animals 2019, 9, 210. https://doi.org/10.3390/ani9050210
Ali AIM, Wassie SE, Korir D, Merbold L, Goopy JP, Butterbach-Bahl K, Dickhoefer U, Schlecht E. Supplementing Tropical Cattle for Improved Nutrient Utilization and Reduced Enteric Methane Emissions. Animals. 2019; 9(5):210. https://doi.org/10.3390/ani9050210
Chicago/Turabian StyleAli, Asep I. M., Shimels E. Wassie, Daniel Korir, Lutz Merbold, John P. Goopy, Klaus Butterbach-Bahl, Uta Dickhoefer, and Eva Schlecht. 2019. "Supplementing Tropical Cattle for Improved Nutrient Utilization and Reduced Enteric Methane Emissions" Animals 9, no. 5: 210. https://doi.org/10.3390/ani9050210